
Effects of Canopy Nitrogen Addition and Understory Vegetation Removal on Nitrogen Transformations in a Subtropical Forest
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design and Measurements
2.3. Nitrogen Mineralization Measurement
2.4. Statistical Analyses
3. Results
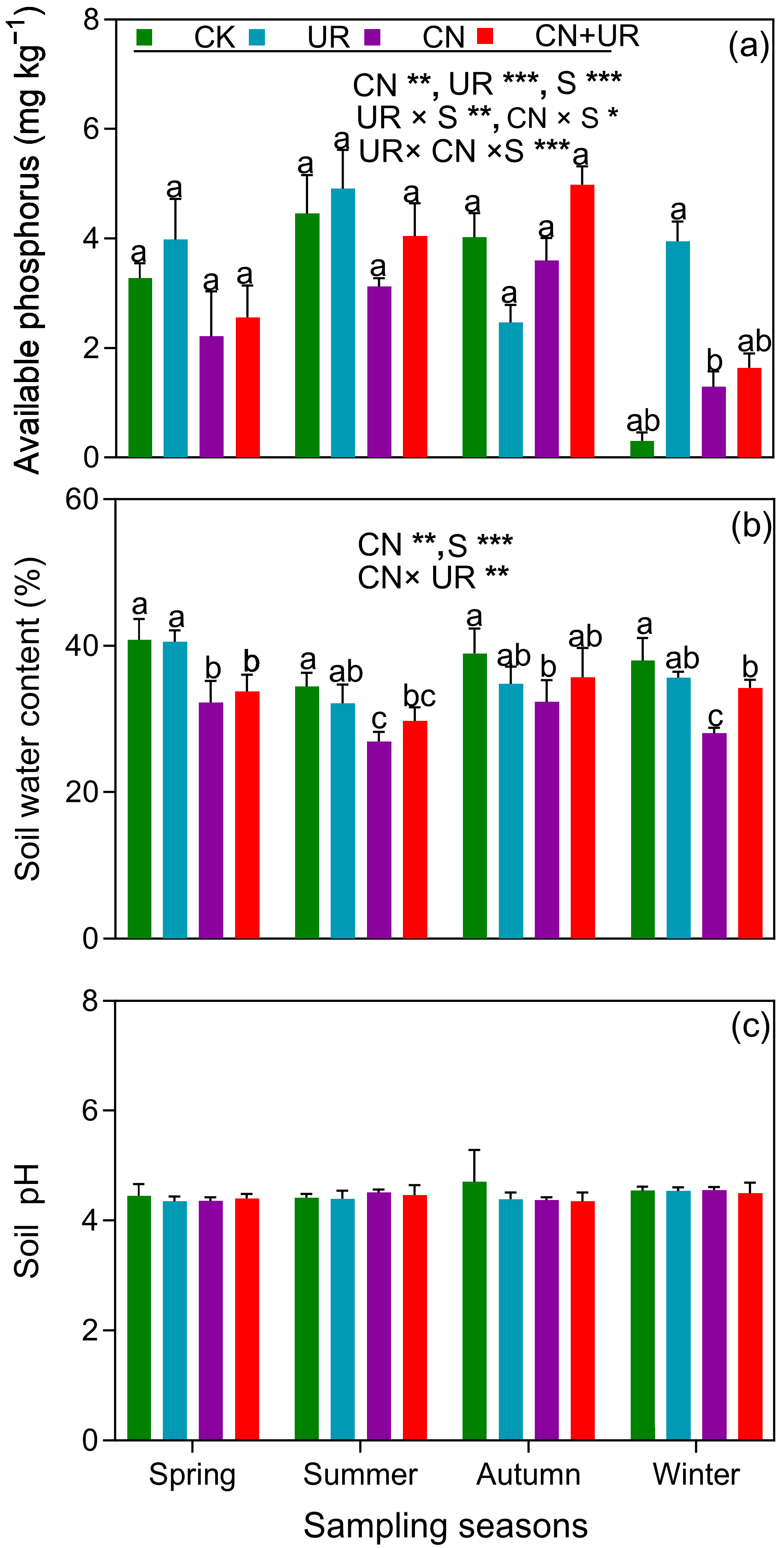
3.1. Soil Physiochemical Properties
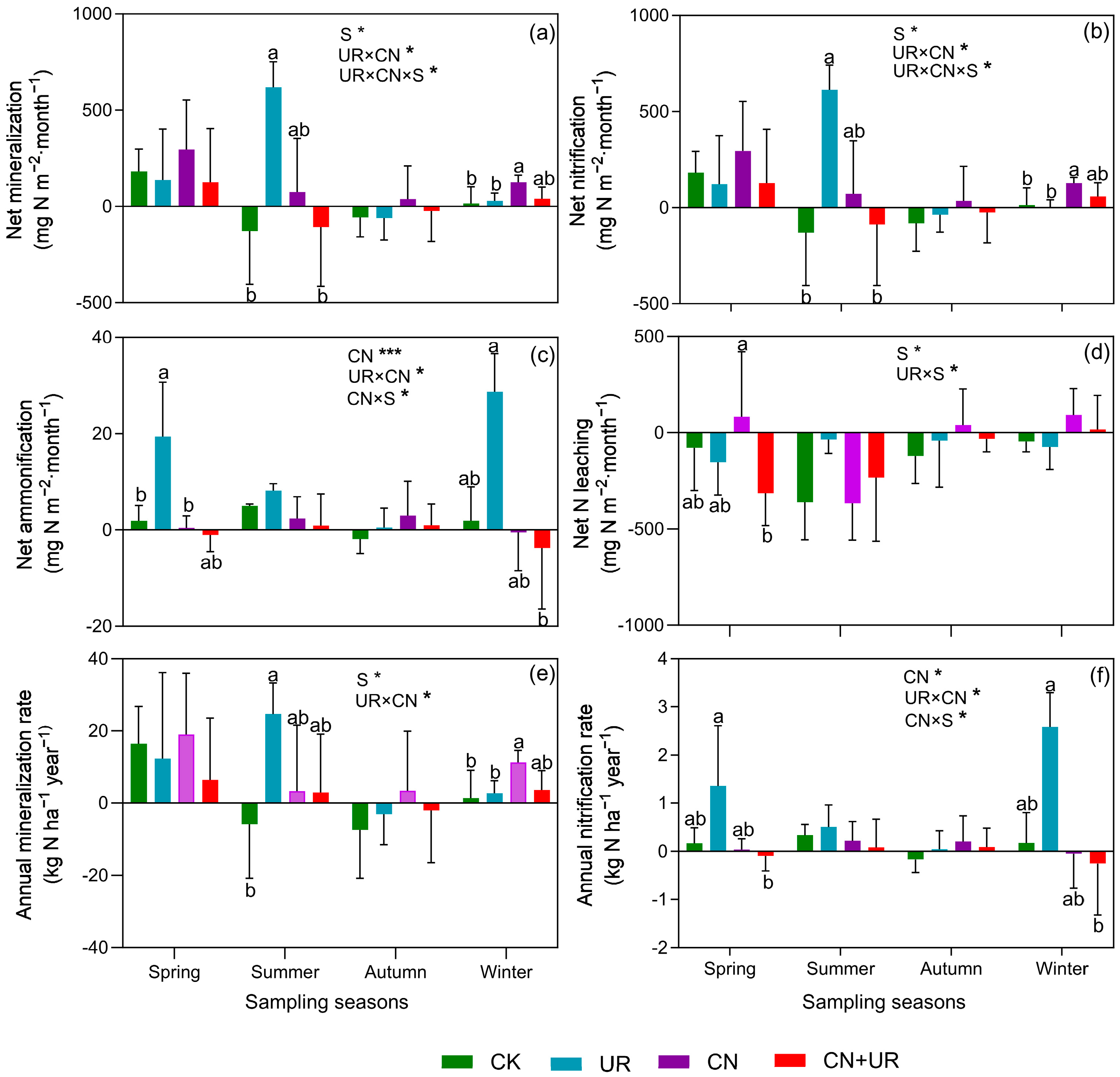
3.2. N Transformation under CN and UR
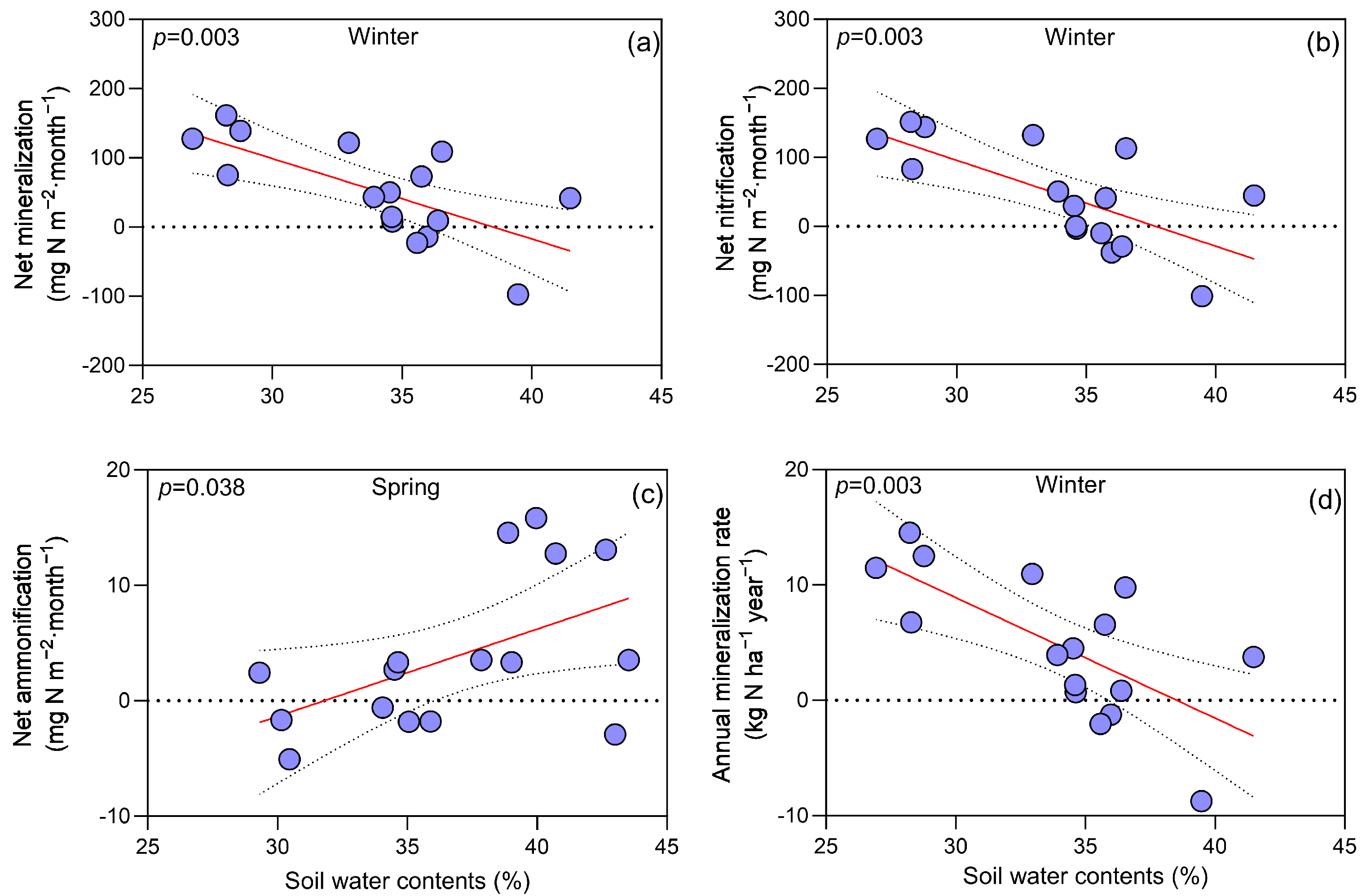
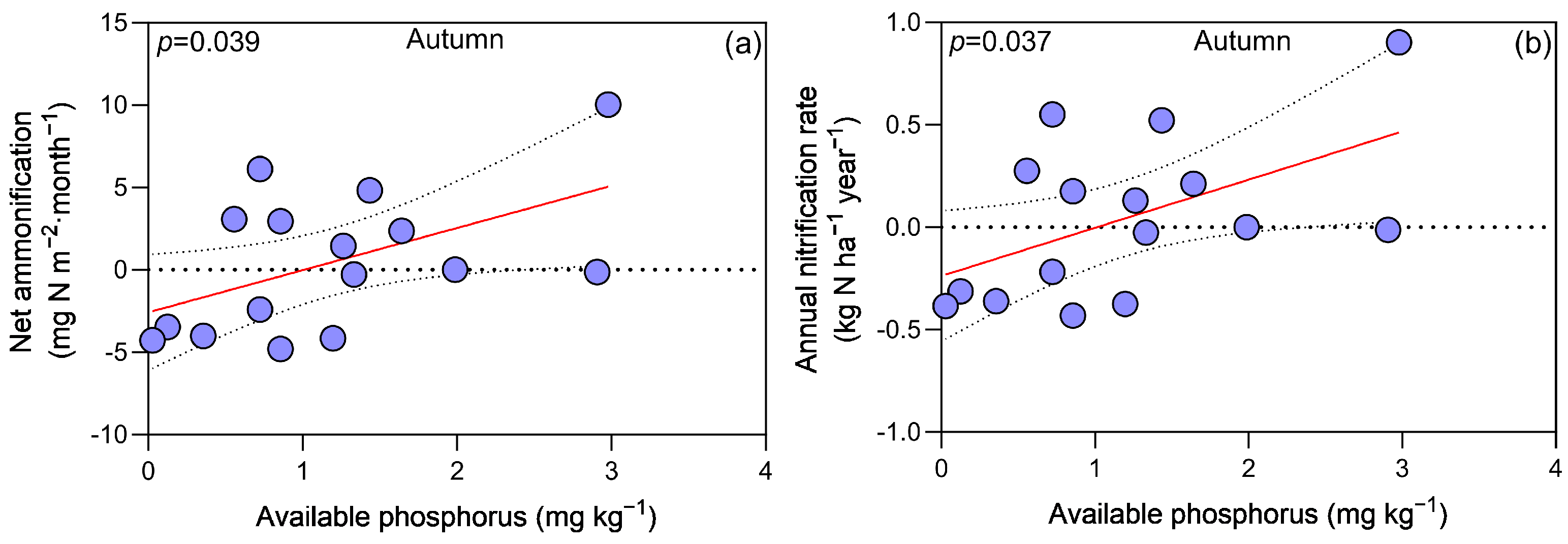
3.3. Regression Analyses between Soil Factors and N Processes Dynamics
4. Discussion
4.1. Seasonal Dynamics and the Impact of CN on N Processes
4.2. Understory Vegetation Removal, Soil Nitrogen Processes, and Seasonal Correlations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gudale, M.J. The impact of anthropogenic nitrogen deposition on global forests: Negative impacts far exceed the carbon benefits. Glob. Chang. Biol. 2022, 28, 690–692. [Google Scholar] [CrossRef]
- Nesha, K.; Herold, M.; Sy, D.; Bruin, S.D.; Araza, A.; Málaga, N.; Gamarra, J.G.P.; Hergoualc’h, K.; Pekkarinen, A.; Ramirez, C.; et al. Exploring characteristics of national forest inventories for integration with global space-based forest biomass data. Sci. Total Environ. 2022, 850, 157788. [Google Scholar] [CrossRef]
- Stevens, C.J. Nitrogen in the environment. Science 2019, 363, 578–580. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Y.; Wang, M.; Wang, K.; Meng, F.; Liu, L.; Zhao, Y.; Ma, L.; Zhu, Q.; Xu, W.; et al. Atmospheric nitrogen deposition: A review of quantification methods and its spatial pattern derived from the global monitoring networks. Ecotoxicol. Environ. Saf. 2021, 216, 112180. [Google Scholar] [CrossRef]
- Lu, X.; Ren, W.; Hou, E.; Tang, S.; Zhang, L.; Liu, Z.; Lin, Y.; Fu, S.; Wen, D.; Kuang, Y. Different effects of canopy and understory nitrogen addition on soil organic carbon and its related processes in a subtropical forest. J. Soils Sediments 2021, 21, 235–244. [Google Scholar] [CrossRef]
- Verma, P.; Sagar, R. Effect of nitrogen (N) deposition on soil-N processes: A holistic approach. Sci. Rep. 2020, 10, 10470. [Google Scholar] [CrossRef]
