Abstract
The silver fir (Abies alba Mill.) is among the most valuable conifers in Europe for ecological and economic reasons. Throughout the course of history, primarily in the 20th century, its share in stands has been declining due to ill-suited management practices, especially clear-cut management, air pollution (SO2 and NOX emissions), and wildlife-induced damage. This literature review compiles findings from 338 scientific papers. It describes futures for silver fir and its distribution, ecological requirements, threats and diseases, seed production and nurseries, and forest management practices with emphasis on ongoing climate change. Based on recent knowledge of fir ecology and population dynamics, small-scale shelterwood and selection management have been introduced in fir stands, which have also stabilized them. Fir is an essential species for maintaining high stability and biodiversity, especially on planosols and in waterlogged habitats. Owing to its shade tolerance and environmental plasticity, it can coexist very well with many tree species in mixtures, which can increase the productive potential of stands within the natural range in Europe. The average stand volume of mature fir stands ranges from 237–657 m3 ha−1. For its successful natural regeneration, it is essential to reduce cloven-hoofed game and thus prevent bud browsing damage. The attractiveness of fir in terms of heavy browsing is the highest of all conifers (52% damage). On the other hand, fir is a species relatively resistant to bark stripping and the spread of secondary rot compared with Norway spruce (Picea abies [L.] Karst.). Under global climate change, fir is expected to shift to higher elevations with sufficient precipitation, while in the southern part of its natural range or at lower elevations, outside water-influenced habitats, it is likely to decline. Climate change is intricately linked to the heightened prevalence of forest pathogens with significant damage potential in Europe, necessitating careful consideration and strategic adaptation within management practices of fir forests.
1. Introduction
From ecological and economic perspectives, silver fir is one of the most significant coniferous tree species in Europe [1,2,3,4,5,6], as well as in Czechia [7,8,9]. Originally, fir was the most abundant coniferous tree species in Czechia; its share of an area in the natural species composition is reported to be 19.8% [10]. According to Málek and Novák and Dušek [11,12], significant changes are evident in the representation of fir throughout history—in the year 1200: 20%, 1600: 30%, 1800: 23%, and 1900: 10% of the total area of Czechia. The increase in fir representation, up to 30%, was likely supported by more widespread grazing under deciduous trees and raking of their litter, thus improving the conditions for the germination of fir seeds [13,14,15,16]. On the other hand, the proportion of fir in Czechia was only 3% in 1950, 2.1% in 1970, and it declined to 0.9% in 1998. Currently, fir grows on an area of 32,272 ha, i.e., 1.2% of the forest area of Czechia [10], and its share in forest regeneration is gradually increasing (approximately 1500 ha annually; [10]). A similar trend is also visible in other European countries, for example, Germany or Italy [17,18].
Fir is a climax species [19] that cannot thrive at lower elevations (below ~500 m a.s.l.) and in warm regions (especially in the Mediterranean—Spain and Italy), as it is limited by lower precipitation [20], but it can be compensated by higher soil water content and sufficient air humidity [21]. However, fir does not grow on permanently (only seasonally or temporarily) waterlogged sites [22]. In Northern Europe and in mountainous areas, silver fir is limited by low temperatures and late frosts that extend into the early part of the growing season [23,24].
As an indicator of various types of atmospheric contamination [25], the distribution of fir was strongly affected by the air pollution crisis (especially high concentration of SO2) in the second half of the 20th century [7,26,27,28]. The synergism of air pollution with the occurrence of silver fir woolly aphid (Dreyfusia normannianae), balsam woolly aphid (Dreyfusia piceae), and poor management practices (inappropriate silviculture practices, clear-cut systems, and insufficient pest control) have also been reported [4,5,8,29]. The convergence of pollution and pest outbreaks, compounded by lower tree vitality, underscores a critical nexus driving fir forest degradation. This interplay not only amplifies the susceptibility of weakened trees to pests, but also amplifies the overall decline in forest resilience [21,30].
Decreases in silver fir stand densities are also hastened by game-induced damage through bud browsing, bark stripping, browsing, and fraying [7,31,32,33,34,35]. These activities not only stunt the growth of silver fir regeneration, which is very palatable for deer and other wildlife, but also leave it vulnerable to diseases and environmental stresses [36,37]. Without proactive conservation efforts to address game damage and active silviculture practices, the future of silver fir populations remains uncertain [38,39].
Current fir dieback is mainly attributed to climate change [5,8,28,40,41]. In particular, warm summers and recurrent drought have a significant negative impact on the health of silver fir [2,21,41,42,43].
The main thesis of this literature review of 338 studies aims to assess the role, opportunities, and risks, with a focus on supporting the planting and silviculture of silver fir in European forestry, when Norway spruce is in decline due to climate change. The secondary objectives focus on a detailed review of (i) species description and distribution, (ii) ecological requirements, (iii) threats and diseases, (iv) habitat and stand conditions, (v) seed production and nursery management, and (vi) close-to-nature forest management with an emphasis on the ongoing climate change.
2. Silver Fir Features and Its Distribution
Silver fir has a sturdy, cylindrical trunk and a conical to cylindrical crown, often becoming a flattened “stork’s nest” in mature trees (Figure 1). It typically grows 30–60 m tall with a trunk diameter of 1–2 m. Its root system, either tapering or heart-shaped, has deep lateral roots and ensures the mechanical stability of the stand. The bark, initially smooth and whitish–grey, darkens and splits with age. Fir wood is yellowish–white with clear annual rings and lacks resin ducts. Its needles, 18–30 mm long and 2 mm wide, are dark-green and glossy on top with pale stomatal bands below. The male strobili measure 2 cm long and female strobili are 2.5–4.5 cm long, appearing on the upper branches. The cones, which are 10–18 cm long, turn from greenish–blue to brown as they mature. The fir flowers in April–May and the cones ripen by September of the same year, with a lifespan of 300–600 years [3,19,44,45,46,47].
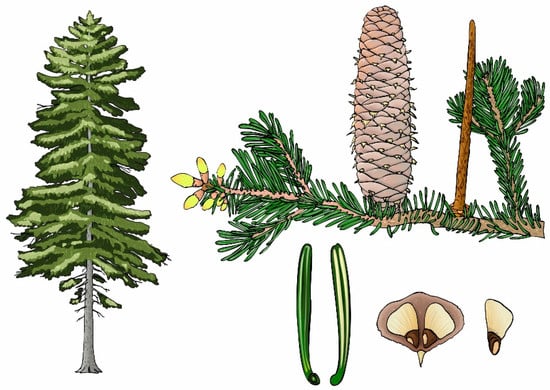
Figure 1.
Tree habitus, branch with cones, needle, and scales with the seed of silver fir (Abies alba Mill.; author: Josef Macek).
Silver fir grows primarily in Central and Southern Europe [48,49]. Its distribution range is relatively small, patchy, and limited to the mountainous regions of Europe, but its potential range is significantly larger (Figure 2). In comparison, the distribution area of Norway spruce or Scots pine is considerably more extensive [50,51]. In the south, fir grows from the Pyrenees through Corsica, Italy, and the Dinaric Mountains, representing the southern edge of silver fir’s natural distribution. Its southernmost limit is in the Southern Apennine Peninsula in Calabria, while the westernmost range closes up in the Eastern Pyrenees, where it also forms the upper boundary of the forest. Further west, there is a small, isolated population in the Normandy hills of northwest France and another in central France. A more continuous distribution begins in the western foothills of the Alps in eastern France and the Jura, Vosges, and German Black Forest. The northern boundary of the fir traverses the Weser Uplands in northwestern Germany, the Thuringian Forest, the foothills of the Ore Mountains (Krušné hory), and the Giant Mountains (Krkonoše), and then the Malopolska and Lublin Uplands in Poland. It reaches its northern limit near Warsaw and in the Białowieża Forest. The eastern boundary continues into the Eastern and Southern Carpathians. Inside the Alpine system and in the Tatra Mountains, it grows in small, scattered patches. However, silver fir also occurs in the lowlands, for example, in France, Poland, and Ukraine [2,3,19,26,45,47,52,53]. It is the leading co-dominant tree, primarily in forest altitudinal zones (FAZ) 4–6 (400–1200 m a.s.l.; [54]). In these typical habitats, silver fir (share 10%–20%) forms mixed forests together with European beech (Fagus sylvatica L.; 20%–40%) and Norway spruce (Picea abies [L.] Karst.; 50%–80%), characterized by a complex internal structure, the so-called Hercynian mixtures [55,56,57]. Montane forests, composed of these three tree species, cover a total area of more than 10 million hectares in Europe [58].
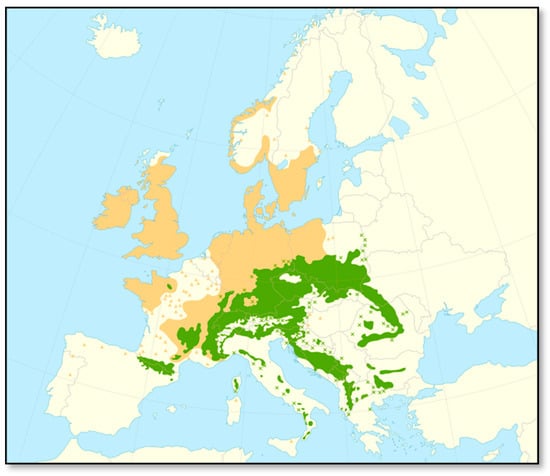
Figure 2.
Map showing the native range (green color) and introduced/naturalized range (orange color) of silver fir (Abies alba Mill.) in Europe [59]. Green crosses represent native isolated populations and orange triangles are introduced/naturalized isolated populations.
3. Ecological Requirements and Productivity
Abies alba grows optimally under a predominantly oceanic temperate cool and humid climate with mild winters, ideally resembling the continental climate in Poland. In Europe, it grows from 135 to 2900 m above sea level [44]. Fir–beech forests in the central part of its distribution range are considered the optimal habitats for silver fir, i.e., to the south from Czechia, at altitudes of about 800–1200 m a.s.l., with precipitation of 1000 mm or more [45]. At lower altitudes, silver fir occurs in cooler and wetter basins and also on alluvial plains at the northern boundary of its range [60]. Severe, dry winters and dry, hot summers are unsuitable for fir. It is sensitive to late frosts, i.e., in spring [19,45,47,61].
Silver fir is a tree species that could benefit from the anticipated climate change, especially in terms of dispersal to higher altitudes with sufficient precipitation, except in areas with severe winters [26,56,62]. However, it has considerable moisture requirements and requires higher air humidity than other species [4,21]. The annual precipitation in its current range varies widely from ~700 to 2500 mm with optimum precipitation of 1500 mm, ideally not falling below 700–800 mm yr−1 [26].
Silver fir is known for its ability to tolerate shade for several decades [63]. Fir undergrowth can grow in heavy shade for as long as 120 years, attaining heights of 1–2 m in Central Europe. Its light requirements are influenced by a complex of other climatic factors (heat, precipitation, humidity, and airflow) and soil factors. The more favorable the habitat conditions, the lower the light requirements of the fir. In contrast, at cooler, higher elevations or on drier and mineral-poor soils, even at the lower limit of its range, the light requirements of fir trees are significantly higher [13,14]. Fir grows primarily on deep, moderately fertile to rich and moist to seasonally or temporarily waterlogged soils. However, it can adapt and grow on stony or peaty soils. In some areas, its optimal habitat is limestone (Western Alps, Jura). This tree species is irreplaceable on heavier loess soils, especially on planosols at mid- and higher altitudes [14]. In mixed forest stands, fir positively influences soil properties because the accumulation of needles stimulates the formation of the humus layer, especially compared with spruce [64]. In mixed stands, silver fir also positively contributes to creating and maintaining a stand environment, especially considering that it can thrive as a component of the lower layer for a long time. Fir in mixed stands balances extremes in temperature, humidity, and airflow limitation [62].
Silver fir is a high biomass-producing tree species (Table 1). For example, at a mean height for the main stand of 30 m, the stock volume reaches 580 m3 ha−1 [65]. The average stock volume of mature fir stands ranges from 237 to 657 m3 ha−1 and the stand basal area from 20.6 to 70.0 m2 ha−1 (Table 1). However, timber processors are not yet able to adequately exploit the high-quality and standard production of fir timber compared with spruce, and therefore, the produced mass is not sufficiently cost-efficient (caused by the low proportion of fir in the species composition). An interesting feature of the change in the timber market during the years of the historically greatest bark beetle outbreaks in Czechia (2016–2020) is the fact that the price of fir timber in roundwood assortments increased significantly and is currently at the same level as that of spruce [66]. Bark beetle salvage logging of coniferous species (especially Norway spruce) increased from 1.5 mil. m3 annually in the period of 2003–2015 to 23 mil. m3 in 2019 [67]. The significant increase in roundwood prices in the last decade (up to 146%) has been the result of a substantial destabilization of the Central European timber market, caused by natural disturbances linked to climate change, a shortage of wood in the context of increased demand, and higher timber harvesting compared with required increments [68]. Furthermore, unaffected mature fir stands at the harvesting age are rarely affected by climate change and salvage logging compared with the vulnerable Norway spruce [21].

Table 1.
Overview of selected available publications related to silver fir (Abies alba Mill.) production parameters in Europe.
Fir is the slowest growing tree species in the first 10–15 years compared with beech and spruce, although it can tolerate a lack of light for a long time. Therefore, it can only succeed in the regeneration process if it has a head start of at least 15–20 years over other tree species that grow more vigorously in their early stages [84,85]. Another prerequisite for its successful growth is sufficiently diverse stands in which fir can maintain a vertically extended crown compared with a short part of the trunk without branches [8,86]. Subsequently, the height increment of fir accelerates until around year 15, peaks at 30–40 (–70) years or much later under unfavorable conditions, and persists for over 100 years. The volume increment peaks at around 55–65 years, i.e., relatively late [45]. As a result of climate change (longer growing season and increase in annual mean temperature), a significant increase in the annual volumetric increment of fir trees, from 7.2 to 11.3 m3 ha−1 y−1, has been recorded in European mountains from 1980–2010 [58].
Fir’s ability to tolerate shading and regenerate under it makes fir suitable for multi-layer, mixed-aged stands and for single and group mixed-selection forests, which are characterized by selective tree harvesting, continuous canopy cover, rich diversity and vertical structure, and sustainable management [4]. In mixed stands, its strengthening function against windthrow and its beneficial effect on soil is highly valued [64,87]. This is also true in the case of the Hercynian mixture, which used to be the most common composition of natural stands at mid and mountain altitudes in Central Europe [88]. In ravines and on scree, fir mixtures evolved, e.g., with maples (Acer spp.), in warmer habitats with European hornbeam (Carpinus betulus L.), and in poorer habitats with Scots pine (Pinus sylvestris L.). Lime (Tilia spp.), sessile oak (Quercus petraea [Matt.] Liebl.), rowan (Sorbus aucuparia L.), or hazel (Corylus avellana L.) occur as subsidiary species. In the Pyrenees, it accompanies mountain pine (Pinus uncinata Ramon ex DC) on the uppermost border in the context of elevation. In other habitats, fir is usually found only as an admixed or interspersed tree species [89]. It maintains its presence here primarily due to its ability to survive in the understory for a long time or its ability to grow on alternately wet and waterlogged soils [8,16,56,90].
