Abstract
Diaspore dispersal is crucial in shaping plant population dynamics, biodiversity, and ecosystem functions. The effect of the vegetation structure on the secondary wind dispersal of diaspores with different appendage types is not well understood. Using a wind tunnel and a high-definition video camera, we accurately measured the lift-off and dispersal velocities of diaspores from sixteen plant species across six wind velocities (2, 4, 6, 8, 10, and 12 m s−1) under six simulated vegetation structures. Vegetation structure and appendage type were pivotal factors, explaining 41.1% and 42.3% of the variance in lift-off velocity and accounting for 12.0% and 25.3% of the variability in diaspore dispersal velocity, respectively. Vegetation coverage was the main factor influencing near-surface wind velocity, and the lift-off and dispersal velocities of diaspores changed significantly when vegetation coverage exceeded 40%. Diaspores with one wing, having high lift-off velocities and low dispersal velocities, adopt the anti-long-distance wind dispersal strategy, whereas diaspores with pappus, having low lift-off velocities and high dispersal velocities, adopt the long-distance wind dispersal strategy. In contrast, diaspores with thorn, discoid, balloon, and four wings adopt the non-long-distance wind dispersal strategy, suitable for environments such as low-coverage deserts and desert steppes but not high-coverage typical steppes. This study could help comprehend the effect of the vegetation structure on the dispersal process of diaspores, which facilitate habitat restoration and biodiversity conservation of grassland and forest ecosystems.
1. Introduction
Diaspore dispersal plays a crucial role in the formation and development of plant populations and communities [1,2,3]. Wind diaspore dispersal proceeds in two phases: initial aerial movement (primary dispersal) and subsequent ground movement (secondary dispersal) [4,5,6,7]. Secondary dispersal is a pivotal process influencing diaspore movement in diverse open habitats [8,9], including deserts [5,10], temperate steppes [11], coastal and post-fire landscapes [4], seashore meadows [12], snow-covered areas [13,14], as well as in urban areas and on roads [15,16]. The efficacy of secondary wind diaspore dispersal is strongly influenced by factors such as vegetation structure and diaspore morphology [8,16,17,18].
Vegetation structure plays a crucial role in modifying wind velocity near the ground surface [4,17]. The two primary factors defining vegetation structure are coverage and height. Coverage predominantly influences the horizontal wind velocity spread, whereas height determines its vertical reach near the ground surface [19]. For example, dense vegetation coverage can obstruct wind flow, thereby reducing diaspore mobility. Conversely, sparse coverage often facilitates greater dispersal distances owing to increased airflow [20]. Despite these insights, the specific ways in which the vegetation structure influences secondary diaspore dispersal by wind require further investigation.
Diaspore morphology is crucial in wind dispersal. Key morphological traits, including shape, size, and appendage type, significantly alter the aerodynamic properties of diaspores, thereby influencing their movement [21]. Researchers assume that these morphological traits adapt to specific wind dispersal modes, increasing the probability and distance of secondary dispersal [22,23]. However, research on the effect of diaspore morphology on secondary dispersal across various vegetation structures is limited.
The lift-off (LV) and diaspore velocities (DV) play crucial roles in the process of secondary wind dispersal. Lift-off velocity serves as a reliable indicator for evaluating the initial movement capability of diaspores and is defined as the critical wind velocity necessary for their initial movement [24]. Diaspore morphological characteristics significantly influence LV [21]. Diaspore velocity is the ability of diaspores to move when wind velocity exceeds the LV [25]. Thus, both LV and DV play significant roles in determining the distance traveled by diaspores in different vegetation structures [26,27].
With the intensification of grassland and savanna desertification, trees and shrubs invade into nature grassland, resulting in habitat fragmentation, forming different spatially heterogeneous patches of herb, shrub–herb, and tree–shrub–herb vegetation structures [28,29]. Therefore, the process of seed secondary wind dispersal in different patches is important for plant colonization, community regeneration, and plant diversity restoration [30,31,32,33]. However, the effects of different vegetation structures on the LV, DV, dispersal distance, and distribution pattern remain unclear.
This study aimed to determine (a) the impact of varying vegetation structures on the LV and DV during secondary wind dispersal and (b) the influence of diaspore morphological traits on the LV and DV across different vegetation structures. We hypothesized that vegetation coverage is a primary factor influencing wind velocity, which in turn affects the LV and DV. Previous studies suggested that lighter diaspores, especially those with wings and pappus, typically exhibit lower lift-off velocities and higher dispersal speeds across various vegetation types [25]. Consequently, we predicted that diaspore morphology might serve as a secondary factor in determining lift-off and dispersal velocities during secondary wind dispersal.
2. Materials and Methods
2.1. Diaspore Selection and Measurement
Our study utilized diaspores from 16 species, and they were collected from grassland and riparian woodland in Inner Mongolia (Table 1). We randomly collected 50 mature and intact diaspores from 10 individual plants per species, obtaining 500 diaspores per species. The diaspores were air-dried and stored in the laboratory for experiments.

Table 1.
Diaspore traits across 16 selected species (mean ± standard deviation). Different letters indicate significance differences between vegetation structures (p < 0.05).
For each species, we randomly selected 20 diaspores from the 500 diaspores to determine diaspore traits. The evaluated traits included mass, length, width, height, projected area, terminal velocity, shape index, and wing loading. The mass (M) of each diaspore was precisely determined using an electronic balance (precision of 0.01 g). The length (longest axis), width (intermediate axis), and height (shortest axis) were measured using a Vernier caliper (precision of 0.01 mm). The projected area of each diaspore was calculated using the Python software library (Open-Source Computer Vision Library) after scanning a three-dimensional diaspore with a digital scanner [24]. The terminal velocity of diaspores was determined via a video camera recording method in a settling tower. A video camera (GC-P100AC, JVC Japan, Yokohama, Japan) was used to record diaspore free fall, with 50 frames per second. The recorded video was analyzed for the observation and calculation of the terminal velocity of the diaspore [34]. The shape index (SI) of each diaspore was calculated using the methodology established by Thompson [35]:
Wing loading, which is the mass of the diaspore divided by its projected area, was calculated as follows [36]:
2.2. Wind Tunnel
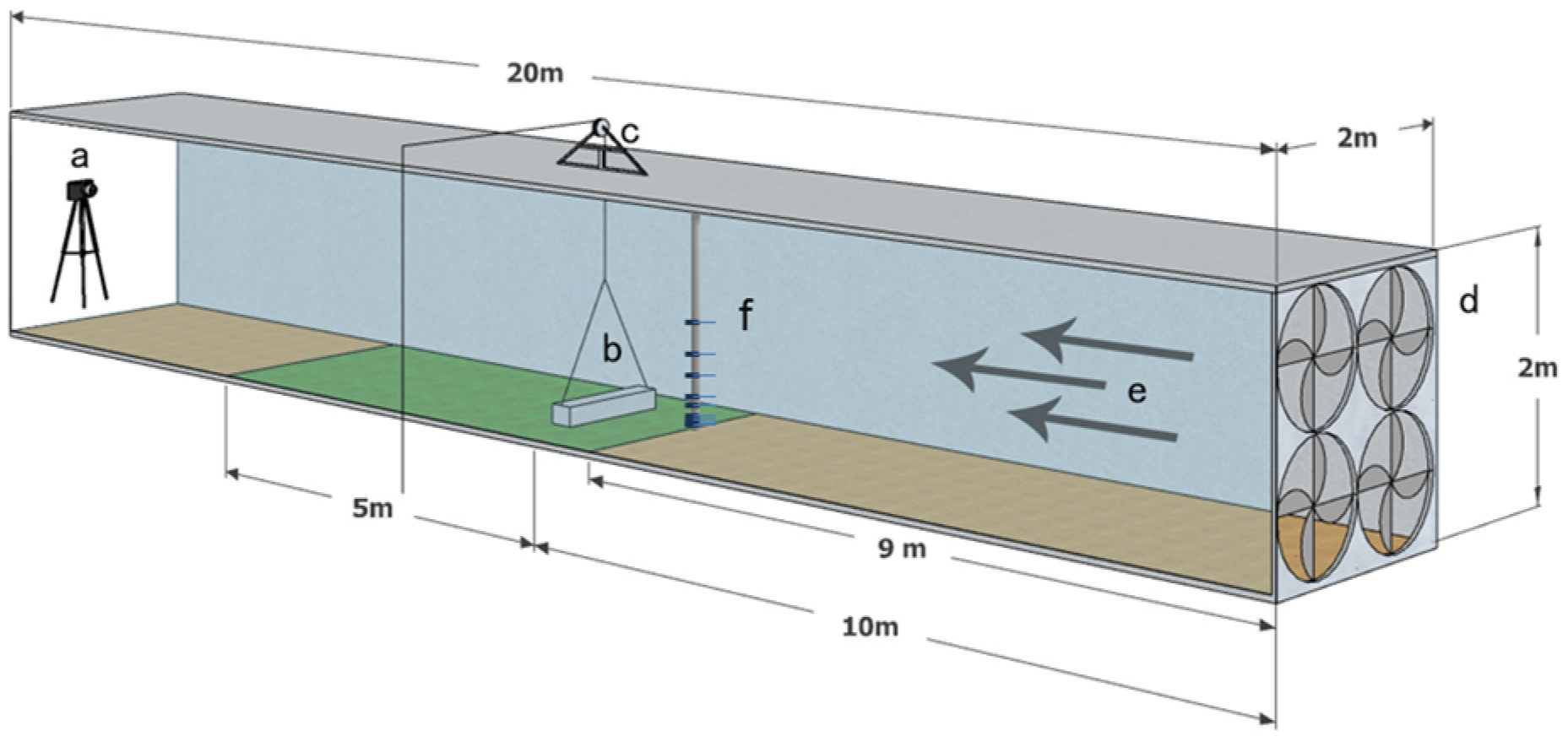
Our experiment was carried out in a wind tunnel measuring 2 m in height and width and 20 m in length. The tunnel included a 10 m transition section and a 5 m experimental section situated 10 m from the air outlet (Figure 1). The large wind tunnel was crucial for ensuring physical similarity conditions, including geometric, kinematic, and dynamic similarity, during diaspore dispersal [24,37].