- Chen, C.; Chen, X.; Chen, H.Y.H. Mapping N deposition impacts on soil microbial biomass across global terrestrial ecosystems. Geoderma 2023, 433, 116429. [Google Scholar] [CrossRef]
- Yu, Q.; Duan, L.; Yu, L.; Chen, X.; Si, G.; Ke, P.; Ye, Z.; Mulder, J. Threshold and multiple indicators for nitrogen saturation in subtropical forests. Environ. Pollut. 2018, 241, 664–673. [Google Scholar] [CrossRef]
- Harris, E.; Yu, L.; Wang, Y.P.; Mohn, J.; Henne, S.; Bai, E.; Barthel, M.; Bauters, M.; Boeckx, P.; Dorich, C.; et al. Warming and redistribution of nitrogen inputs drive an increase in terrestrial nitrous oxide emission factor. Nat. Commun. 2022, 13, 4310. [Google Scholar] [CrossRef]
- Liu, M.; Gan, B.; Li, Q.; Xiao, W.; Song, X. Effects of nitrogen and phosphorus addition on soil extracellular enzyme activity and stoichiometry in Chinese fir (Cunninghamia lanceolata) forests. Front. Plant Sci. 2022, 13, 834184. [Google Scholar] [CrossRef]
- Chen, H.; Gurmesa, G.A.; Zhang, W.; Zhu, X.; Zheng, M.; Mao, Q.; Zhang, T.; Mo, J. Nitrogen saturation in humid tropical forests after 6 years of nitrogen and phosphorus addition: Hypothesis testing. Funct. Ecol. 2016, 30, 305–313. [Google Scholar] [CrossRef]
- Hu, P.; Zhao, Y.; Xiao, D.; Xu, Z.; Zhang, W.; Xiao, J.; Wang, K. Dynamics of soil nitrogen availability following vegetation restoration along a climatic gradient of a subtropical karst region in China. J. Soils Sediments 2021, 21, 2167–2178. [Google Scholar] [CrossRef]
- Houlton, B.Z.; Driscoll, C.T.; Fahey, T.J.; Likens, G.E.; Groffman, P.M.; Bernhardt, E.S.; Buso, D.C. Nitrogen dynamics in ice storm-damaged forest ecosystems: Implications for nitrogen limitation theory. Ecosystems 2003, 6, 431–443. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bilbrough, C.; Welker, J.M. Increased snow depth affects microbial activity and nitrogen mineralization in two Arctic tundra communities. Soil Biol. Biochem. 2004, 36, 217–227. [Google Scholar] [CrossRef]
- Hu, S.; Chapin, F.S.; Firestone, M.K.; Field, C.B.; Chiariello, N.R. Nitrogen limitation of microbial decomposition in a grassland under elevated CO2. Nature 2001, 409, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Lin, W.; Gao, R.; Yin, Y.; Peng, Y. Nitrogen addition change soil N pools with litter removal or not in subtropical forest. J. Soil Sci. Plant Nutr. 2020, 66, 421–428. [Google Scholar] [CrossRef]
