
Prediction of Potential Suitable Distribution of Liriodendron chinense (Hemsl.) Sarg. in China Based on Future Climate Change Using the Optimized MaxEnt Model



Abstract
1. Introduction
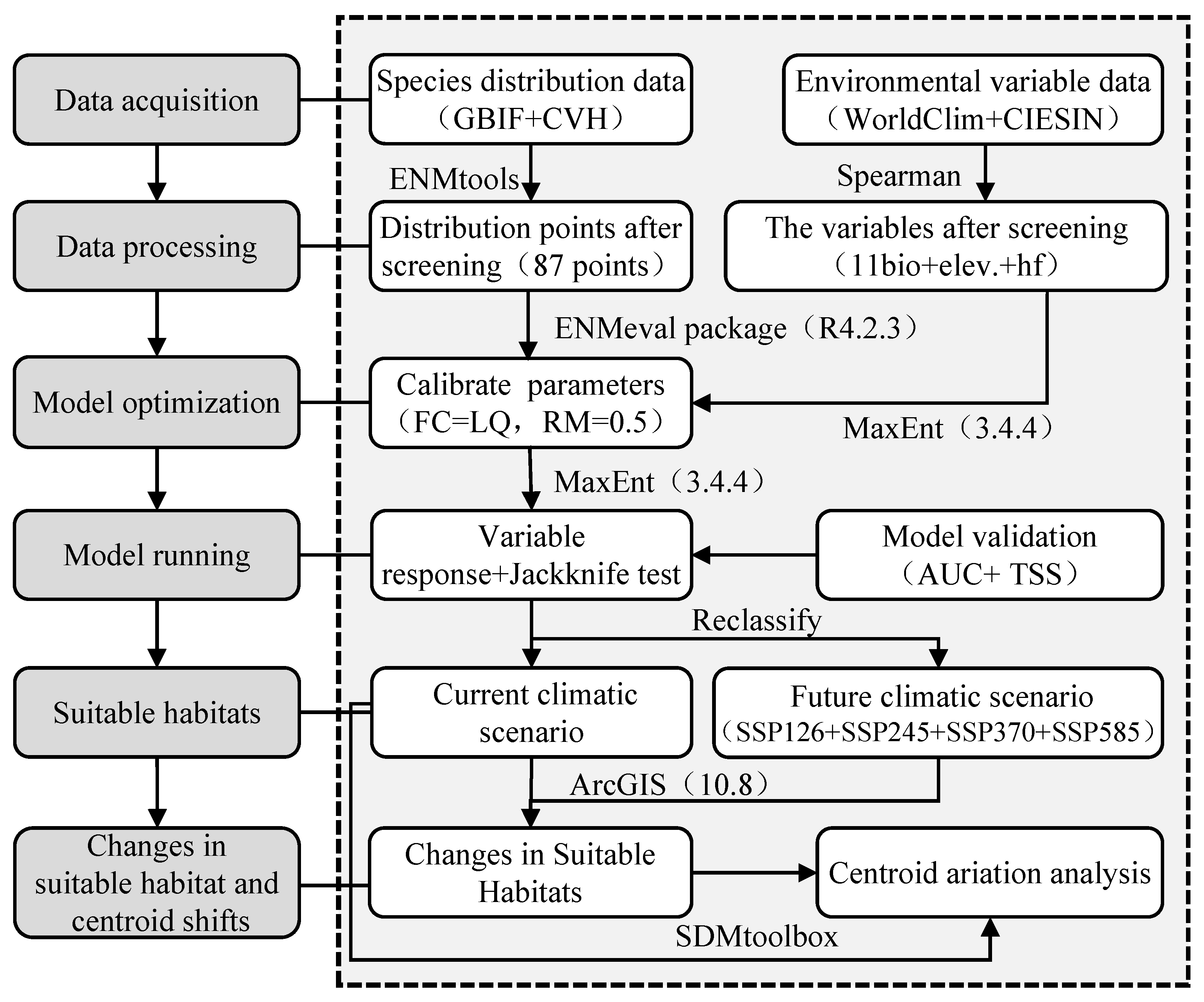
2. Materials and Methods
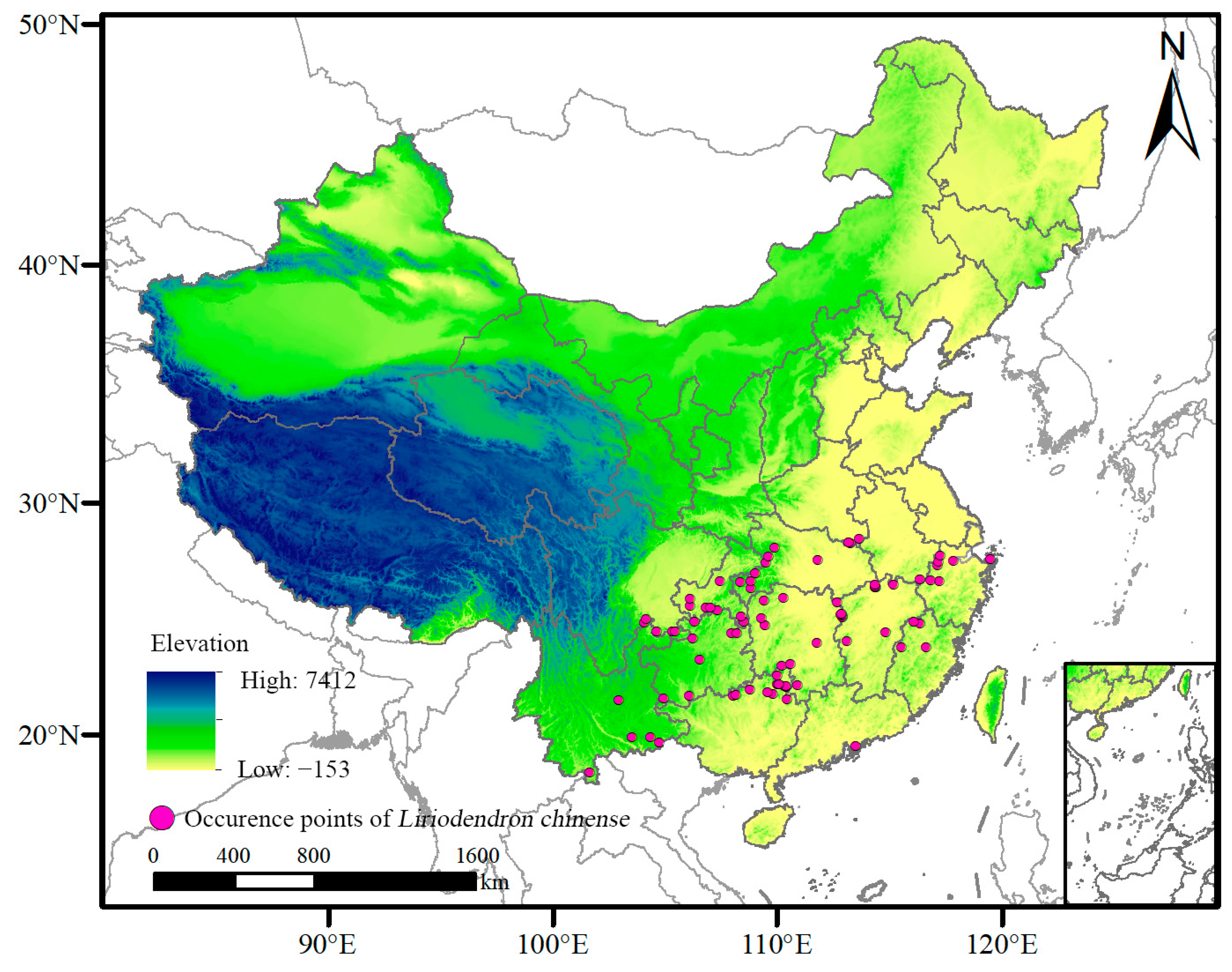
2.1. Sources and Processing of Species Distribution Data
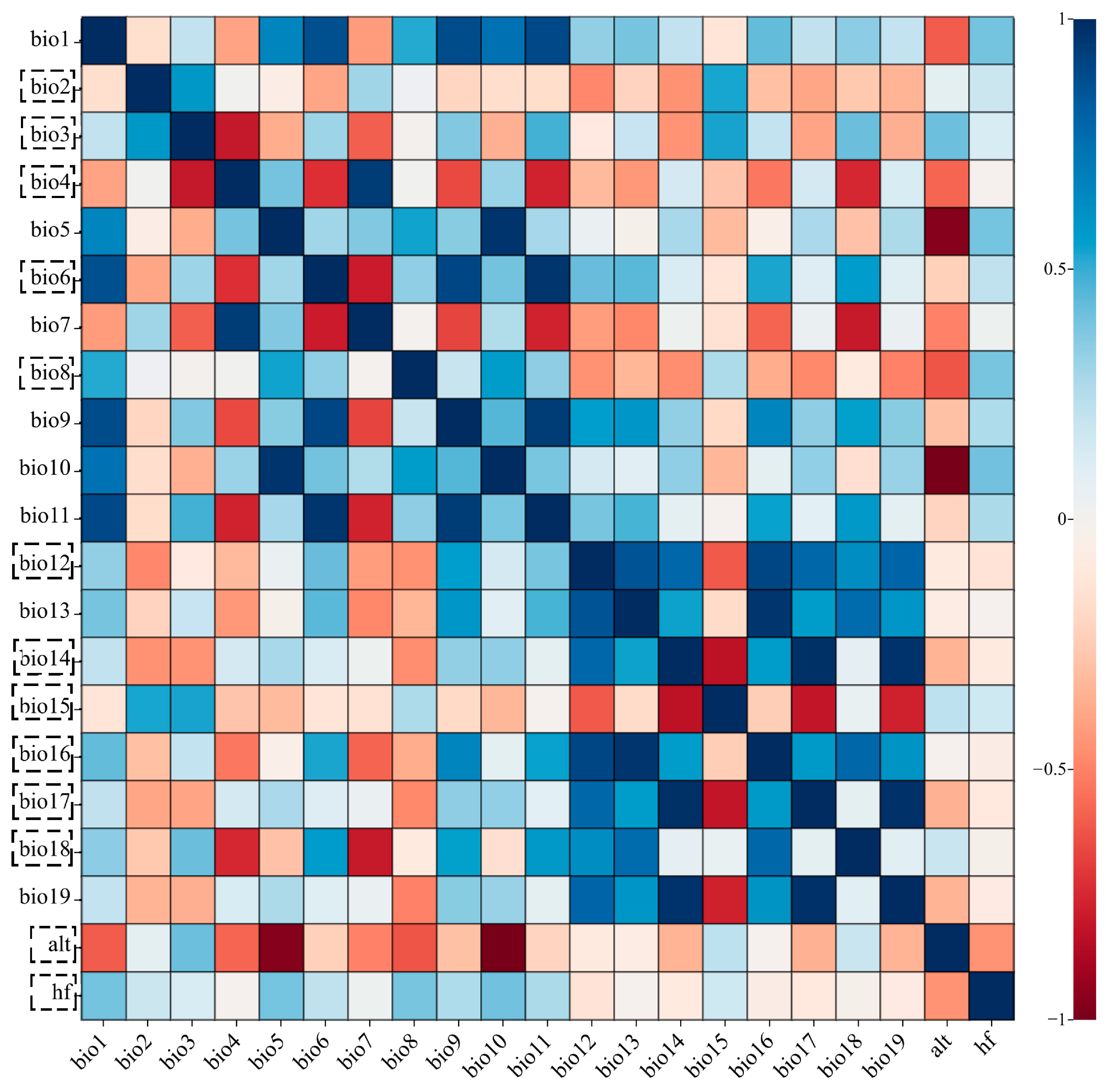
2.2. Collection and Processing of Environmental Variable Data
2.3. Model Optimization and Accuracy Evaluation
2.4. Suitability Classification and Analysis of Dominant Environmental Variables