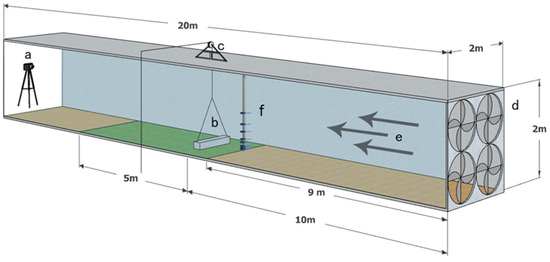
Figure 1.
Diagram of the wind tunnel used in the experiments: a, camera; b, diaspore cover; c, pulley system; d, fan assembly; e, wind direction; f, hot-wire anemometers.
Other apparatus equipped in the wind tunnel included a pulley, camera, and anemometers system. The pulley system comprised a pulley, a rope, and a diaspore cover. The pulley was securely mounted atop a sturdy, tripod-like structure. The rope was used to lift the diaspore cover through a pulley to measure the DV of the diaspores. The camera (DCR-SR100, Sony, Tokyo, Japan) was mounted on a tripod at the wind tunnel outlet for real-time monitoring of diaspore movement. The anemometers system monitored wind velocity using hot-wire anemometers (AirPro Velocity Meter AP500, TSI Incorporated, Shoreview, MN, USA) fixed at different heights above the ground in the middle of the wind tunnel in front of the experimental section.
2.3. Simulation of Vegetation Structures
We used plastic artificial grasses, shrubs, and a combination of the two to simulate the desert, typical, and meadow steppes. The grasses were simulated by short plastic artificial Stipa tianschanica var. gobica (Roshev.) P.C.Kuo & Y.H.Sun (165.6 cm2 crown area and 15 cm high) and tall plastic artificial Pennisetum alopecuroides Ham., Prodr. Pl. Ind. Occid (724 cm2 crown area and 60 cm high). The shrubs were simulated using plastic artificial Ammopiptanthus mongolicus (Maxim.) S.H.Cheng (1962 cm2 crown area and 60 cm height). We calculated the density of short grass, high grass, and shrub in different vegetation structures according to their single coverage. Top-view photographs and Adobe Photoshop were used to measure the vegetation coverage in order to obtain the correct density of plants. We used bare ground covered with aeolian sandy soil as a control (V0), randomly distributed short grass (12.1 m−2 density, 15 cm mean height, and 20% coverage) to simulate sparse low-grass desert steppe (V1), randomly distributed tall grass (density 2.8 m−2, 60 cm mean height, and 20% coverage) to simulate sparse high-grass desert steppe (V2), a randomly distributed short grass (density 7.3 m−2) and shrub (density 0.3 m−2) combination (with a mean height of 17 cm and coverage of 20%) to simulate sparse shrub–grass desert steppe (V3), a tall grass (density 2.4 m−2) and short grass (density 19 m−2) combination (with a mean height of 20 cm and coverage of 40%) to simulate the typical steppe (V4), and a short grass (density 79 m−2) and tall grass (density 22.6 m−2) combination (with a mean height of 25 cm and coverage of 80%) to simulate the meadow steppe (V5).
2.4. Measurement of Wind Profiles
The wind velocity at the central axis of the wind tunnel (1 m distance from the side walls and floor) was the highest free-stream wind velocity because it was the least affected by the side walls. Nine hot-wire anemometers were fixed above the tunnel floor at 1, 3, 7, 10, 20, 30, 50, 70, and 100 cm to record wind velocities (5 duplicates, once every 2–3 s) in six vegetation structures [25]. The data of wind velocity were automatically transmitted to a wirelessly connected computer. In total, there were 24 different wind profiles corresponding to free-stream wind velocities of 4, 6, 8, and 10 m s−1 across 6 vegetation structures.
2.5. Measurement of Diaspore Lift-Off Velocity across Six Vegetation Structures
A row of 10 diaspores from the same species, spaced 3 cm apart and 50 cm away from the sidewalls, was placed in each vegetation structure. Diaspores were continuously monitored as wind velocity gradually increased from 0 m s−1. When more than two diaspores began to move, the specific wind velocity was recorded as the LV. The experiment was repeated five times for each species within the vegetation structure.
2.6. Measurement of Diaspore Dispersal Velocity in Six Vegetation Structures
A row of 10 diaspores from the same species, spaced 3 cm apart, was positioned centrally in the test section. The free-stream wind velocity was set to 2, 4, 6, 8, 10, and 12 m s−1 to measure the dispersal velocity of diaspores [38]. Prior to reaching the pre-set wind velocity, the diaspores were shielded with a cover. The cover was lifted as soon as the wind reached a predetermined velocity. Diaspore movement was recorded via camera upon being exposed to airflow. For each species, five replicate tests were conducted for each vegetation structure at each specified wind velocity.
2.7. Data Analysis
The videos captured by the camera were analyzed frame-by-frame using the Adobe Premiere Pro software (18.0) [39]. Diaspores can enter one of three final states: (a) remaining stationary, (b) crossing the finish line, or (c) stopping before the finish line. In the first scenario, where the diaspores remained stationary, their velocity was recorded as 0 m s−1. The diaspore velocity was determined using the following formula for diaspores that crossed the finish line:
where DV is the diaspore velocity, L is the length of the experimental section (5 m), and T is the movement time of the diaspore.
For diaspores that stopped before the finish line (third scenario), we determined their movement distance and time through video analysis. Our analysis revealed that the movement time for these diaspores varied between 0.42 s and 12 s. Consequently, we defined the diaspore velocity in this scenario as the ratio of the moving distance to the longest observed movement time (12 s).
Redundancy analysis was conducted using Canoco 5.0 (version 5.0, Microcomputer Power, Ithaca, NY, USA) to determine the relationship between the explanatory variables of wind velocity, appendage type (AT), terminal velocity (TV), projected area (PA), wing loading (WL), shape index (SI), mass (M), and vegetation structure (VS), and the response variables of the lift-off velocity (LV) and diaspore velocity (DV). To address multicollinearity, we removed explanatory variables that exhibited significantly high variance inflation factors (VIF > 10).
Because appendage type and vegetation structure are not scaled attributes, seven variables were defined for the appendage type occurrence: A1, A2, A3, A4, A5, A6, and A7. Similarly, six vegetation structure variables were defined as V0, V1, V2, V3, V4, and V5. The Ai variable was assigned a value of 1 if the appendage type corresponded to the following: balloon (A1), discoid (A2), four wings (A3), pappus (A4), one wing (A5), thorn (A6), or two wings (A7); otherwise, Ai was 0. The vegetation structure variables had the same assignment as the appendage type.
Descriptive statistical analysis and analysis of variance (ANOVA) were performed using Predictive Analytics Software (PASW 18.0 IBM Corp., Armonk, NY, USA). Histograms were generated using Python (3.9.10).
3. Results
3.1. Diaspore Traits across 16 Selected Species
Eight morphological and wind dynamics indicators of the sixteen species of diaspores were significantly different (p < 0.01). According to appendage types, the diaspores was classified into 7 groups, those with one wing (3 species), pappus (2 species), thorn (3 species), two wings (1 species), discoid (3 species), balloon (2 species), and four wings (2 species). Diaspores with one wing were characterized by a high shape index (SI), terminal velocity (TV), and wing loading (WL). In contrast, diaspores with pappus showed a low SI, TV, and WL. Diaspores with thorn, two wings, discoid, balloon, and four wings generally showed a low SI, and a high TV and WL. Except for the group with two wings, most of the diaspore indicators within the groups presented significant differences (p < 0.01); however, the variation of indicators within a group was generally lower than the total variation across all groups (Table 1).