- Karki, H.; Bargali, K.; Bargali, S.S. Spatial and temporal trends in soil N-mineralization rates under the agroforestry systems in Bhabhar belt of Kumaun Himalaya, India. Agrofor. Syst. 2021, 95, 1603–1617. [Google Scholar] [CrossRef]
- Zhou, L.; Li, S.; Jia, Y.; Heal, K.V.; He, Z.; Wu, P.; Ma, X. Spatiotemporal distribution of canopy litter and nutrient resorption in a chronosequence of different development stages of Cunninghamia lanceolata in southeast China. Sci. Total Environ. 2021, 762, 143153. [Google Scholar] [CrossRef]
- Piñeiro, J.; Ochoa, H.R.; Serrano, G.L.; Power, S.A. Phosphorus and water supply independently control productivity and soil enzyme activity responses to elevated CO2 in an understorey community from a Eucalyptus woodland. Plant Soil 2022, 483, 643–657. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, J.; Lambers, H.; Li, Y.; Li, Y.; Qin, G.; Wang, M.; Wang, J.; Li, Z.; Wang, F. Nitrogen and phosphorus addition exerted different influences on litter and soil carbon release in a tropical forest. Sci. Total Environ. 2022, 832, 155049. [Google Scholar] [CrossRef]
- Baral, B.R.; Kuyper, T.W.; Van Groenigen, J.W. Liebig’s law of the minimum applied to a greenhouse gas: Alleviation of P-limitation reduces soil N2O emission. Plant Soil 2014, 374, 539–548. [Google Scholar] [CrossRef]
- Turner, B.L.; Joseph, W.S. The response of microbial biomass and hydrolytic enzymes to a decade of nitrogen, phosphorus, and potassium addition in a lowland tropical rain forest. Biogeochemistry 2014, 117, 115–130. [Google Scholar] [CrossRef]
- Fang, X.M.; Wang, G.G.; Xu, Z.J.; Zong, Y.Y.; Zhang, X.L.; Li, J.J.; Wang, H.; Chen, F.S. Litter addition and understory removal influenced soil organic carbon quality and mineral nitrogen supply in a subtropical plantation forest. Plant Soil 2021, 460, 527–540. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, X.; Li, D.; Li, S.; Chen, Z.; Wu, J. A meta-analysis of understory plant removal impacts on soil properties in forest ecosystems. Geoderma 2022, 426, 116116. [Google Scholar] [CrossRef]