In forest stands, fir affects soil mainly by its quality of litter and its relatively low decomposition rate [76,91,92]. Specifically, litter of deciduous species with rapid decomposition rates typically break down within a few months (C/N ratio = 12–25), whereas slower decomposition of litter is observed in conifers, such as silver fir, which may take several years due to their higher C/N ratio (>40) [93]. Spatial structure also influences soil conditions through litter [94]. The amount of litter is higher under the tree canopy than in the openings [95,96]. Differences in the spatial structure of stands greatly influence thermal, light, and moisture conditions, and thus affect the rate of litter decomposition [97,98,99]. Stand gaps are characterized not only by increased light, heat, and precipitation, but also by faster decomposition of organic litter [100,101] and higher nutrient concentrations in the soil solution [102]. In addition, soil heterogeneity is enhanced by the cluster distribution of concentrated roots and, thus, higher water and nutrient consumption in mixed stands [103,104].
Generally, the success of natural regeneration, apart from ground vegetation [105], depends on the properties of the topsoil horizons [106], which is a condition for seed germination, root development, relationships with soil microflora, and the availability of water and nutrients [99]. Compared with spruce, fir is characterized by a lower acidification capacity and a higher C/N ratio [107]. Třeštík and Podrázský [108] reported a lower (54%) accumulation of forest floor humus in fir stands and significantly higher total nitrogen and calcium contents compared with spruce. A higher accumulation of total carbon in the Ah soil horizon was observed beneath spruce than beneath fir [109]. In tree biomass, fir fixes a lower amount of carbon compared with spruce, because fir allocates more carbon to the crown and spruce to the stem [110].
In the natural species composition, fir reaches the highest proportion in the ecological series of gleysols and planosols, up to 70%. The groups of forest habitat types include (Fageto-) Abietum variohumidum mesotrophicum, Abietum piceosum variohumidum acidophilum, Abietum piceosum variohumidum oligotrophicum, Abietum quercino-piceosum paludosum mesotrophicum, and Abietum quercino-piceosum paludosum oligotrophicum [111]. In these habitats, fir forms a full spectrum of mixed stands, with beech, spruce, oaks, sycamore maple (Acer pseudoplatanus L.), black alder (Alnus glutinosa [L.] Gaertn.), silver birch (Betula pendula Roth.), European aspen (Populus tremola L.), and Scots pine [89]. It is often the main species in montane forest phytocenoses, such as silver fir and European beech (Abieti-Fagetum), subcontinental silver fir forests (Galio-Abietion), upland fir forests (Querco-Abietetum), and slope forests of silver fir (Abietetum albae) in Switzerland [112]. It also forms forest phytocenoses with spruce Piceetum subalpinum sphagnetosum [112].
As a long-lived species, fir is considered an important ecological and functional stabilizer of European forests [3]. It stabilizes soil, retains water, and is less susceptible to snow and ice damage than Norway spruce [113,114]. Silver fir is an essential species for maintaining high biodiversity in forest ecosystems due to its tolerance to shade, ability to survive extended periods in the understory and respond when light conditions become more favorable, plasticity to environmental conditions, and ability to coexist with numerous tree species [7,63,115].
Fir is normally the most differentiated tree species in terms of age, height, and diameter, which makes natural forests with a higher proportion of fir trees close to selection forests in their structure [116,117]. With a higher representation of beech at the expense of spruce, the regeneration and growth of fir is more continuous, creating a vertical and multi-layered canopy [118]. A higher spruce share creates a typical horizontal fir and spruce canopy at the optimum stage as a consequence of the fact that the lifespan of trees is longer than the duration of their height growth. The optimum stage, also characterized by stagnation of natural regeneration, usually occupies about 20% of the area of natural forests, and its duration is expressed in the same period over the entire development cycle, i.e., about 80 years according to studies from Slovakia and the Czechia [90,119].
At the main level of the stand of both selection and cultural forests (extensively managed by human activities over time in terms of density, species composition, and structure), there is typically a gradual rotation of the dominant tree species occurring in differently sized areas [116,120]. It is clear that, especially in forest altitudinal zones 5 (600–900 m a.s.l.) and 6 (800–1200 m a.s.l.), there is a gradual rotation of generations of fir, spruce, and beech in their typical stand mix at the main stand level. This is likely due to the shorter lifespan of beech and its requirements for specific light and soil conditions [55,90,121,122,123].
The interspecific substitution of trees in the same stand is also seen in the context of their different light uses [124]. Firs primarily use the short-wavelength blue component of the solar spectrum (400–430 nm; [125]) and are more sensitive to the lack of this component than to the overall reduction in light intensity that occurs in shaded conifer stands [89,116,126]. This fact also explains the better regeneration of firs under spruce than under fir canopy, a historically observed phenomenon [4,61,127].
The competitive abilities of tree species in mixed stands also change depending on the soil properties [128]. Acidic soils reduce the vigor of beech, and calcareous soil reduces the vigor of spruce. With increasing acidity and excessive soil moisture, fir and spruce establish at the beech optimum, while nitrogen-rich soils reduce conifer vigor [55]. According to Ellenberg [129] and Tinner and Lotter [130], fir is more competitive than beech in locations with low temperatures and high summer rainfall. According to historical records, abundant summer precipitation is more crucial for fir than low temperatures [130].
Complex layered stands, where the growth space is fully utilized, exhibit an inverse relationship between the amount and biomass production of the understory (lower story) and the biomass of the main stand (upper stories; [76,131,132,133,134]). Changes in the upper story, combined with canopy disturbance, are quickly reflected in a changed light regime of the understory, and thus in an increase in its biomass. The main cause of overyielding in multi-layered stands is their light-use efficiency [135]. Hence, the reduction in the canopy, whether natural or artificial, affects the structure and vigor of natural regeneration [84]. In forests of the typical Hercynian mixture of forest altitudinal zones 5 and 6, the requirements of fir, spruce, and beech for light, nutrients, and water do not differ substantially, while light intensity plays the most important role in growth [56,136,137]. Tree species differ significantly in their ability to survive in the long term under reduced light conditions while maintaining fully functional and efficient photosynthetic processes [138]. Fir, an extremely shade-tolerant species, has a distinct competitive advantage, especially over spruce, in low light conditions [55,131,137]. This fact was also confirmed by the preserved montane mixed forests of the Romanian Carpathians by Stancioiu and O’Hara [133]. These studied stands are located at an altitude of 800–1300 m a.s.l., and the age of the main stand is 70–350 years. Their results showed that, at low light intensity (percentage of the above-canopy light, PACL < 20%–35%; BA > 30 m2 ha−1), fir and beech clearly outgrow spruce, and the latter can even be eliminated from the regeneration as it develops. Under medium-light conditions (PACL = 35%–70%; BA = 15–35 m2 ha−1), the growth abilities of all three species are equal, while under open conditions (PACL > 80%–90%; BA < 15–20 m2 ha−1), all of the three species show the same development, with spruce tending to outgrow the other two shade-tolerant species [133]. Another factor is the reduction in the height increment of fir under direct sunlight (PACL > 80%–90%) compared with the maximum growth when shaded (PACL = 50%–80%; [133]). The fact that strong interventions into the main stand canopy are more favorable for spruce and beech than for fir has been noted by many authors [38,116,131,138,139,140].
Numerous studies confirmed that mixed stands can have higher biomass productivity than monocultures in suitable habitats [122,141,142,143,144]. Silver fir and Norway spruce also grow faster in mixed stands than in monocultures, and their complementary effect increases with improved growing conditions, i.e., resource availability or climatic conditions [145,146,147]. However, an increase in the complementarity and productivity of these species occurs because of their different light requirements, which also affects the absorption of photosynthetically active compounds by radiation or light-use efficiency [142].
In the last 40–60 years, a significant decline in fir in forest stands and its gradual replacement by spruce or naturally spreading beech have been observed, both in the Hercynian and Carpathian regions [7,148,149,150,151,152,153]. Especially in forest altitudinal zones 5 and 6, the predominance of beech regeneration over fir regeneration has been documented. European beech has profited (acceleration of growth, fructification, and regeneration abundance) more from climate-changed growing conditions compared with other tree species and could have changed the competition relationship in favor of beech [154,155]. In the well-preserved Dobroč Primeval Forest in Slovakia, the percentage of natural regeneration of fir was around 60% in 1935, followed by a significant decline. Korpel [90] reported only a 20% share of fir in the late 1970s and 1980s. This phenomenon has been interpreted as a common substitution of tree species occurring during the development cycle in virgin forests [90,119]. However, the negative influence of game on fir regeneration was evident at the time, and as the regeneration continued to decrease in many areas, this has been assessed as an unnatural and negative development that is unlikely to be reversed naturally within the evolution of the forest stand in Switzerland, Czechia, France, and other countries [7,36,118,156].
4. Threats and Diseases
One of the most striking aspects of the ecology of silver fir is its recurring decline, which has been observed in Europe since the 1500s [127,157,158,159]. The rapid decline of fir in Central Europe has been associated primarily with the intensification of human activity in forests and the development of industry [160,161,162,163,164]. However, data on fir dieback dates from well before the mass industrial expansion of the 20th century [7,8,16,26,116,165,166]. Fir dieback and its reduced natural regeneration have been observed from the 1960s to the 1990s. The decline was first interpreted as a consequence of the natural range of fir reaching its outer boundaries [167], but in the 1970s and 1980s, fir dieback of varying intensities was observed across the entire natural range of the species [158,168].
Owing to their high sensitivity to air pollution [169,170], fir trees have been reported to be in decline as a result of the ensuing stress [8,27,28,61,171,172,173,174]. In particular, SO2 pollution has been a critical factor in the decline of silver fir in the 20th century [173]. The worsening situation was reflected in reduced growth and increased tree mortality [5,173]. There were predictions that silver fir would eventually experience general dieback due to air pollution [175], but in the interim, the SO2 concentrations declined significantly. In Europe, SO2 emissions peaked in the early 1980s and from that point until 1995, decreased by 50% [176,177], which triggered a rapid recovery in fir growth and vigor [27,43,178]. Fir stands regenerate in the most affected areas following a reduction in air pollution [7,173]. According to Bošeľa et al. [27], Bountgen et al. [43], and Mikulenka et al. [8], this is due to the combination of air pollution decrease and an increase in temperature. Bošela et al. [27] stated that the most significant factors that positively affect radial growth in the Western Carpathians are the decreases in the air concentrations of SO2 and NO3 and the increases in temperature in April, June, and July. Although there are differences between the areas in all four regions, a rapid acceleration of the increment in the last two to three decades was observed, reaching values between 150 and 300% compared with previous periods. Similarly, in the forests of the Sudeten Mountains, there has been a significant regeneration of silver fir since the high annual SO2 concentrations (30–50 μg·m−3) subsided [179].
Natural silver fir regeneration has increased its dominance in Pyrenean subalpine forests in recent years, as well as in some mixed forests in Spain [180] and in other European forests [70]. However, some studies suggest a different response of silver fir along the edges of its natural range [181]. A retreat of silver fir from warmer and drier areas has been observed in Slovenia, especially in fragmented forests and at the limits of fir’s distribution range [182]. Its dieback is often attributed to climate change [28,40,41,183,184,185,186]. The negative impact of climate warming has been observed in southwestern Europe [2], chiefly in the Mediterranean region, where the decline in silver fir is related to increased aridity [2,178,187]. In particular, warm summers and recurrent drought have had a significant impact on the health of silver firs [2,41,42,43,188]. The narrow genetic variation of silver fir in Europe may have limited its adaptability to current conditions in terms of climate change [158,189,190,191]. Adaptation to drought is coupled with slow growth, but independent from phenology in marginal silver fir populations [192]. Populations with “start early and grow slowly” strategies have higher water use efficiency [193]. The provenances’ ranking by resistance, recovery, and resilience in radial growth revealed that a number of provenances from Bulgaria, Italy, Romania, and Czech Republic placed in the top ranks. Assisted migration may support the adaptation process and help to conserve and increase genetic diversity, especially at the species distribution edges [194].
Infestation by bark beetles (Pityographus pityographus Ratz.; Pityokteines vorontzovi Jac.; Pityokteines spinidens Reitt.) has been observed in southern France, which could partly explain the high mortality [195,196]. The decline in firs during the period of the ecological disaster (1980s and 1990s) has often been associated with the damage to fir stands by the silver fir woolly aphid (Dreyfusia normannianae) and balsam woolly aphid (Dreyfusia piceae; [197,198].
In the northern Carpathians (Czechia and Slovakia), Slovenia, and Croatia, silver fir has declined due to the spread of beech and, to a lesser extent, Norway spruce, and due to the failed regeneration as a result of the cloven-hoofed game population increase [7,118,148,182,199,200]. The retreat of silver fir from its natural stands, as well as from artificial regeneration, is aggravated in many locations by game-induced damage (especially red deer—Cervus elaphus L., roe deer—Capreolus capreolus L., sika deer—Cervus nippon Temminck, European muflon—Ovis aries musimon Pallas, and fallow deer—Dama dama L.) through bud browsing, bark stripping, browsing, and fraying [7,31,33,34,35,124,201,202]. Table 2 clearly shows the high attractiveness of silver fir in terms of bud browsing damage (52%) across Europe. Higher damage in montane forests was also recorded for sycamore (56%) and rowan (55%), compared with minimal damage in Scots pine (5%) and Norway spruce (17%).
Table 2.
Overview of selected available publications related to the proportion of browsing damage (%) in selected tree species with an emphasis on silver fir (Abies alba Mill.).
One of the most serious threats, not only to fir, but to all conifer saplings, is the removal of bark from the tree trunks by cloven-hoofed animals (browsing and bark stripping). As a result of this damage, not only is the vascular cambium disrupted, but above all, the quality of the timber is compromised by secondary infestation with fungal pathogens and the development of stem rot. This is especially true for Norway spruce, whose value as merchantable timber decreases rapidly due to bark damage and subsequent decay [213,214,215,216,217]. In contrast, there is minimal knowledge of the effects and consequences of game-induced bark removal on silver fir. Bazzigher and Schmid [218] and Kohnle and Kändler [219] only reported that bark damage in silver fir was less detrimental than that in Norway spruce, while also suggesting that fir timber is less susceptible to stem rot. However, neither the reasons for the increased susceptibility of Norway spruce to decay nor the mechanisms of resistance of silver fir to the spread of rot and subsequent decay are currently known. Metzler et al. [220] reported that, unlike silver fir, Norway spruce has resin canals, which may be the reason for the spread of fungal pathogens after bark damage by game. Game-induced bark damage to silver fir was partially studied in terms of histological changes by Oven and Torelli [221,222], and concerning growth and vitality by Pach [223,224,225,226,227], who reported a decrease in timber quality, reduced vitality and growth of fir trees, and the spread of rot. This rot can manifest itself in timber discoloration, a more advanced level of its development, and even the decomposition of the wood mass (so-called soft rot), which very negatively affects the mechanical stability of the affected stands. Timber discoloration, as the initial stage of rot, can be the result of fungi, bacteria, and the reaction of wood to pathogens [214,228,229,230].
