

Seed Propagation of Carapa amorphocarpa W. Palacios Using Various Treatments of Substrates and Mechanical Seed Scarification in a Nursery in the Andean Area of Northwestern Ecuador
 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Location
2.2. Substrate Preparation
- ▪
- Substrate 1 (S1): Forest soil mixed with sand in a 3:1 ratio.
- ▪
- Substrate 2 (S2): Forest soil alone.
2.3. Fruits and Seeds Collection
2.4. Treatments of the Carapa amorphocarpa Seeds
2.5. Statistical Analysis
3. Results
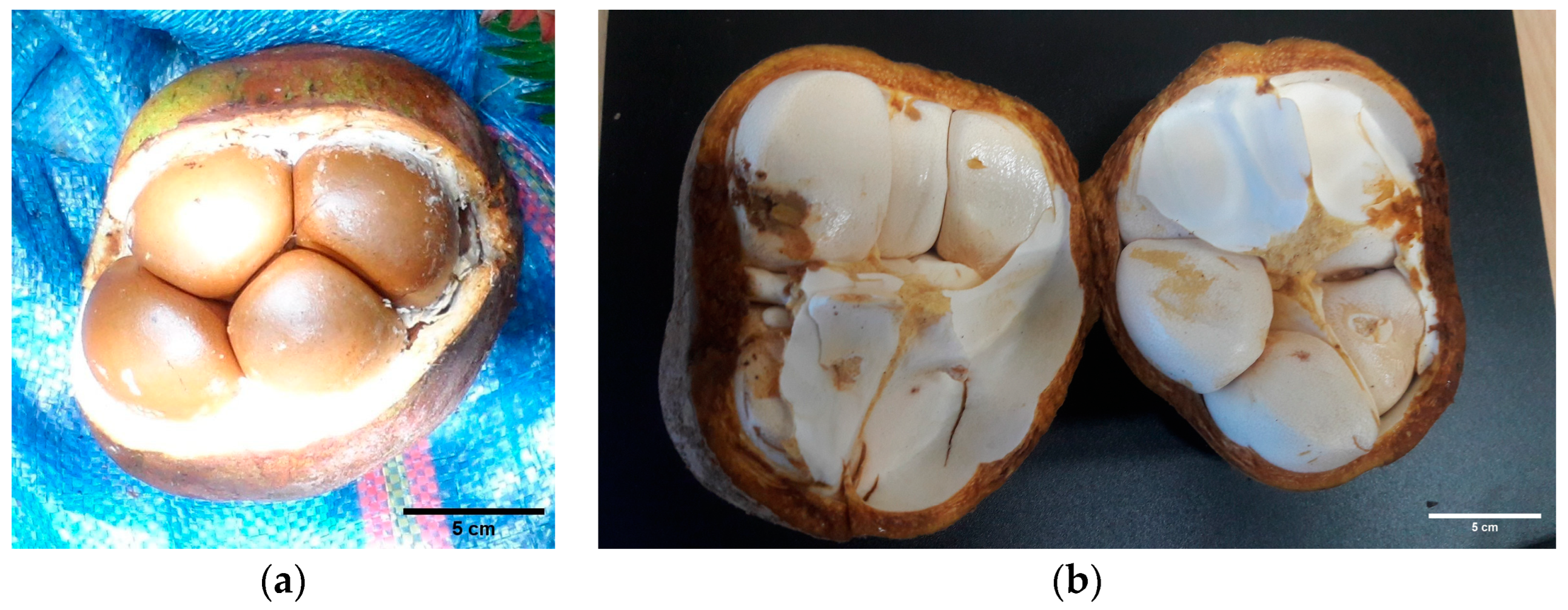
3.1. General Characteristics of Fruits and Seeds
3.2. Preliminary Germination Tests
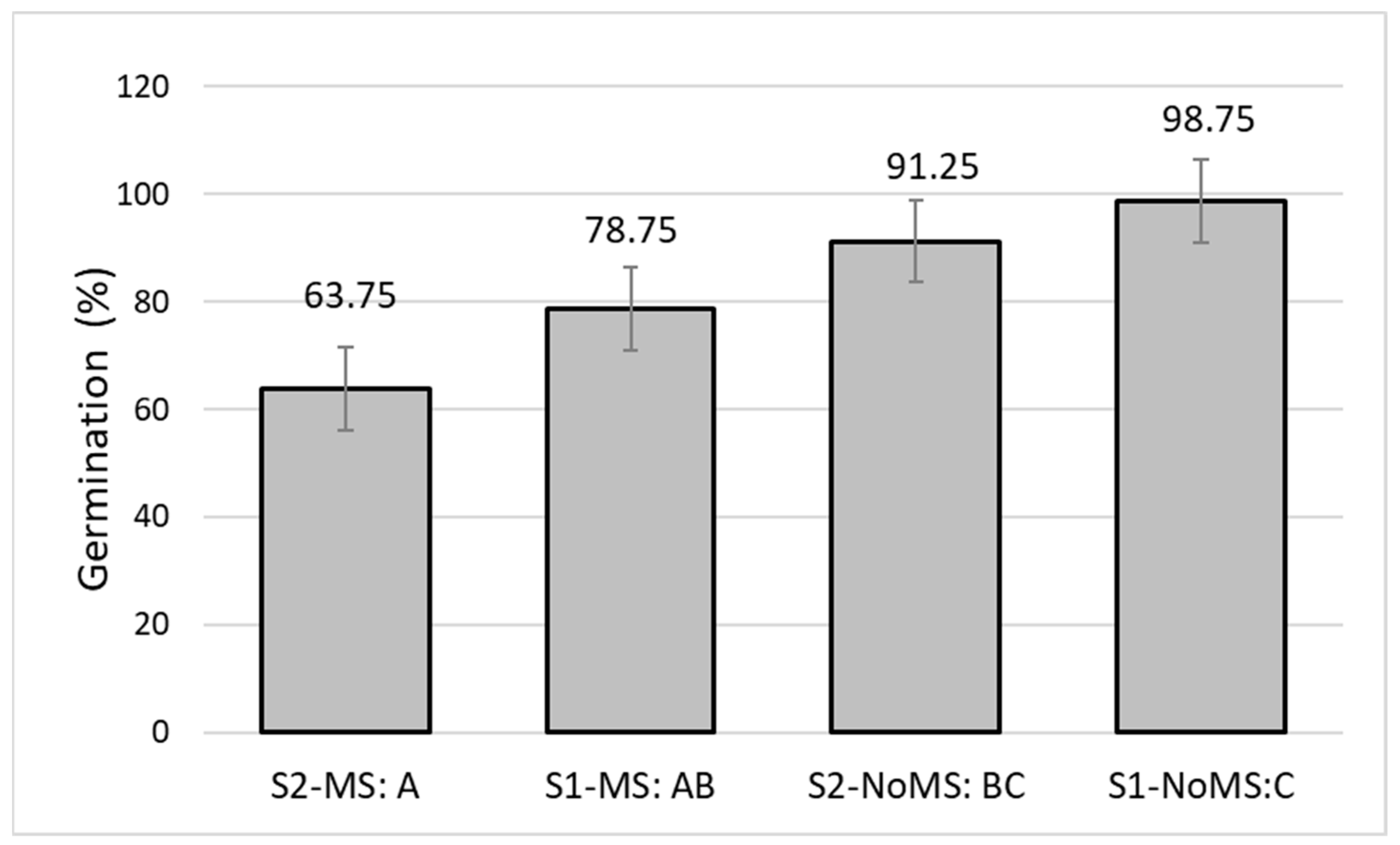
3.3. Germination Capacity in the Main Test
3.4. Growth in Height
3.5. Growth in Diameter
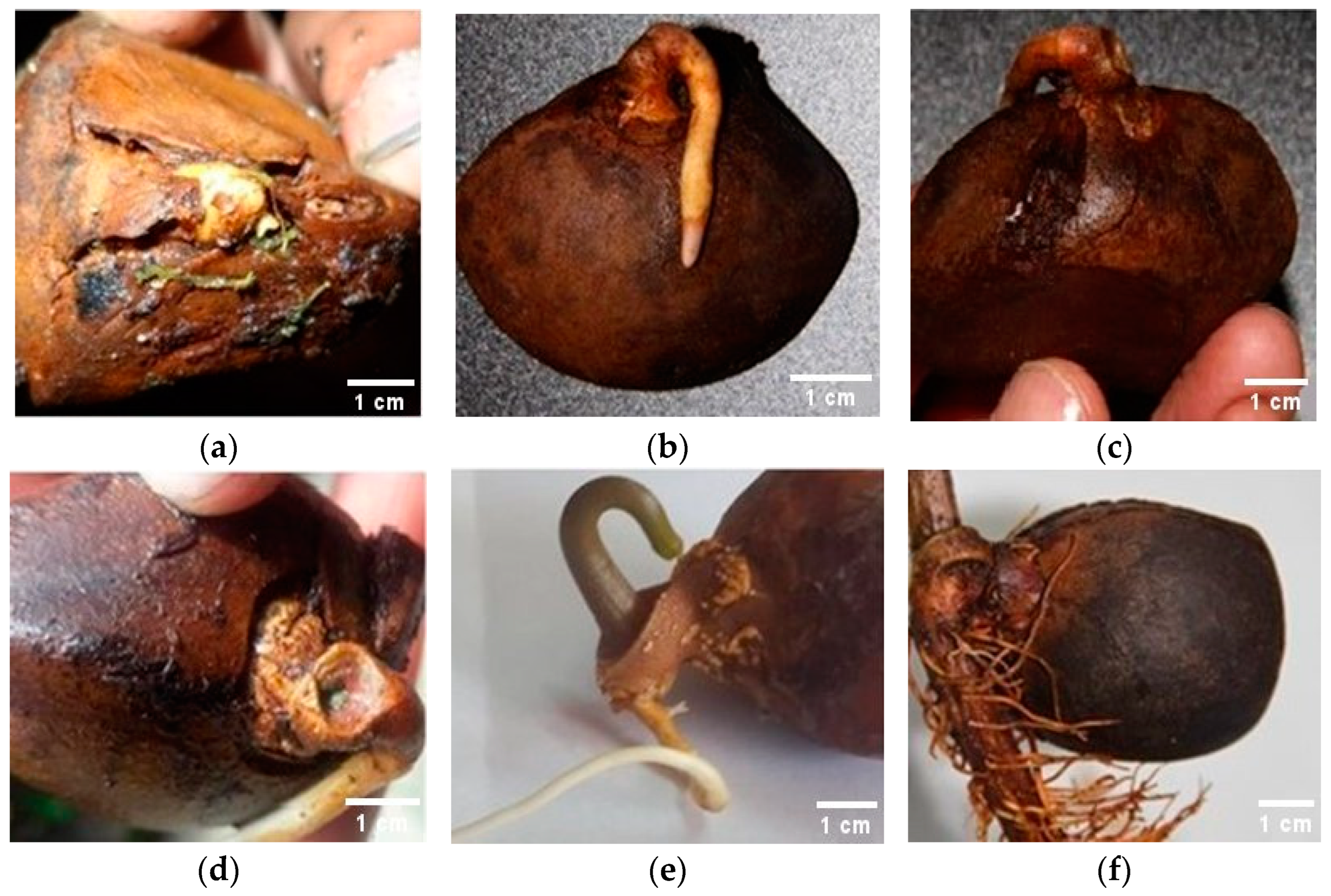
3.6. Phytosanitary Status and Shape of the Seedlings
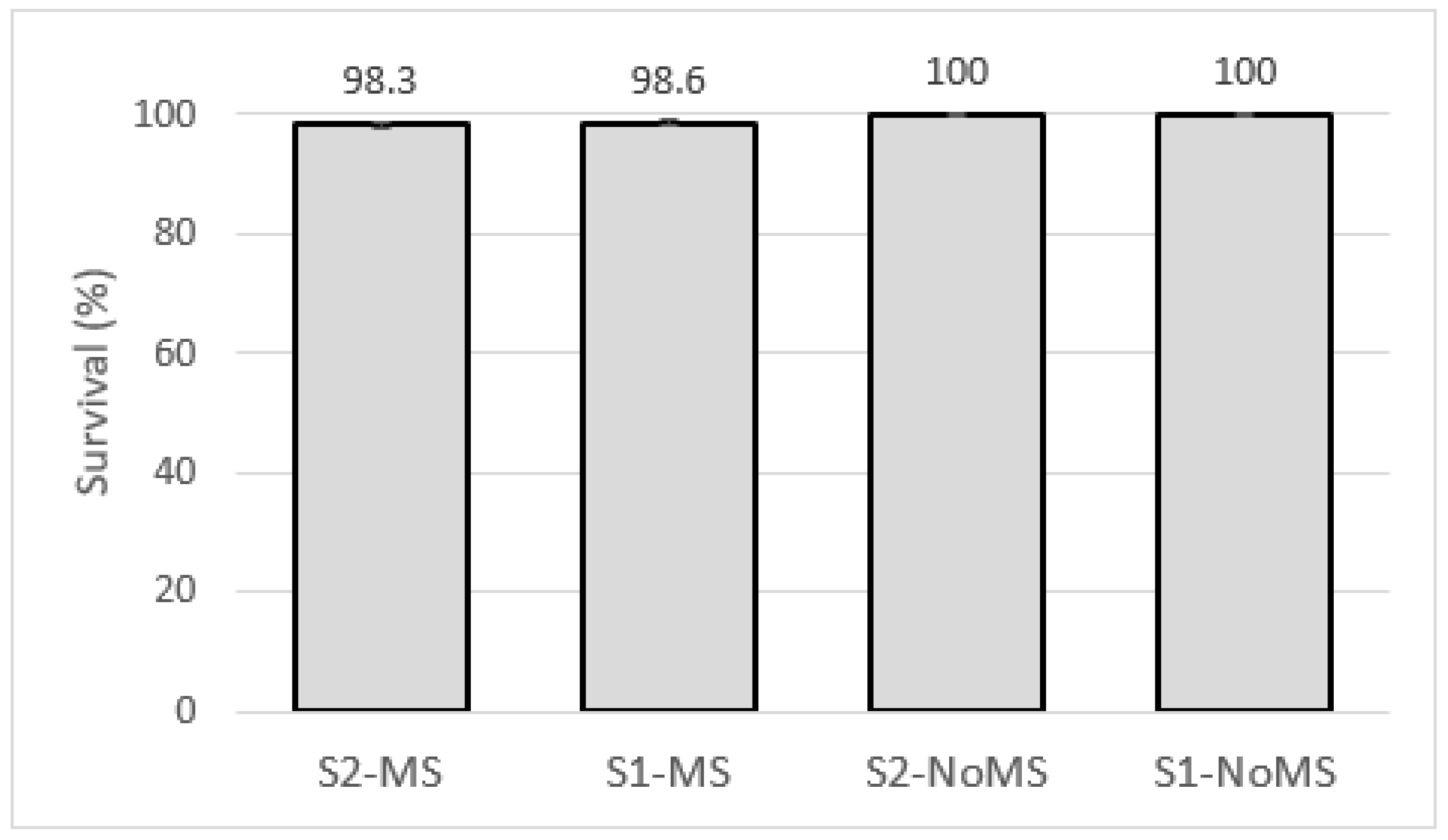
3.7. Survival
3.8. Correlations
3.9. Propagation Costs
4. Discussion
4.1. General Characteristics of Fruits and Seeds of Carapa amorphocarpa
4.2. Germination Trials and Establishment of the Experiment
4.3. Germination of Carapa amorphocarpa Seeds and Seedling Growth in Height and Diameter
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Oguh, C.E.; Obiwulu, E.N.O.; Umezinwa, O.J.; Ameh, S.E.; Ugwu, C.V.; Sheshi, I.M. Ecosystem and ecological services; need for biodiversity conservation-a critical review. Asian J. Biol. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Palacios, W.; Jaramillo, N. Árboles amenazados del Chocó ecuatoriano. ACI Av. Cienc. Ing. 2016, 8, 1. [Google Scholar]
- Astudillo-Sánchez, E.; Pérez, J.; Troccoli, L.; Aponte, H.; Tinoco, O. Flora leñosa del bosque de garúa de la cordillera Chongón Colonche, Santa Elena-Ecuador. Ecol. Aplicada 2019, 18, 155–169. [Google Scholar] [CrossRef]
- Alarcón-Aguirre, J.S.; Aguirre-Mejía, P.M.; Palacios-Hinestroza, H.; Sulbarán-Rangel, B. Evaluation of the forestry administrative system that regulates the activity of extracting wood from the Northern Ecuadorian amazon. Land. Use Policy 2020, 99, 104852. [Google Scholar] [CrossRef]