- Chen, W.; Su, F.; Pang, Z.; Mao, Q.; Zhong, B.; Xiong, Y.; Mo, J.; Lu, X. The removal of understory vegetation can rapidly alter the soil microbial community structure without altering the community assembly in a primary tropical forest. Geoderma 2023, 429, 116180. [Google Scholar] [CrossRef]
- Xu, X.; Yin, L.; Duan, C.; Jing, Y. Effect of N addition, moisture, and temperature on soil microbial respiration and microbial biomass in forest soil at different stages of litter decomposition. J. Soils Sediments 2016, 16, 1421–1439. [Google Scholar] [CrossRef]
- Shah, J.A.; Liu, W.; Ullah, S.; Duan, H.; Shen, F.; Liao, Y.; Huang, G.; Wu, J. Linkages among leaf nutrient concentration, resorption efficiency, litter decomposition and their stoichiometry to canopy nitrogen addition and understory removal in subtropical plantation. Ecol. Process 2024, 13, 27. [Google Scholar] [CrossRef]
- Gao, W.; Yang, H.; Kou, L.; Li, S. Effects of nitrogen deposition and fertilization on N transformations in forest soils: A review. J. Soils Sediments 2015, 15, 863–879. [Google Scholar] [CrossRef]
- Nair, R.K.F.; Perks, M.P.; Weatherall, A.; Baggs, E.M.; Mencuccini, M. Does canopy nitrogen uptake enhance carbon sequestration by trees? Glob. Chang. Biol. 2016, 22, 875–888. [Google Scholar] [CrossRef]
- Tian, Y.; Wang, J.; Zhou, L.; Tao, L.; Lin, Y.; Hui, D.; Ren, H.; Lu, H. Nitrogen budgets of a lower subtropical forest as affected by 6 years of over-canopy and understory nitrogen additions. Sci. Total Environ. 2022, 852, 158546. [Google Scholar] [CrossRef]
- Sievering, H.; Tomaszewski, T.; Torizzo, J. Canopy uptake of atmospheric N deposition at a conifer forest: Part I-canopy N budget, photosynthetic efficiency and net ecosystem exchange. Tellus Ser. B Chem. Phys. Meteorol. 2007, 59, 483–492. [Google Scholar] [CrossRef]
- Schwarz, M.T.; Bischoff, S.; Blaser, S.; Boch, S.; Schmitt, B.; Thieme, L.; Fischer, M.; Michalzik, B.; Schulze, E.D.; Siemens, J.; et al. More efficient aboveground nitrogen use in more diverse central European forest canopies. For. Ecol. Manag. 2014, 313, 274–282. [Google Scholar] [CrossRef]