3.2. Wind Profiles across Six Vegetation Structures under Different Wind Velocities
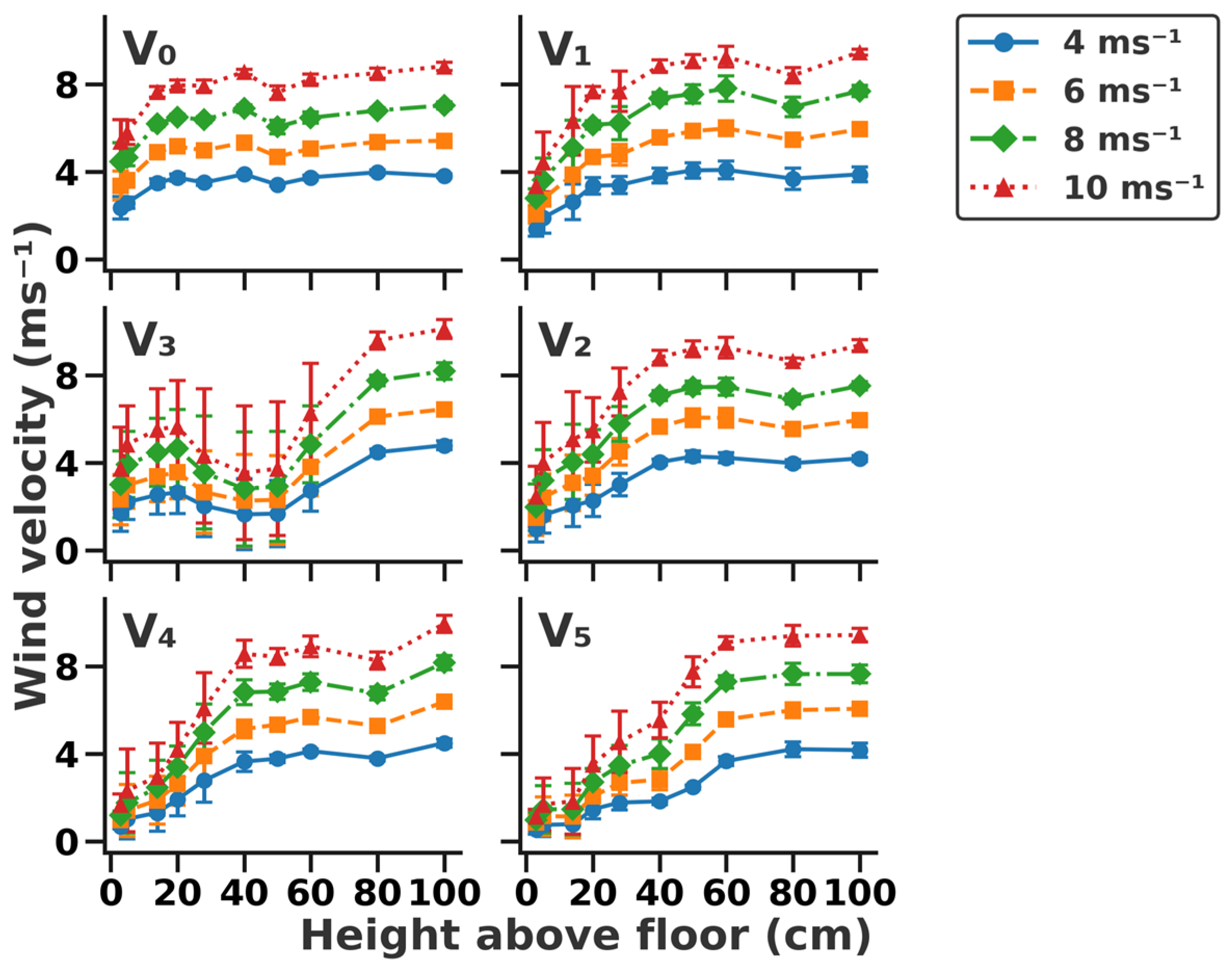
Wind profiles for all vegetation structures under the tested wind velocities are shown in Figure 2. The thickness of the boundary layer (TBL) on bare ground was 20 cm above the ground and was mainly influenced by the vegetation type present on the surface. The TBL varied with different vegetation structures: 40 cm for V1, 80 cm for V3, 50 cm for V2, and 60 cm for both V4 and V5. In particular, the TBL of the vegetation structure with shrubs (V3) was higher than that with grass. Vegetation coverage significantly influenced wind velocity near the ground surface. For instance, wind velocities were 2.3, 3.4, 4.4, and 5.4 ms−1 at 3 cm above the ground on V0, corresponding to those of 4, 6, 8, and 10 m s−1 at 100 cm above the ground (p = 0.015), respectively. Wind velocities decreased to 1.4, 2.1, 2.8, and 3.4 m s−1 in V1 at 3 cm above the ground. Furthermore, wind velocities were 0.6, 0.9, 1.0, and 1.1 ms−1 in V5 at a height of 3 cm above the ground.
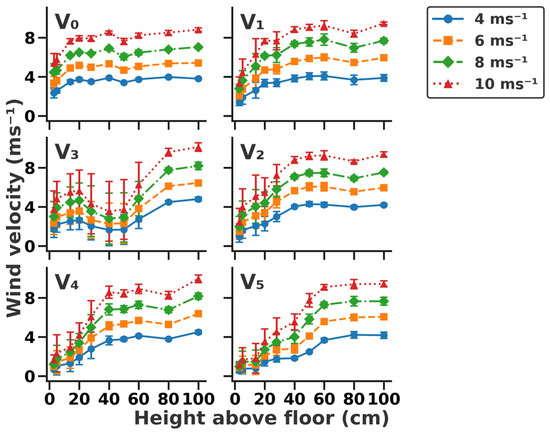
Figure 2.
Wind profiles across six vegetation structures under different wind velocities. (V0) Bare ground, (V1) simulated sparse low-grass desert steppe, (V2) simulated sparse high-grass desert steppe, (V3) simulated sparse shrub–grass desert steppe, (V4) simulated typical steppe, and (V5) simulated meadow steppe.
3.3. Lift-Off Velocity of Seven Diaspore Types across Six Vegetation Structures
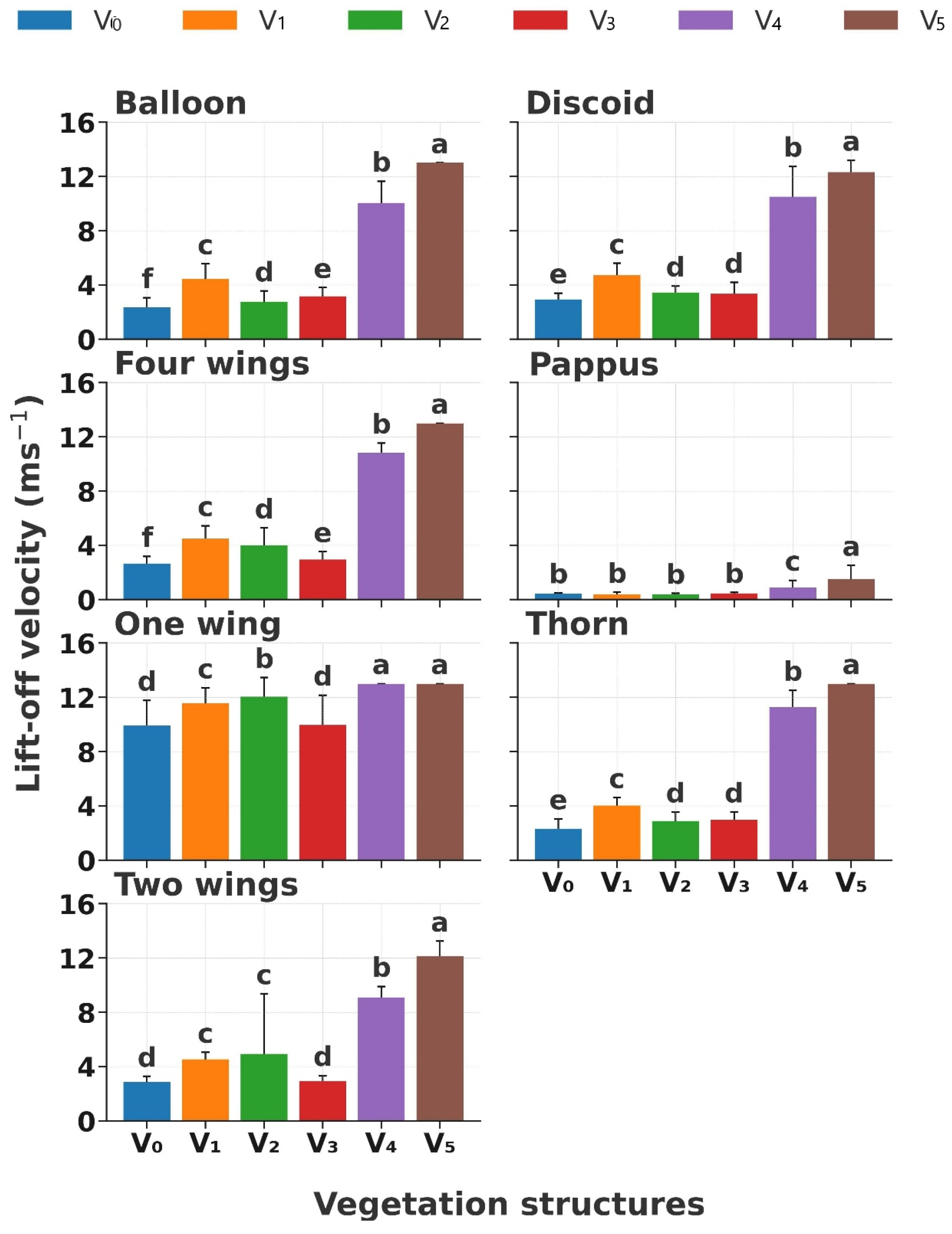
Vegetation structure significantly influenced diaspore lift-off velocity (LV, p < 0.01; Figure 3). The LV increased with increasing vegetation coverage in the following order: V0, V3, V2, V1, V4, and V5 (p < 0.01); especially, the LV showed a conspicuous increase when the coverage was higher than 40% (Figure 3). Significant differences in LVs were observed according to diaspore appendage types (p < 0.01). Diaspores with balloon, discoid, four wings, thorn, and two wings experienced a slight rise in LV (p < 0.05, from 3 m s−1 to 5 m s−1) from V0, V3, V2, to V1 when grass coverage < 20%; however, the LV increased to 10 m s−1 to 14 m s−1 when the coverage was higher than 40% (Figure 3). Nevertheless, the LV of diaspores with pappus did not exhibit a slight variation (p < 0.05, from 0.5 m s−1 to 2 m s−1) across all the vegetation structures. In contrast, diaspores with one wing showed the highest LV (from 10 m s−1 to 14 m s−1) across all the vegetation structures.