2.5. Centroid Analysis of Suitable Areas
3. Results
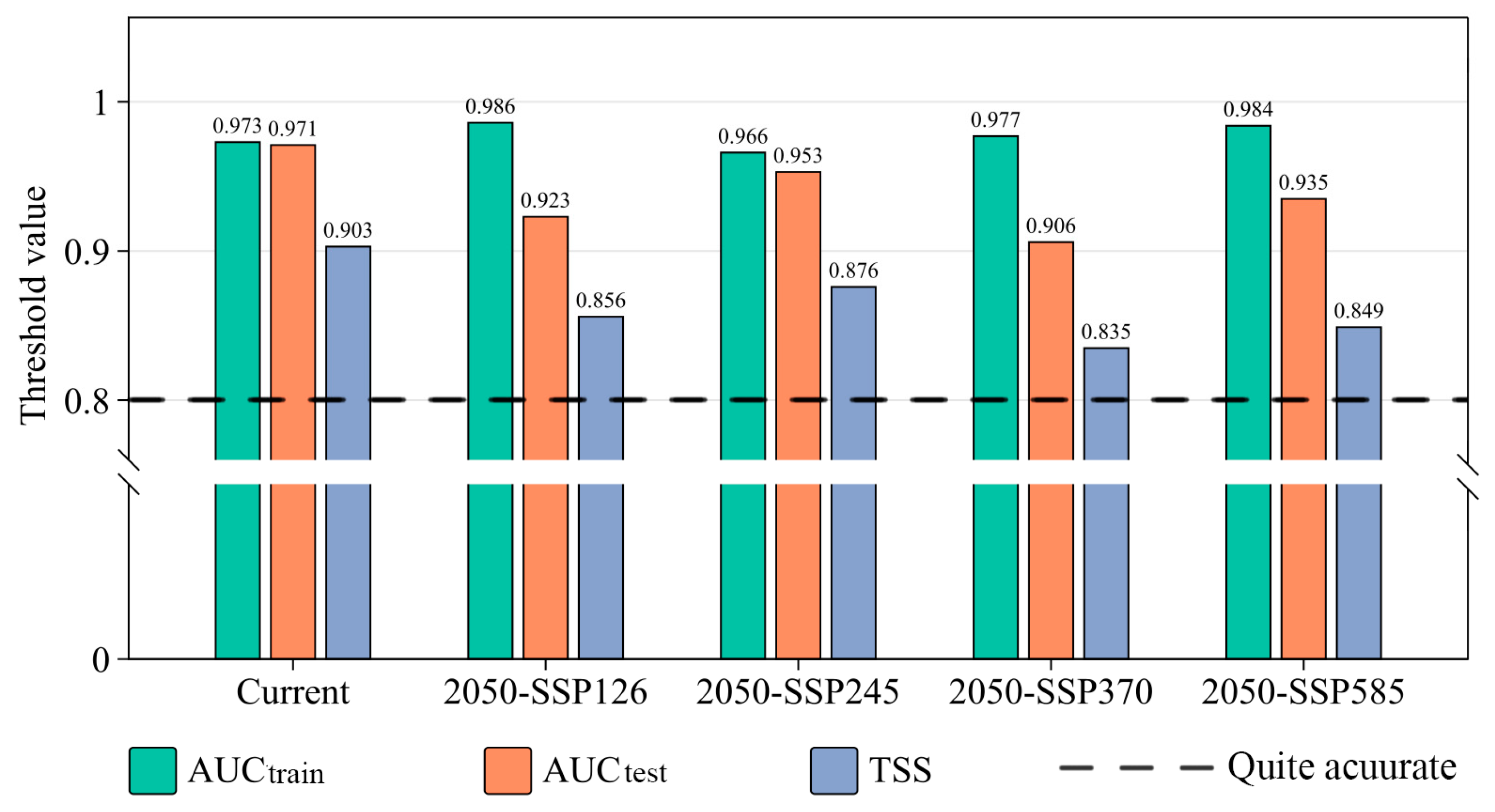
3.1. Optimization and Accuracy Evaluation of the MaxEnt Model
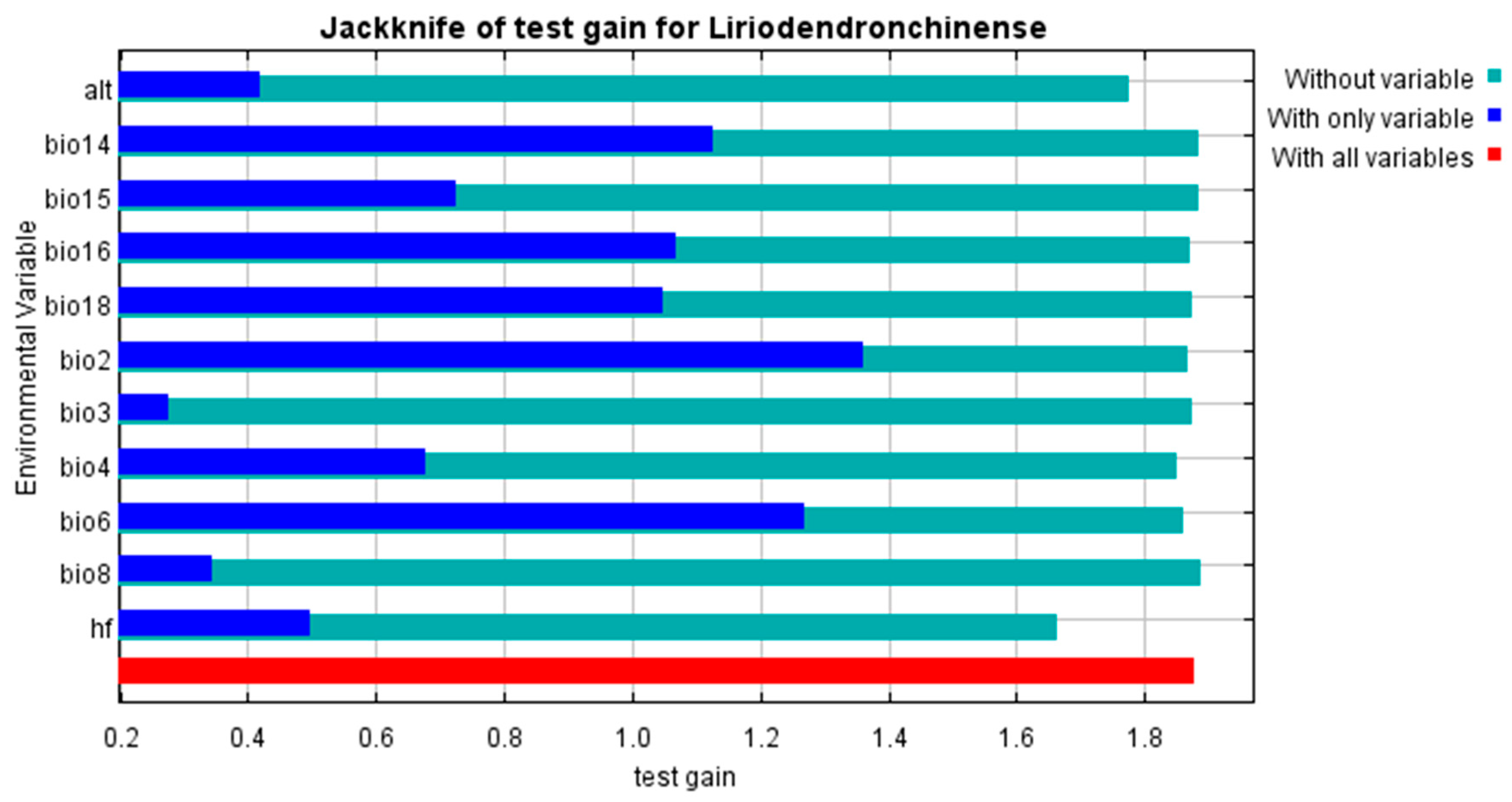
3.2. Dominant Environmental Variables Influencing the Distribution of L. chinense
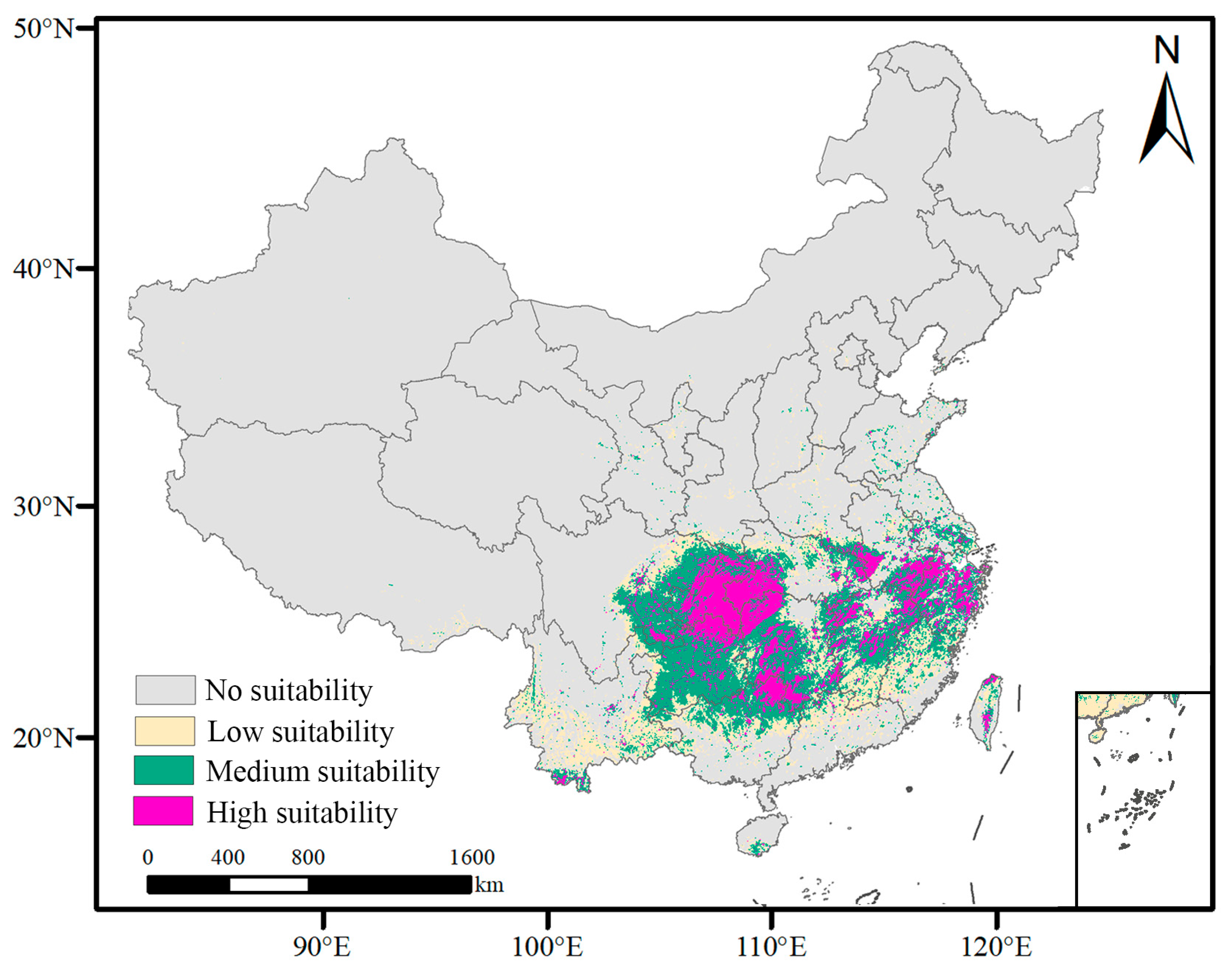
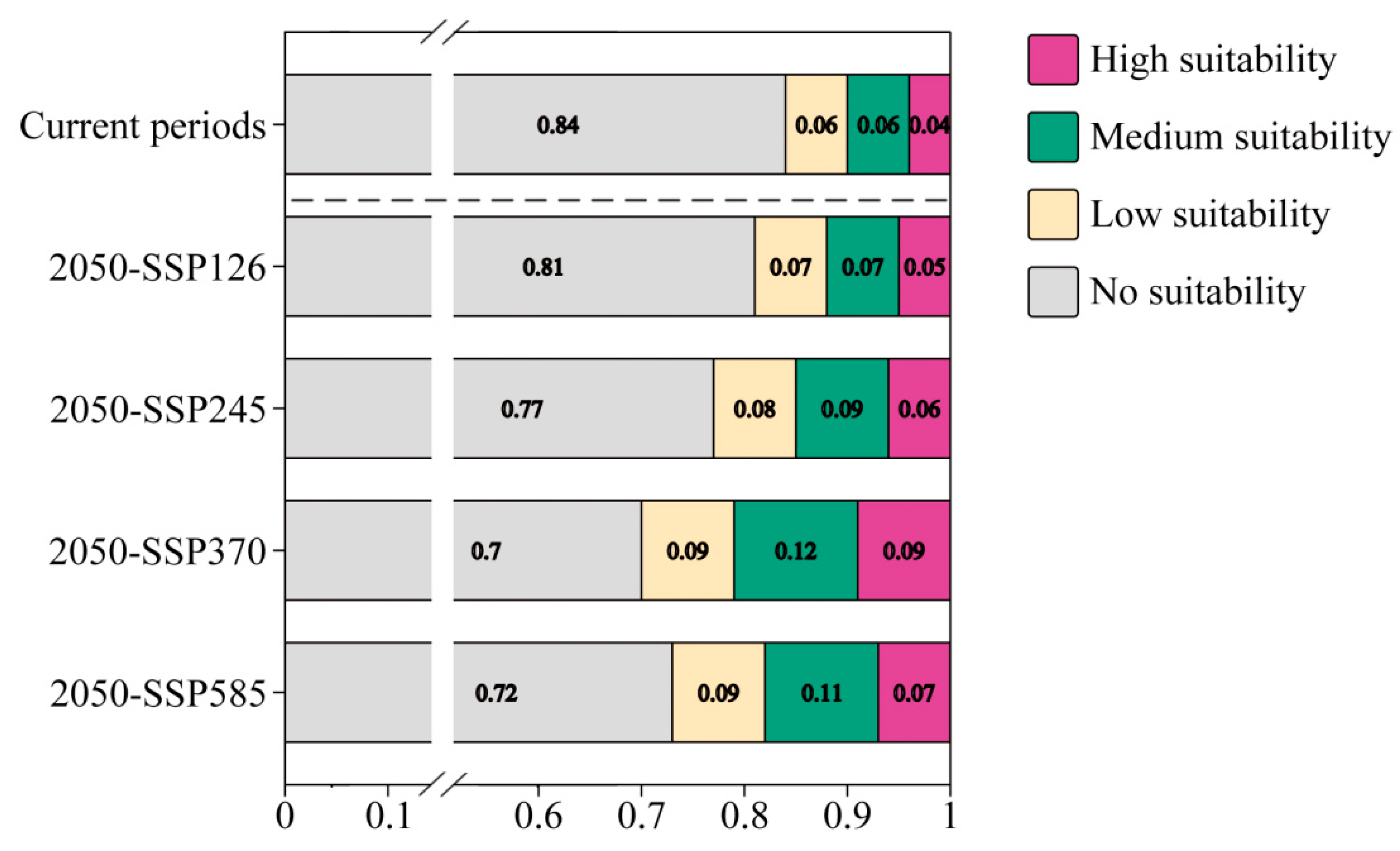
3.3. Potential Suitable Distribution Areas of L. chinense under Current Climate Conditions
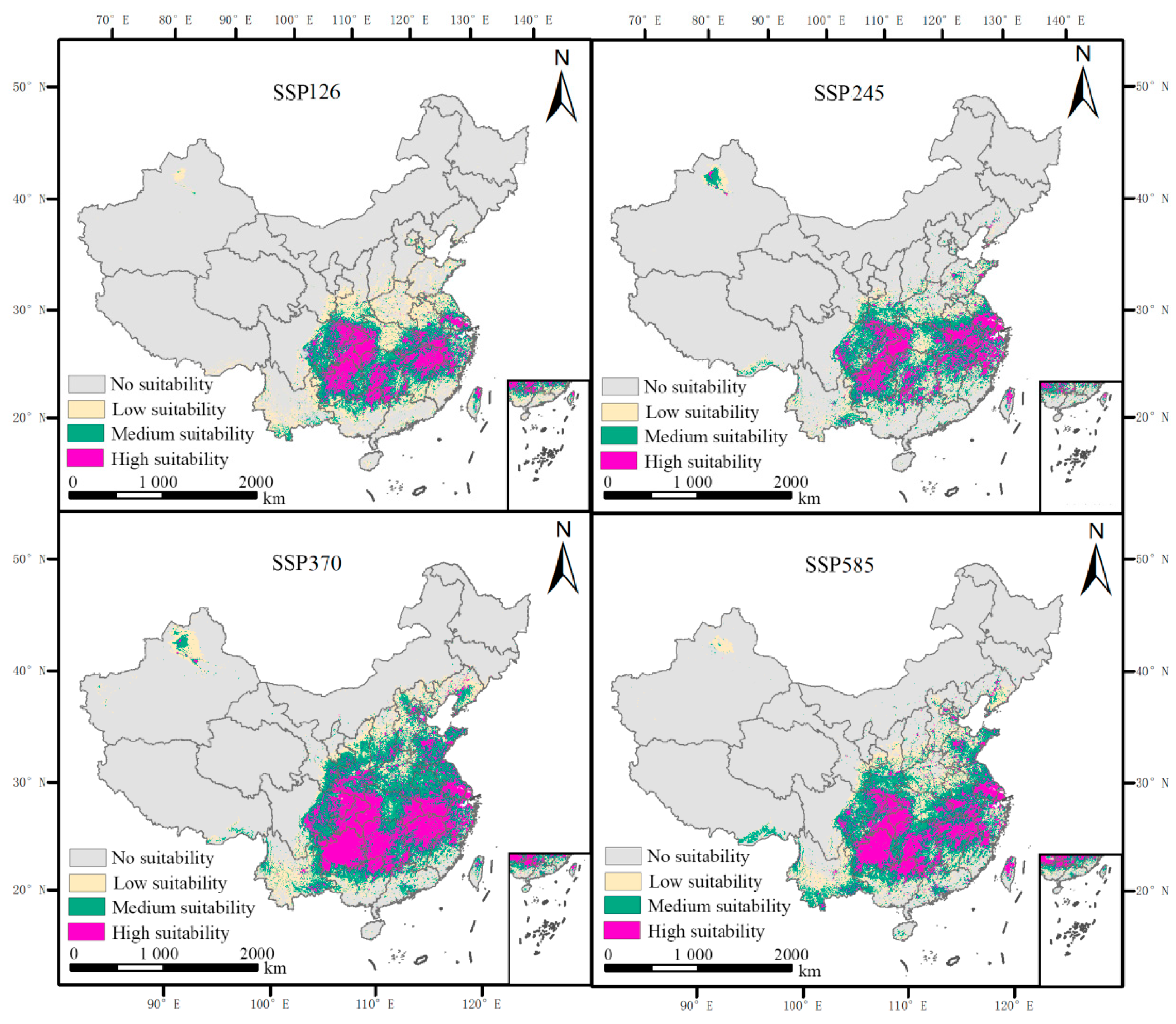
3.4. Forecasting the Impact of Future Climate Change on the Potential Geographic Distribution of L. chinense