- Bastin, J.-F.; Finegold, Y.; Garcia, C.; Mollicone, D.; Rezende, M.; Routh, D.; Zohner, C.M.; Crowther, T.W. The global tree restoration potential. Science 2019, 365, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Cué García, J.L.; Añazco, M.J.; Orlando Paredes, H. Producción y conservación de semillas forestales: Situación actual y perspectivas en Ecuador. Rev. Cuba. Cienc. For. 2019, 7, 365–376. [Google Scholar]
- Hua, F.; Bruijnzeel, L.A.; Meli, P.; Martin, P.A.; Zhang, J.; Nakagawa, S.; Miao, X.; Wang, W.; McEvoy, C.; Peña-Arancibia, J.L. The biodiversity and ecosystem service contributions and trade-offs of forest restoration approaches. Science 2022, 376, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Tonguç, M.; Önder, S.; Gülcemal, N.; Tonguç, F. Seed, germination, and seed-reserve traits differ along an altitudinal gradient. J. Res. 2022, 33, 1903–1912. [Google Scholar] [CrossRef]
- Akaffou, S.D.; Kouame, A.K.; Gore, N.B.B.; Abessika, G.Y.; Kouassi, H.K.; Hamon, P.; Sabatier, S.; Duminil, J. Effect of the seeds provenance and treatment on the germination rate and plants growth of four forest trees species of Côte d’Ivoire. J. Res. 2021, 32, 161–169. [Google Scholar] [CrossRef]
- Li, R.; Yan, Q.; Xie, J.; Wang, J.; Zhang, T.; Zhu, J. Effects of logging on the trade-off between seed and sprout regeneration of dominant woody species in secondary forests of the Natural Forest Protection Project of China. Ecol. Process 2022, 11, 16. [Google Scholar] [CrossRef]
- Palacios, W. Cuatro especies nuevas de árboles del Ecuador. Caldasia 2012, 34, 75–85. [Google Scholar]
- Londres, M.; Schulze, M.; Staudhammer, C.L.; Kainer, K.A. Population structure and fruit production of Carapa guianensis (Andiroba) in Amazonian floodplain forests: Implications for community-based management. Trop. Conserv. Sci. 2017, 10, 1940082917718835. [Google Scholar] [CrossRef]