Thus, bark damage is less harmful in silver fir than in Norway spruce, and its timber is also less susceptible to stem rot [219]. Trees damaged by browsing and bark stripping are generally more vulnerable to a lack of precipitation, while healthy trees are more responsive to temperature [231], which may play a significant role in terms of climate change. Healthy trees can increase its production potential in cooler and higher-elevation areas due to the increasing average air temperature, while damaged trees will have a higher risk of mortality due to drought in lowlands. Various measures are taken in an attempt to prevent damage to forest stands by wildlife, notably methods of individual or group protection and the use of commercial repellents [232,233,234,235]. However, these short-term measures do not address the long-term problem of continuous increases in population density as well as the distribution range of wild ungulates [236,237]. These changes in the population dynamics of ungulates are driven by many factors, including changing climatic conditions [238,239].
Silver fir decay results from the spread of rot through injured bark. Heterobasidion abietinum infects primarily silver fir and causes stem rot, particularly in Southern Europe [240,241,242], where firs are often affected by drought during summer. However, in Central Europe, this fungus does not cause any tangible problems in fir, likely due to the more favorable climatic conditions or to the low abundance of this fungus [241,243]. Phellinus hartigii occurs on silver firs with injured bark in Central Europe ([244,245]; Figure 3). Widespread deterioration in the health and dieback of the genus Abies Mill. due to other pathogens have been observed in Europe in recent years. Increased forest degradation of silver fir forests, presumably due to climate change, was found, for example, due to infection of the phytopathogenic bacterium Lelliottia nimipressuralis in Ukraine [246] or the fungi responsible for Herpotrichia needle browning, including Nematostoma parasiticum [247]. Fir stands can also be infested by the hemiparasitic mistletoe (Viscum album L.), which attacks a wide range of woody plants [248,249,250]. This parasite and the subsequent invasion of microorganisms—such as fungi or bacteria—cause mechanical and aesthetic damage to the timber, reducing its increment and commercial value [251,252,253,254].

Figure 3.
Intensive, repeated bud browsing by game (left), earlier damage by browsing and bark stripping (middle), and subsequent infestation by Phellinus hartigii (right) (Photo: Stanislav Vacek).
Natural and anthropogenic disturbances play a principal role in the dynamics of forest ecosystems and influence stand structure and regeneration processes. In Europe, wind and fire are the most severe abiotic disturbances [2,255], and insect infestation is the primary driver of biotic disturbance [123]. Silver fir is relatively resistant to wind, snow, and ice storms compared with other dominant tree species, such as Norway spruce [4,194]. Disturbance regimes in forests dominated by silver fir are characterized predominantly by small-scale events, such as a single tree falling, while large-scale events, such as windstorms or forest fires, are rare [256]. Large-scale events have been increasing, especially in natural silver fir sites in recent years. Annual disturbance-based tree mortality of silver fir has been accelerating and was the highest (compared to Norway spruce, Douglas-fir, European beech, and oak) in Germany during the period of 1929–2014 [257]. Extensive mortality of fir trees was observed after the drought in 2018 in Czechia [258].
5. Impact of Climate Change
Ongoing climate change is currently exhibiting widespread impacts on forest ecosystems and thus on all forest management, which should adapt to changing conditions. Existing climate models predict an increase in average air temperatures during the 21st century, as well as an increase in the frequency of extreme weather events, such as storms, floods, heat waves, and dry periods [123,194,259,260]. Appropriate forest management measures include abandoning monocultures, establishing mixed stands, and promoting the cultivation of natural regeneration [261,262]. A large number of studies report positive relationships between species diversity and forest productivity [122,263], and it has also been confirmed that diversity in forest species composition increases resistance to biotic insect pest calamities [261,264]. Therefore, declining forest stands in Central Europe are often replaced by mixtures of tree species in an attempt to adapt forests to climate change [255,262,265,266]. Much of the Central European forest landscape is still dominated by Norway spruce, despite its documented susceptibility to drought, wind, bark beetles, etc. Numerous studies from across Central Europe confirm that the area suitable for growing Norway spruce will continue to decrease with ongoing climate change [40], even at higher altitudes [267]. Compared with Norway spruce, silver fir and European beech are less susceptible to short summer droughts [265,268,269].
Fir is not considered a dominant tree species in most European countries at present, nor will it be in the future in relation to ongoing climate change, but only an interspersed or admixed species in mixed stands. The range of silver fir, together with Norway spruce and European beech, shows a decline under climate change scenarios. Conversely, sweet chestnut (Castanea sativa Mill.), wild service tree (Sorbus torminalis Crantz), and European white elm (Ulmus laevis Pall.) expand their range potential, and along with European hornbeam, they hold the potential to contribute significantly to the sustainable adaptation of European forests [270]. In nutrient-rich and gleyed habitats at mid- and higher altitudes, a slightly higher proportion of fir is expected due to its ameliorative and stabilizing properties [271]. However, in stands with a high proportion of fir, it is necessary to maintain its share through natural regeneration [8]. Therefore, adaptation measures (monitoring and management, silviculture practices, genetic selection, etc.), taking into account the variability in climatic conditions, must be adopted to mitigate the negative impacts of climate fluctuations [189,272,273,274,275,276].
Living organisms can choose between two strategies for survival in a rapidly changing environment: migration to more suitable habitats or local persistence through adaptation [277]. The third option is extinction. These potential strategies apply to trees in forest ecosystems that have limited migratory capabilities, although significant climate change makes them very relevant [278]. Tree populations begin to migrate when existing habitat conditions are unsuitable for survival [279]. Our knowledge of tree migration rates under global climate change is still limited. However, the differences between the observed migration rates and tree habitat displacement rates under ongoing climate change are considerable [280]. Among others, Feurdean et al. [281,282] estimated the maximum migration rate for the early successional stages of pioneer trees, such as birch, willow, or Scots pine, to be 225–540 m yr−1, while for climax trees, such as silver fir and European beech, it ranges between 115 and 385 m yr−1 depending on the mode of seed dispersal by wind or animals. Numerous climate change scenarios predict a horizontal spread of up to several kilometers and a vertical spread of up to tens of meters per year [283]. For these reasons, tree migration is significantly slower than the rate of forest habitat change [284]. Moreover, the significant forest fragmentation of much of Europe has substantially reduced the rate of tree migration [285].
6. Seed Production and Nursery Management in the Context of Climate Change
Climate change is altering worldwide forests by influencing tree mortality, yet regeneration is pivotal in shaping the forthcoming tree generation and will dictate the configuration and makeup of future stands. Regeneration hinges not only on average seed production, but also on the yearly fluctuations and synchronization among plants in seed production, a phenomenon recognized as mast seeding [286]. In the case of fir, fertility occurs at 25–35 years on isolated trees and at 60–70 years in a dense forest stand [287,288]. Depending on latitude and altitude, fir trees flower from May to mid-July. Seed years occur at two-year intervals at lower altitudes, while at higher altitudes, they occur every 3–5 years [289,290]. The differences in fir fertility of cones in Romania reached up to 11.2 times when comparing good mass years to poor years [291]. The mean dispersal distances of seeds from trees ranged from 9.9 m in pure fir stands to 21.5 m in mixed forest in Spain [292].
The cones mature from September to October. They grow on the upper third of the crown, from where, after disintegration, the seeds are released by May of the following year [293]. Therefore, the best time to harvest cones is in September, ideally just before they fully ripen, while the cones are still intact [294,295,296,297]. At this time, the cones contain 30%–70% water [294,298] and the seeds contain about 40% water [299]. The harvesting period depends on the altitude and the amount of precipitation in a given year. The cones can be harvested as early as mid-August and at higher altitudes as late as October [289,300]. The harvesting period is closely related to the cone-to-seed ratio and the germination capacity of the seed. The later the harvest, the higher the cone-to-seed ratio and germination. A recurrent level of 14% has been reported for fir cones [301,302]. Fir seeds should be collected at morphological (hard) maturity, which precedes physiological maturity, when the seeds can germinate [298]. Fir cones are usually collected manually by picking from standing or felled trees [303] or from seed orchards established for this purpose [291]. Using Czechia as an example, the expanding production of silver fir seed material points to rising demand for fir seed material in the context of an increased need for seedlings for the reforestation of large clearings following the 2017–2019 bark beetle calamity ([67]; Table 3).
Table 3.
Production of seed material, estimated weight of seed (in kg) (cone-to-seed ratio of 10%), and average seedling yield according to the standard of the silver fir quantity in Czechia in 2010–2022 [10].
In Czechia, the share of artificial regeneration converted to hectares ranged from 79.8 to 83.6% for 2010–2022. The seedling numbers in Table 3 show that fir’s reproductive material can be used for the regeneration of approximately 5000 ha per year according to the minimum numbers during afforestation determined by law. The annual artificial regeneration of fir in Czechia ranges from 872 to 1635 ha per year. These figures show a substantial overproduction and the supply of other EU member states with silver fir reproductive material. The seed productivity of stands under ongoing global climate change is highly variable across various locations in Czechia. The need for collecting high-quality reproduction material is constantly increasing [10]. In Central Europe, inter-annual variations in the population-level seed production of fir increased during the past half-century. However, the population-level temporal autocorrelation in fir seed production became more negative during the last decades in the context of climate change [304]. Seed quality is influenced by the forest altitudinal zone, which reflects the amount of precipitation and air temperature during the growing season. The highest seed quality in Czechia was recorded in forest altitudinal zones 2 and 3 (200–500 m a.s.l.), where water-affected habitats suitable for fir are often found [305]. The germination of fir seeds, which depends on provenance and pre-sowing preparation [306], is generally low, around 40% [307,308], and on the southern border of the distribution range, it is even lower, reaching only ca. 28% [309]. Provenances from Calabria in Italy, for example, performed very well and should be the first choice of origin for future planting because of the deleterious effects of climate change in Britain [310]. The Carpathian provenances from Rogów were evaluated according to the index of cultivation and breeding as very good in Poland [311]. Fir exhibited greater resistance to heat stress for the provenance cultivated in the warmer sites [41].
Bezděčková and Řezníčková [306] found that fir seeds stratified for 3 to 4 weeks at 3 °C germinated better at an alternating temperature of 20/30 °C than at a constant 20 °C. The long-term storage of fir seeds is accomplished by gradually reducing the water content in the seeds to 9%–11% and storing them at −8 to −15 °C in cooling boxes [302]. Seedlings are generally grown on substrates sowings with a specific pH (4.2–6.0), favoring soil-based substrates for bare-root seedlings and peat-based substrates for containerized ones [292,293,312]. Inoculation with ectomycorrhizal fungi, such as Lactarius spp., improves seedling quality and survival [294,295,296]. Sowing on mineral soils is usually performed in autumn to benefit from natural seed stratification, although artificial stratification in the spring at 3–5 °C for 21 days is essential to avoid spring frosts [281,282,297]. Fir seedlings need constant moisture, adequate shading, and are susceptible to frost damage, but grow to 5 cm in the first year and 8–25 cm in the second year [280,298,299].
In nurseries, bare-root seedlings are usually grown in shaded conditions for two to three years to promote growth and vigor [299,300]. Seedlings are usually collected manually or mechanically in the spring, with precautions to minimize root hair damage [301,302]. Root system reductions are performed to prevent growth deformities after replanting [302].
It is essential to conduct health inspections related to pests prior to transplanting, especially for seedlings that have been stored [303,304,305]. After collection, roots should be immediately protected and possibly treated with anti-desiccants to improve seedling survival [303,307]. The standard growing period for bare-root seedlings is up to seven years, with innovative methods using air cushions to expedite growth [308,309]. Proper handling and protection from weather conditions from nursery to planting are critical to prevent damage and ensure the survival of transplanted seedlings [310,311,313].
7. Close-to-Nature Silvicultural Systems and Climate Change
The basic principle of close-to-nature forestry, defined as the integration of different silvicultural systems, such as selection, irregular shelterwood, and free-style silviculture, each adapted to the specific conditions and goals of a particular site, is the continuous and gradual improvement of forest stands and sites, promoting a holistic view of forest ecosystems [314]. The study by Čater et al. [315] deepened our understanding of optimizing silvicultural systems for light conditions, essential for the growth and regeneration of tree species, such as silver fir and European beech. It found significant variations in the assimilation rates of beech and fir under varying light intensities in the Carpathian and Dinaric Mountains, indicating that each tree species exhibits distinct growth responses to available light [315,316].
Recent research by Adamič et al. [317] underscores the varying responses of silver fir to climate factors across different regions of the Carpathian Massive. This study revealed that the regeneration efforts may significantly depend on local climatic conditions, particularly precipitation and air temperature patterns. These findings suggest that site-specific climate considerations are crucial in planning silvicultural practices for fir, especially under the impacts of climate change [317]. In addition to the climate, the most crucial factors for fir regeneration, its survival for advanced growth, and further development are suitable habitat, light, soil, and air humidity. For the light requirements of fir, Jaworski [313] stated that seedlings emerge at light intensities of 1%–5% of maximum light. They require at least 5% of maximum light to survive until the following year. Up to the age of 15, the optimum conditions for fir are 15%–25% of maximum light. In the shade, growth is suspended, and saplings tend to form a flat crown [318]. Its emergence in the third year of life is an indicator of adequate light conditions of regenerating stands. For the next 10 to 20 years, the height growth of fir remains relatively low. When a height of about 50 to 80 cm is reached, illumination should be increased by the slow, gradual opening of the parent stand, initiating the height increment and following close-to-nature forestry principles. The transition to full release must be slow and smooth [319,320]. The critical consequences of changes in the growth rhythm of trees in Saxony were addressed by Meyer [127], who searched for the causes of fir dieback at the northern limit of its distribution. Informed by numerous analyses of fir trunks with normal crowns and growth dependencies based on Backman’s function, Meyer derived the course of a “normal development line” [321]. He did the same for diseased and poorly growing fir trees. From the developmental line of healthy and diseased trees, he concluded that the development of all healthy trees was slower in their early years than that of diseased trees with deformed crowns. This implies that fir trees in the studied same-aged stands did not experience a period of suppressed growth in their early years. Too rapidly releasing growth and even stimulating seedling growth in the nursery alters the growth course of fir trees in the early years of development [56,322]. This presumably sets a specific growth rhythm for fir trees, whereby, as a shade tree, they respond to altered conditions. It is possible that, in this way, a climax tree can become a pioneer tree, i.e., with an altered growth rhythm in its early years and early maturation, but also with premature senescence and a lower final wood mass. An abrupt change in the light regime can cause reduced resistance, increased susceptibility to diseases, and a likely change in hereditary characteristics [321].