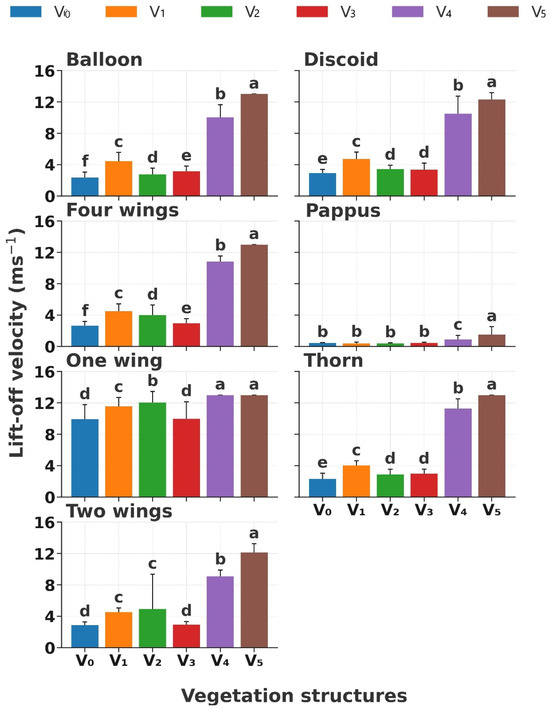
Figure 3.
Lift-off velocity of seven types of diaspores across six vegetation structures. V0, bare ground; V1, simulated sparse low-grass desert steppe; V2, simulated sparse high-grass desert steppe; V3, simulated sparse shrub–grass desert steppe; V4, simulated typical steppe; V5, simulated meadow steppe. Different letters indicate significance differences between the lift-off velocities of each appendage type across different vegetation structures (p < 0.05).
3.4. Relationships between Diaspore Lift-Off Velocity and Explanatory Factors
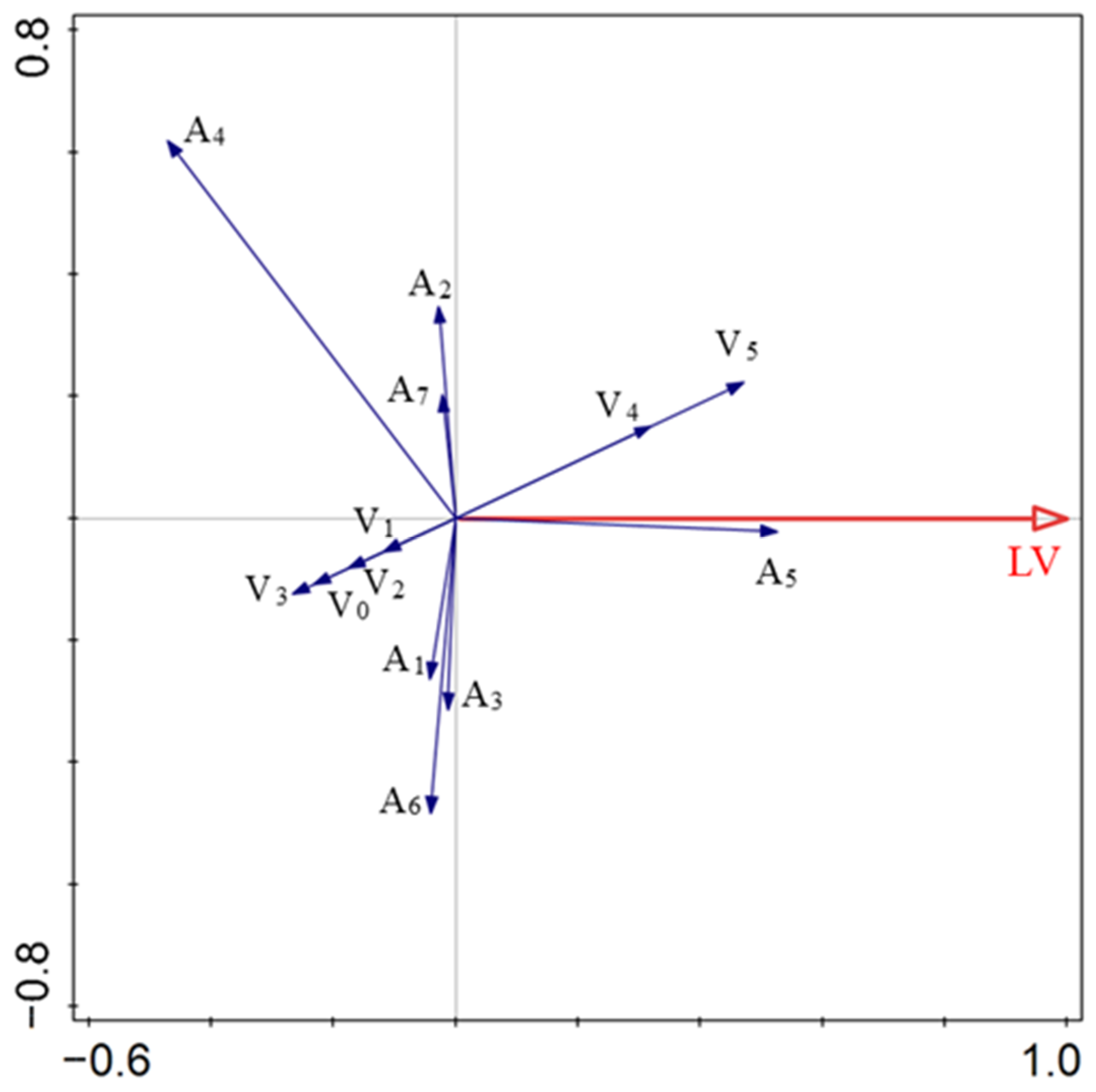
Vegetation structure, appendage type, and diaspore traits explained 83.5% of the variation in LV. A1, A3, A6, A4, A2, A7, V3, V0, V1, and V2 had negative effects on the LV. A5, V4, and V5 positively correlated with the LV. The four factors with the highest levels of explained variation were A5, V5, V4, and A4, which accounted for 27.7%, 22.2%, 17.9%, and 14.6% of the total variation, respectively (Figure 4; Table 2).
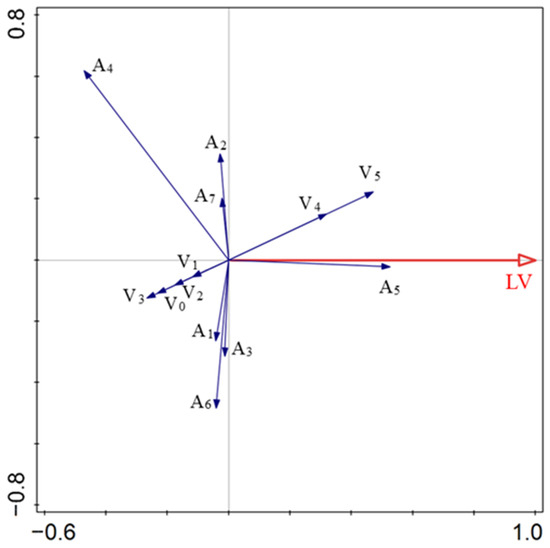
Figure 4.
Relationships between diaspore lift-off velocity and explanatory factors. V0, bare ground; V1, simulated sparse low-grass desert steppe; V2, simulated sparse high-grass desert steppe; V3, simulated sparse shrub–grass desert steppe; V4, simulated typical steppe; V5, simulated meadow steppe; LV, lift-off velocity; A1, balloon; A2, discoid; A3, four wings; A4, pappus; A5, one wing; A6, thorn; A7, two wings.
Table 2.
Variation in diaspore lift-off velocity explained by vegetation structure, diaspore appendage type, diaspore morphology, and wind dynamic traits for all species using redundancy analysis.
