
Transcriptome Analysis Reveals the Impact of Arbuscular Mycorrhizal Symbiosis on Toona ciliata var. pubescens Seedlings

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material and Experimental Design
2.2. Harvest
2.3. Quantification of AMF Colonization
2.4. RNA Extraction and Sequencing
2.5. Assembly and Annotation
2.6. Differential Expression Analysis
2.7. GO (Gene Ontology) and KEGG (Kyoto Encyclopedia of Genes and Genomes) Enrichment Analysis
2.8. Statistical Analyses
2.9. Data Availability
3. Results
3.1. AM Colonization and Plant Growth
3.2. Sequencing Data Quality
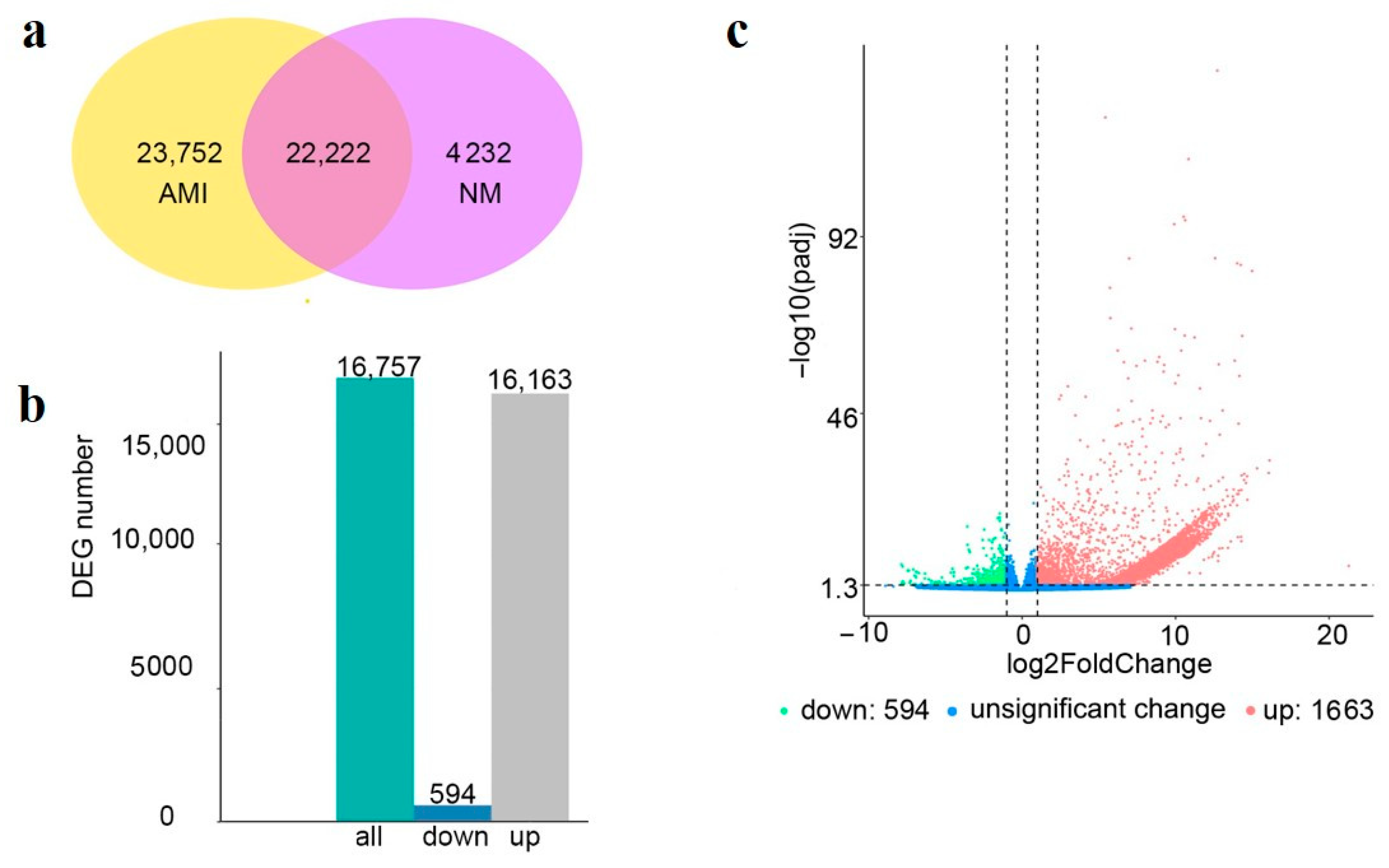
3.3. Analysis of DEGs
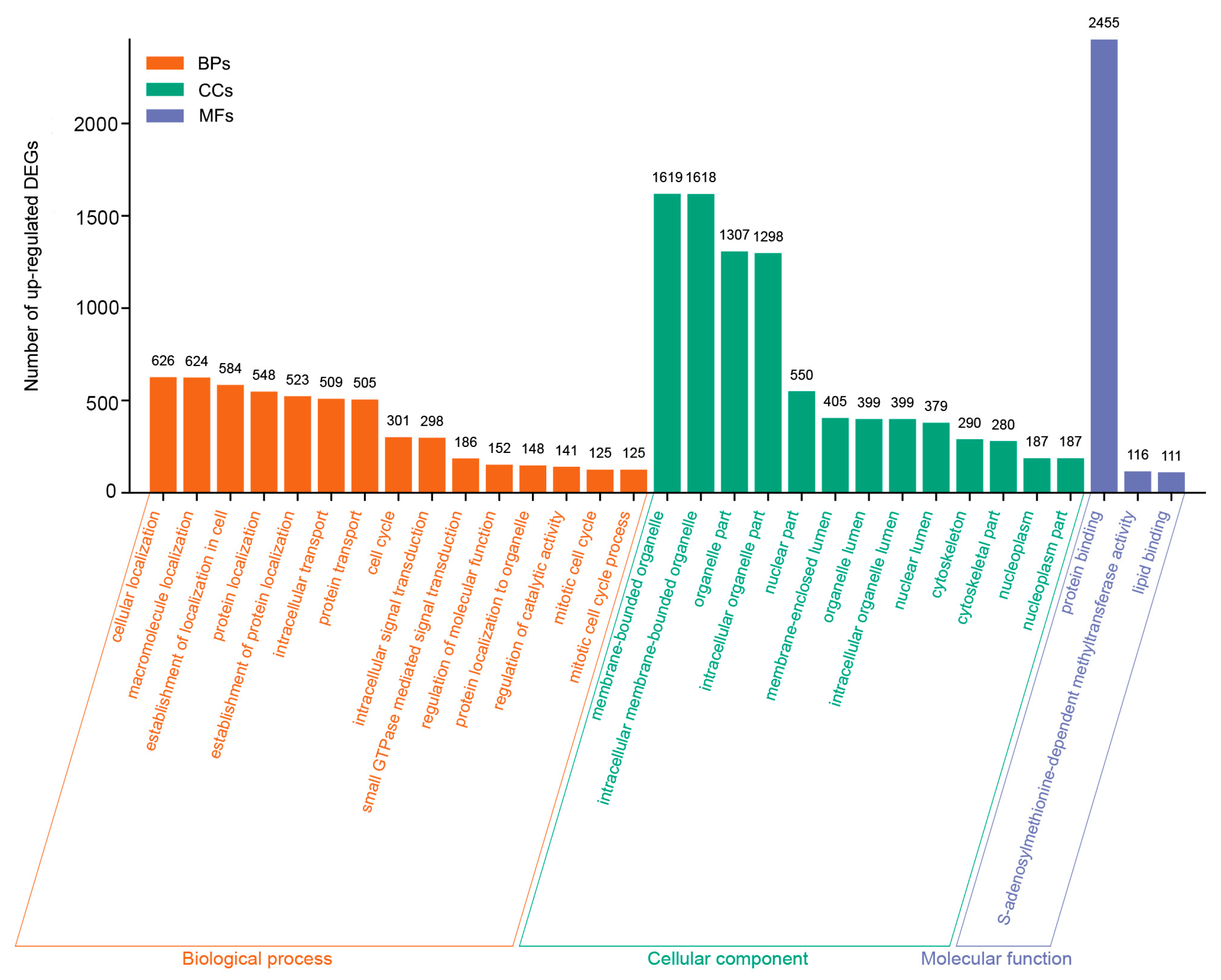
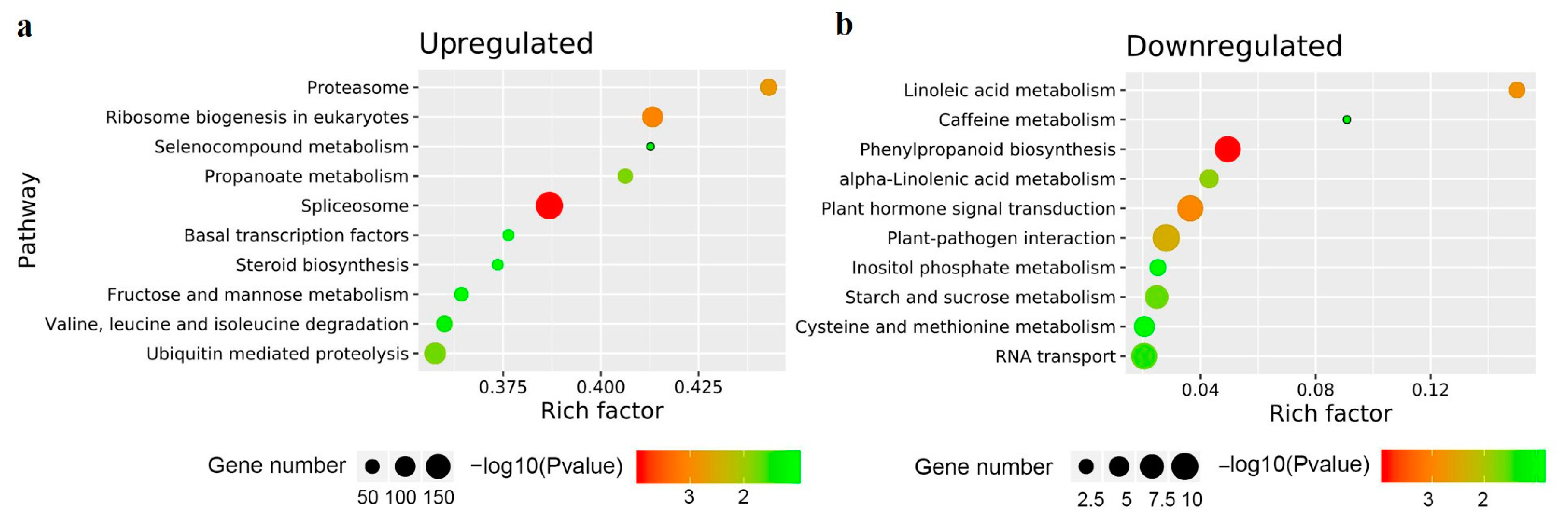
3.4. GO and KEGG Enrichment Analysis of the DEGs
3.5. Upregulated DEGs Related to the Oxidative Stress Response and Nutrient Transport
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, H.L.; Zhang, L.; Liao, C.K. Seed rain, soil seed bank, and natural regeneration of natural Toona ciliata var. pubescens forest. J. Appl. Ecol. 2012, 23, 972–978. [Google Scholar]
- Pan, J.; Wang, Q.; Guo, X.; Jiang, X.; Cheng, Q.; Fu, L.; Liu, W.; Zhang, L. Local patterns of arbuscular mycorrhizal fungal diversity and community structure in a natural Toona ciliata var. pubescens forest in South Central China. PeerJ 2021, 9, e11331. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.Y.; Fu, L.; Zhang, L.; Su, H.; Liang, Y.L. Effects of forest soil and soil-borne fungi on seed germination and seedling survival of Toona ciliate var. pubescens. For. Res. 2017, 30, 871–877. [Google Scholar]
- Liu, W.; Jin, J.; Li, Y.; Tan, Z.; Jiang, J.; Liu, J. Effects of waters, fertilization with different nitrogen and phosphorus ratios on the growth, nutrient distribution and chlorophyll fluorescence characteristics of Toona ciliate var. pubescens seedlings. J. West China For. Sci. 2021, 50, 83–90. [Google Scholar]
- Sun, B.; Zhang, T.; Zhao, Q. Comprehensive evaluation of soil fertility in the hilly and mountainous region of Southeastern China. Acta Pedol. Sin. 1995, 32, 362–369. [Google Scholar]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Ezawa, T.; Saito, K. How do arbuscular mycorrhizal fungi handle phosphate? New insight into fine-tuning of phosphate metabolism. New Phytol. 2018, 220, 1116–1121. [Google Scholar] [CrossRef] [PubMed]