Fir can tolerate long periods of suppression without weakening their vital energy under natural conditions. However, firs that evolved in clear-cut forests for several generations did not experience long-term shelter from the parent stand, i.e., they changed their nature and required increased illumination from the beginning of their growth. If they are suddenly exposed to the shade of the shelterwood for a longer period, they fail to adapt and become critically weakened [116]. Annual ring analyses indicate that withering firs with dried-out crowns and a more pronounced decline in recent increments generally showed significantly higher diameter increments in their youth than relatively healthier firs with less weathered crowns in the same stand [323]. This implies a greater threat to fir trees that were released more rapidly in the seedling or sapling stages than those that were gradually thinned and experienced slower development.
Fir’s requirements for the other component of radiation—heat—must be considered in close connection with light. From this point of view, fir is a relatively demanding tree species, especially when compared with spruce [116]. Because fir assimilates well in the shade, it also requires a reasonable moisture regime, as it transpires more and has a higher demand for CO2 and water. Higher transpiration is predominantly characteristic of self-seeding firs. They transpire more than older trees [324,325]. Therefore, their regeneration and quality growth depend not only on light, but also on the soil and air humidity, and—above all—sufficient precipitation during the growing season of at least 350 to 400 mm [319]. Firs are susceptible to short-term droughts, severe winters, late frosts, and air currents [326,327]. Therefore, fir can be considered one of the most sensitive and demanding coniferous tree species because, in addition to the already mentioned requirements for light, moisture, and heat, it needs deep, nutrient-rich, loose soils with sufficient water in the upper soil layers for successful growth, preferably around springs in flat terrain, on slopes with water-holding soil, and generally on sites influenced by water [328,329].
Regarding silvicultural practices, fir is not suitable for the clear-cutting management method with artificial regeneration (due to specific light requirements, soil condition preferences, limited regeneration capacity, and competition with other tree species), which was widespread in Central Europe in the past [86,118,330,331,332]. Contrarily, shelterwood, selection, or even border (group) management methods are suitable [4,7,205,333] or as underplanting under pioneer tree species in the restoration of salvage clearings [334]. The recommended management of silver fir forests involves either single-tree selection systems or an irregular shelterwood method, with a regeneration phase lasting between 20 and 40 years [335,336].
From the perspective of future forest management principles, it is therefore essential to know the forest types that will be most suitable in terms of their species, age, and spatial composition under future climatic conditions [123,231,337]. Determining this is particularly challenging given the increasing pressure of rapid climate change [338].
8. Conclusions
Silver fir has the potential to play a crucial role in the future composition of Central European forests, particularly due to its relative resistance to abiotic and biotic factors. Its adaptability and shade-tolerant nature enable it to fulfill both productive and non-productive functions in mountainous forests and water-influenced habitats at lower altitudes. Adaptive management, which supports the establishment of mixed stands with silver fir, is essential for enhancing the stability and biodiversity of these ecosystems. The use of small-scale clear-cutting and selection-cutting methods that mimic natural processes is key to promoting structural differentiation within forest stands. Additionally, addressing high game densities is vital to ensure successful natural regeneration. Research indicates that silver fir is less competitive in areas undergoing rapid and extensive changes, thus recommending selective cutting or irregular shelterwood systems for better adaptation.
Given the anticipated climate changes, including extreme weather events such as storms, droughts, frosts, and increasing average temperatures, the distribution of silver fir is expected to shift to higher elevations and northwards. It is essential to actively manage the genetic pool of silver fir, considering the transfer of highly variable and vital populations from regions like the Carpathians to Central Europe. This genetic management is crucial for mitigating the negative effects of climate change and ensuring the species’ survival.
The conservation of silver fir requires a holistic approach that integrates active forestry and wildlife management. This includes implementing strategies to cope with climate change, such as increasing species and structural diversity, maintaining and enhancing genetic variation within tree species, and increasing the resistance of individual trees to biotic and abiotic stress. Shorter cutting cycles and a lower intensity per cut, focusing on silver fir trees of various sizes and ages, are suggested under warmer climate conditions.
The successful conservation and regeneration of silver fir will depend on a combination of adaptive silvicultural practices, genetic management, and effective wildlife control measures. By adopting these strategies, forest managers can enhance the resilience of forest ecosystems, ensuring that silver fir continues to thrive in the face of ongoing environmental changes.
Funding
The paper was funded by the Technology Agency of the Czech Republic (Project No. TQ03000107), the National Agency of Agricultural Research of the Czech Republic (Project No. QL24010275), and institutional support from MZE-RO0123.
Acknowledgments
The authors thank Jitka Šišáková, an expert in the field, and Richard Lee Manore, a native speaker, for checking the English. The authors’ also thank Josef Macek for his graphical works.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Prpic, B. Obicna jela (Abies alba Mill.) u Hrvatskoj; Akademija sumarskih znanosti: Zagreb, Croatia, 2001. [Google Scholar]
- Gazol, A.; Camarero, J.J.; Gutiérrez, E.; Ionel, P.; Andreu-Hayles, L.; Motta, R.; Nola, P.; Ribas, M.; Sangüesa-Barreda, G.; Urbinati, C.; et al. Distinct effects of climate warming on populations of silver fir (Abies alba) across Europe. J. Biogeogr. 2015, 42, 1150–1162. [Google Scholar] [CrossRef]
- Mauri, A.; de Rigo, D.; Caudullo, G. Abies alba in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; p. E01493b+. [Google Scholar]
- Dobrowolska, D.; Bončina, A.; Klumpp, R. Ecology and silviculture of silver fir (Abies alba Mill.): A review. J. For. Res. 2017, 22, 326–335. [Google Scholar] [CrossRef]
- Bošeľa, M.; Lukac, M.; Castagneri, D.; Sedmák, R.; Biber, P.; Carrer, M.; Konôpka, B.; Nola, P.; Nagel, T.; Ionel, P.; et al. Contrasting effects of environmental change on the radial growth of co-occurring beech and fir trees across Europe. Sci. Total Environ. 2018, 615, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Dinca, L.; Marin, M.; Vlad, R.; Murariu, G.; Drasovean, R.; Cretu, R.; Georgescu, L.; Voichița, T.-G. Which are the best site and stand conditions for silver fir (Abies alba Mill.) located in the Carpathian Mountains? Diversity 2022, 14, 547. [Google Scholar] [CrossRef]
- Vacek, S.; Vacek, Z.; Bulušek, D.; Bílek, L.; Schwarz, O.; Simon, J.; Štícha, V. The role of shelterwood cutting and protection against game browsing for the regeneration of silver fir. Aust. J. For. Sci. 2015, 132, 81–102. [Google Scholar]
- Mikulenka, P.; Prokůpková, A.; Vacek, Z.; Vacek, S.; Bulušek, D.; Simon, J.; Šimůnek, V.; Hájek, V. Effect of climate and air pollution on radial growth of mixed forests: Abies alba Mill. vs. Picea abies (L.) Karst. Cent. Eur. For. J. 2020, 66, 23–36. [Google Scholar]
- Šimůnek, V.; Prokůpková, A.; Vacek, Z.; Vacek, S.; Cukor, J.; Remeš, J.; Hájek, V.; D’Andrea, G.; Šálek, M.; Nola, P.; et al. Silver fir tree-ring fluctuations decrease from north to south latitude—Total solar irradiance and NAO are indicated as the main influencing factors. For. Ecosyst. 2023, 10, 100150. [Google Scholar] [CrossRef]
- MZE. Zpráva o Stavu Lesa a Lesního Hospodářství České Republiky v Roce 2021; Ministerstvo zemědělství: Praha, Czech Republic, 2022. [Google Scholar]
- Málek, J. Problematika Ekologie Jedle Bělokoré a Jejího Odumírání; Československá Akademie Věd Praha: Studie ČSAV: Praha, Czech Republic, 1983; Volume 11, p. 108. [Google Scholar]
- Novák, J.; Dušek, D. Thinning of silver fir stands—Review. Rep. For. Res. 2021, 66, 176–187. [Google Scholar]
- Klika, J. Lesní Dřeviny; Československá matice lesnická: Písek, Czech Republic, 1947; p. 394. [Google Scholar]
- Svoboda, P. Život Lesa; Brázda: Praha, Czech Republic, 1952; p. 894. [Google Scholar]
- Zlatník, A. Lesnická Fytocenologie; SZN: Praha, Czech Republic, 1976; p. 495. [Google Scholar]
- Málek, J. Problematik der Ökologie der Tanne (Abies alba Mill.) und ihres Sterbens in der ČSSR. Forstw. Cbl. 1981, 100, 170–174. [Google Scholar] [CrossRef]
- Santopuoli, G.; Lasserre, B.; Di Martino, P.; Marchetti, M. Dynamics of the silver fir (Abies alba Mill.) natural regeneration in a mixed forest in the Central Apennine. Plant Biosyst. 2014, 150, 217–226. [Google Scholar] [CrossRef]
- Dănescu, A.; Kohnle, U.; Bauhus, J.; Weiskittel, A.; Albrecht, A.T. Long-term development of natural regeneration in irregular, mixed stands of silver fir and Norway spruce. For. Ecol. Manag. 2018, 430, 105–116. [Google Scholar] [CrossRef]
- Farjon, A. A Handbook of the World’s Conifers; Leiden & Boston: Brill, The Netherlands, 2017; Volume 1, p. 1112. [Google Scholar]
- Pinto, P.E.; Gégout, J.-C.; Hervé, J.-C.; Dhôte, J.-F. Respective importance of ecological conditions and stand composition on Abies alba Mill. dominant height growth. For. Ecol. Manag. 2008, 255, 619–629. [Google Scholar] [CrossRef]
- Vitasse, Y.; Bottero, A.; Rebetez, M.; Conedera, M.; Augustin, S.; Brang, P.; Tinner, W. What is the potential of silver fir to thrive under warmer and drier climate? Eur. J. For. Res. 2019, 138, 547–560. [Google Scholar] [CrossRef]
- Kučeravá, B.; Dobrovolný, L.; Remeš, J. Responses of Abies alba seedlings to different site conditions in Picea abies plantations. Dendrobiology 2013, 69, 49–58. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Colangelo, M.; de Luis, M.; Martinez del Castillo, E.; Serra-Maluquer, X. Summer drought and spring frost, but not their interaction, constrain European beech and silver fir growth in their southern distribution limits. Agric. For. Meteorol. 2019, 278, 107695. [Google Scholar] [CrossRef]
- Maxime, C.; Hendrik, D. Effects of climate on diameter growth of co-occurring Fagus sylvatica and Abies alba along an altitudinal gradient. Trees 2011, 25, 265–276. [Google Scholar] [CrossRef]
- Świercz, A.; Świątek, B.; Pietrzykowski, M. Changes in the concentrations of trace elements and supply of nutrients to silver fir (Abies alba Mill.) needles as a bioindicator of industrial pressure over the past 30 years in Świętokrzyski National Park (Southern Poland). Forests 2022, 13, 718. [Google Scholar] [CrossRef]
- Tinner, W.; Colombaroli, D.; Heiri, O.; Henne, P.; Steinacher, M.; Untenecker, J.; Vescovi, E.; Allen, J.; Carraro, G.; Conedera, M.; et al. The past ecology of Abies alba provides new perspectives on future responses of silver fir forests to global warming. Ecol. Monogr. 2013, 83, 419–439. [Google Scholar] [CrossRef]
- Bošeľa, M.; Petráš, R.; Sitková, Z.; Priwitzer, T.; Pajtík, J.; Hlavatá, H.; Sedmák, R.; Tobin, B. Possible causes of the recent rapid increase in the radial increment of silver fir in the Western Carpathians. Environ. Pollut. 2014, 184, 211–221. [Google Scholar] [CrossRef]
- Boettger, T.; Haupt, M.; Friedrich, M.; Waterhouse, J.S. Reduced climate sensitivity of carbon, oxygen and hydrogen stable isotope ratios in tree-ring celulose of silver fir (Abies alba Mill.) influenced by background SO2 in Franconia (Germany, Central Europe). Environ. Pollut. 2014, 185, 281–294. [Google Scholar] [CrossRef]
- Vacek, S.; Černý, T.; Vacek, Z.; Podrázský, V.; Mikeska, M.; Králíček, I. Long-term changes in vegetation and site conditions in beech and spruce forests of lower mountain ranges of Central Europe. For. Ecol. Manag. 2017, 398, 75–90. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Prokůpková, A.; Bulušek, D.; Podrázský, V.; Hunova, I.; Putalova, T.; Král, J. Long-term effect of climate and air pollution on health status and growth of Picea abies (L.) Karst. peaty forests in the Black Triangle region. Dendrobiology 2020, 83, 1–19. [Google Scholar] [CrossRef]
- Gill, R.M.A. A review of damage by mammals in north temperate forests: 3. Impact on trees and forests. Forestry 1992, 65, 363–388. [Google Scholar] [CrossRef]