3.5. Diaspore Velocities by Appendage Type in Different Vegetation Structures
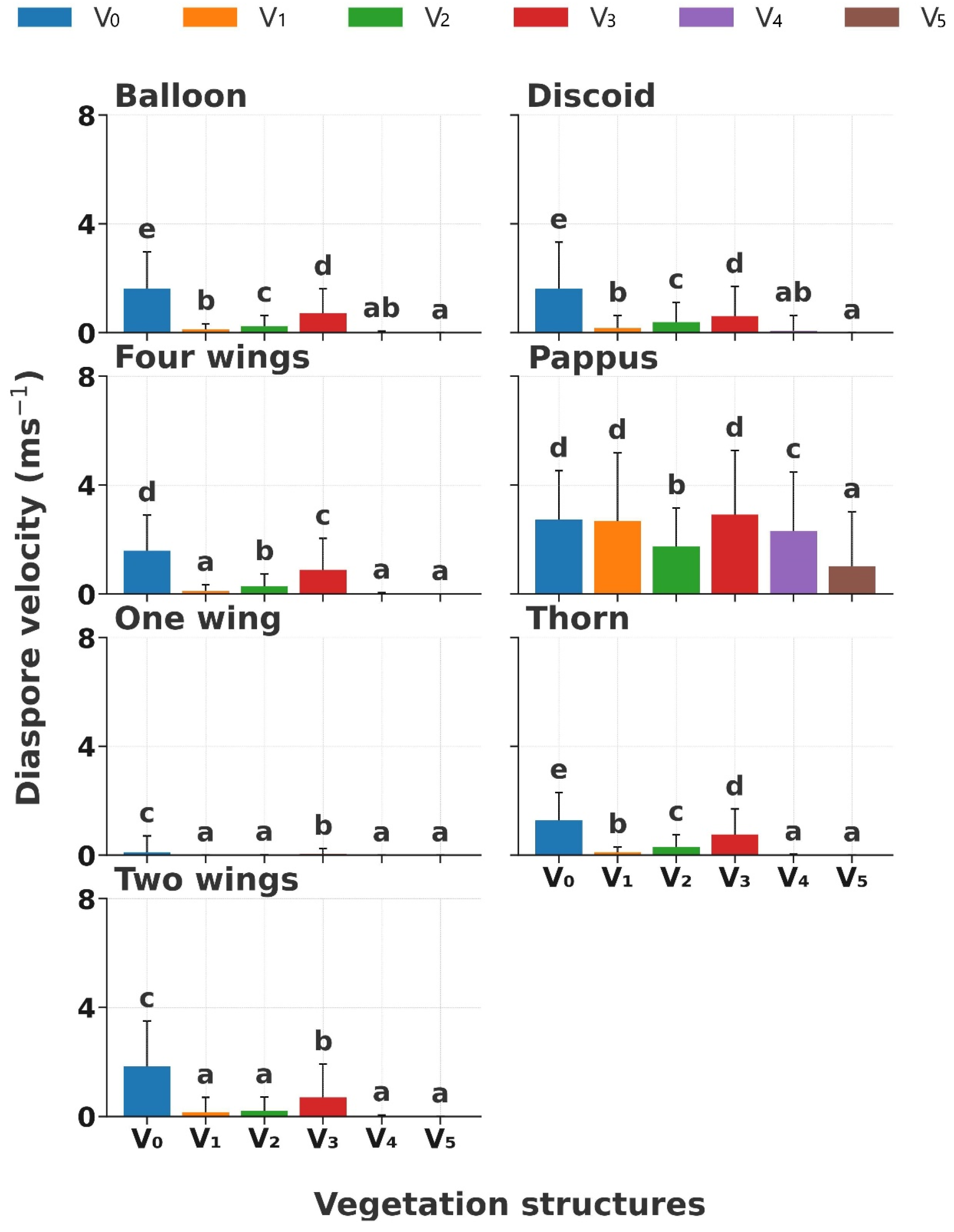
The diaspore velocity (DV) significantly decreased with increasing vegetation coverage in the order of V0, V3, V2, V1, V4, and V5 (p < 0.01); especially, the DV significantly decreased when the coverage was >40% (Figure 5). Significant variations in DVs were observed in different diaspore appendage types (p < 0.01). The DV of diaspores with balloon, discoid, four wings, thorn, and two wings showed a significant decrease (p < 0.05, from 2 m s−1 to 0.3 m s−1) from V0, V3, V2, to V1 when grass coverage < 20%; however, the DV tended to 0 m s−1 when the coverage was higher than 40% (Figure 5). Nevertheless, the DV of diaspores with pappus had a high velocity and a significant variation (p < 0.05, from 1 m s−1 to 3 m s−1) across all the vegetation structures. In contrast, diaspores with one wing showed the lowest DV (from 0 m s−1 to 0.1 m s−1) across all the vegetation structures.
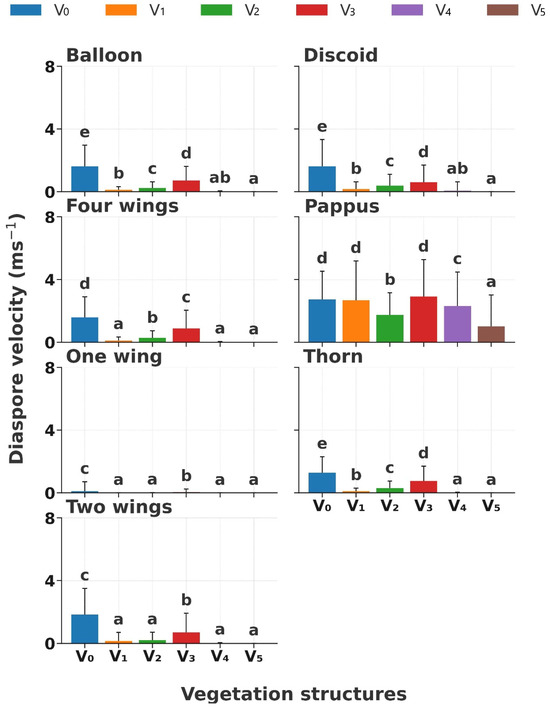
Figure 5.
Diaspore velocity of seven diaspore appendage types across six vegetation structures. V0, bare ground; V1, simulated sparse low-grass desert steppe; V2, simulated sparse high-grass desert steppe; V3, simulated sparse shrub–grass desert steppe; V4, simulated typical steppe; V5, simulated meadow steppe. Different letters indicate significance differences between the diaspore velocities of each appendage type across different structures of vegetation (p < 0.05).
3.6. Relationships between Diaspore Velocity and Explanatory Factors
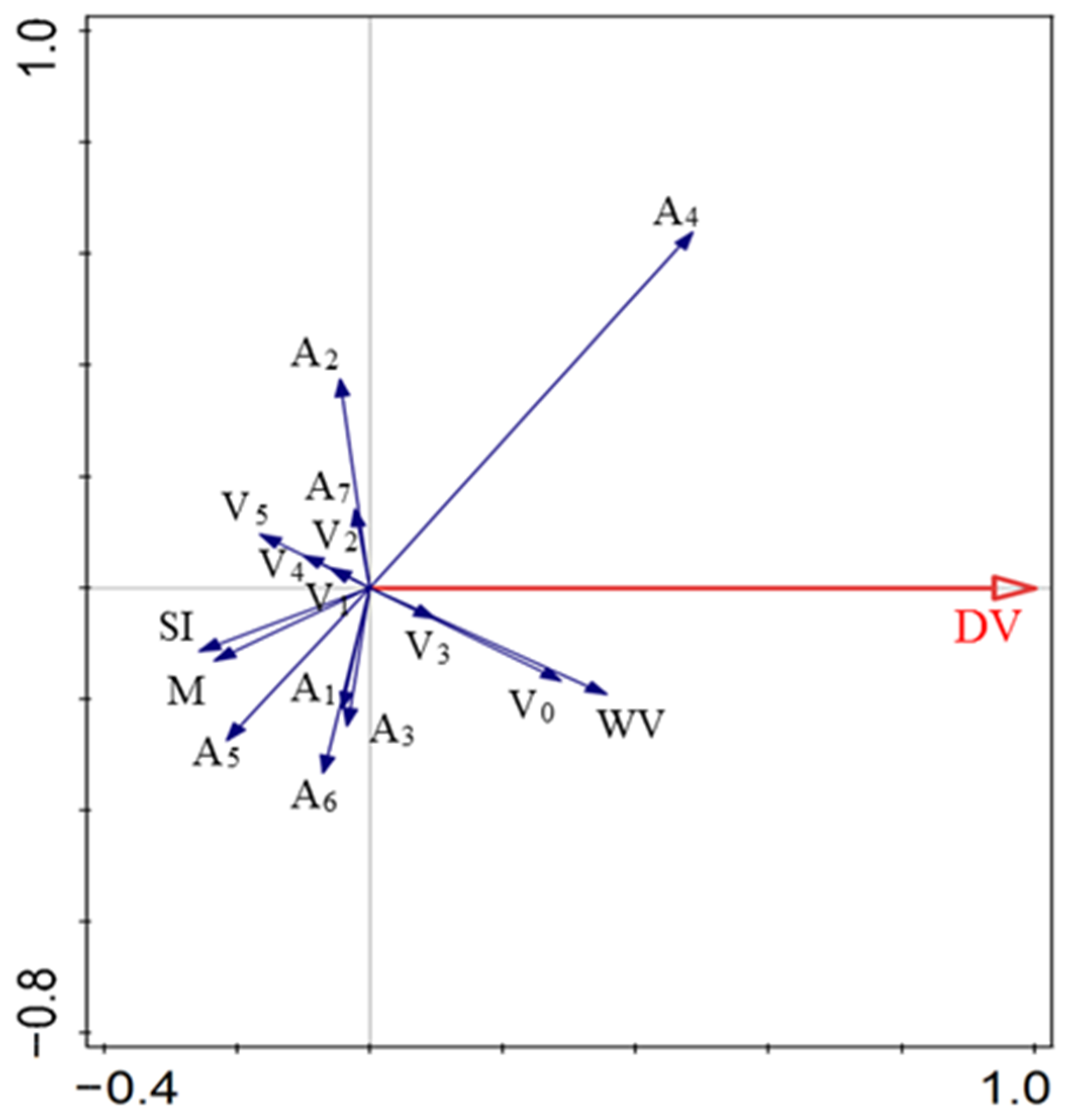
The combination of wind velocity, vegetation structure, and diaspore appendage type explained 49.5% of the variation in the diaspore velocity. A4, V3, V0, and wind velocity positively affected DV. A1, A2, A3, A5, A6, V4, V3, V1, V2, V5, M, and SI were negatively correlated with DV. A4, wind velocity, and V0 accounted for 23.6%, 12.7%, and 8.3% of the total variation, respectively, and contributed 47.1%, 25.3%, and 16.6%, respectively (Figure 6; Table 3).