- Liu, T.; Mao, P.; Shi, L.L.; Eisenhauer, N.; Liu, S.J.; Wang, X.L.; He, X.X.; Wang, Z.Y.; Zhang, W.; Liu, Z.F.; et al. Forest canopy maintains the soil community composition under elevated nitrogen deposition. Soil Biol. Biochem. 2020, 143, 107733. [Google Scholar] [CrossRef]
- Wei, X.; Blanco, J.A.; Jiang, H.; Kimmins, J.P. Effects of nitrogen deposition on carbon sequestration in Chinese fir forest ecosystems. Sci. Total Environ. 2012, 416, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Raison, R.; Connell, M.; Khanna, P. Methodology for studying fluxes of soil mineral-N in situ. Soil Biol. Biochem. 1987, 19, 521–530. [Google Scholar] [CrossRef]
- Subler, S.; Parmelee, R.W.; Allen, M.F. Comparison of buried bag and PVC core methods for in situ measurement of nitrogen mineralization rates in an agricultural soil. Commun. Soil Sci. Plant Anal. 1995, 26, 2369–2381. [Google Scholar] [CrossRef]
- Wang, S.; Chen, X.; Li, D.; Wu, J. Effects of soil organism interactions and temperature on carbon use efficiency in three different forest soils. Soil Ecol. Lett. 2021, 3, 156–166. [Google Scholar] [CrossRef]
- Wang, R.; Bicharanloo, B.; Hou, E.; Jiang, Y.; Dijkstra, F.A. Phosphorus supply Increases nitrogen transformation rates and retention in soil: A global meta-analysis. Earths Future 2022, 10, e2021EF002479. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Reichmann, L.G.; Sala, O.E.; Peters, D.P.C. Water controls on nitrogen transformations and stocks in an arid ecosystem. Ecosphere 2013, 4, art11. [Google Scholar] [CrossRef]
- Contosta, A.R.; Frey, S.D.; Cooper, A.B. Seasonal dynamics of soil respiration and N mineralization in chronically warmed and fertilized soils. Ecosphere 2011, 2, art36. [Google Scholar] [CrossRef]
- Homyak, P.M.; Allison, S.D.; Huxman, T.E.; Goulden, M.L.; Treseder, K.K. Effects of drought manipulation on soil nitrogen cycling: A meta-analysis. J. Geophys. Res. Biogeosci 2017, 122, 3260–3272. [Google Scholar] [CrossRef]
- Lei, X.; Shen, F.; Lei, X.; Liu, W.; Duan, H.; Fan, H.; Wu, J. Assessing influence of simulated canopy nitrogen deposition and understory removal on soil microbial community structure in a Cunninghamia lanceolata plantation. Biodivers. Sci. 2018, 26, 962–971. [Google Scholar] [CrossRef]