- Vacek, S.; Prokůpková, A.; Vacek, Z.; Bulušek, D.; Šimůnek, V.; Králíček, I.; Prausová, R.; Hájek, V. Growth response of mixed beech forests to climate change, various management and game pressure in Central Europe. J. For. Sci. 2019, 65, 331–345. [Google Scholar] [CrossRef]
- Huth, F.; Wehnert, A.; Tiebel, K.; Wagner, S. Direct seeding of silver fir (Abies alba Mill.) to convert Norway spruce (Picea Abies L.) forests in Europe: A review. For. Ecol. Manag. 2017, 403, 61–78. [Google Scholar] [CrossRef]
- Kupferschmid, A.; Zimmermann, S.; Bugmann, H. Browsing regime and growth response of naturally regenerated Abies alba saplings along light gradients. For. Ecol. Manag. 2013, 310, 393–404. [Google Scholar] [CrossRef]
- Kupferschmid, A. Selective browsing behaviour of ungulates influences the growth of Abies alba differently depending on forest type. For. Ecol. Manag. 2018, 429, 317–326. [Google Scholar] [CrossRef]
- Bernard, M.; Boulanger, V.; Dupouey, J.-L.; Laurent, L.; Montpied, P.; Morin, X.; Picard, J.-F.; Said, S. Deer browsing promotes Norway spruce at the expense of silver fir in the forest regeneration phase. For. Ecol. Manag. 2017, 400, 269–277. [Google Scholar] [CrossRef]
- Konôpka, B.; Šebeň, V.; Pajtík, J. Bark browsing and recovery: A comparative study between Douglas fir and silver fir species in the Western Carpathians. Sustainability 2024, 16, 2293. [Google Scholar] [CrossRef]
- Heuze, P.; Schnitzler, A.; Klein, F. Is browsing the major factor of silver fir decline in the Vosges Mountains of France? For. Ecol. Manag. 2005, 217, 219–228. [Google Scholar] [CrossRef]
- Häsler, H.; Senn, J. Ungulate browsing on European silver fir Abies alba: The role of occasions, food shortage and diet preferences. Wildl. Biol. 2012, 18, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Hanewinkel, M.; Cullmann, D.; Schelhaas, M.-J.; Nabuurs, G.J.; Zimmermann, N. Climate change may cause severe loss in economic value of European forestland. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Konôpková, A.; Kurjak, D.; Kmeť, J.; Klumpp, R.; Longauer, R.; Ditmarová, Ľ.; Gömöry, D. Differences in photochemistry and response to heat stress between silver fir (Abies alba Mill.) provenances. Trees—Struct. Funct. 2018, 32, 73–86. [Google Scholar] [CrossRef]
- Linares, J.C.; Camarero, J.J. From pattern to process: Linking intrinsic water-use efficiency to drought-induced forest decline. Glob. Chang. Biol. 2012, 18, 1000–1015. [Google Scholar] [CrossRef]
- Büntgen, U.; Tegel, W.; Kaplan, J.; Schaub, M.; Hagedorn, F.; Bürgi, M.; Brázdil, R.; Helle, G.; Carrer, M.; Heussner, K.-U.; et al. Placing unprecedented recent fir growth in a European-wide and Holocene-long context. Front. Ecol. Environ. 2014, 12, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Wolf, H. EUFORGEN Technical Guidelines for Genetic Conser-Vation and Use for Silver Fir (Abies alba); International PlantGenetic Resources Institute: Rome, Italy, 2003; p. 6. [Google Scholar]
- Musil, I.; Hamerník, J. Jehličnaté Dřeviny: Přehled Nahosemenných (i Výtrusných) Dřevin; Academia: Praha, Czech Republic, 2007; p. 352. [Google Scholar]
- Svoboda, M.; Nagel, T.A. Gap disturbance regime in an old-growth Fagus-Abies forest in the Dinaric Mountains, Bosnia-Herzegovina. Can. J. For. Res. 2008, 38, 2728–2737. [Google Scholar]
- Úradníček, L.; Madera, P.; Tichá, S.; Koblížek, J. Dřeviny České Republiky; Lesnická práce, s.r.o.: Kostelec nad Černými lesy, Czech Republic, 2009; p. 367. [Google Scholar]
- Ballian, D.; Bogunić, F.; Bajric, M.; Kajba, D.; Kraigher, H.; Monika, K. The genetic population study of Balkan silver fir (Abies alba Mill.). Period. Biol. 2012, 114, 55–65. [Google Scholar]
- Volařík, D.; Hédl, R. Expansion to abandoned agricultural land forms an integral part of silver fir dynamics. For. Ecol. Manag. 2013, 292, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Caudullo, G.; Tinner, W.; de Rigo, D. Picea abies in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; pp. 114–116. [Google Scholar]
- Brichta, J.; Vacek, S.; Vacek, Z.; Cukor, J.; Mikeska, M.; Bílek, L.; Šimůnek, V.; Gallo, J.; Brabec, P.; Štefančík, I. Importance and potential of Scots pine (Pinus sylvestris L.) in 21 st century. Cent. Eur. For. J. 2023, 69, 3–20. [Google Scholar]
- Xiang, X.-G.; Cao, M.; Zhou, Z.-K. Fossil history and modern distribution of the genus Abies (Pinaceae). Front. For. China 2007, 2, 355–365. [Google Scholar] [CrossRef]
- Debreczy, Z.; Rácz, I. Conifers around the World: Conifers of the Temperate Zones and Adjacent Regions; Dendro Press: Wellesley, MA, USA, 2011. [Google Scholar]
- Žárník, M.; Holuša, O. Silver fir (Abies alba) in the forest-typological altitudinal vegetation zones of the Czech massif, Western and Eastern Carpathy Mts. In Jedle Bělokorá—2005. Proceedings of the Jedle Bělokorá—2005, Srní, Czech Republic, 31 October–1 November 2005; Neuhöferová, P., Ed.; ČZU FLE v Praze, Katedra Pěstování Lesů a Správa Národního Parku a Chráněné Krajinné Oblasti Šumava: Praha, Czech Republic, 2005; pp. 83–87. [Google Scholar]
- Míchal, I. Dynamika Přírodního Lesa I.—VI; Živa: Bled, Slovenia, 1983; Volume 31, pp. 8–12, 48–51, 85–88, 128–133, 163–168, 233–238. [Google Scholar]
- Poleno, Z.; Vacek, S.; Podrázský, V.; Remeš, J.; Štefančík, I.; Mikeska, M.; Kobliha, J.; Kupka, I.; Malík, V.; Turčáni, M.; et al. Pěstování Lesů III. Praktické Postupy Pěstování Lesů; Lesnická práce, s.r.o.: Kostelec nad Černými lesy, Czech Republic, 2009; p. 952. [Google Scholar]
- Zamora-Pereira, J.C.; Yousefpour, R.; Cailleret, M.; Bugmann, H.; Hanewinkel, M. Magnitude and timing of density reduction are key for the resilience to severe drought in conifer-broadleaf mixed forests in Central Europe. Ann. For. Sci. 2021, 78, 68. [Google Scholar] [CrossRef]
- Hilmers, T.; Avdagić, A.; Bartkowicz, L.; Bielak, K.; Binder, F.; Boncina, A.; Dobor, L.; Forrester, D.; Hobi, M.; Ibrahimspahic, A.; et al. The productivity of mixed mountain forests comprised of Fagus sylvatica, Picea abies, and Abies alba across Europe. Forestry 2019, 92, 512–522. [Google Scholar] [CrossRef]
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Brief 2017, 12, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Botany.cz. Available online: https://botany.cz/cs/abies-alba/ (accessed on 19 May 2024).
- Dobrowolska, D. Structure of silver fir (Abies alba Mill.) natural regeneration in the ‘Jata’ reserve in Poland. For. Ecol. Manag. 1998, 110, 237–247. [Google Scholar] [CrossRef]
- Novák, J.; Kacálek, D.; Fulín, M.; Čáp, J.; Beran, F.; Cafourek, J.; Kotrla, P.; Brožovičová, K.; Mačejovský, V.; Bezděčková, L.; et al. Podpora a Perskpetiva Jedle Bělokoré v Českých Zemích; Lesnická práce: Kostelec nad Černými lesy, Czech Republic, 2023; p. 240. [Google Scholar]
- Ferlin, F. The growth potential of understorey silver fir and Norway spruce for uneven-aged forest management in Slovenia. Forestry 2002, 75, 375–383. [Google Scholar] [CrossRef]
- Kacálek, D.; Mauer, O.; Podrázský, V.; Slodičák, M.; Houšková, K.; Špulák, O.E.A. Meliorační a Zpěvňující Funkce Lesních Dřevin; Lesnická Práce: Kostelec nad Černými lesy, Czech Republic, 2017; p. 300. [Google Scholar]
- Bercha, J. Konference: Jedle bělokorá—2005. Lesnická Práce 2006, 1, 10–11. [Google Scholar]
- Bledý, M. Využití Jedle Bělokoré (Abies alba Mill.) v Přírodě Blízkém Hospodaření v Podmínkách 2.-4. Lesního Vegetačního Stupně. Dissertation Thesis, Czech University of Life Sciences, Prague, Czech Republic, 2023. [Google Scholar]
- Hlásny, T.; Zimová, S.; Merganičová, K.; Štěpánek, P.; Modlinger, R.; Turčáni, M. Devastating outbreak of bark beetles in the Czech Republic: Drivers, impacts, and management implications. For. Ecol. Manag. 2021, 490, 119075. [Google Scholar] [CrossRef]
- Březina, D.; Michal, J.; Hlaváčková, P. The impact of natural disturbances on the Central European timber market—An analytical study. Forests 2024, 15, 592. [Google Scholar] [CrossRef]
- Jović, G.; Dukić, V.; Stajic, B.; Kazimirović, M.; Petrović, D. A dendroclimatological analysis of fir (Abies alba Mill.) growth in the Borja Mountain area of Bosnia and Herzegovina. Glas. Sumar. Fak. 2018, 118, 27–45. [Google Scholar] [CrossRef]
- Prokůpková, A.; Vacek, Z.; Vacek, S.; Bulušek, D. Natural regeneration potential of mixed forests in Kronoše Mts. National Park: Structure, dynamics and effect of game. In Proceedings of Central European Silviculture; Houšková, K., Jan, D., Eds.; Publishing Centre of Mendel University in Brno: Brno, Czech Republic, 2019; pp. 80–90. [Google Scholar]
- Hofmeister, Š.; Vacek, S.; Simon, J.; Minx, T. Struktura a vývoj přírodě blízkých porostů s jedlí bělokorou v genové základně Jánské Lázně v Krkonoších. In Increase of Close-to Nature Stand Component of Forests with Special Protection Status; Vacek, S., Ed.; Ústav Hospodářské Úpravy Lesů LDF MZLU v Brně a Katedra Pěstování Lesů FLE ČZU v Praze: Brno, Czech Republic, 2006. [Google Scholar]
- Šindelář, J.; Frýdl, J.; Novotný, P. Results of evaluation of the oldest provenance plot of the FGMRI Jíloviště-Strnady with silver fir established in 1961 on the locality Jíloviště, Baně. Rep. For. Res. 2005, 50, 24–32. [Google Scholar]
- Marage, D.; Lemperiere, G. The management of snags: A comparison in managed and unmanaged ancient forests of the Southern French Alps. Ann. For. Sci. 2005, 62, 135–142. [Google Scholar] [CrossRef]
- Motta, R.; Garbarino, F. Stand history and its consequences for the present and future dynamic in two silver fir (Abies alba Mill.) stands in the high Pesio Valley (Piedmont, Italy). Ann. For. Sci. 2003, 60, 361–370. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Dyderski, M.K.; Gęsikiewicz, K.; Horodecki, P. Tree and stand level estimations of Abies alba Mill. aboveground biomass. Ann. For. Sci. 2019, 76, 56. [Google Scholar] [CrossRef]
- Paluch, J. The influence of the spatial pattern of trees on forest floor vegetation and silver fir (Abies alba Mill.) regeneration in uneven-aged forests. For. Ecol. Manag. 2005, 205, 283–298. [Google Scholar] [CrossRef]
- Paluch, J. Ground seed density patterns under conditions of strongly overlapping seed shadows in Abies alba Mill. stands. Eur. J. For. Res. 2011, 130, 1009–1022. [Google Scholar] [CrossRef]
- Prokupková, A.; Brichta, J.; Vacek, Z.; Bielak, K.; Andrzejczyk, T.; Vacek, S.; Štefančík, I.; Bílek, L.; Fuchs, Z. Effect of vegetation on natural regeneration of mixed silver fir forests in lowlands: A case study from the Rogów region in Poland. Sylwan 2021, 165, 779–795. [Google Scholar]
- Tudoran, G.-M.; Avram, C.; Ciceu, A.; Dobre, A.-C. Growth relationships in silver fir stands at their lower-altitude limit in Romania. Forests 2021, 12, 439. [Google Scholar] [CrossRef]
- Štefančík, I. Comparison of growth of silver fir (Abies alba Mill.) in pure and mixed spruce, fir and beech stands. Rep. For. Res. 2019, 64, 94–101. [Google Scholar]
- Kobal, M.; Grčman, H.; Zupan, M.; Levanič, T.; Simončič, P.; Kadunc, A.; Hladnik, D. Influence of soil properties on silver fir (Abies alba Mill.) growth in the Dinaric Mountains. For. Ecol. Manag. 2015, 337, 77–87. [Google Scholar] [CrossRef]
- Sopushynskyi, I. Intraspecific structural signs of curly silver fir (Abies alba Mill.) growing in the Ukrainian Carpathians. J. For. Sci. 2020, 66, 299–308. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sc. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Peřina, V.; Kadlus, Z.; Jirkovský, V. Přirozená Obnova Lesních Porostů; SZN: Praha, Czech Republic, 1964; p. 167. [Google Scholar]
- Míchal, I.; Petříček, V.E.A. Péče o Chráněná Území, II. Lesní Společenstva; AOPK: Praha, Czech Republic, 1999; p. 713. [Google Scholar]
- Zakopal, V. Pěstovaní jedle ve světle nových poznatků. Rep. For. Res. 1970, 16, 24–32. [Google Scholar]
- Schütt, P. Tannenarten Europas und Klein Asiens; Ecomed Verlagsgesellschaft: Landsberg am Lech, Germany, 1994; pp. 1–132. [Google Scholar]
- Pretzsch, H.; Hilmers, T.; Biber, P.; Avdagić, A.; Binder, F.; Boncina, A.; Bošeľa, M.; Dobor, L.; Forrester, D.; Lévesque, M.; et al. Evidence of elevation-specific growth changes of spruce, fir, and beech in European mixed-mountain forests during the last three centuries. Can. J. For. Res. 2020, 50, 689–703. [Google Scholar] [CrossRef]
- Průša, E. Pěstování lesů na Typologických Základech; Lesnická Práce, s.r.o.: Kostelec nad Černými lesy, Czech Republic, 2001; p. 593. [Google Scholar]
- Korpeľ, Š. Pralesy Slovenska; Veda: Bratislava, Slovakia, 1989; p. 328. [Google Scholar]
- Tingey, D.T.; Phillips, D.L.; Johnson, M.G.; Rygiewicz, P.T.; Beedlow, P.A.; Hogsett, W.A. Estimates of douglas-fir fine root production and mortality from minirhizotrons. For. Ecol. Manag. 2005, 204, 359–370. [Google Scholar] [CrossRef]
- Weintraub, M.N.; Scott-Denton, L.E.; Schmidt, S.K.; Monson, R.K. The effects of tree rhizodeposition on soil exoenzyme activity, dissolved organic carbon, and nutrient availability in a subalpine forest ecosystem. Oecologia 2007, 154, 327–338. [Google Scholar] [CrossRef]