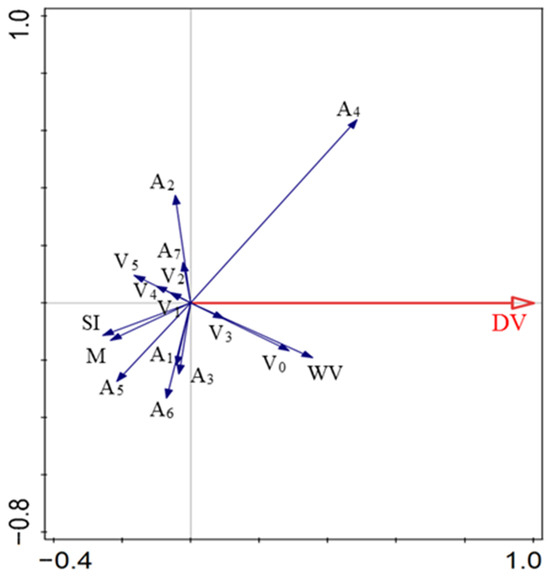
Figure 6.
Relationships between diaspore velocity and explanatory factors. V0, bare ground; V1, simulated sparse low-grass desert steppe; V2, simulated sparse high-grass desert steppe; V3, simulated sparse shrub–grass desert steppe; V4, simulated typical steppe; V5, simulated meadow steppe; DV, diaspore velocity; A1, balloon; A2, discoid; A3, four wings; A4, pappus; A5, one wing; A6, thorn; A7, two wings; SI, shape index; M, mass; WV, wind velocity.
Table 3.
Variation in diaspore velocity explained by vegetation structure, diaspore appendage type, morphology, and wind dynamic traits in all species using redundancy analysis.
4. Discussion
4.1. Effect of Vegetation Structure on Diaspore Lift-Off Velocity with Different Morphological Characteristics
The results showed that the taller the vegetation, the thicker the boundary layer, and the greater the vegetation coverage, the lower the near-surface wind velocity (Figure 2). Furthermore, secondary wind dispersal is known to be primarily influenced by near-surface wind velocity [40]; thus, the effect of vegetation coverage on the secondary wind dispersal of diaspores is greater than that of vegetation height. Our findings revealed that vegetation structure was the primary contributor to the LV, accounting for nearly 50% of the variance (Table 2), and that the LV significantly increased with increasing vegetation coverage (Figure 3). This increase in LV can be attributed to two factors. Firstly, the reduction in near-surface wind velocity with denser vegetation coverage increased the LV. Our results support previous conclusions that vegetation coverage is the most important factor affecting the lift-off velocity (LV) of seeds [37]. Secondly, the physical obstruction caused by plants or the surface matrix, which hinders diaspore movement [24,25,40,41,42,43]. Our results showed that diaspores with balloon, discoid, thorn, two wings, and four wings exhibited lower lift-off velocities in V3 than in V2. Possible reasons might be that the sparse shrubs could potentially create areas of accelerated airflow. The obstruction from the vegetation canopy might increase the wind speed at the ground surface, thereby reducing the lift-off velocity needed for the diaspores. We also found that when vegetation coverage was more than 40% (close to the coverage of a typical steppe), the LV noticeably increased (>10 m s−1; Figure 4), suggesting that secondary wind dispersal of diaspores is more likely to occur in open habitats, such as bare ground (active sand dunes) and desert steppes [25].
Appendage type significantly influenced LV, contributing nearly 50%, a factor only surpassed by vegetation structure (Table 2). This finding is consistent with previous reports [37]. Diaspores with pappus had a much lower LV than other types of diaspores (Figure 4) because of their low mass, slow terminal velocity, and small wing loading, which promote dispersion [44,45]. However, diaspores with one wing had a much higher LV than other types of diaspores (Figure 4), likely because of their greater mass, terminal velocity, wing loading, long and flat shape, and minimal wind resistance due to the reduced surface area facing the wind when resting on the ground [46]. Diaspores with thorns, discoid, balloons, and four wings had lower lift-off velocities in low-coverage (≤20%) vegetation structures, whereas in high-coverage (≥40%) vegetation structures, their lift-off velocities rapidly increased (Figure 4). Several factors influence the lift-off wind velocities of these types of diaspores. Disc-shaped diaspores exhibit low lift-off velocities because of their overall lightness, which makes them susceptible to wind forces. Those with thorns, balloons, and four wings have low lift-off velocities because of their near-spherical shape, which allows them to easily roll on the ground despite their high mass, terminal velocity, and wing loading [47]. Secondly, the three-dimensional structures of these diaspores provide a larger surface area exposed to horizontal wind, which allows them to be moved more readily by the wind. However, as vegetation cover thickens, the wind velocity near the surface significantly decreases, reducing the pulling force of the wind on the diaspores [37]. Consequently, diaspores with these types of appendages are likely more suited for secondary wind dispersal in low-coverage environments, such as deserts and desert steppes [48,49].
4.2. Effect of Vegetation Structure on Diaspore Dispersal Velocity with Different Morphological Characteristics
Appendage type, vegetation structure, and wind velocity were the primary factors influencing diaspore movement velocity (Table 3). Our findings are consistent with previous findings [50,51,52]. However, the contribution of these three factors to the DV was less than 50% (Table 3), indicating that other factors, such as the soil matrix, initial angle of diaspores, and movement mode (i.e., creeping, saltation, or rolling), may also affect diaspore dispersal velocity (DV) [4,36]. In this study, the most significant factor contributing to DV was diaspore appendage type, which differs from previous results suggesting that the most important factor was wind speed [25]. One reason might be that the study was only focusing on the effect of the surface matrix without vegetation. Additionally, different results gained by previous studies may be attributed to the limited variety and diversity of diaspores utilized [53]. Many studies suggest that using few diaspore types in wind dispersal research may cause errors or biases in the results, as such an experimental design does not fully reflect the complexity of wind dispersal [54,55,56,57,58,59].
Wind velocity was the second most important factor affecting DV (Table 3), indicating that it is the driving force for diaspore movement [25,38]. Generally, there was a positive correlation between wind velocity and DV, which is also consistent with previous research findings [25]. Vegetation structure was the third most significant factor affecting DV (Table 3), and DV decreased as vegetation coverage increased (Figure 6). These results indicated that increased vegetation coverage not only decreased wind velocity in the soil surface [12,45,60,61], but increased direct obstruction of diaspores by plants [37]. Therefore, deserts and desert steppes with low coverage are conducive for increasing diaspore DV, thereby facilitating secondary wind dispersal [20,25]. Compared to herbaceous structures, the grass–shrub combination vegetation structure had a favorable effect on diaspore DV (Figure 6), possibly because the surface under the vegetation of grass–shrub combinations was sparser, and the wind velocity was higher under the same coverage. Our results revealed a sharp decline in DV when vegetation coverage surpassed 40% (Figure 6). Consistent with the results of wind velocity profiles (Figure 3), the near-surface wind velocity also decreased drastically when vegetation coverage ≥ 40%, which limits the continual movement of diaspores. Previous studies have shown that diaspores with greater mass are more difficult to move compared to smaller diaspores [17,62]. However, in our study, diaspore mass contributed the least to their DV (Table 3), suggesting that the impact of mass on DV may vary due to morphological differences; for example, spherical diaspores with greater mass bear less force from rolling friction, thereby reducing the effect of mass on DV.
4.3. Diaspore Adaptation Strategies with Different Morphological Characteristics in Different Vegetation Structures in Secondary Wind Dispersal
The morphological characteristics of different diaspores represent adaptation strategies for wind dispersal in various vegetation structures. Dispersal strategies are classified into three types: long-distance wind dispersal, anti-long-distance wind dispersal, and non-long-distance wind dispersal [60,63,64]. Diaspores adopting the long-distance dispersal strategy generally exhibit morphological features that facilitate extended dispersal [65,66]. Diaspores with pappus demonstrate the long-distance dispersal strategy. They typically possess a large projected area, low mass, small wing loading, low density, and low terminal velocity (Table 1), as well as high DV and low lift-off velocity (LV) in different vegetation structures (Figure 3 and Figure 5). They can adapt to a variety of habitats, from active dunes to meadow grasslands. Diaspores showing anti-long-distance wind dispersal strategies are limited by their initial position or morphological structure, inhibiting long-distance dispersal [65]. Diaspores with one wing possess the anti-long-distance strategy. The diaspores often have characteristics such as a low projected area, heavy mass, high wing loading, density, and terminal velocity (Table 1), as well as low DV and high LV (Figure 4 and Figure 6). Although they possess wind dispersal traits, they tend to become trapped by surface structures or obstructed by vegetation during secondary dispersal. Diaspores with a non-long-distance wind dispersal strategy lack structures conducive to long-distance dispersal [65]. Diaspores with thorns, discoid, balloons, and four wings adapt to the non-long-distance wind dispersal strategy. They are easily saltated or rolled on the ground surface, and their secondary wind dispersal is influenced more by the vegetation structure. Thus, they are more suited to low-coverage deserts and desert steppes than to high-coverage typical steppes and meadows [49,67].