- Mori, T.; Zhou, K.; Wang, S.; Zhang, W.; Mo, J. Effect of nitrogen addition on DOC leaching and chemical exchanges on canopy leaves in Guangdong Province. China. J. For. Res. 2019, 30, 1707–1713. [Google Scholar] [CrossRef]
- Wu, G.; Chen, D.; Zhou, Z.; Ye, Q.; Wu, J. Canopy nitrogen addition enhance the photosynthetic rate of canopy species by improving leaf hydraulic conductivity in a subtropical forest. Front. Plant Sci. 2022, 13, 942851. [Google Scholar] [CrossRef]
- Wallace, A.J.; Armstrong, R.D.; Grace, P.R.; Scheer, C. Nitrogen use efficiency and N2O emissions vary according to seasonal water supply across different cereal production systems of south eastern Australia. Geoderma Reg. 2022, 29, e00498. [Google Scholar] [CrossRef]
- Peralta, A.L.; Ludmer, S.; Matthews, J.W.; Kent, A.D. Bacterial community response to changes in soil redox potential along a moisture gradient in restored wetlands. Ecol. Eng. 2014, 73, 246–253. [Google Scholar] [CrossRef]
- Davidson, E.A.; Nepstad, D.C.; Ishida, F.Y.; Brando, P.M. Effects of an experimental drought and recovery on soil emissions of carbon dioxide, methane, nitrous oxide, and nitric oxide in a moist tropical forest. Glob. Chang. Biol. 2008, 14, 2582–2590. [Google Scholar] [CrossRef]
- Sardar, M.F.; Chen, Z.; Tang, C.; Zhang, S.; Fang, L.; Miao, D.; Li, Y.; Zhang, Q.; Li, Y. Seasonal linkages between soil nitrogen mineralization and the microbial community in broadleaf forests with Moso bamboo (Phyllostachys edulis) invasion. Sci. Total Environ. 2023, 899, 165557. [Google Scholar] [CrossRef]
- Shibata, M.; Sangsompaisarn, N.; Watanabe, S.; Funakawa, S. Gross nitrogen transformation rates in shifting cultivation systems in northern Thailand: Controlling factors and implications for inorganic nitrogen availability. Geoderma Reg. 2023, 34, e00692. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, W.; Gurmesa, G.A.; Zhu, X.; Li, D.; Mo, J. Phosphorus addition affects soil nitrogen dynamics in a nitrogen-saturated and two nitrogen-limited forests. Eur. J. Soil Sci. 2017, 68, 472–479. [Google Scholar] [CrossRef]