- Ferrol, N.; Azcón-Aguilar, C.; Pérez-Tienda, J. Review: Arbuscular mycorrhizas as key players in sustainable plant phosphorus acquisition: An overview on the mechanisms involved. Plant Sci. 2019, 280, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Fang, F.; Wu, N.; Li, L.; Tang, M. Nitrate transporter gene expression and kinetics of nitrate uptake by Populus × canadensis ‘Neva’ in relation to arbuscular mycorrhizal fungi and nitrogen availability. Front. Microbiol. 2020, 11, 176. [Google Scholar] [CrossRef]
- Bennett, A.E.; Groten, K. The costs and benefits of plant-arbuscular mycorrhizal fungal interactions. Annu. Rev. Plant Biol. 2022, 73, 649–672. [Google Scholar] [CrossRef]
- Begum, N.; Cheng, Q.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef]
- Fusconi, A. Regulation of root morphogenesis in arbuscular mycorrhizae: What role do fungal exudates, phosphate, sugars and hormones play in lateral root formation? Annu. Rev. 2014, 113, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Evelin, H.; Kapoor, R. Arbuscular mycorrhizal symbiosis modulates antioxidant response in salt-stressed Trigonella foenum-graecum plants. Mycorrhiza 2014, 24, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Zhai, S.; Wu, Y.; Xu, C.; Chen, W.; Feng, J.; Zheng, Q.; Meng, Y.; Yang, H. Symbiotic soil fungi suppress N2O emissions but facilitate nitrogen remobilization to grains in sandy but not clay soils under organic amendments. Appl. Soil Ecol. 2021, 167, 104012. [Google Scholar] [CrossRef]
- Thirkell, T.J.; Charters, M.D.; Elliott, A.J.; Sait, S.M.; Field, K.J. Are mycorrhizal fungi our sustainable saviours? Considerations for achieving food security. J. Ecol. 2017, 105, 921–929. [Google Scholar] [CrossRef]
- Liu, N.; Shao, C.; Sun, H.; Liu, Z.; Guan, Y.; Wu, L.; Zhang, L.; Pan, X.; Zhang, Z.; Zhang, Y.; et al. Arbuscular mycorrhizal fungi biofertilizer improves American ginseng (Panax quinquefolius L.) growth under the continuous cropping regime. Geoderma 2020, 363, 114155. [Google Scholar] [CrossRef]
- Loo, W.T.; Chua, K.O.; Mazumdar, P.; Cheng, A.; Osman, N.; Harikrishna, J.A. Arbuscular mycorrhizal symbiosis: A strategy for mitigating the impacts of climate change on tropical legume crops. Plants 2022, 11, 2875. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; Wiemken, A.; Sanders, I.R. Different arbuscular mycorrhizal fungal species are potential determinants of plant community structure. Ecology 1998, 79, 2082–2091. [Google Scholar]