- Dias, K.K.B.; Cardoso, A.L.; da Costa, A.A.F.; Passos, M.F.; da Costa, C.E.F.; da Rocha Filho, G.N.; de Aguiar Andrade, E.H.; Luque, R.; do Nascimento, L.A.S.; Noronha, R.C.R. Biological activities from andiroba (Carapa guianensis Aublet.) and its biotechnological applications: A systematic review. Arab. J. Chem. 2023, 16, 104629. [Google Scholar] [CrossRef]
- Kenfack, D. A synoptic revision of Carapa (Meliaceae). Harv. Pap. Bot. 2011, 16, 171–231. [Google Scholar] [CrossRef]
- Gobierno Provincial del Carchi. Plan de Desarrollo y Ordenamiento Territorial de la Provincia del Carchi Actualización 2019–2023. Fase Diagnóstico 2019. Available online: https://carchi.gob.ec/2016f/index.php/mega-menu-2/category/115-plan-de-desarrollo-y-ordenamiento-territorial.html (accessed on 25 May 2022).
- Pennington, T.D. A Monograph of Neotropical Meliaceae (with Accounts of the Subfamily Swietenioideae by BT Styles and the Chemotaxonomy by DAH Taylor); Flora Neotropica, No. 28; New York Botanical Garden: Bronx, NY, USA, 1981; 470p, ISBN 0-89327-235-3. [Google Scholar]
- Salazar, R.; Soihet, C.; Méndez, J.M. Manejo de Semillas de 100 Especies Forestales de América Latina; Centro Agronómico de Investigación y Enseñanza (CATIE): Turrialba, Costa Rica, 2000. [Google Scholar]
- Klimas, C.A.; Wadt, L.H.d.O.; Castilho, C.V.d.; Lira-Guedes, A.C.; da Costa, P.; da Fonseca, F.L. Variation in Seed Harvest Potential of Carapa guianensis Aublet in the Brazilian Amazon: A Multi-Year, Multi-Region Study of Determinants of Mast Seeding and Seed Quantity. Forests 2021, 12, 683. [Google Scholar] [CrossRef]
- Orlovic Lovren, V. Traditional and Indigenous Knowledge: Bridging Past and Future Sustainable Development. In Life on Land; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1033–1041. [Google Scholar]
- Camara, B.; Ndiaye, S.; Diédhiou, M.A.A.; Goudiaby, A.O.K.; Kande, M.; Fall, F.; Ngom, D. Croissance et Développement de Carapa procera DC. sur différents types de terreau en pépinière en Basse Casamance (Sénégal). Int. J. Biol. Chem. Sci. 2023, 17, 1006–1019. [Google Scholar] [CrossRef]