- Röhrig, E.; Bartsch, N. Waldbau auf Ökologischer Grundlage, 6th ed.; Parey: Hamburg, Germany; Berlin, Germany, 1992. [Google Scholar]
- Whelan, M.J.; Sanger, L.J.; Baker, M.; Anderson, J.M. Spatial patterns of throughfall and mineral ion deposition in a lowland Norway spruce (Picea abies) plantation at the plot scale. Atmos. Environ. 1998, 20, 3493–3501. [Google Scholar] [CrossRef]
- Bartsch, N.; Bauhus, J.; Vor, T. Effects of group selection and liming on nutrient cycling in European beech forest on acidic soils. In Forest Development. Succession, Environmental Stress and Forest Management. Case Studies; Dohrenbush, A., Bartsch, N., Eds.; Springer: Berlin, Germany, 2002; pp. 109–166. [Google Scholar]
- Penne, C.; Ahrends, B.; Deurer, M.; Böttcher, J. The impact of the canopy structure on the spatial variability in forest floor carbon stocks. Geoderma 2010, 158, 282–297. [Google Scholar] [CrossRef]
- Morris, D.M.; Gordon, A.G.; Gordon, A.M. Patterns of canopy interception and throughfall along a topographic sequence for black spruce dominated forest ecosystems in northwestern Ontario. Can. J. For. Res. 2003, 33, 1046–1060. [Google Scholar] [CrossRef]
- Staelens, J.; De Schrijver, A.; Verheyen, K.; Verhoest, N.E.C. Spatial variability and temporal stability of throughfall water under a dominant beech (Fagus sylvatica L.) tree in relationship to canopy cover. J. Hydrol. 2006, 330, 651–662. [Google Scholar] [CrossRef]
- Paluch, J.; Gruba, P. Inter-crown versus under-crown area: Contribution of local configuration of trees to variation in topsoil morphology, pH and moisture in Abies alba Mill. forests. Eur. J. For. Res. 2012, 131, 857–870. [Google Scholar] [CrossRef]
- Zhang, Q.; Zak, J.C. Effects of gap size on litter decomposition and microbial activity in a subtropical forest. Ecology 1995, 76, 2196–2204. [Google Scholar] [CrossRef]
- Collins, B.S.; Battaglia, L.L. Microenvironmental heterogeneity and Quercus michauxii regeneration in experimental gaps. For. Ecol. Manag. 2002, 155, 279–290. [Google Scholar] [CrossRef]
- Bauhus, J. C and N mineralization in an acid forest soil along a gap-stand gradient. Soil Biol. Biochem. 1996, 28, 923–932. [Google Scholar] [CrossRef]
- Carvalheiro, K.O.; Nepstad, D.C. Deep soil heterogeneity and fine root distribution in forests and pastures of eastern Amazonia. Plant Soil 1996, 182, 279–285. [Google Scholar] [CrossRef]
- Pärtel, M.; Wilson, S.D. Root dynamics and spatial pattern in prairie and forest. Ecology 2002, 83, 1199–1203. [Google Scholar] [CrossRef]
- Vacek, S.; Nosková, I.; Bílek, L.; Vacek, Z.; Schwarz, O. Regeneration of forest stands on permanent research plots in the Krkonoše Mts. J. For. Sci. 2010, 56, 541–554. [Google Scholar] [CrossRef]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Conifer seedling recruitment in a southeastern Canadian boreal forest: The importance of substrate. J. Veg. Sci. 1998, 9, 575–582. [Google Scholar] [CrossRef]
- Augusto, L.; Ranger, J.; Binkley, D.; Rothe, A. Impact of several common tree species of European temperate forests on soil fertility. Ann. For. Sci. 2002, 59, 233–253. [Google Scholar] [CrossRef]
- Třeštík, M.; Podrázský, V. Soil improving role of the silver fir (Abies alba Mill.): A case study. Rep. For. Res. 2017, 62, 182–188. [Google Scholar]
- Podrázský, V.; Vacek, Z.; Kupka, I.; Vacek, S.; Třeštík, M.; Cukor, J. Effects of silver fir (Abies alba Mill.) on the humus forms in Norway spruce (Picea abies (L.) H. Karst.) stands. J. For. Sci. 2018, 64, 245–250. [Google Scholar] [CrossRef]
- Čihák, T.; Vejpustková, M. Comparison of nutrient and carbon stocks in the aboveground biomass of mature silver fir (Abies alba Mill.) and Norway spruce (Picea abies L. Karst) stands. J. For. Sci. 2023, 69, 334–347. [Google Scholar] [CrossRef]
- Viewegh, J.; Kusbach, A.; Mikeska, M. Czech forest ecosystem classification. J. For. Sci. 2003, 49, 74–94. [Google Scholar] [CrossRef]
- Ellenberg, H. Vegetation Ecology of Central Europe; Cambridge University Press: Cambridge, MA, USA, 1988; p. 731. [Google Scholar]
- Senn, J.; Suter, W. Ungulate browsing on silver fir (Abies alba) in the Swiss Alps: Beliefs in search of supporting data. For. Ecol. Manag. 2003, 181, 151–164. [Google Scholar] [CrossRef]
- Klopčič, M.; Simončič, T.; Bončina, A. Comparison of regeneration and recruitment of shade-tolerant and light-demanding tree species in mixed uneven-aged forests: Experiences from the Dinaric region. Forestry 2015, 88, 552–563. [Google Scholar] [CrossRef]
- Schütz, J.-P. Silvicultural tools to develop irregular and diverse forest structures. Forestry 2002, 75, 329–337. [Google Scholar] [CrossRef]
- Korpeľ, Š.; Vinš, B. Pestovanie Jedle; Slovenské Vydavaťelstvo Pódohospodárskej Literatury: Bratislava, Slovakia, 1966; p. 342. [Google Scholar]
- Míchal, I. Obnova Ekologické Stability Lesů; Academia: Praha, Czech Republic, 1992; p. 169. [Google Scholar]
- Vacek, Z. Structure and dynamics of spruce-beech-fir forests in Nature Reserves of the Orlické hory Mts. in relation to ungulate game. Cent. Eur. For. J. 2017, 63, 23–34. [Google Scholar] [CrossRef]
- Korpeľ, Š. Die Urwälder der Westkarpaten; Gustav Fischer Verlag: Stuttgart, Germany; Jena, Germany; New York, NY, USA, 1995. [Google Scholar]
- Vacek, S.; Bílek, L.; Schwarz, O.; Hejcmanová, P.; Mikeska, M. Effect of air pollution on the health status of spruce stands. Mt. Res. Dev. 2013, 33, 40–50. [Google Scholar] [CrossRef]
- Hladík, M.; Korpeľ, Š.; Lukáč, T.; Tesař, V. Hospodárenie v Lesoch Horských Oblastí; VŠZ—Lesnická Fakulta Praha a Matice Lesnická Písek: Praha, Czech Republic; Písek, Czech Republic, 1993; p. 123. [Google Scholar]
- Vacek, Z.; Prokůpková, A.; Vacek, S.; Bulušek, D.; Šimůnek, V.; Hájek, V.; Králíček, I. Effect of Norway spruce and European beech mixing in relation to climate change: Structural and growth perspectives of mountain forests in Central Europe. For. Ecol. Manag. 2021, 488, 119019. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Cukor, J. European forests under global climate change: Review of tree growth processes, crises and management strategies. J. Environ. Manag. 2023, 332, 117353. [Google Scholar] [CrossRef]
- Dobrowolska, D. Growth and development of silver fir (Abies alba Mill.) regeneration and restoration of the species in the Karkonosze Mountains. J. For. Sci. 2008, 54, 398–408. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2002; p. 690. [Google Scholar]
- Chmelař, J. Přirozená obnova jedle (Abies alba Mill.) v pralesové rezervaci „Mionší“ v Moravskoslezských Beskydech. Lesnictví 1959, 5, 225–238. [Google Scholar]
- Meyer, H. Beitrag zur Frage der Rückgängigkeitserscheinungen der Weisstanne (Abies alba Mill.) am Nordrand ihres Naturareals. Arch. Forstw. 1957, 6, 719–787. [Google Scholar]
- Podrázský, V.; Vacek, S.; Vacek, Z.; Raj, A.; Mikeska, M.; Boček, M.; Schwarz, O.; Hošek, J.; Šach, F.; Černohous, V.; et al. Půdy Lesů a Ekosystémů nad Horní Hranicí Lesa v Národních Parcích Krkonoš; Lesnická Práce, s. r. o.: Kostelec nad Černými lesy, Czech Republic, 2010; p. 304. [Google Scholar]
- Ellenberg, H. Vegetation Mitteleuropas mit den Alpen; Verlag Eugen Ulmer: Stuttgart, Germany, 1996; Volume 6, p. 1095. [Google Scholar]
- Tinner, W.; Lotter, A. Holocene expansion of Fagus sylvatica and Abies alba in Central Europe: Where are we after eight decades of debate? Quat. Sci. Rev. 2006, 25, 526–549. [Google Scholar] [CrossRef]
- Kadlus, Z. Struktura a vývoj zmlazení smrku, jedle a buku v Orlických horách. Lesnictví 1969, 15, 381–399. [Google Scholar]
- Lieffers, V.J.; Macmillan, R.B.; MacPherson, D.; Branter, K.; Stewart, J.D. Semi-natural and intensive silvicultural systems for the boreal mixedwood forest. For. Chron. 1996, 72, 286–292. [Google Scholar] [CrossRef]
- Stancioiu, P.T.; O’Hara, K.L. Regeneration growth in different light environments of mixed species, multiaged, mountainous forests of Romania. Eur. J. For. Res. 2006, 125, 151–162. [Google Scholar] [CrossRef]
- Paluch, J. Spatial distribution of regeneration in West-Carpathian uneven-aged silver fir forests. Eur. J. For. Res. 2005, 124, 47–54. [Google Scholar] [CrossRef]
- Ligot, G.; Ameztegui, A.; Courbaud, B.; Coll, L.; Kneeshaw, D. Tree light capture and spatial variability of understory light increase with species mixing and tree size heterogeneity. Can. J. For. Res. 2016, 46, 968–977. [Google Scholar] [CrossRef]
- Košulič, M. Cesta k Přírodě Blízkému Hospodářskému Lesu; FSC ČR: Brno, Czech Republic, 2010; p. 449. [Google Scholar]
- Forrester, D.I.; Albrecht, A.T. Light absorption and light-use efficiency in mixtures of Abies alba and Picea abies along a productivity gradient. For. Ecol. Manag. 2014, 328, 94–102. [Google Scholar] [CrossRef]
- Grassi, G.; Bagnaresi, U. Foliar morphological and physiological plasticity in Picea abies and Abies alba saplings along a natural light gradient. Tree Physiol. 2001, 21, 959–967. [Google Scholar] [CrossRef]
- Hynek, V. Opatření k záchraně a reprodukci genofondu jedle bělokoré v ČSR. Práce VÚLHM 1987, 71, 39–66. [Google Scholar]
- Heuze, P.; Schnitzler, A.; Klein, F. Consequences of increased deer browsing winter on silver fir and spruce regeneration in the Southern Vosges mountains: Implications for forest management. Ann. For. Sci. 2005, 62, 175–181. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Vanclay, J.K. Mixed-species plantations of Eucalyptus with nitrogen fixing trees: A review. For. Ecol. Manag. 2006, 233, 211–230. [Google Scholar] [CrossRef]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Pretzsch, H.; Steckel, M.; Heym, M.; Biber, P.; Ammer, C.; Ehbrecht, M.; Bielak, K.; Bravo, F.; Ordóñez, C.; Collet, C.; et al. Stand growth and structure of mixed-species and monospecific stands of Scots pine (Pinus sylvestris L.) and oak (Q. robur L., Quercus petraea (Matt.) Liebl.) analysed along a productivity gradient through Europe. Eur. J. For. Res. 2020, 139, 349–367. [Google Scholar] [CrossRef]
- Del Río, M.; Pretzsch, H.; Ruiz-Peinado, R.; Jactel, H.; Coll, L.; Löf, M.; Aldea, J.; Ammer, C.; Avdagić, A.; Barbeito, I.; et al. Emerging stability of forest productivity by mixing two species buffers temperature destabilizing effect. J. Appl. Ecol. 2022, 59, 2730–2741. [Google Scholar] [CrossRef]
- Pretzsch, H.; Block, J.; Dieler, J.; Dong, P.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zingg, A. Comparison between the productivity of pure and mixed stands of Norway spruce and European beech along an ecological gradient. Ann. For. Sci. 2010, 67, 712. [Google Scholar] [CrossRef]
- Coates, K.D.; Lilles, E.B.; Astrup, R. Competitive interactions across a soil fertility gradient in a multispecies forest. J. Ecol. 2013, 101, 806–818. [Google Scholar] [CrossRef]
- Forrester, D.I.; Kohnle, U.; Albrecht, A.T.; Bauhus, J. Complementarity in mixed-species stands of Abies alba and Picea abies varies with climate, site quality and stand density. For. Ecol. Manag. 2013, 304, 233–242. [Google Scholar] [CrossRef]
- Vrška, T.; Hort, L.; Odehnalová, P.; Adam, D.; Horal, D. Prales Mionší—Historický vývoj a současný stav. J. For. Sci. 2000, 46, 411–424. [Google Scholar]
- Vrška, T.; Hort, L.; Odehnalová, P.; Adam, D.; Horal, D. Dynamika Vývoje Pralesovitých Rezervací v České Republice. Sv. 1—Českomoravská Vrchovina—Polom, Žákova hora, 1st ed.; Academia: Praha, Czech Republic, 2002; p. 213. [Google Scholar]
- Jaworski, A.; Kołodziej, Z.; Porada, K. Structure and dynamics of stands of primeval character in selected areas of the Bieszczady National Park. J. For. Sci. 2002, 48, 185–201. [Google Scholar] [CrossRef]
- Štefančík, I. Growth and development of fir (Abies alba Mill.) in mixed spruce, fir and beech stands. Ekológia 2004, 23, 144–151. [Google Scholar]
- Štefančík, I. Changes in tree species composition, stand structure, qualitative and quantitative production of mixed spruce, fir and beech stand on Stará Píla research plot. J. For. Sci. 2006, 52, 74–91. [Google Scholar] [CrossRef]
- Jaworski, A.; Podlaski, R. Processes of loss, recruitment, and increment in stands of a primeval character in selected areas of the Pieniny National Park (southern Poland). J. For. Sci. 2007, 53, 278–289. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef] [PubMed]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dauber, E. Long-term stand dynamics of managed spruce-fir-beech mountain forests in Central Europe: Structure, productivity and regeneration success. Forestry 2015, 88, 407–428. [Google Scholar] [CrossRef]
- Kupferschmid, A.; Wasem, U.; Bugmann, H. Light availability and ungulate browsing determine growth, height and mortality of Abies alba saplings. For. Ecol. Manag. 2014, 318, 359–369. [Google Scholar] [CrossRef]
- Cramer, H.H. On the predisposition to disorders of MiddleEuropean forests. Pflanzenschutz-Nachrichten Bayer 1984, 37, 97–207. [Google Scholar]
- Larsen, J.B. Das Tannensterben: Eine neue Hypothese zur Klärung des Hintergrundes dieser rätselhaften Komplexkrankheit der Weißtanne (Abies alba Mill.). Forstw. Cbl. 1986, 105, 381–396. [Google Scholar] [CrossRef]