- Gillespie, A.W.; Farrell, R.E.; Walley, F.L.; Ross, A.R.; Leinweber, P.; Eckhardt, K.U.; Regier, T.Z.; Blyth, R.I. Glomalin-related soil protein contains non-mycorrhizal-related heat-stable proteins, lipids and humic materials. Soil Biol. Biochem. 2011, 43, 766–777. [Google Scholar] [CrossRef]
- Huang, Y.; Jiang, Y.; Wang, Z.; Song, F.; He, L.; Tian, R.; Wu, L. The suppression effect and mechanism of arbuscular mycorrhizal fungi against plant root rot. J. Appl. Ecol. 2021, 32, 1890–1902. [Google Scholar]
- Fu, L. Study on the community characteristics of arbuscular mycorrhizal fungi in soil of the community of natural forest of Toon ciliata. Master’s Thesis, Jiangxi Agricultural University, Nanchang, China, 2018. [Google Scholar]
- Redecker, D.; Schüßler, A.; Stockinger, H.; Stürmer, S.L.; Morton, J.B.; Walker, C. An evidence-based consensus for the classification of arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza 2013, 23, 515–531. [Google Scholar] [CrossRef]
- Tedone, L.; Ruta, C.; De Cillis, F.; De Mastro, G. Effects of Septoglomus viscosum inoculation on biomass yield and steviol glycoside concentration of some Stevia rebaudiana chemotypes. Sci. Hortic. 2020, 262, 109026. [Google Scholar] [CrossRef]
- Tarraf, W.; Ruta, C.; Tagarelli, A.; De Cillis, F.; De Mastro, G. Influence of arbuscular mycorrhizae on plant growth, essential oil production and phosphorus uptake of Salvia officinalis L. Ind. Crop. Prod. 2017, 102, 144–153. [Google Scholar] [CrossRef]
- Li, J.; Wang, Q.; Wang, Y.; Zhang, Q.; Luo, J.; Jiang, X.; Liu, W.; Zhao, X. Effects of arbuscular mycorrhizal fungi on improvement of degraded landscape soil in an ionized rare earth mining area, subtropical China. Soil Sci. Soc. Am. J. 2022, 86, 275–292. [Google Scholar] [CrossRef]
- Liu, W.; Zhao, X.; Zhang, Q.; Li, J.; Xue, H.; Zhang, J.; Wu, J.; Deng, G.; Zhang, W.; Li, X.; et al. A Strain of Septoglomus viscosum, including Its Microbial Agent and Application. Chinese Patent CN110819542B, 9 February 2021. [Google Scholar]