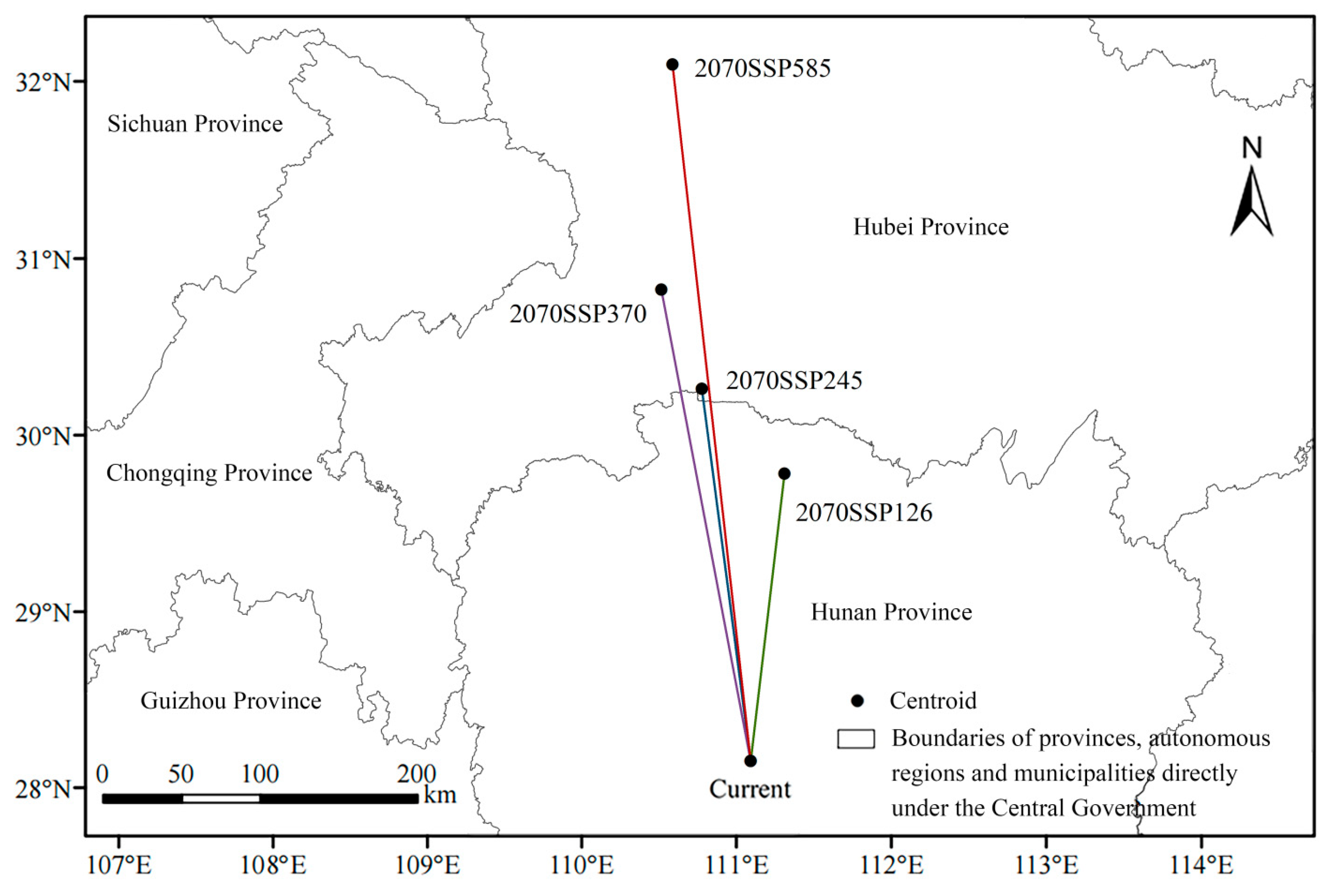
3.5. Shift in the Centroid of L. chinense’s Suitable Distribution Area
4. Discussion
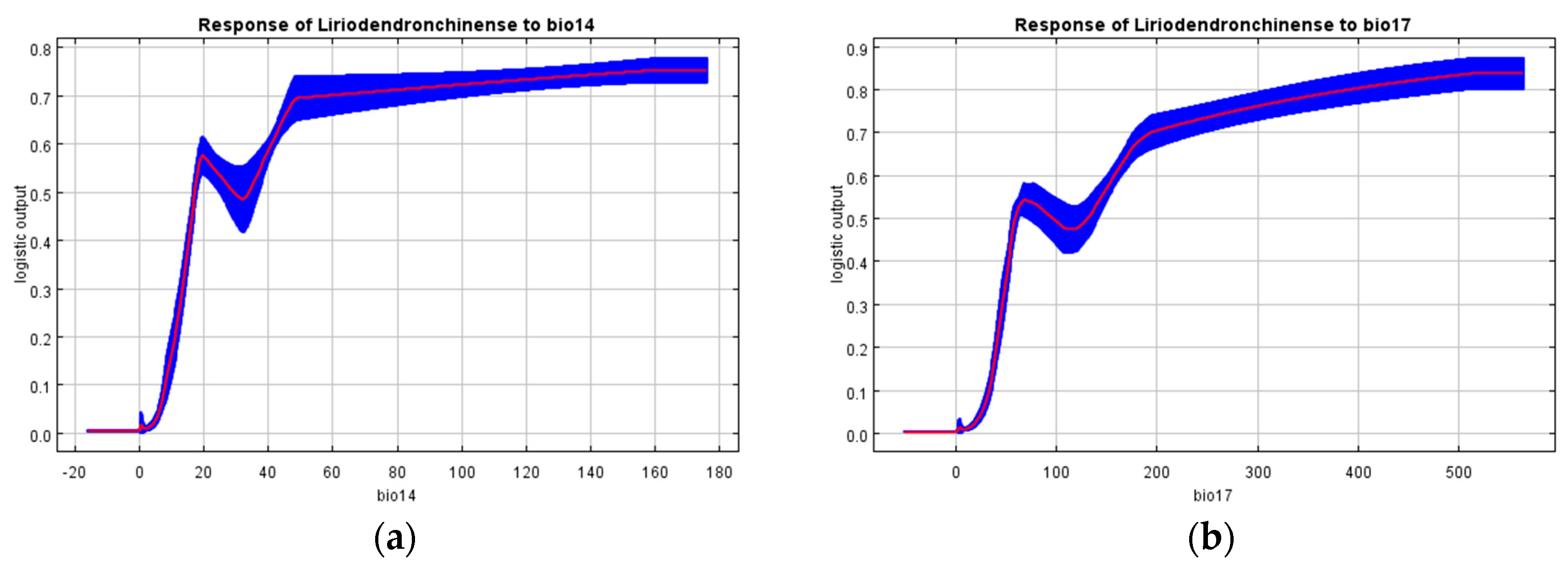
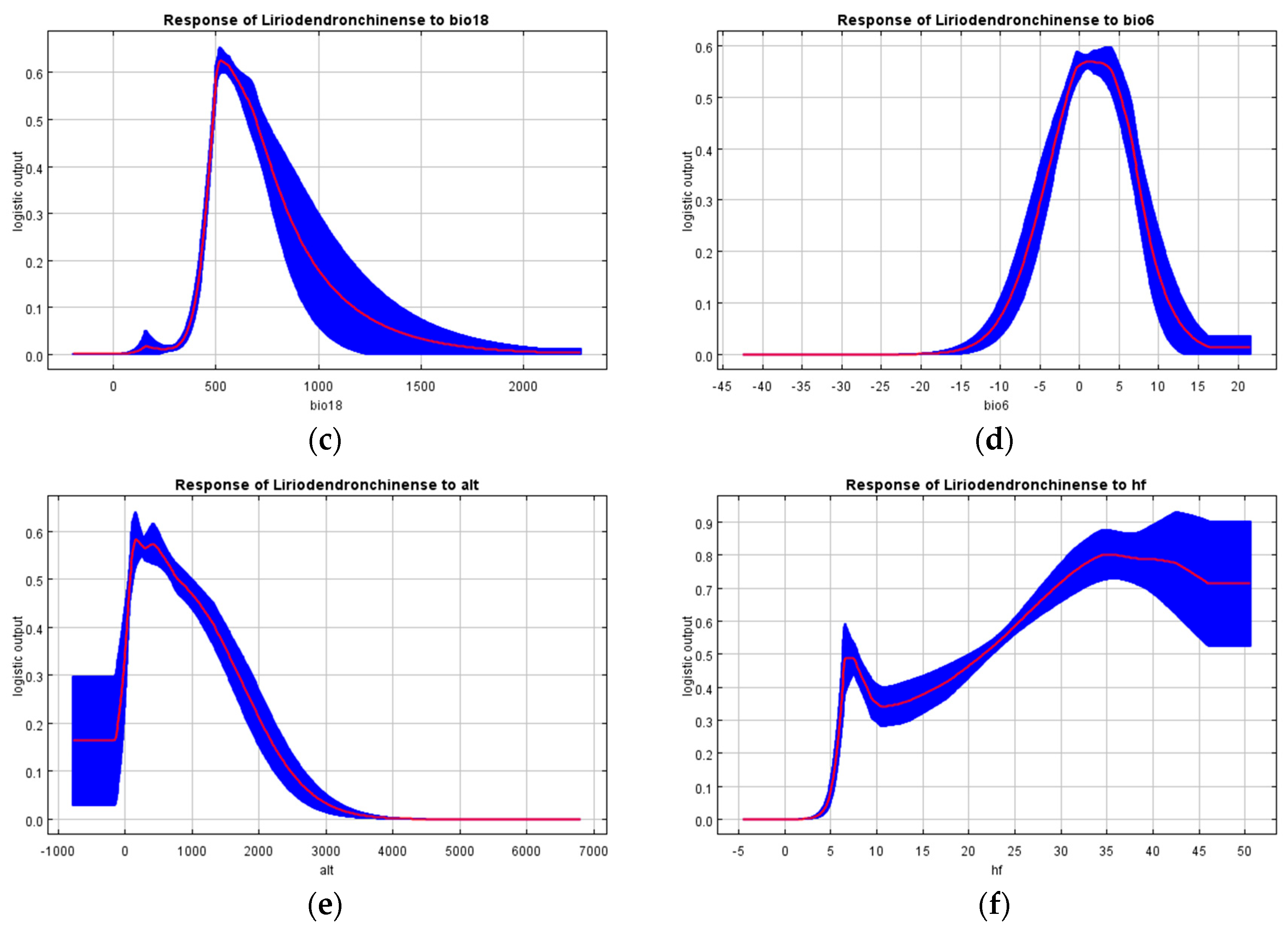
4.1. The Influence of Environmental Variables on the Modern Potential Geographic Distribution of L. chinense
4.2. Changes in the Geographic Distribution of L. chinense under Different Climate-Change Scenarios
4.3. Methodological Limitation
- (1)
- Data collection: This study only collected and analyzed distribution data within China. Hence, the environmental factor response curves may not fully reflect the response of L. chinense to environmental changes. Moreover, the species distribution data covers the years 1970–2020, which does not completely synchronize with the climate data spanning from 1970 to 2000. This temporal mismatch is particularly relevant given that L. chinense, as a woody plant, exhibits a longer growth cycle, necessitating a broader time range to gather a comprehensive dataset. Although such temporal discrepancies are common in ecological research and generally do not significantly alter the analytical outcomes—especially in studies focused on long-term climatic effects [65]—they could potentially limit the precision of our findings. For future studies, expanding the data collection to include global distribution data of L. chinense would enhance our understanding of its responses to diverse environmental conditions. Additionally, ensuring tighter temporal alignment between species distribution and climate data will improve the accuracy of our response curve analyses and make our conclusions more robust.