- Gonçalves, M.C.A.; Yokomizo, G.K.I.; Segovia, J.F.O.; dos Santos, E.C. Seedlings germination aspects of the Carapa guianensis and its ecological conservation for the Amazonian peoples. Mundo Amaz. 2023, 14, e101653. [Google Scholar] [CrossRef]
- Voui, B.B.N.B.; Coulibaly, S. Vegetative propagation trial of Carapa procera CD (Meliaceae), a spontaneous species with multiple uses (Daloa, Central-West, Côte d’Ivoire). GSC Adv. Res. Rev. 2022, 13, 158–169. [Google Scholar]
- Bettoni, J.C.; van der Walt, K.; Souza, J.A.; McLachlan, A.; Nadarajan, J. Sexual and asexual propagation of Syzygium maire, a critically endangered Myrtaceae species of New Zealand. N. Z. J. Bot. 2024, 62, 35–52. [Google Scholar] [CrossRef]
- Sagwal, S.S. Forest Tree Seeds: Handbook; Scientific Publishers: Jodhpur, India, 2020. [Google Scholar]
- Nascimento, G.O.; Souza, D.P.; Santos, A.S.; Batista, J.F.; Rathinasabapathi, B.; Gagliardi, P.R.; Gonçalves, J.F.C. Lipidomic profiles from seed oil of Carapa guianensis Aubl. and Carapa vasquezii Kenfack and implications for the control of phytopathogenic fungi. Ind. Crops Prod. 2019, 129, 67–73. [Google Scholar] [CrossRef]
- Claire, A.-L. Remoción y germinación de semillas de Dipteryx panamensis y Carapa guianensis en bosques fragmentados de Sarapiquí, Costa Rica. Recur. Nat. Y Ambiente 2001, 10, 42–46. [Google Scholar]
- Forget, P.-M. Removal of seeds of Carapa procera (Meliaceae) by rodents and their fate in rainforest in French Guiana. J. Trop. Ecol. 1996, 12, 751–761. [Google Scholar] [CrossRef]
- Yadok, B.G.; Forget, P.-M.; Gerhard, D.; Aliyu, B.; Chapman, H. Seed nutrient content rather than size influences seed dispersal by scatterhoarding rodents in a West African montane forest. J. Trop. Ecol. 2020, 36, 174–181. [Google Scholar] [CrossRef]
- Morales Puentes, M.E. El género Carapa Aubl.(Meliaceae) en Colombia. Caldasia 1997, 19, 397–407. [Google Scholar]