- Chen, W.; Li, J.; Zhu, H.; Xu, P.; Chen, J.; Yao, Q. Arbuscular mycorrhizal fungus enhances lateral root formation in Poncirus trifoliata (L.) as revealed by RNA-Seq analysis. Front Plant Sci. 2017, 8, 2039. [Google Scholar] [CrossRef]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.; Piché, Y. Ink and vinegar., a simple staining technique for arbuscular-mycorrhizal fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef] [PubMed]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 123. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level—The DESeq Package; European Molecular Biology Laboratory (EMBL): Heidelberg, Germany, 2012. [Google Scholar]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Vangelisti, A.; Natali, L.; Bernardi, R.; Sbrana, C.; Turrini, A.; Hassani-Pak, K.; Hughes, D.; Cavallini, A.; Giovannetti, M.; Giordani, T. Transcriptome changes induced by arbuscular mycorrhizal fungi in sunflower (Helianthus annuus L.) roots. Sci. Rep. 2018, 8, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, R.; Tian, H.; Gao, Y. Transcriptome responses in wheat roots to colonization by the arbuscular mycorrhizal fungus Rhizophagus irregularis. Mycorrhiza 2018, 28, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.K.; Sharma, M.; Pandey, A.; Shanmugam, T. Overview of small GTPase signaling proteins in plants. In GTPases; Springer Briefs in Plant Science; Springer: Cham, Switzerland, 2015; pp. 9–14. [Google Scholar]
- Yuksel, B.; Memon, A.R. Legume small GTPases and their role in the establishment of symbiotic associations with Rhizobium spp. Plant Signal. Behav. 2009, 4, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, E. The Small GTPase Superfamily in Plants: A conserved regulatory module with novel functions. Annu. Rev. Plant Biol. 2020, 71, 247–272. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress., antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Jajoo, A.; Mathur, S. Role of arbuscular mycorrhizal fungi as an underground savior for protecting plants from abiotic stresses. Physiol. Mol. Biol. Plants 2021, 27, 2589–2603. [Google Scholar] [CrossRef]
- Sharma, S.; Anand, G.; Singh, N.; Kapoor, R. Arbuscular mycorrhiza augments arsenic tolerance in wheat (Triticum aestivum L.) by strengthening antioxidant defense system and thiol metabolism. Front. Plant Sci. 2017, 8, 906. [Google Scholar] [CrossRef]