- (2)
- Temporal scope and forecasting limits: The future of climate change holds many uncertainties, and this study only utilizes the BCC-CMS2-MR GCM to simulate the potentially suitable habitats for L. chinense, which increases the instability of the MaxEnt model’s predictions and leads to certain errors in geographic distribution and area occurrence. This can affect decision-makers’ ability to formulate ecological protection measures. Previous studies have demonstrated that employing multiple GCMs significantly diminishes uncertainties and errors in predictions [66,67]. Future research on predicting the potential suitable habitats for L. chinense should also adopt this approach. Furthermore, our study only utilized climate scenario data for 2050, which may not adequately reflect changes in the suitable habitat of L. chinense over different future time periods. While we acknowledge that there are high-quality climate datasets available for nearer periods, such as 2020–2040, our choice was guided by the need to provide insights into future climatic conditions that match the time frames most relevant to policy and decision-making processes. Using the 2041–2060 period provides a balance between near-term forecasting and long-term strategic planning. To more accurately predict the potential geographic distribution changes in L. chinense, it is recommended that future research employs dynamic analysis and comparison at multiple time points.
- (3)
- Consideration of ecological factors: In predicting the potential suitable areas for L. chinense, this study primarily focused on factors such as climate, altitude, and human footprint without fully considering biological factors like interspecies interactions. Given the existence of interactions and dependencies between species [68], these complex ecological relationships could impact the actual climate adaptability and survival conditions of species [69]. Therefore, the predicted potentially suitable areas might deviate from actual conditions. Future research should explore a wider range of ecological factors, including soils, land cover, and interspecies interactions, in greater detail to enhance the accuracy of predictions regarding the potentially suitable areas for L. chinense.
5. Conclusions
- (1)
- Under current climate conditions, the potential suitable habitat area for L. chinense spans 147.55 × 104 km2, accounting for 15.01% of China’s land area. It is primarily distributed in central, southern, and southwestern China, with cultivation also present in Taiwan Province.
- (2)
- Precipitation in the driest month, precipitation in the driest quarter, precipitation in the warmest quarter, minimum temperature of the coldest month, altitude, and human footprint are the dominant factors influencing the suitable habitat distribution of L. chinense. Among these, precipitation in the driest month (bio14) is the most critical determinant of the suitable environment for L. chinense.
- (3)
- Under future climate scenarios, with increasing carbon concentrations, the distribution range of L. chinense is expected to initially expand and then contract, reaching its maximum expansion under the SSP370 emission scenario.
- (4)
- In the future 2050s climate scenarios, the centroid of L. chinense’s distribution is predicted to shift from lower latitudes to higher latitudes in the north and from lower to higher elevations.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021. [Google Scholar]
- Xiang, J.; Zhang, L.; Deng, Y.; She, D.; Zhang, Q. Projection and evaluation of extreme temperature and precipitation in major regions of China by CMIP6 models. Eng. J. Wuhan Univ. 2021, 54, 46–57. [Google Scholar]
- Ostad-Ali-Askar, K.; Su, R.; Liu, L. Editorial: Water resources and climate change. J. Water Clim. Change 2018, 9, 239. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Berry, J.A.; Smith, D.D.; Sperry, J.S.; Anderegg, L.D.L.; Field, C.B. The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off. Proc. Natl. Acad. Sci. USA 2012, 109, 233–237. [Google Scholar] [CrossRef]
- Scholze, M.; Knorr, W.; Arnell, N.W.; Prentice, I.C. A climate-change risk analysis for world ecosystems. Proc. Natl. Acad. Sci. USA 2006, 103, 13116–13120. [Google Scholar] [CrossRef]
- Vermeiren, P.; Reichert, P.; Schuwirth, N. Integrating uncertain prior knowledge regarding ecological preferences into multi-species distribution models: Effects of model complexity on predictive performance. Ecol. Model. 2020, 420, 108956. [Google Scholar] [CrossRef]
- Chen, W.; Wei, J.; Zhu, K.; Lu, Y.; Cai, Y.; Wu, Q.; Huang, Z.; Wang, Y. Predicting potential distribution of Emmenopterys henryi in Southwest China based on the Maxent model and influencing factors. Trop. Ecol. 2022, 63, 572–583. [Google Scholar] [CrossRef]
- He, Y.; Ma, J.; Chen, G. Potential geographical distribution and its multi-factor analysis of Pinus massoniana in China based on the maxent model. Ecol. Indic. 2023, 154, 110790. [Google Scholar] [CrossRef]
- Xu, L.; Fan, Y.; Zheng, J.; Guan, J.; Lin, J.; Wu, J.; Liu, L.; Wu, R.; Liu, Y. Impacts of climate change and human activity on the potential distribution of Aconitum leucostomum in China. Sci. Total Environ. 2024, 912, 168829. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar] [CrossRef]
- Zhao, X.; Lei, M.; Wei, C.; Guo, X. Assessing the suitable regions and the key factors for three Cd-accumulating plants (Sedum alfredii, Phytolacca americana, and Hylotelephium spectabile) in China using MaxEnt model. Sci. Total Environ. 2022, 852, 158202. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. BIOCLIM: The first species distribution modelling package, its early applications and relevance to most current MAXENT studies. Divers Distrib 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Qin, X.; Li, M. Predicting the Potential Distribution of Oxalis debilis Kunth, an Invasive Species in China with a Maximum Entropy Model. Plants 2023, 12, 3999. [Google Scholar] [CrossRef]
- Wang, G.; Geng, Q.; Xiao, M.; Zhang, M.; Wang, Z. Predicting Pseudolarix amabilis potential habitat based on four Niche models. Acta Ecol. Sin. 2020, 40, 6096–6104. [Google Scholar]
- Ashraf, U.; Peterson, A.T.; Chaudhry, M.N.; Ashraf, I.; Saqib, Z.; Ahmad, S.R.; Ali, H. Ecological niche model comparison under different climate scenarios: A case study of Olea spp. in Asia. Ecosphere 2017, 8, e01825. [Google Scholar] [CrossRef]
- Ye, X.; Zhao, G.; Zhang, M.; Cui, X.; Fan, H.; Liu, B. Distribution Pattern of Endangered Plant Semiliquidambar cathayensis (Hamamelidaceae) in Response to Climate Change after the Last Interglacial Period. Forests 2020, 11, 434. [Google Scholar] [CrossRef]
- Liu, C.; Huo, H.; Tian, L.; Dong, X.; Qi, D.; Zhang, Y.; Xu, J.; Cao, Y. Potential geographical distribution of Pyrus calleryana under different climate change scena-rios based on the MaxEnt model. Chin. J. Appl. Ecol. 2018, 29, 3696–3704. [Google Scholar]
- Dirr, M.A. Manual of Woody Landscape Plants: Their Identification, Ornamental Characteristics, Culture, Propagation and Uses; Stipes Publishing CoS: Champaign, IL, USA, 1990; p. 1007. [Google Scholar]
- Wei, P.; Jia, C.; Zhou, Y.; Gu, Y.; Luo, J. A study of wood physical and mechanical properties and vertical variation characteristics of natural Liriodendron. J. Sichuan For. Sci. Technol. 2018, 39, 27–31. [Google Scholar]
- Xue, W.; LI, C.; Ai, X.; Yao, L.; Guo, Q.; Zhu, J. Niche and interspecific association of dominant tree species in Liriodendron chinense natural forest. J. For. Environ. 2023, 43, 26–34. [Google Scholar]
- IUCN Species and Climate Change. Available online: https://www.iucn.org/resources/issues-brief/species-and-climate-change (accessed on 2 April 2024).