Our results showed that there is variation in the aerodynamic traits within each appendage type group, resulting from differences in species selection (Table 1). The variation between different species may vary the dispersal strategies of diaspores with the same appendage type. For example, diaspores with one wing adopt the anti-long-distance strategy, as a whole, whereas the Acer ginnala might tend toward the non-long-distance wind dispersal strategy because of its lower terminal velocity and wing loading than Fraxinus chinensis and Acer truncatum. In contrast, diaspores with four wings adopt the non-long-distance strategy, while Calligonum rubicundum tends to adopt the anti-long-distance strategy because its terminal velocity and wing loading are higher than those of Calligonum leucocladum.
5. Conclusions
Vegetation structure and diaspore appendage type are crucial factors that significantly influence lift-off and dispersal velocities in secondary wind dispersal. Vegetation coverage is the primary factor affecting near-surface wind velocity. Notably, 40% vegetation cover acted as a threshold, significantly affecting the secondary wind dispersal of diaspores across varying vegetation structures. Diaspores adopt diverse dispersal strategies according to their morphological and wind dynamic characteristics, such as terminal velocity, wing loading, and shape index, to affect lift-off and dispersal velocities. This study not only helps understand the relationship between the vegetation structure and the processes and strategies of diaspores’ secondary dispersal by wind, but also facilitates habitat restoration and biodiversity conservation of grassland and forest ecosystems.
Author Contributions
Conceptualization, Q.Z. and Z.L.; formal analysis, L.T. and L.Z.; methodology, Y.W. and X.L.; software, S.C.; supervision, J.Z.; visualization, H.Y. and Y.J.; writing—original draft, X.L.; writing—review and editing, Q.Z. and Z.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (grant number 31971732 and 41601588), the Strategic Priority Research Program of Chinese Academy of Sciences (grant number XDA26020104).
Data Availability Statement
The data that support the findings of this study are available upon request from the corresponding author.
Acknowledgments
We thank the Experimental Center of Desert Forestry, Chinese Academy of Forestry, and the First Forestry Station for providing accommodation during the experiment.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cousens, R.; Dytham, C.; Law, R. Dispersal in Plants: A Population Perspective; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Levin, S.A.; Muller-Landau, H.C.; Nathan, R.; Chave, J. The ecology and evolution of seed dispersal: A theoretical perspective. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 575–604. [Google Scholar] [CrossRef]
- Nathan, R.; Muller-Landau, H.C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 2000, 15, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Schurr, F.M.; Bond, W.J.; Midgley, G.F.; Higgins, S.I. A mechanistic model for secondary seed dispersal by wind and its experimental validation. J. Ecol. 2005, 93, 1017–1028. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, M.; Xin, Z.; Liu, Z.; Schurr, F.M. A trade-off between primary and secondary seed dispersal by wind. Plant Ecol. 2019, 220, 541–552. [Google Scholar] [CrossRef]
- Nathan, R.; Katul, G.G.; Bohrer, G.; Kuparinen, A.; Soons, M.B.; Thompson, S.E.; Trakhtenbrot, A.; Horn, H.S. Mechanistic models of seed dispersal by wind. Theor. Ecol. 2011, 4, 113–132. [Google Scholar] [CrossRef]
- Willson, M.F.; Rice, B.; Westoby, M. Seed dispersal spectra: A comparison of temperate plant communities. J. Veg. Sci. 1990, 1, 547–562. [Google Scholar] [CrossRef]
- Chambers, J.C.; MacMahon, J.A. A day in the life of a seed: Movements and fates of seeds and their implications for natural and managed systems. Annu. Rev. Ecol. Syst. 1994, 25, 263–292. [Google Scholar] [CrossRef]
- der Weduwen, D.; Ruxton, G.D. Secondary dispersal mechanisms of winged seeds: A review. Biol. Rev. 2019, 94, 1830–1838. [Google Scholar] [CrossRef] [PubMed]
- Reichman, O. Spatial and temporal variation of seed distributions in Sonoran Desert soils. J. Biogeogr. 1984, 11, 1–11. [Google Scholar] [CrossRef]
- Vander Wall, S.B.; Joyner, J.W. Secondary dispersal by the wind of winged pine seeds across the ground surface. Am. Midl. Nat. 1998, 139, 365–373. [Google Scholar] [CrossRef]
- Redbo-Torstensson, P.; Telenius, A. Primary and secondary seed dispersal by wind and water in Spergularia salina. Ecography 1995, 18, 230–237. [Google Scholar] [CrossRef]
- Greene, D.F.; Johnson, E.A. Secondary dispersal of tree seeds on snow. J. Ecol. 1997, 85, 329–340. [Google Scholar] [CrossRef]
- Matlack, G.R. Diaspore size, shape, and fall behavior in wind-dispersed plant-species. Am. J. Bot. 1987, 74, 1150–1160. [Google Scholar] [CrossRef]
- Kowarik, I.; Von der Lippe, M. Secondary wind dispersal enhances long-distance dispersal of an invasive species in urban road corridors. NeoBiota 2011, 9, 49. [Google Scholar] [CrossRef]
- von der Lippe, M.; Bullock, J.M.; Kowarik, I.; Knopp, T.; Wichmann, M. Human-mediated dispersal of seeds by the airflow of vehicles. PLoS ONE 2013, 8, e52733. [Google Scholar] [CrossRef]
- Johnson, E.A.; Fryer, G.I. Physical characterization of seed microsites—Movement on the ground. J. Ecol. 1992, 80, 823–836. [Google Scholar] [CrossRef]
- Wang, J.-h.; Ma, Q.-l.; Liu, H.-j.; Yang, Z.-h.; Zhang, D.-k. Effect of wind-breaking and sand-fixing of vegetation in progressive succession on desertification land in arid area. J. Desert Res. 2006, 26, 908–909. [Google Scholar]
- Maurer, K.D.; Hardiman, B.S.; Vogel, C.S.; Bohrer, G. Canopy-structure effects on surface roughness parameters: Observations in a Great Lakes mixed-deciduous forest. Agric. For. Meteorol. 2013, 177, 24–34. [Google Scholar] [CrossRef]
- Monty, A.; Brown, C.S.; Johnston, D.B. Fire promotes downy brome (Bromus tectorum L.) seed dispersal. Biol. Invasions 2013, 15, 1113–1123. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, Y.; Li, X.; Liu, Z.; Wu, J.; Musa, A.; Ma, Q.; Yu, H.; Cui, X.; Wang, L. Geographical distribution and determining factors of different invasive ranks of alien species across China. Sci. Total Environ. 2020, 722, 137929. [Google Scholar] [CrossRef] [PubMed]
- Jongejans, E.; Telenius, A. Field experiments on seed dispersal by wind in ten umbelliferous species (Apiaceae). Plant Ecol. 2001, 152, 67–78. [Google Scholar] [CrossRef]
- Nathan, R. The challenges of studying dispersal. Trends Ecol. Evol. 2001, 16, 481–483. [Google Scholar] [CrossRef]
- Zong, L.; Liang, W.; Liu, Z.; Liu, M.; Baskin, C.C.; Tian, L.; Xin, Z.; Zhou, Q.; Ba, C. Diaspore burial during wind dispersal depends on particle size of the underlying substrate. Plant Soil 2023, 482, 163–174. [Google Scholar] [CrossRef]
- Zhou, Q.; Liu, Z.; Xin, Z.; Daryanto, S.; Wang, L.; Li, X.; Wang, Y.; Liang, W.; Qin, X.; Zhao, Y.; et al. Responses of secondary wind dispersal to environmental characteristics and diaspore morphology of seven Calligonum species. Land Degrad. Dev. 2019, 31, 842–850. [Google Scholar] [CrossRef]
- Zhou, Q.; Liu, Z.; Xin, Z.; Qian, J.; Wang, Y.; Liang, W.; Qin, X.; Zhao, Y.; Li, X.; Cui, X.; et al. How do wind speed, release height, seed morphology interact to determine seed dispersal trajectory of Calligonum (Polygonaceae) species. bioRxiv 2018, 362434. [Google Scholar] [CrossRef]
- Qin, X.; Liu, Z.; Liu, M.; Liang, W.; Baskin, C.C.; Baskin, J.M.; Xin, Z.; Li, X.; Wang, Z.; Zhou, Q. Shrub canopy interception of diaspores dispersed by wind. Seed Sci. Res. 2020, 30, 310–318. [Google Scholar] [CrossRef]
- Liu, Z.M.; Shou, W.K.; Qian, J.Q.; Wu, J.; Busso, C.A.; Hou, X.Z. Level of habitat fragmentation determines its non-linear relationships with plant species richness, frequency and density at desertified grasslands in Inner Mongolia, China. J. Plant Ecol. 2018, 11, 866–876. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Z.; Qian, J. Non-linear effect of habitat fragmentation on plant diversity: Evidence from a sand dune field in a desertified grassland in northeastern China. Ecol. Eng. 2013, 54, 90–96. [Google Scholar] [CrossRef]