- Jia, T.; Wang, J.; Chang, W.; Fan, X.; Sui, X.; Song, F. Proteomics analysis of Echinacea angustifolia seedlings inoculated with arbuscular mycorrhizal fungi under salt stress. Int. J. Mol. Sci. 2019, 20, 788. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Wu, Q.; Chang, W.; Fan, X.; Song, F. Proteomic analysis of the response of Funnelifor mismosseae/Medicago sativa to atrazine stress. BMC Plant Biol. 2018, 18, 289. [Google Scholar] [CrossRef]
- Rivera-Becerril, F.; van Tuinen, D.; Martin-Laurent, F.; Metwally, A.; Dietz, K.J.; Gianinazzi, S.; Gianinazzi-Pearson, V. Molecular changes in Pisum sativum L. roots during arbuscular mycorrhiza buffering of cadmium stress. Mycorrhiza 2005, 16, 51–60. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.Y.; Martini, M.Y.; Yusuff, O.A.; Ismail, M.R.; Miah, G. Molecular analysis of Hsp70 mechanisms in plants and their function in response to stress. Biotechnol. Genet. Eng. Rev. 2017, 33, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Rui, W.; Mao, Z.; Li, Z. The roles of phosphorus and nitrogen nutrient transporters in the arbuscular mycorrhizal symbiosis. Int. J. Mol. Sci. 2022, 23, 11027. [Google Scholar] [CrossRef]
- Silvia, C.; Loic, C.; Alexis, S.; Annette, N.; Alexander, E.; Daphnée, B.; Ghislaine, R.; Daniel, W.; Christophe, R.; Joachim, K.; et al. Imbalanced regulation of fungal nutrient transports according to phosphate availability in a symbiocosm formed by poplar, sorghum, and Rhizophagus irregularis. Front. Plant Sci. 2019, 10, 1617. [Google Scholar]
- Bücking, H.; Kafle, A. Role of arbuscular mycorrhizal fungi in the nitrogen uptake of plants: Current knowledge and research Gaps. Agronomy 2015, 5, 587–612. [Google Scholar] [CrossRef]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef]
- Daram, P.; Brunner, S.; Rausch, C.; Steiner, C.; Amrhein, N.; Bucher, M. Pht2;1 Encodes a low-affinity phosphate transporter from Arabidopsis. Plant Cell 1999, 11, 2153–2166. [Google Scholar] [CrossRef]
- Guerinot, M.L. The ZIP family of metal transporters. Biochim. Biophys. Acta (BBA) Biomembr. 2000, 1465, 190–198. [Google Scholar] [CrossRef]