- Gough, R.E. Seed Quality: Basic Mechanisms and Agricultural Implications; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Franić, I.; Cleary, M.; Aday Kaya, A.G.; Bragança, H.; Brodal, G.; Cech, T.L.; Chandelier, A.; Doğmuş-Lehtijärvi, T.; Eschen, R.; Lehtijärvi, A. The Biosecurity Risks of International Forest Tree Seed Movements. Curr. For. Rep. 2023, 10, 89–102. [Google Scholar] [CrossRef]
- Pillai, P.K.C. Seed Handling of Tropical Forestry Species. In Diversity and Dynamics in Forest Ecosystems; Apple Academic Press: Williston, VT, USA, 2021; pp. 323–349. [Google Scholar]
- Douglas, J.E. Successful Seed Programs: A Planning and Management Guide; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Delgado, F. Especies Forestales Nativas en los Andes Ecuatorianos: Resultados Preliminares de Algunas Experiencias, 2nd ed.; CESA: Quito, Ecuador, 1989; p. 50. [Google Scholar]
- R Core Team. RStudio: Integrated Development for R, Version 4.1.1; RStudio, PBC: Boston, MA, USA, 2021.
- Hanusz, Z.; Tarasinska, J.; Zielinski, W. Shapiro-Wilk test with known mean. REVSTAT-Stat. J. 2016, 14, 89–100. [Google Scholar]
- Tukey, J. Multiple comparisons. J. Am. Stat. Assoc. 1953, 48, 624–625. [Google Scholar]
- Zimmerman, D.W.; Zumbo, B.D. Relative power of the Wilcoxon test, the Friedman test, and repeated-measures ANOVA on ranks. J. Exp. Educ. 1993, 62, 75–86. [Google Scholar] [CrossRef]
- Bashalkhanov, S.; Pandey, M.; Rajora, O.P. A simple method for estimating genetic diversity in large populations from finite sample sizes. BMC Genet. 2009, 10, 1–10. [Google Scholar] [CrossRef]
- Ferraz, I.D.K.; Camargo, J.L.C.; Sampaio, P.d.T.B. Sementes e plântulas de andiroba (Carapa guianensis Aubl. e Carapa procera DC): Aspectos botânicos, ecológicos e tecnológicos. Acta Amaz. 2002, 32, 647. [Google Scholar]
- Vozzo, J.A. Manual de Semillas de Árboles Tropicales; Departamento de Agricultura de Estados Unidos, Servicio Forestal: Washington, DC, USA, 2010. [Google Scholar]
- McHargue, L.A.; Hartshorn, G.S. Seed and seedling ecology of Carapa guianensis. Turrialba 1983, 33, 399–404. [Google Scholar]
- Smith, C.E. Flora of Panama. Part VI. Family 92. Meliaceae. Ann. Mo. Bot. Gard. 1965, 52, 55–79. [Google Scholar] [CrossRef]