- Krehan, H. Das Tannensterben in Europa: Eine Literaturstudie Mitkritischer Stellungnahme; FBVA-Berichte 39, Forstliche Bundesversuchsanstalt in Wien, Österreichischer Agrarverlag: Wien, Germany, 1989. [Google Scholar]
- Kramer, W. Die Weißtanne (Abies alba Mill.) in Ost- und Südosteuropa; Gustav Fischer Verlag: Stuttgart, Germany; Jena, Germany; New York, NY, USA, 1992. [Google Scholar]
- Levanic, T. Growth depression of silver fir (Abies alba Mill.) in the Dinaric phytogeographic region between 1960–1995. Zbornik Gozdarstva Lesar. 1997, 52, 137–164. [Google Scholar]
- Svenning, J.-C.; Skov, F. Limited filling of the potential range in European tree species. Ecol. Lett. 2004, 7, 565–573. [Google Scholar] [CrossRef]
- Hockenjos, W. Tannenbäume: Eine Zukunft für Abies alba; DRW-Verlag Weinbrenner GmbH & Co., KG: Leinfelden-Echterdingen, Germany, 2008; p. 232. [Google Scholar]
- Allen, J.R.M.; Huntley, B. Last interglacial palaeovegetation, palaeoenvironments and chronology: A new record from Lago Grande di Monticchio, southern Italy. Quat. Sci. Rev. 2009, 28, 1521–1538. [Google Scholar] [CrossRef]
- Opravil, E. Jedle bělokorá (Abies alba Mill.) v československém kvartéru. Časopis Slez. Muz. 1976, 25, 45–67. [Google Scholar]
- Tinner, W.; Hubschmid, P.; Wehrli, M.; Ammann, B.; Conedera, M. Long-term forest fire ecology and dynamics in southern Switzerland. J. Ecol. 1999, 87, 273–289. [Google Scholar] [CrossRef]
- Dannecker, K. Aus der Hohen Schule Weisstannenwaldes; J.D. Sauerländer: Frankfurt, Germany, 1955; p. 206. [Google Scholar]
- Manion, P.; Lachance, D. Forest Decline Concepts; APS Press: St. Paul, MN, USA; University of Minnesota: Minneapolis, MN, USA, 1992; p. 249. [Google Scholar]
- Wentzel, K.F. Weissitanne = immissionsempfindlichste einheimische Baumart. Allg. Forstztg. 1980, 35, 373–374. [Google Scholar]
- Ulrich, B. Eine ökosystemare Hypothese über die Ursachen des Tannensterbens (Abies alba Mill.). Forstwiss. Cbl. 1981, 100, 228–236. [Google Scholar] [CrossRef]
- Longauer, R. Genetic variation of European silver fir (Abies alba Mill.) in the Western Carpathians. J. For. Sci. 2001, 47, 429–438. [Google Scholar]
- Ficko, A.; Boncina, A. Silver fir (Abies alba Mill.) distribution in Slovenian forests. Zb. Gozdarstva Lesar. 2006, 79, 19–35. [Google Scholar]
- Elling, W.; Dittmar, C.; Pfaffelmoser, K.; Rötzer, T. Dendroecological assessment of the complex causes of decline and recovery of the growth of silver fir (Abies alba Mill.) in Southern Germany. For. Ecol. Manag. 2009, 257, 1175–1187. [Google Scholar] [CrossRef]
- Diaci, J. Silver fir decline in mixed old-growth forests in Slovenia: An interaction of air pollution, changingforest matrix and climate. In AirPollution—New Developments; Moldoveanu, A., Ed.; InTech: London, UK, 2011; pp. 263–274. [Google Scholar]
- Frank, G.; Mayer, H. Waldschadensinventur im Fichten-Tannen-Buchen-Urwaldrest Neuwald; Cbl. f. d. ges. Forstw: Wien, Germany, 1988; pp. 104–123. [Google Scholar]
- Berge, E.; Bartnicki, J.; Olendrzynski, K.; Tsyro, S.G. Long-termtrends in emissions and transboundary transport of acidifying airpollution in Europe. J. Environ. Manag. 1999, 57, 31–50. [Google Scholar] [CrossRef]
- Stern, D.I. Global sulfur emissions from 1850 to 2000. Chemosphere 2005, 58, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Čavlović, J.; Bončina, A.; Božić, M.; Goršić, E.; Simončič, T.; Teslak, K. Depression and growth recovery of silver fir in uneven-aged Dinaric forests in Croatia from 1901 to 2001. For. Int. J. For. Res. 2015, 88, 586–589. [Google Scholar] [CrossRef]
- Balcar, V.; Vacek, S.; Henžlík, V. Poškození a úhyn lesních porostů v Sudetských horách. In Protection of Forest Ecosystems, Selected Problems of Forestry in Sudety Mts.; Paschalis, P., Zajaczkowski, S., Eds.; Biuro GEF: Warszawa, Poland, 1997; pp. 29–57. [Google Scholar]
- Hernández, L.; Camarero, J.J.; Gil-Pelegrín, E.; Sánchez, M.A.S.; Cañellas, I.; Montes, F. Biotic factors and increasing aridity shape the altitudinal shifts of marginal Pyrenean silver fir populations in Europe. For. Ecol. Manag. 2019, 432, 558–567. [Google Scholar] [CrossRef]
- Diaci, J.; Roženbergar, D.; Anic, I.; Mikac, S.; Saniga, M.; Kucbel, S.; Višnjić, Ć.; Ballian, D. Structural dynamics and synchronous silver fir decline in mixed old-growth mountain forests in Eastern and Southeastern Europe. Forestry 2011, 84, 479–491. [Google Scholar] [CrossRef]
- Ficko, A.; Poljanec, A.; Boncina, A. Do changes in spatial distribution, structure and abundance of silver fir (Abies alba Mill.) indicate its decline? For. Ecol. Manag. 2011, 261, 844–854. [Google Scholar] [CrossRef]
- Brinar, M. Življenjska kriza jelke na slovenskem ozemlju v zvezi s klimatičnimi fluktuacijami. Gozdarski vestnik 1964, 22, 97–144. [Google Scholar]
- Schütt, P. Die gegenwärtige Epidemic des Tannensterbens. Eur. J. Plant Pathol. 1978, 7, 187–190. [Google Scholar]
- Wick, L.; Möehl, A. The mid-Holocene extinction of silver fir (Abies alba) in the Southern Alps: A consequence of forest fires? Palaeobotanical records and forest simulations. Veg. Hist. Archaeobot. 2006, 15, 435–444. [Google Scholar] [CrossRef]
- Anic, I.; Vukelic, J.; Mikac, S.; Baksic, D.; Ugarkovic, D. Utjecaj globalnih klimatskih promjena na ekološku nišu obične jele (Abies alba Mill.) u Hrvatskoj. Šumarski List 2009, 133, 135–144. [Google Scholar]
- Cailleret, M.; Nourtier, M.; Amm, A.; Durand-Gillmann, M.; Davi, H. Drought-induced decline and mortality of silver fir differ among three sites in Southern France. Ann. For. Sci. 2014, 71, 643–657. [Google Scholar] [CrossRef]
- Linares, J.C. Biogeography and evolution of Abies (Pinaceae) in the Mediterranean Basin: The roles of long-term climatic change and glacial refugia. J. Biogeogr. 2011, 38, 619–630. [Google Scholar] [CrossRef]
- Bošeľa, M.; Ionel, P.; Gömöry, D.; Longauer, R.; Tobin, B.; Kyncl, J.; Kyncl, T.; Nechita, C.; Petráš, R.; Sidor, C.; et al. Effects of postglacial phylogeny and genetic diversity on the growth variability and climate sensitivity of European silver fir. J. Ecol. 2016, 104, 716–724. [Google Scholar] [CrossRef]
- Leonarduzzi, C.; Piotti, A.; Spanu, I.; Giovanni Giuseppe, V. Effective gene flow in a historically fragmented area at the southern edge of silver fir (Abies alba Mill.) distribution. Tree Genet. Genomes 2016, 12, 95. [Google Scholar] [CrossRef]
- Ondrejčík, R.; Krajmerová, D.; Longauer, R. Genetické riziká v cykle produkcie lesného reprodukčného materiálu na príklade uznaného porastu a sadeníc jedle bielej. In Adaptívny Manažment Pestovania Lesov v Procese Klimatickej Zmeny a Globálneho Otepľovania: Adaptive Management of Silviculture in the Process of Climate Change ang Global Warming; Jaloviar, P., Saniga, M., Eds.; Proceedings of Central European Silviculture; Technická univerzita vo Zvolene: Zvolen, Slovakia, 2017; pp. 87–94. [Google Scholar]
- Csilléry, K.; Buchmann, N.; Fady, B. Adaptation to drought is coupled with slow growth, but independent from phenology in marginal silver fir (Abies alba Mill.) populations. Evol. Appl. 2020, 13, 2357–2376. [Google Scholar] [CrossRef] [PubMed]
- Csilléry, K.; Ovaskainen, O.; Sperisen, C.; Buchmann, N.; Widmer, A.; Gugerli, F. Adaptation to local climate in multi-trait space: Evidence from silver fir (Abies alba Mill.) populations across a heterogeneous environment. Heredity 2020, 124, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Mihai, G.; Alexandru, A.M.; Stoica, E.; Birsan, M.V. Intraspecific growth response to drought of Abies alba in the Southeastern Carpathians. Forests 2021, 12, 387. [Google Scholar] [CrossRef]
- Durand-Gillmann, M.; Cailleret, M.; Boivin, T.; Nageleisen, L.-M.; Davi, H. Individual vulnerability factors of silver fir (Abies alba Mill.) to parasitism by two contrasting biotic agents: Mistletoe (Viscum album L. ssp. Abietis) and bark beetles (Coleoptera: Curculionidae: Scolytinae) during a decline process. Ann. For. Sci. 2012, 71, 659–673. [Google Scholar]
- Lebourgeois, F.; Eberlé, P.; Mérian, P.; Seynave, I. Social status-mediated tree-ring responses to climate of Abies alba and Fagus sylvatica shift in importance with increasing stand basal area. For. Ecol. Manag. 2014, 328, 209–218. [Google Scholar] [CrossRef]
- Mrkva, R. Korovnice kavkazska (Dreyfusia nordmannianae Eckstein), obrana proti ní a její podíl na ústupu jedle. Lesnictvi—Forestry 1994, 40, 361–370. [Google Scholar]
- Zubrík, M. Kôrovnica kaukazská—Významný škodca jedle. Les 1994, 50, 21–22. [Google Scholar]
- Ujházy, K.; Križová, E.; Vančo, M.; Freňáková, E.; Ondruš, M. Herblayer dynamics of primeval fir–beech forests in central Slovakia. In Natural Forests in the Temperate Zone of Europe—Values and Utilisation; Commarmot, B., Hamor, F.D., Eds.; Federal Research Institute WSL, Birmensdorf & Carpathian Biosphere Reserve: Rakhiv, Ukraine; Birmensdorf, Switzerland, 2005. [Google Scholar]
- Ficko, A.; Roessiger, J.; Boncina, A. Can the use of continuous cover forestry alone maintain silver fir (Abies alba Mill.) in central European mountain forests? Forestry 2016, 89, 412–421. [Google Scholar] [CrossRef]
- Motta, R. Impact of wild ungulates on forest regeneration and tree composition of mountain forests in the Western Italian Alps. For. Ecol. Manag. 1996, 88, 93–98. [Google Scholar] [CrossRef]
- Klopčič, M.; Jerina, K.; Bončina, A. Long-term changes of structure and tree species composition in Dinaric uneven-aged forests: Are red deer an important factor? Eur. J. For. Res. 2010, 129, 277–288. [Google Scholar] [CrossRef]
- Čermák, P.; Grundmann, P. Effects of browsing on the condition and development of regeneration of trees in the region of Rýchory (KRNAP). Acta Univ. Agric. Silvic. Mendel. Brun. 2006, 54, 7–14. [Google Scholar] [CrossRef]
- Homolka, M.; Heroldová, M. Impact of large herbivores on mountain forest stands in the Beskydy Mountains. For. Ecol. Manag. 2003, 181, 119–129. [Google Scholar] [CrossRef]
- Slanař, J.; Vacek, Z.; Vacek, S.; Bulušek, D.; Cukor, J.; Štefančík, I.; Bílek, L.; Král, J. Long-term transformation of submontane spruce-beech forests in the Jizerské hory Mts.: Dynamics of natural regeneration. Cent. Eur. For. J. 2017, 63, 212–224. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Slanař, J.; Bílek, L.; Bulušek, D.; Štefančík, I.; Králíček, I.; Vančura, K. Adaption of Norway spruce and European beech forests under climate change: From resistance to close-to-nature silviculture. Cent. Eur. For. J. 2019, 65, 129–144. [Google Scholar] [CrossRef]
- Schwegmann, S.; Mörsdorf, M.; Bhardwaj, M.; Storch, I. Effects of understory characteristics on browsing patterns of roe deer in central European mountain forests. Ecol. Evol. 2023, 13, e10431. [Google Scholar] [CrossRef] [PubMed]
- Motta, R. Wild ungulate browsing, natural regeneration and silviculture in the Italian Alps. J. Sustain. For. 1998, 8, 35–53. [Google Scholar] [CrossRef]
- Caudullo, G.; de Battisti, R.; Colpi, C.; Vazzola, C.; da Ronch, F. Ungulate damage and silviculture in the Cansiglio Forest (Veneto Prealps, NE Italy). J. Nat. Conserv. 2003, 10, 233–241. [Google Scholar] [CrossRef]
- Borowski, Z.; Gil, W.; Bartoń, K.; Zajączkowski, G.; Łukaszewicz, J.; Tittenbrun, A.; Radliński, B. Density-related effect of red deer browsing on palatable and unpalatable tree species and forest regeneration dynamics. For. Ecol. Manag. 2021, 496, 119442. [Google Scholar] [CrossRef]
- Szwagrzyk, J.; Gazda, A.; Muter, E.; Pielech, R.; Szewczyk, J.; Zięba, A.; Zwijacz-Kozica, T.; Wiertelorz, A.; Pachowicz, T.; Bodziarczyk, J. Effects of species and environmental factors on browsing frequency of young trees in mountain forests affected by natural disturbances. For. Ecol. Manag. 2020, 474, 118364. [Google Scholar] [CrossRef]
- Kupferschmid, A.D.; Greilsamer, R.; Brang, P.; Bugmann, H. Assessment of the impact of ungulate browsing on tree regeneration. Schweiz. Z. Forstwes. 2022, 170, 125–134. [Google Scholar] [CrossRef]
- Vasiliauskas, R. Damage to trees due to forestry operations and its pathological significance in temperate forests: A literature review. Forestry 2001, 74, 319–336. [Google Scholar] [CrossRef]
- Čermák, P.; Mrkva, R.; Horsák, P.; Špiřík, M.; Beranová, P.; Orálková, J.; Kadlec, M.; Zárybnický, O.; Svatoš, M. Impact of ungulate browsing on forest dynamics. Folia For. Bohem. 2011, 20, 80. [Google Scholar]
- Vlad, R.; Sidor, C.G. Research for the estimate of rotten stem wood volume in Norway spruce stands damaged by deer species. Rev. Pădurilor 2013, 128, 27–32. [Google Scholar]
- Cukor, J.; Vacek, Z.; Linda, R.; Sharma, R.P.; Vacek, S. Afforested farmland vs. forestland: Effects of bark stripping by Cervus elaphus and climate on production potential and structure of Picea abies forests. PLoS ONE 2019, 14, e0221082. [Google Scholar] [CrossRef]
- Cukor, J.; Zeidler, A.; Vacek, Z.; Vacek, S.; Šimůnek, V.; Gallo, J. Comparison of growth and wood quality of Norway spruce and European larch: Effect of previous land use. Eur. J. For. Res. 2020, 139, 459–472. [Google Scholar] [CrossRef]