- Watts-Williams, S.J.; Cavagnaro, T.R. Arbuscular mycorrhizal fungi increase grain zinc concentration and modify the expression of root ZIP transporter genes in a modern barley (Hordeum vulgare) cultivar. Plant Sci. 2018, 274, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Watts-Williams, S.J.; Tyerman, S.D.; Cavagnaro, T.R. The dual benefit of arbuscular mycorrhizal fungi under soil zinc deficiency and toxicity: Linking plant physiology and gene expression. Plant Soil 2017, 420, 375–388. [Google Scholar] [CrossRef]
- Zhu, J.N.; Huang, H.; Du, Y.; Tang, J.J.; Chen, X. The transgenerational effect of arbuscular mycorrhizal fungi on root phosphatase activity of host plant Medicago truncatula. Chin. J. Ecol. 2022, 41, 912–918. [Google Scholar]
- Wilkens, S. Structure and mechanism of ABC transporters. F1000Prime Rep. 2015, 7, 14. [Google Scholar] [CrossRef]
- Lane, T.S.; Rempe, C.S.; Davitt, J.; Staton, M.E.; Peng, Y.; Soltis, D.E.; Melkonian, M.; Deyholos, M.; Leebens-Mack, J.H.; Chase, M.; et al. Diversity of ABC transporter genes across the plant kingdom and their potential utility in biotechnology. BMC Biotechnol. 2016, 16, 47. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Plant Height (cm) | Branch Number | Mycorrhizal Colonization (%) |
|---|---|---|---|
| NM | 6.21 ± 0.56 | 3.28 ± 0.48 | 0.00 ± 0.00 |
| AMI | 15.07 ± 1.09 *** | 5.28 ± 0.47 *** | 24.63 ± 6.70 *** |
| Sample | Raw Reads | Clean Reads | Clean Bases | Error Rate | Q ≥ 20 | Q ≥ 30 | GC pct |
|---|---|---|---|---|---|---|---|
| NM_1 | 90483268 | 86062588 | 6.45G | 0.02 | 98.36 | 95.09 | 42.54 |
| NM_2 | 95951276 | 91312188 | 6.85G | 0.02 | 98.43 | 95.1 | 43.14 |
| NM_3 | 96105596 | 91035756 | 6.83G | 0.02 | 98.46 | 95.16 | 43.19 |
| AMI_1 | 84660684 | 79156164 | 5.94G | 0.02 | 98.48 | 95.34 | 41.29 |
| AMI_2 | 90570192 | 85901912 | 6.44G | 0.02 | 98.29 | 94.73 | 42.66 |
| AMI_3 | 96210968 | 90386296 | 6.78G | 0.02 | 98.38 | 94.96 | 42.63 |
| Gene Function Annotated by KO | Number | log2FC |
|---|---|---|
| Glutathione S-transferase | 17 (11) | 1.35–16.22 |
| Heat shock 70 kDa protein 1/8 | 9 (9) | 5.31–10.53 |
| Heat shock transcription factor, other eukaryote | 7 (7) | 5.68–10.03 |
| Stress-induced phosphoprotein 1 | 5 (5) | 5.71–10.39 |
| Heat shock 70 kDa protein 4 | 4 (3) | 1.05–9.01 |
| Heat shock 70 kDa protein 5 | 4 (4) | 6.06–11.06 |
| Superoxide dismutase, Fe-Mn family | 4 (4) | 5.22–8.54 |
| Superoxide dismutase, Cu-Zn family | 4 (4) | 6.8–10.72 |
| Glutathione peroxidase | 3 (3) | 7.14–11.63 |