- Rishi, R.R.; Pavithra, S.; Sundararaj, R.; Athulya, R.; Prasad, N.R. Mahogany shoot borer, Hypsipyla robusta Moore (Lepidoptera: Pyralidae) and its food plants-A review. J. Entomol. Res. 2022, 46, 805–809. [Google Scholar] [CrossRef]
- De Vitis, M.; Hay, F.R.; Dickie, J.B.; Trivedi, C.; Choi, J.; Fiegener, R. Seed storage: Maintaining seed viability and vigor for restoration use. Restor. Ecol. 2020, 28, S249–S255. [Google Scholar] [CrossRef]
- Bacca Acosta, P.P.; Zuluaga Peláez, J.J.; Castaño Colorado, Á.A.; Ardila Fernández, A.F. Growth potential of Carapa guianensis (Aubl.) in Tumaco, Colombia. Rev. Chapingo Ser. Cienc. For. 2021, 27, 401–412. [Google Scholar]
- Pritchard, H.W.; Tsan, F.Y.; Wen, B.; Jaganathan, G.K.; Calvi, G.; Pence, V.C.; Mattana, E.; Ferraz, I.D.K.; Seal, C.E. Regeneration in recalcitrant-seeded species and risks from climate change. In Plant Regeneration from Seeds; Elsevier: Amsterdam, The Netherlands, 2022; pp. 259–273. [Google Scholar]
- Connor, K.F.; Bonner, F.T.; Vozzo, J.A.; Kossmann-Ferraz, I.D. Changes in desiccating seeds of temperate and tropical forest tree species. In Frontiers of Forest Biology; CRC Press: Boca Raton, FL, USA, 2021; pp. 319–326. [Google Scholar]
- Cordero, J.; Boshier, D.; Barrance, A. Árboles de Centroamérica: Un Manual Para Extensionistas; Orangutan; CATIE: Turrialba, Costa Rica, 2003. [Google Scholar]
- Silva, C.C.; de Souza, R.C.; da Silva Ferraz, J.B. Estudo da diferenciação das espécies de andiroba: Carapa guianensis e Carapa procera através do método quimiométrico. Rev. Contemp. 2023, 3, 2202–2227. [Google Scholar] [CrossRef]
- Bacca, P.P.; Zuluaga, J.J.; Ivez Perez, J.; Palacio, M.R. Evaluation of pre-germination treatments in Carapa guianensis AUBL. Rev. Cienc. Agrícolas 2020, 37, 56–66. [Google Scholar] [CrossRef]
- Scarano, F.R.; Pereira, T.S.; Rôças, G. Seed germination during floatation and seedling growth of Carapa guianensis, a tree from flood-prone forests of the Amazon. Plant Ecol. 2003, 168, 291–296. [Google Scholar] [CrossRef]