- Soons, M.; Messelink, J.; Jongejans, E.; Heil, G. Habitat fragmentation reduces grassland connectivity for both short-distance and long-distance wind-dispersed forbs. J. Ecol. 2005, 93, 1214–1225. [Google Scholar] [CrossRef]
- Szitár, K.; Tölgyesi, C.; Deák, B.; Gallé, R.; Korányi, D.; Batáry, P. Connectivity and fragment size drive plant dispersal and persistence traits in forest steppe fragments. Front. Ecol. Evol. 2023, 11, 1155885. [Google Scholar] [CrossRef]
- Aavik, T.; Helm, A. Restoration of plant species and genetic diversity depends on landscape-scale dispersal. Restor. Ecol. 2018, 26, S92–S102. [Google Scholar] [CrossRef]
- Escribano-Avila, G.; Calviño-Cancela, M.; Pías, B.; Virgos, E.; Valladares, F.; Escudero, A. Diverse guilds provide complementary dispersal services in a woodland expansion process after land abandonment. J. Appl. Ecol. 2014, 51, 1701–1711. [Google Scholar] [CrossRef]
- Liu, M.; Xin, Z.; Su, Z.; Zhao, Y.; Li, X.; Liu, Z.; Cony, M.A.; Liang, W.; Qin, X.; Qian, J. A video camera recording method for measuring terminal velocity of seed dispersal by wind. J. For. Res. 2021, 32, 81–90. [Google Scholar] [CrossRef]
- Thompson, K.; Band, S.R.; Hodgson, J.G. Seed size and shape predict persistence in soil. Funct. Ecol. 1993, 7, 236–241. [Google Scholar] [CrossRef]
- Matlack, G.R. Influence of fruit size and weight on wind dispersal in betula-lenta, a gap-colonizing tree species. Am. Midl. Nat. 1992, 128, 30–39. [Google Scholar] [CrossRef]
- Tian, L.; Liang, W.; Liu, Z.; Liu, M.; Phartyal, S.S.; Zong, L.; Xin, Z.; Ba, C.; Li, X.; Liu, Y. The complexity of vegetation structure decreases the seed-hanging ability of the vegetation layer during secondary wind dispersal. Trees 2023, 37, 1671–1680. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, M.; Xin, Z.; Zhao, Y.; Liu, Z. Which factors have stronger explanatory power for primary wind dispersal distance of winged diaspores: The case of Zygophyllum xanthoxylon (Zygophyllaceae)? J. Plant Ecol. 2016, 9, 346–356. [Google Scholar] [CrossRef]
- Nielsen, T.; Khodabakhsh, A.; Busch, C. Unit-Selection Based Facial Video Manipulation Detection. In Proceedings of the 2020 International Conference of the Biometrics Special Interest Group (BIOSIG), Darmstadt, Germany, 16–18 September 2020; pp. 1–5. [Google Scholar]
- Skarpaas, O.; Auhl, R.; Shea, K. Environmental variability and the initiation of dispersal: Turbulence strongly increases seed release. Proc. R. Soc. B Biol. Sci. 2006, 273, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Musick, H.B.; Trujillo, S.M.; Truman, C.R. Wind-tunnel modelling of the influence of vegetation structure on saltation threshold. Earth Surf. Process. Landf. 1996, 21, 589–605. [Google Scholar] [CrossRef]
- Yang, L.; Huai, W.; Guo, Y. Stochastic simulation of the suspended sediment deposition in the channel with vegetation and its relevance to turbulent kinetic energy. Water Resour. Res. 2021, 57, e2021WR030380. [Google Scholar] [CrossRef]
- Wolfe, S.A.; Nickling, W. Shear stress partitioning in sparsely vegetated desert canopies. Earth Surf. Process. Landf. 1996, 21, 607–619. [Google Scholar] [CrossRef]
- Hensen, I.; Müller, C. Experimental and structural investigations of anemochorous dispersal. Plant Ecol. 1997, 133, 169–180. [Google Scholar] [CrossRef]
- Pounden, E.; Greene, D.; Quesada, M.; Contreras Sánchez, J. The effect of collisions with vegetation elements on the dispersal of winged and plumed seeds. J. Ecol. 2008, 96, 591–598. [Google Scholar] [CrossRef]
- Zong, L.; Liang, W.; Liu, Z.; Liu, M.; Baskin, C.C.; Baskin, J.M.; Tian, L.; Li, X.; Xin, Z.; Luo, W. Lift-off velocity of diaspores during secondary wind dispersal varies with particle size of the underlying surface matrix. Plant Soil 2023, 482, 529–542. [Google Scholar] [CrossRef]
- Shan-shan, S.; Yong, S.; Shun-ying, J. Experimental study of rolling-sliding transition and friction coefficients of particles. Rock Soil Mech. 2009, 30, 110–115. [Google Scholar]
- Baskin, J.M.; Lu, J.J.; Baskin, C.C.; Tan, D.Y.; Wang, L. Diaspore dispersal ability and degree of dormancy in heteromorphic species of cold deserts of northwest China: A review. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 93–99. [Google Scholar] [CrossRef]
- Maddox, J.C.; Carlquist, S. Wind dispersal in californian desert plants: Experimental studies and conceptual considerations. Aliso 1985, 11, 77–96. [Google Scholar]
- Straatsma, M.; Warmink, J.J.; Middelkoop, H. Two novel methods for field measurements of hydrodynamic density of floodplain vegetation using terrestrial laser scanning and digital parallel photography. Int. J. Remote Sens. 2008, 29, 1595–1617. [Google Scholar] [CrossRef]
- Cheng, T.l.; Pan, Y.x.; Wang, X.p.; Li, Y. Community plant height modulated by aridity promotes spatial vegetation patterns in Alxa plateau in Northwest China. Ecol. Evol. 2023, 13, e9823. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.K.; Almlöf, K. Adapting boreal streams to climate change: Effects of riparian vegetation on water temperature and biological assemblages. Freshw. Sci. 2016, 35, 984–997. [Google Scholar] [CrossRef]
- Liang, H.; Wang, J.; Lu, Q.; Yang, W.; Dong, H.; Zhao, A.; Zhang, Z. Windbreak effects and wind velocity flow field of low density arbor with two spatial distribution patterns in wind tunnel. J. Desert Res. 2009, 29, 1021–1028. [Google Scholar]
- Abdurahman, M.; Sabirhazi, G.; Liu, B.; Yin, L.; Pan, B. Comparison of five Calligonum species in tarim basin based on morphological and molecular data. Excli J. 2012, 11, 776–782. [Google Scholar] [PubMed]
- Trakhtenbrot, A.; Katul, G.G.; Nathan, R. Mechanistic modeling of seed dispersal by wind over hilly terrain. Ecol. Model. 2014, 274, 29–40. [Google Scholar] [CrossRef]
- Jongejans, E.; Pedatella, N.M.; Shea, K.; Skarpaas, O.; Auhl, R. Seed release by invasive thistles: The impact of plant and environmental factors. Proc. R. Soc. B Biol. Sci. 2007, 274, 2457–2464. [Google Scholar] [CrossRef] [PubMed]
- Schenk, J.J. Evolution of limited seed dispersal ability on gypsum islands. Am. J. Bot. 2013, 100, 1811–1822. [Google Scholar] [CrossRef] [PubMed]
- Greene, D.F.; Quesada, M. The differential effect of updrafts, downdrafts and horizontal winds on the seed abscission of Tragopogon dubius. Funct. Ecol. 2011, 25, 468–472. [Google Scholar] [CrossRef]
- Thompson, S.E.; Katul, G.G. Implications of nonrandom seed abscission and global stilling for migration of wind-dispersed plant species. Glob. Change Biol. 2013, 19, 1720–1735. [Google Scholar] [CrossRef]
- Heydel, F.; Cunze, S.; Bernhardt-Roemermann, M.; Tackenberg, O. Long-distance seed dispersal by wind: Disentangling the effects of species traits, vegetation types, vertical turbulence and wind speed. Ecol. Res. 2014, 29, 641–651. [Google Scholar] [CrossRef]
- Schenkeveld, A.J. The Ecology of Short-Lived Forbs in Chalk Grasslands. Ph.D. Thesis, University of Utrecht, Utrecht, The Netherlands, 1984. [Google Scholar]
- Zhou, Q.; Liu, Z.; Xin, Z.; Daryanto, S.; Wang, L.; Qian, J.; Wang, Y.; Liang, W.; Qin, X.; Zhao, Y.; et al. Relationship between seed morphological traits and wind dispersal trajectory. Funct. Plant Biol. FPB 2019, 46, 1063–1071. [Google Scholar] [CrossRef]
- Venable, D.L.; Flores-Martinez, A.; Muller-Landau, H.C.; Barron-Gafford, G.; Becerra, J.X. Seed dispersal of desert annuals. Ecology 2008, 89, 2218–2227. [Google Scholar] [CrossRef] [PubMed]
- Quick, Z.I.; Houseman, G.R.; Bueyuektahtakin, I.E. Assessing wind and mammals as seed dispersal vectors in an invasive legume. Weed Res. 2017, 57, 35–43. [Google Scholar] [CrossRef]
- Zhu, J.; Buchmann, C.M.; Schurr, F.M. Effects of seed morphology and orientation on secondary seed dispersal by wind. J. Plant Ecol. 2022, 15, 1257–1272. [Google Scholar] [CrossRef]
- van Oudtshoorn, K.v.R.; van Rooyen, M.W. Dispersal Biology of Desert Plants; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Bonte, D.; Hovestadt, T.; Poethke, H.J. Evolution of dispersal polymorphism and local adaptation of dispersal distance in spatially structured landscapes. Oikos 2010, 119, 560–566. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).