| Glutathione reductase (NADPH) | 2 (2) | 8.65–8.98 |
| Glutathione gamma-glutamylcysteinyltransferase | 2 (1) | 1.76–5.9 |
| Peroxidase | 2 (0) | 2.58–4.36 |
| Glutathione reductase (NADPH) | 2 (2) | 8.65–8.98 |
| Glutathione synthase | 2 (2) | 6.37–7.41 |
| Heat shock protein 90 kDa beta | 2 (2) | 6.85–8.46 |
| HSP20 family protein | 1 (1) | 5.63 |
| Gene Function Annotated by KO | Number | log2FC |
|---|---|---|
| Solute carrier family 30 (zinc transporter), member 2 | 6 (6) | 5.07–9.37 |
| Zinc transporter, ZIP family | 4 (4) | 6.41–8.11 |
| Solute carrier family 39 (zinc transporter), member 1/2/3 | 2 (1) | 4.41–5.38 |
| MFS transporter, PHS family, inorganic phosphate transporter | 5 (4) | 1.28–15.42 |
| Solute carrier family 20 (sodium-dependent phosphate transporter) | 3 (2) | 5.03–12.21 |
| Phosphate transporter | 2 (2) | 8.57–8.98 |
| MFS transporter, ACS family, solute carrier family 17 (sodium-dependent inorganic phosphate cotransporter), other | 1 (1) | 8.65 |
| Ammonium transporter, Amt family | 5 (1) | 2.52–14.69 |
| MFS transporter, NNP family, nitrate/nitrite transporter | 4 (2) | 1.8–9.54 |
| Solute carrier family 7 (cationic amino acid transporter), member 1 | 1 | 11.59 |
| Sulfate transporter 1, high affinity | 1 (1) | 6.82 |
| V-type H+-transporting ATPase | 19 (19) | 6.28–10.7 |
| H+-transporting ATPase | 4 (3) | 2.52–12.71 |
| Mitochondrial ABC transporter ATM | 6 (6) | 5.18–9.21 |
| Ca2+-transporting ATPase | 17 (16) | 2.41–10.52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, X.-R.; Pan, J.-F.; Zhao, M.; Guo, X.-Y.; Wang, Q.; Zhang, L.; Liu, W. Transcriptome Analysis Reveals the Impact of Arbuscular Mycorrhizal Symbiosis on Toona ciliata var. pubescens Seedlings. Forests 2024, 15, 673. https://doi.org/10.3390/f15040673
Jiang X-R, Pan J-F, Zhao M, Guo X-Y, Wang Q, Zhang L, Liu W. Transcriptome Analysis Reveals the Impact of Arbuscular Mycorrhizal Symbiosis on Toona ciliata var. pubescens Seedlings. Forests. 2024; 15(4):673. https://doi.org/10.3390/f15040673
Chicago/Turabian StyleJiang, Xue-Ru, Jian-Feng Pan, Ming Zhao, Xiao-Yan Guo, Qiong Wang, Lu Zhang, and Wei Liu. 2024. "Transcriptome Analysis Reveals the Impact of Arbuscular Mycorrhizal Symbiosis on Toona ciliata var. pubescens Seedlings" Forests 15, no. 4: 673. https://doi.org/10.3390/f15040673