- Lin, C.; Lin, W.; Chen, S.; Peng, J.; Guo, J.; Yang, Y. Phosphorus addition accelerates fine root decomposition by stimulating extracellular enzyme activity in a subtropical natural evergreen broad-leaved forest. Eur. J. For. Res. 2019, 138, 917–928. [Google Scholar] [CrossRef]
- Mori, T.; Lu, X.; Aoyagi, R.; Mo, J. Reconsidering the phosphorus limitation of soil microbial activity in tropical forests. Funct. Ecol. 2018, 32, 1145–1154. [Google Scholar] [CrossRef]
- Qiu, L.; Zhang, Y.; Mao, R.; Chen, F.; Liu, J.; Yang, G.; Wan, S. Understory removal accelerates nucleic phosphorus release but retards residual phosphorus release in decomposing litter of Phyllostachys edulis in subtropical China. Land. Degrad. Dev. 2021, 32, 2695–2703. [Google Scholar] [CrossRef]
- Wang, Q.W.; Pieristè, M.; Liu, C.; Kenta, T.; Robson, T.M.; Kurokawa, H. The contribution of photodegradation to litter decomposition in a temperate forest gap and understorey. New Phytol. 2021, 229, 2625–2636. [Google Scholar] [CrossRef] [PubMed]
- Hogan, J.A.; Sharpe, J.M.; Ashley, V.B.; Stankavich, S.; Carmona, S.M.; Bithorn, J.E.; Díaz, J.T.; González, G.; Zimmerman, J.K.; Shiels, A.B. Solar radiation and soil moisture drive tropical forest understory responses to experimental and natural hurricanes. Ecosphere 2022, 13, e4150. [Google Scholar] [CrossRef]
- Wang, F.; Zou, B.; Li, H.; Li, H. The effect of understory removal on microclimate and soil properties in two subtropical lumber plantations. J. For. Res. 2014, 19, 238–243. [Google Scholar] [CrossRef]
- Xiong, L.; Yuan, C.; Wu, Q.; Fornara, D.A.; Heděnec, P.; Chen, S.; Peng, Y.; Zhao, Z.; Wu, F.; Yue, K. Understory vegetation removal significantly affected soil biogeochemical properties in forest ecosystems. Appl. Soil Ecol. 2024, 193, 105132. [Google Scholar] [CrossRef]
- Zhu, H.; Gong, L.; Ding, Z.; Li, Y. Effects of litter and root manipulations on soil carbon and nitrogen in a Schrenk’s spruce (Picea schrenkiana) forest. PLoS ONE 2021, 16, e0247725. [Google Scholar] [CrossRef]
- Takahashi, K.S.; Uemura, S.; Suzuki, J.I.; Hara, T. Effects of understory dwarf bamboo removal on soil water and the growth of overstory trees in a dense secondary Betula ermanii forest, northern Japan. Ecol. Res. 2003, 18, 767–774. [Google Scholar] [CrossRef]