- Vianna, N.G. Conservação de sementes de andiroba (Carapa guianensis Aubl.). EMBRAPA-CPATU. Circular Técnica 1982, 34, 1–10. [Google Scholar]
- Román, F.; De Liones, R.; Sautu, A.; Deago, J.; Hall, J.S. Guía para la Propagación de 120 Especies de Árboles Nativos de Panamá y el Neotrópico; Environmental Leadership and Training Initiative—ELTI, Yale School of Forestry & Environmental Studies: New Haven, CT, USA, 2012; ISBN 978-9962-05-347-7. [Google Scholar]
- Wu, Y.; Sun, X.-R.; Pritchard, H.W.; Shen, Y.-B.; Wu, X.-Q.; Peng, C.-Y. The metagenomics of soil bacteria and fungi and the release of mechanical dormancy in hard seeds. Front. Plant Sci. 2023, 14, 1187614. [Google Scholar] [CrossRef]
- Stoian-Dod, R.L.; Dan, C.; Morar, I.M.; Sestras, A.F.; Truta, A.M.; Roman, G.; Sestras, R.E. Seed germination within genus Rosa: The complexity of the process and influencing factors. Horticulturae 2023, 9, 914. [Google Scholar] [CrossRef]
- Jarrar, H.; El-Keblawy, A.; Ghenai, C.; Abhilash, P.C.; Bundela, A.K.; Abideen, Z.; Sheteiwy, M.S. Seed enhancement technologies for sustainable dryland restoration: Coating and scarification. Sci. Total Environ. 2023, 904, 166150. [Google Scholar] [CrossRef] [PubMed]
- Barbaro, L.; Karlanian, M.; Rizzo, P.; Riera, N. Caracterización de diferentes compost para su uso como componente de sustratos. Chil. J. Agric. Anim. Sci. 2019, 35, 126–136. [Google Scholar] [CrossRef]
- Polláková, N.; Šimanský, V.; Jonczak, J. The relationships of texture and hydrophysical properties in soil profiles under selected exotic trees in the context of climate change in Central Europe. J. Ecol. Eng. 2021, 22, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Howard, F.W.; Merida, M.A. El taladrador de las meliaceas, Hypsipyla grandella (Zellar) (Insecta: Lepidoptera: Pyralidae: Phycitinae EENY-337/IN614; Featured Creatures, Entomology and Nematology Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 2004; pp. 1–10. Available online: https://edis.ifas.ufl.edu/publication/IN614 (accessed on 18 August 2023).