- IUCN The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 1 January 2024).
- Cao, Y.; Feng, J.; Hwarari, D.; Ahmad, B.; Wu, H.; Chen, J.; Yang, L. Alterations in Population Distribution of Liriodendron chinense (Hemsl.) Sarg. and Liriodendron tulipifera Linn. Caused by Climate Change. Forests 2022, 13, 488. [Google Scholar] [CrossRef]
- Qiu, H.; Sun, J.; Xu, D.; Shen, A.; Jiang, B.; Yuan, W.; Li, S. MaxEnt model-based prediction of potential distribution of Liriodendron chinense in China. J. Zhejiang A F Univ. 2020, 37, 1–8. [Google Scholar]
- Li, D.; Li, Z.; Liu, Z.; Yang, Y.; Khoso, A.G.; Wang, L.; Liu, D. Climate change simulations revealed potentially drastic shifts in insect community structure and crop yields in China’s farmland. J. Pest Sci. 2023, 96, 55–69. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Clim. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Venter, O.; Sanderson, E.W.; Magrach, A.; Allan, J.R.; Beher, J.; Jones, K.R.; Possingham, H.P.; Laurance, W.F.; Wood, P.; Fekete, B.M.; et al. Global terrestrial Human Footprint maps for 1993 and 2009. Sci. Data 2016, 3, 160067. [Google Scholar] [CrossRef] [PubMed]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2024, 9, 1937–1958. [Google Scholar] [CrossRef]
- Lee, H.; Calvin, K.; Dasgupta, D. Climate Change 2023: Synthesis Report; Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; The Australian National University: Canberra, ACT, Australia, 2023. [Google Scholar]
- Shi, X.; Wang, J.; Zhang, L.; Chen, S.; Zhao, A.; Ning, X.; Fan, G.; Wu, N.; Zhang, L.; Wang, Z. Prediction of the potentially suitable areas of Litsea cubeba in China based on future climate change using the optimized MaxEnt model. Ecol. Indic. 2023, 148, 110093. [Google Scholar] [CrossRef]
- Jarnevich, C.S.; Stohlgren, T.J.; Kumar, S.; Morisette, J.T.; Holcombe, T.R. Caveats for correlative species distribution modeling. Ecol. Inf. 2015, 29, 6–15. [Google Scholar] [CrossRef]
- Li, Y.; Shao, W.; Jiang, J. Predicting the potential global distribution of Sapindus mukorossi under climate change based on MaxEnt modelling. Environ. Sci. Pollut. Res. 2022, 29, 21751–21768. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Deng, C.; Duan, G.; Wang, Z.; Zhang, Y.; Fan, G. Potentially suitable habitats of Daodi goji berry in China under climate change. Front. Plant Sci. 2024, 14, 1279019. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, X.; Li, R.; Wang, X.; Cheng, J.; Yang, Q.; Kong, H. AHP-GIS and MaxEnt for delineation of potential distribution of Arabica coffee plantation under future climate in Yunnan, China. Ecol. Indic. 2021, 132, 108339. [Google Scholar] [CrossRef]
- Kass, J.M.; Muscarella, R.; Galante, P.J.; Bohl, C.L.; Pinilla-Buitrago, G.E.; Boria, R.A.; Soley-Guardia, M.; Anderson, R.P. ENMeval 2.0: Redesigned for customizable and reproducible modelling of species’ niches and distributions. Methods Ecol. Evol. 2021, 12, 1602–1608. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MAXENT ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Wan, G.; Wang, L.; Jin, L.; Chen, J. Evaluation of environmental factors affecting the quality of Codonopsis pilosula based on chromatographic fingerprint and MaxEnt model. Ind. Crop. Prod. 2021, 170, 113783. [Google Scholar] [CrossRef]
- Hou, J.; Xiang, J.; Li, D.; Liu, X. Prediction of Potential Suitable Distribution Areas of Quasipaa spinosa in China Based on MaxEnt Optimization Model. Biology 2023, 12, 366. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.C.S.; Lorini, M.L.; Minsky, E.M.C.; Figueiredo, M.S.L.; Santos, L.N. Far beyond the Amazon: Global distribution, environmental suitability, and invasive potential of the two most introduced peacock bass. Biol. Invasions 2022, 24, 2851–2872. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Preau, C.; Trochet, A.; Bertrand, R.; Isselin-Nondedeu, F. Modeling Potential Distributions of Three European Amphibian Species Comparing ENFA and Maxent. Herpetol. Conserv. Biol. 2018, 13, 91–104. [Google Scholar]
- Xu, Z.; Ding, G.; Liu, B.; Chi, C.; Xiao, W.; JIn, X.; Li, C. Geographic distribution and predicton of potential suitable regions of Iva xanthifolia. Acta Pratacult. Sin. 2012, 21, 75–83. [Google Scholar]
- Yang, J.; Huang, Y.; Jiang, X.; Chen, H.; Liu, M.; Wang, R. Potential geographical distribution of the edangred plant Isoetes under human activities using MaxEnt and GARP. Glob. Ecol. Conserv. 2022, 38, e02186. [Google Scholar] [CrossRef]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Roberts, D.R.; Bahn, V.; Ciuti, S.; Boyce, M.S.; Elith, J.; Guillera-Arroita, G.; Hauenstein, S.; Lahoz-Monfort, J.J.; Schroeder, B.; Thuiller, W.; et al. Cross-validation strategies for data with temporal, spatial, hierarchical, or phylogenetic structure. Ecography 2017, 40, 913–929. [Google Scholar] [CrossRef]
- Thuiller, W. Biodiversity-Climate change and the ecologist. Nature 2007, 448, 550–552. [Google Scholar] [CrossRef]
- Lopez-Pujol, J.; Zhang, F.; Sun, H.; Ying, T.; Ge, S. Centres of plant endemism in China: Places for survival or for speciation ? J. Biogeogr. 2011, 38, 1267–1280. [Google Scholar] [CrossRef]
- Elliott, K.J.; Miniat, C.F.; Pederson, N.; Laseter, S.H. Forest tree growth response to hydroclimate variability in the southern Appalachians. Glob. Change Biol. 2015, 21, 4627–4641. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Sarwar, R.; Zhang, W.; Geng, R.; Zhu, K.; Tan, X. Research progress on the physiological response and molecular mechanism of cold response in plants. Front. Plant Sci. 2024, 15, 1334913. [Google Scholar] [CrossRef] [PubMed]
- Cano-Ramirez, D.L.; Carmona-Salazar, L.; Morales-Cedillo, F.; Ramirez-Salcedo, J.; Cahoon, E.B.; Gavilanes-Ruiz, M. Plasma Membrane Fluidity: An Environment Thermal Detector in Plants. Cells 2021, 10, 2778. [Google Scholar] [CrossRef]
- Amat, M.E.; Vargas, P.; Gomez, J.M. Effects of human activity on the distribution and abundance of an endangered Mediterranean high-mountain plant (Erysimum penyalarense). J. Nat. Conserv. 2013, 21, 262–271. [Google Scholar] [CrossRef]
- Ghini, R.; Hamada, E.; Angelotti, F.; Costa, L.B.; Bettiol, W. Research approaches, adaptation strategies, and knowledge gaps concerning the impacts of climate change on plant diseases. Trop. Plant Pathol. 2012, 37, 5–24. [Google Scholar]
- Jo, W.S.; Kim, H.; Kim, B.J. Climate change alters diffusion of forest pest: A model study. J. Korean Phys. Soc. 2017, 70, 108–115. [Google Scholar] [CrossRef]
- Guan, X.; Shi, W.; Cao, K. Effect of Climate Change in Future on Geographical Distribution of Widespread Quercus acutissima and Analysis of Dominant Climatic Factors. J. Trop. Subtrop. Bot. 2018, 26, 661–668. [Google Scholar]