- Giuggiola, A.; Zweifel, R.; Feichtinger, L.M.; Vollenweider, P.; Bugmann, H.; Haeni, M.; Rigling, A. Competition for water in a xeric forest ecosystem—Effects of understory removal on soil micro-climate, growth and physiology of dominant Scots pine trees. For. Ecol. Manag. 2018, 409, 241–249. [Google Scholar] [CrossRef]
- Turner, B.L.; Yavitt, J.B.; Harms, K.E.; Garcia, M.N.; Wright, J. Seasonal changes in soil organic matter after a decade of nutrient addition in a lowland tropical forest. Biogeochemistry 2015, 123, 221–235. [Google Scholar] [CrossRef]
- Chen, X.; Fang, K.; Chen, C. Available phosphorus in soils of Taihu Lake. Water Environ. J. 2012, 26, 392–398. [Google Scholar] [CrossRef]
- Gao, X.; Gao, Y.; Li, X.; Zhang, C.; Zeng, Q.; Yuan, X.; Chen, Y.; Yu, Y.; Fu, S. Eleven-year canopy nitrogen addition enhances the uptake of phosphorus by plants and accelerates its depletion in soil. Forests 2024, 15, 416. [Google Scholar] [CrossRef]
- Song, Y.; Xing, J.; Hu, C.; Song, C.; Wang, Q.; Wang, S. Decomposition and carbon and nitrogen releases of twig and leaf litter were inhibited by increased level of nitrogen deposition in a subtropical evergreen broad-leaved forest in Southwest China. Forests 2024, 15, 492. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, S.; Liu, W.; Shah, J.A.; Shen, F.; Liao, Y.; Duan, H.; Huang, G.; Wu, J. Effects of Canopy Nitrogen Addition and Understory Vegetation Removal on Nitrogen Transformations in a Subtropical Forest. Forests 2024, 15, 962. https://doi.org/10.3390/f15060962
Ullah S, Liu W, Shah JA, Shen F, Liao Y, Duan H, Huang G, Wu J. Effects of Canopy Nitrogen Addition and Understory Vegetation Removal on Nitrogen Transformations in a Subtropical Forest. Forests. 2024; 15(6):962. https://doi.org/10.3390/f15060962
Chicago/Turabian StyleUllah, Saif, Wenfei Liu, Jawad Ali Shah, Fangfang Shen, Yingchun Liao, Honglang Duan, Guomin Huang, and Jianping Wu. 2024. "Effects of Canopy Nitrogen Addition and Understory Vegetation Removal on Nitrogen Transformations in a Subtropical Forest" Forests 15, no. 6: 962. https://doi.org/10.3390/f15060962