- Hilje, L. In Search of a Preventive Approach to Management of the Meliaceous Borer (Hypsipyla grandella). Rev. Cienc. Ambient. 2020, 54, 211–229. [Google Scholar] [CrossRef]
- Villacorta Barbarán, M.A. Ensayo de Propagación y Crecimiento Inicial de Carapa guianensis, en Vivero, CIEFOR-Puerto Almendras, Loreto, Perú. Bachelor’s Thesis, Universidad Nacional de la Amazonía Peruana, Iquitos, Peru, 2010. Available online: http://repositorio.unapiquitos.edu.pe/handle/20.500.12737/1875 (accessed on 2 April 2023).
- Nautiyal, P.C.; Sivasubramaniam, K.; Dadlani, M. Seed dormancy and regulation of germination. In Seed Science and Technology; Springer: Berlin/Heidelberg, Germany, 2023; pp. 39–66. [Google Scholar]
- Chenge, I.B. Height–diameter relationship of trees in Omo strict nature forest reserve, Nigeria. Trees For. People 2021, 3, 100051. [Google Scholar] [CrossRef]
- Quoreshi, A.M.; Piché, Y.; Khasa, D.P. Field performance of conifer and hardwood species 5 years after nursery inoculation in the Canadian Prairie Provinces. New For. 2008, 35, 235–253. [Google Scholar] [CrossRef]
- Armas, M.L.T.; Paredes, L.U.N. Aprovechamiento de plantaciones forestales en Imbabura, Ecuador. Rev. Amaz. Cienc. Tecnol. 2019, 8, 98–106. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fruits | Origin of the Fruits | Number of Fruits | Number of Seeds | Utilization |
|---|---|---|---|---|
| Mature | Harvested from the tree | 23 | 324 | Main test |
| Immature | Process of maturing in water | 8 | 89 | Preliminary |
| Process of maturation in forest soil | 8 | 94 | Germination test | |
| Total | 39 | 507 | ||
| Code | Category | Characteristics |
|---|---|---|
| 0 | Bad | Dead plants, dry |
| 1 | Regular | Plants with ≤50% green leaves |
| 2 | Good | Plants with ≥50% green leaves |
| 3 | Excellent | Plants with 100% green leaves |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villota-González, F.H.; Palacios, W.; Villota González, E.J.; Bernaola-Paucar, R.M.; Sulbarán-Rangel, B. Seed Propagation of Carapa amorphocarpa W. Palacios Using Various Treatments of Substrates and Mechanical Seed Scarification in a Nursery in the Andean Area of Northwestern Ecuador. Forests 2024, 15, 987. https://doi.org/10.3390/f15060987
Villota-González FH, Palacios W, Villota González EJ, Bernaola-Paucar RM, Sulbarán-Rangel B. Seed Propagation of Carapa amorphocarpa W. Palacios Using Various Treatments of Substrates and Mechanical Seed Scarification in a Nursery in the Andean Area of Northwestern Ecuador. Forests. 2024; 15(6):987. https://doi.org/10.3390/f15060987
Chicago/Turabian StyleVillota-González, Freddy Hernán, Walter Palacios, Edison Javier Villota González, Rosario Marilú Bernaola-Paucar, and Belkis Sulbarán-Rangel. 2024. "Seed Propagation of Carapa amorphocarpa W. Palacios Using Various Treatments of Substrates and Mechanical Seed Scarification in a Nursery in the Andean Area of Northwestern Ecuador" Forests 15, no. 6: 987. https://doi.org/10.3390/f15060987
APA StyleVillota-González, F. H., Palacios, W., Villota González, E. J., Bernaola-Paucar, R. M., & Sulbarán-Rangel, B. (2024). Seed Propagation of Carapa amorphocarpa W. Palacios Using Various Treatments of Substrates and Mechanical Seed Scarification in a Nursery in the Andean Area of Northwestern Ecuador. Forests, 15(6), 987. https://doi.org/10.3390/f15060987