- Kim, G.N.; Han, S. Effects on Growth, Photosynthesis and Pigment Contents of Liriodendron tulipifera under Elevated Temperature and Drought. Korean J. Agric. For. Meteorol. 2015, 1, 75–84. [Google Scholar] [CrossRef]
- Liu, M.; Yang, L.; Su, M.; Gong, W.; Liu, Y.; Yang, J.; Huang, Y.; Zhao, C. Modeling the potential distribution of the energy tree species Triadica sebifera in response to climate change in China. Sci. Rep. 2024, 14, 1220. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhang, X.; Wang, J. IPCC-AR4 Model Projections of Climate Change Scenarios for China in the 21st Century. Geogr. Res.-Aust. 2008; 7, 787–799. [Google Scholar]
- Schaufler, G.; Kitzler, B.; Schindlbacher, A.; Skiba, U.; Sutton, M.A.; Zechmeister-Boltenstern, S. Greenhouse gas emissions from European soils under different land use: Effects of soil moisture and temperature. Eur. J. Soil Sci. 2010, 61, 683–696. [Google Scholar] [CrossRef]
- Li, X.Z.; Lilt, X.D. Numerical Simulations of Extreme Precipitation in Eastern China under A1B Scenario. J. Trop. Meteorol. 2014, 20, 121–134. [Google Scholar]
- Zhang, H.; Zhao, H.; Wang, H. Potential geographical distribution of Populus euphratica in China under future climate change scenarios based on Maxent model. Acta Ecol. Sin. 2020, 40, 6552–6563. [Google Scholar]
- Prevey, J.S.; Parker, L.E.; Harrington, C.A.; Lamb, C.T.; Proctor, M.F. Climate change shifts in habitat suitability and phenology of huckleberry (Vaccinium membranaceum). Agric. For. Meteorol. 2020, 280, 107803. [Google Scholar] [CrossRef]
- Yu, D.; Fan, Z.; Ma, C. Distribution of Suitable Areas of Torreya yunnanensis under Different Carbon Emission Mode. J. Northwest For. Univ. 2023, 38, 40–46. [Google Scholar]
- Esperon-Rodriguez, M.; Power, S.A.; Tjoelker, M.G.; Beaumont, L.J.; Burley, H.; Caballero-Rodriguez, D.; Rymer, P.D. Assessing the vulnerability of Australia’s urban forests to climate extremes. Plants People Planet 2019, 1, 387–397. [Google Scholar] [CrossRef]
- Guan, J.; Li, M.; Ju, X.; Lin, J.; Wu, J.; Zheng, J. The potential habitat of desert locusts is contracting: Predictions under climate change scenarios. Peerj 2021, 9, e12311. [Google Scholar] [CrossRef]
- Li, J.; Chang, H.; Liu, T.; Zhang, C. The potential geographical distribution of Haloxylon across Central Asia under climate change in the 21st century. Agric. For. Meteorol. 2019, 275, 243–254. [Google Scholar] [CrossRef]
- Faeth, S.H.; Warren, P.S.; Shochat, E.; Marussich, W.A. Trophic dynamics in urban communities. Bioscience 2005, 55, 399–407. [Google Scholar] [CrossRef]
- Jenerette, G.D.; Clarke, L.W.; Avolio, M.L.; Pataki, D.E.; Gillespie, T.W.; Pincetl, S.; Nowak, D.J.; Hutyra, L.R.; McHale, M.; McFadden, J.P.; et al. Climate tolerances and trait choices shape continental patterns of urban tree biodiversity. Glob. Ecol. Biogeogr. 2016, 25, 1367–1376. [Google Scholar] [CrossRef]
- Zhao, Z.; Xiao, N.; Shen, M.; Li, J. Comparison between optimized MaxEnt and random forest modeling in predicting potential distribution: A case study with Quasipaa boulengeri in China. Sci. Total Environ. 2022, 842, 156867. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Description | Abbreviation | Unit |
|---|---|---|
| Annual mean temperature | bio1 | °C |
| Mean diurnal range | bio2 | °C |
| Isothermality (×100) | bio3 | - |
| Temperature seasonality | bio4 | - |
| Max temperature of warmest month | bio5 | °C |
| Min temperature of coldest month | bio6 | °C |
| Temperature annual range | bio7 | °C |
| Mean temperature of wettest quarter | bio8 | °C |
| Mean temperature of driest quarter | bio9 | °C |
| Mean temperature of warmest quarter | bio10 | °C |
| Mean temperature of coldest quarter | bio11 | °C |
| Annual precipitation | bio12 | mm |
| Precipitation of wettest month | bio13 | mm |
| Precipitation of driest month | bio14 | mm |
| Precipitation seasonality | bio15 | mm |
| Precipitation of wettest quarter | bio16 | mm |
| Precipitation of driest quarter | bio17 | mm |
| Precipitation of warmest quarter | bio18 | mm |
| Precipitation of coldest quarter | bio19 | mm |
| Elevation | elev. | m |
| Human footprint | hf | - |
| Parameter Settings | Regularization Multiplier | Feature Combination | Δ.AICc | auc.diff.avg | or.10p.avg |
|---|---|---|---|---|---|
| Default | 1 | LQPHT | 153.929412 | 0.053008106 | 0.321969697 |
| Optimized | 0.5 | LQ | 0 | 0.059019351 | 0.207251082 |
| Environment Variables | Percent Contribution (%) | Permutation Importance (%) | Total Suitable Range |
|---|---|---|---|
| bio2 | 1.4 | 1.6 | 5.96–7.93 °C |
| bio3 | 4.5 | 2.4 | 23.07%–26.76%, 52.33%–56.78% |
| bio4 | 3.1 | 3.7 | 678–843, 200–377 |
| bio6 | 5.0 | 21.8 | −1.38–6.70 °C |
| bio8 | 0.9 | 1.2 | 20.52–25.60 °C |
| bio12 | 8.2 | 4.4 | 1140.55–2175.97 mm |
| bio14 | 34.9 | 7.1 | 18.18–177.02 mm |
| bio15 | 0.9 | 1.2 | 44.34%–67.94% |
| bio16 | 0.4 | 4.4 | 510.45–966.62 mm |
| bio17 | 21 | 9.4 | 127.22–564.30 mm |
| bio18 | 1.2 | 23.8 | 520.37–718.70 mm |
| Elev. | 9.0 | 10.1 | 80–900 m |
| Hf | 10.3 | 9.8 | 22.45–49.44 |
| Climate Periods | Highly Suitable Area | Medium-Suitable Area | Low Suitable Area | Total Suitable Area |
|---|---|---|---|---|
| Current | 35.52 | 61.03 | 55.00 | 151.55 |
| 2050-SSP126 | 51.34 | 71.82 | 62.49 | 185.65 |
| 2050-SSP245 | 55.35 | 87.88 | 74.02 | 217.24 |
| 2050-SSP370 | 86.03 | 113.93 | 86.92 | 286.88 |
| 2050-SSP585 | 69.04 | 109.17 | 87.36 | 265.57 |
| Climate Scenarios | Migration Distance of Centroid Shift (m) | Latitude (°) | Longitude (°) | Elevation (m) |
|---|---|---|---|---|
| Current | — | 111.123288 | 28.000177 | 189 |
| 2050-SSP126 | 184,214.8732 | 111.466436 | 29.619396 | 57 |
| 2050-SSP245 | 239,097.4455 | 110.951573 | 30.130432 | 983 |
| 2050-SSP370 | 446,488.6247 | 110.883370 | 31.975295 | 731 |
| 2050-SSP585 | 305,669.3452 | 110.718422 | 30.706310 | 1165 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, J.; Wang, H.; Hu, Y. Prediction of Potential Suitable Distribution of Liriodendron chinense (Hemsl.) Sarg. in China Based on Future Climate Change Using the Optimized MaxEnt Model. Forests 2024, 15, 988. https://doi.org/10.3390/f15060988
Bai J, Wang H, Hu Y. Prediction of Potential Suitable Distribution of Liriodendron chinense (Hemsl.) Sarg. in China Based on Future Climate Change Using the Optimized MaxEnt Model. Forests. 2024; 15(6):988. https://doi.org/10.3390/f15060988
Chicago/Turabian StyleBai, Jieyuan, Hongcheng Wang, and Yike Hu. 2024. "Prediction of Potential Suitable Distribution of Liriodendron chinense (Hemsl.) Sarg. in China Based on Future Climate Change Using the Optimized MaxEnt Model" Forests 15, no. 6: 988. https://doi.org/10.3390/f15060988
APA StyleBai, J., Wang, H., & Hu, Y. (2024). Prediction of Potential Suitable Distribution of Liriodendron chinense (Hemsl.) Sarg. in China Based on Future Climate Change Using the Optimized MaxEnt Model. Forests, 15(6), 988. https://doi.org/10.3390/f15060988