- Bazzigher, G.; Schmid, P. Sturmschäden und Fäule. Schweiz. Z. Forstwes. 1969, 10, 521–535. [Google Scholar]
- Kohnle, U.; Kändler, G. Is Silver fir (Abies alba) less vulnerable to extraction damage than Norway spruce (Picea abies)? Eur. J. For. Res. 2007, 126, 121–129. [Google Scholar] [CrossRef]
- Metzler, B.; Hecht, U.; Nill, M.; Brüchert, F.; Fink, S.; Kohnle, U. Comparing Norway spruce and silver fir regarding impact of bark wounds. For. Ecol. Manag. 2012, 274, 99–107. [Google Scholar] [CrossRef]
- Oven, P.; Torelli, N. Wound response of the bark in healthy and declining silver firs (Abies alba). IAWA J. 1994, 15, 407–415. [Google Scholar] [CrossRef]
- Oven, P.; Torelli, N. Response of the cambial zone in conifers to wounding. Phyton Ann. Rei Bot. 1999, 39, 133–137. [Google Scholar]
- Pach, M. Spałowanie jodły na terenie Leśnego Zakładu Doświadczalnego w Krynicy (Beskid Sądecki) oraz jego wpływ na wybrane cechy morfologiczne koron. Acta Agr. Silv. Ser. Silv. 2002, 40, 31–47. [Google Scholar]
- Pach, M. Wpływ spałowania powodowanego przez jelenie na szerokość słojów rocznych pni jodeł. Acta Agr. Silv. Ser. Silv. 2003, 41, 75–82. [Google Scholar]
- Pach, M. Wpływ spałowania powodowanego przez jelenie na przyrost wysokości i miąższości jodeł (Abies alba Mill.). Acta Agr. Silv. Ser. Silv. 2004, 42, 35–48. [Google Scholar]
- Pach, M. Zasięg i dynamika rozprzestrzeniania się zgnilizny wewnątrz pni jodeł w wyniku ich spałowania przez jeleniowate. Extent and dynamics of wood decay spreading inward fir stems as a result of bark stripping by ungulates. Sylwan 2005, 149, 23–35. [Google Scholar]
- Pach, M. Tempo zarastania spał na jodle oraz niektóre czynniki na nie wpływające. The rate of bark-stripping wound closure in fir and some factors affecting it. Sylwan 2008, 152, 46–57. [Google Scholar]
- Isomäki, A.; Kallio, T. Consequences of injury caused by timber harvesting machines on the growth and decay of spruce (Picea abies (L.) Karst.). Acta For. Fenn. 1974, 136, 1–25. [Google Scholar] [CrossRef]
- Gregory, S.C. The development of stain in wounded Sitka spruce stems. Forestry 1986, 59, 199–208. [Google Scholar] [CrossRef]
- Barszcz, P.; Jamrozy, G. Deprecjacja drewna jodeł i jesionów spałowanych przez jelenie w lasach Beskidu Sądeckiego. Sylwan 2001, 145, 47–57. [Google Scholar]
- Vacek, Z.; Cukor, J.; Linda, R.; Vacek, S.; Šimůnek, V.; Brichta, J.; Gallo, J.; Prokůpková, A. Bark stripping, the crucial factor affecting stem rot development and timber production of Norway spruce forests in Central Europe. For. Ecol. Manag. 2020, 474, 118360. [Google Scholar] [CrossRef]
- Pietrzykowski, E.; McArthur, C.; Fitzgerald, H.; Goodwin, A.N. Influence of patch characteristics on browsing of tree seedlings by mammalian herbivores. J. Appl. Ecol. 2003, 40, 458–469. [Google Scholar] [CrossRef]
- Marada, P.; Cukor, J.; Linda, R.; Vacek, Z.; Vacek, S.; Havránek, F. Extensive orchards in the agricultural landscape: Effective protection against fraying damage caused by roe deer. Sustainability 2019, 11, 3738. [Google Scholar] [CrossRef]
- Stutz, R.; Croak, B.; Leimar, O.; Bergvall, U. Borrowed plant defences: Deterring browsers using a forestry by-product. For. Ecol. Manag. 2017, 390, 1–7. [Google Scholar] [CrossRef]
- Spake, R.; Bellamy, C.; Graham, L.; Watts, K.; Wilson, T.; Norton, L.; Wood, C.; Schmucki, R.; Bullock, J.; Eigenbrod, F. An analytical framework for spatially targeted management of natural capital. Nat. Sustain. 2019, 2, 90–97. [Google Scholar] [CrossRef]
- Carpio, A.J.; Apollonio, M.; Acevedo, P. Wild ungulate overabundance in Europe: Contexts, causes, monitoring and management recommendations. Mammal Rev. 2021, 51, 95–108. [Google Scholar] [CrossRef]
- Valente, A.M.; Acevedo, P.; Figueiredo, A.M.; Fonseca, C.; Torres, R.T. Overabundant wild ungulate populations in Europe: Management with consideration of socio-ecological consequences. Mammal Rev. 2020, 50, 353–366. [Google Scholar] [CrossRef]
- Ruprecht, J.S.; Koons, D.N.; Hersey, K.R.; Hobbs, N.T.; MacNulty, D.R. The effect of climate on population growth in a cold-adapted ungulate at its equatorial range limit. Ecosphere 2020, 11, e03058. [Google Scholar] [CrossRef]
- Peláez, M.; San Miguel, A.; Rodríguez-Vigal, C.; Moreno-Gómez, Á.; del Rincón Garoz, A.G.; García-Calvo, R.P. Using retrospective life tables to assess the effect of extreme climatic conditions on ungulate demography. Ecol. Evol. 2022, 12, e8218. [Google Scholar] [CrossRef]
- Capretti, P.; Korhonen, K.; Mugnai, L.; Romagnoli, C. An Intersterility group of Heterobasidion annosum specialized to Abies alba. Eur. J. Plant Pathol. 1990, 20, 231–240. [Google Scholar] [CrossRef]
- Korhonen, K.; Capretti, P.; Karjaleinen, R.; Stenlid, J. Distribution of Heterobasidion annosum intersterility groups in Europe. In Heterobasidion Annosum: Biology, Ecology, Impact and Control; Woodward, S.J., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: Wallingford, UK, 1998; pp. 93–104. [Google Scholar]
- Oliva, J.; Colinas, C. Epidemiology of Heterobasidion abietinum and Viscum album on silver fir (Abies alba) stands of the Pyrenees. For. Pathol. 2010, 40, 19–32. [Google Scholar] [CrossRef]
- Korhonen, K.; Holdenrieder, O. Neue Erkenntnisse über den Wurzelschwamm (Heterobasidion annosum s. l.)—Eine Literaturübersicht. Forst Holz 2005, 60, 206–211. [Google Scholar]
- Holdenrieder, O. Krankheiten der Tanne Abies spp. Schweiz. Beitr. Dendrol. 1994, 43, 11–21. [Google Scholar]
- Krieglsteiner, G.J.; Kaiser, A. Die Großpilze Baden-Württembergs. Allgemeiner Teil: Ständerpilze: Gallert-, Rinden-, Stachel- und Porenpilze; Die Großpilze Baden-Württemberg, 1, Verlag Eugen Ulmer: Stuttgart, Germany, 2000. [Google Scholar]
- Kulbanska, I.M.; Plikhtyak, P.P.; Shvets, M.V.; Soroka, M.I.; Goychuk, A.F. Lelliottia nimipressuralis (Carter 1945) Brady et al. 2013 as the causative agent of bacterial wetwood disease of common silver fir (Abies alba mill.). Folia For. Pol. 2022, 64, 173–183. [Google Scholar] [CrossRef]
- Pusz, W.; Baturo-Ciesniewska, A.; Kaczmarek-Pieńczewska, A.; Zwijacz-Kozica, T.; Patejuk, K. The mycobiota of needles and shoots of silver fir (Abies alba Mill.) with symptoms of Herpotrichia needle browning in the Tatra Mts. (Poland). Ann. For. Res. 2020, 63, 45–56. [Google Scholar] [CrossRef]
- Janssen, T.; Wulf, A. Zur Bedeutung von Misteln im Forstschutz; Biologische Bundesanstalt für Land und Forstschutz: Berlin, Germany, 1999; p. 129. [Google Scholar]
- Procházka, F. A centre of occurrence of Viscum album subsp. album in eastern Bohemia and an overview of the diversity of its host plants in the Czech Republic. Preslia 2004, 76, 349–359. [Google Scholar]
- Iszkulo, G.; Armatys, L.; Dering, M.; Ksepko, M.; Tomaszewski, D.; Wazna, A.; Giertych, M.J. Mistletoe as a threat to the health state of coniferous forest. Sylwan 2020, 164, 226–236. [Google Scholar]
- Tubeuf, K. Monographie der Mistel; München & Berlin: Oldenbourg, Germany, 1923. [Google Scholar]
- Knuchel, H. Die Holzfehler; Classen: Zürich, Switzerland, 1947. [Google Scholar]
- König, E. Fehler des Holzes; Holz-Zentralblatt Verlag: Stuttgart, Germany, 1957; 256p. [Google Scholar]
- Noetzli, K.P.; Müller, B.; Sieber, T.N. Impact of population dynamics of white mistletoe (Viscum album ssp. abietis) on European silver fir (Abies alba). Ann. For. Sci. 2003, 60, 773–779. [Google Scholar]
- Brang, P.; Spathelf, P.; Larsen, J.; Bauhus, J.; Boncina, A.; Chauvin, C.; Drössler, L.; García-Güemes, C.; Heiri, C.; Kerr, G.; et al. Suitability of close-to-nature silviculture for adapting temperate European forests to climate change. Forestry 2014, 87, 492–503. [Google Scholar] [CrossRef]
- Nagel, T.A.; Mikac, S.; Dolinar, M.; Klopčič, M.; Keren, S.; Svoboda, M.; Diaci, J.; Bončina, A.; Paulic, V. The natural disturbance regime in forests of the Dinaric Mountains: A synthesis of evidence. For. Ecol. Manag. 2017, 388, 29–42. [Google Scholar] [CrossRef]
- Maringer, J.; Stelzer, A.S.; Paul, C.; Albrecht, A.T. Ninety-five years of observed disturbance-based tree mortality modeled with climate-sensitive accelerated failure time models. Eur. J. For. Res. 2021, 140, 255–272. [Google Scholar] [CrossRef]
- Vejpustková, M.; Čihák, T.; Fišer, P. The increasing drought sensitivity of silver fir (Abies alba Mill.) is evident in the last two decades. J. For. Sci. 2023, 69, 67–79. [Google Scholar] [CrossRef]
- Koffi, B.; Koffi, E. Heat waves across Europe by the end of the 21st century: Multiregional climate simulations. Clim. Res. 2008, 36, 153–168. [Google Scholar] [CrossRef]
- Kyselý, J.; Gaál, L.; Beranová, R.; Plavcová, E. Climate change scenarios of precipitation extremes in Central Europe from ENSEMBLES regional climate models. Theor. Appl. Climatol. 2011, 104, 529–542. [Google Scholar] [CrossRef]
- Bauhus, J.; Forrester, D.I.; Gardiner, B.; Jactel, H.; Vallejo, R.; Pretzsch, H. Ecological stability of mixed-species forests. In Mixed-Species Forests; Pretzsch, H., Forrester, D.I., Bauhus, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 337–382. [Google Scholar]
- Steckel, M.; Rio, M.; Heym, M.; Aldea, J.; Bielak, K.; Brazaitis, G.; Černý, J.; Coll, L.; Collet, C.; Ehbrecht, M.; et al. Species mixing reduces drought susceptibility of Scots pine (Pinus sylvestris L.) and oak (Quercus robur L., Quercus petraea (Matt.) Liebl.)—Site water supply and fertility modify the mixing effect. For. Ecol. Manag. 2020, 461, 117908. [Google Scholar] [CrossRef]
- Forrester, D.; Bauhus, J. A review of processes behind diversity—Productivity relationships in forests. Curr. For. Rep. 2016, 2, 45–61. [Google Scholar] [CrossRef]
- Jactel, H.; Bauhus, J.; Boberg, J.; Bonal, D.; Castagneyrol, B.; Gardiner, B.; Olabarria, J.; Koricheva, J.; Meurisse, N.; Brockerhoff, E. Tree diversity drives forest stand resistance to natural disturbances. Curr. For. Rep. 2017, 3, 223–243. [Google Scholar] [CrossRef]
- Vitali, V.; Büntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Chang. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef]
- Ciceu, A.; Bronisz, K.; García-Duro, J.; Badea, O. Age-independent diameter increment models for mixed mountain forests. Eur. J. For. Res. 2022, 141, 781–800. [Google Scholar] [CrossRef]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate forest trees: Climatic drivers and levels of variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef] [PubMed]
- Zang, C.; Rothe, A.; Weis, W.; Pretzsch, H. Zur Baumarteneignung bei Klimawandel: Ableitung der Trockenstress-Anfälligkeit wichtiger Waldbaumarten aus Jahrringbreiten. Allg. Forst Jagdztg. 2011, 182, 98–112. [Google Scholar]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Koch, O.; de Avila, A.L.; Heinen, H.; Albrecht, A.T. Retreat of major European tree species distribution under climate change—Minor natives to the rescue? Sustainability 2022, 14, 5213. [Google Scholar] [CrossRef]
- Šindelář, J.; Frýdl, J. Perspektivy jedle bělokoré (Abies alba Mill.) v lesním hospodářství České republiky. In Jedle bělokorá—2005. Proceedings of the Jedle Bělokorá—2005, Srní, Czech Republic, 31 October–1 November 2005; Neuhöferová, P., Ed.; ČZU FLE v Praze, Katedra Pěstování Lesů a Správa Národního Parku a Chráněné krajinné Oblasti Šumava: Praha, Czech Republic, 2005. [Google Scholar]
- Spittlehouse, D.L.; Stewart, R.B. Adaptation to climate change in forest management. J. Ecosyst. Manag. 2003, 4, 1–11. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.-J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive forest management in central Europe: Climate change impacts, strategies and integrative concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Hiltunen, M.; Strandman, H.; Kilpeläinen, A. Optimizing forest management for climate impact and economic profitability under alternative initial stand age structures. Biomass Bioenergy 2021, 147, 106027. [Google Scholar] [CrossRef]
- Sterck, F.; Vos, M.; Hannula, S.; de Goede, S.; de Vries, W.; Ouden, J.; Nabuurs, G.-J.; var der Putten, W.; Veen, C. Optimizing stand density for climate-smart forestry: A way forward towards resilient forests with enhanced carbon storage under extreme climate events. Soil Biol. Biochem. 2021, 162, 108396. [Google Scholar] [CrossRef]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef]
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Barnard, P.; Moomaw, W.R. World scientists’ warning of a climate emergency. BioScience 2020, 70, 8–12. [Google Scholar] [CrossRef]
- Woodall, C.W.; Oswalt, C.M.; Westfall, J.A.; Perry, C.H.; Nelson, M.D.; Finley, A.O. An indicator of tree migration in forests of the eastern United States. For. Ecol. Manag. 2009, 257, 1434–1444. [Google Scholar] [CrossRef]
- Gömöry, D.; Krajmerova, D.; Hrivnák, M.; Longauer, R. Assisted migration vs. close-to-nature forestry: What are the prospects for tree populations under climate change? Cent. Eur. For. J. 2020, 66, 63–70. [Google Scholar] [CrossRef]
- Feurdean, A.; Tămaş, T.; Tanţău, I.; Farcas, S. Elevational variation in regional vegetation responses to late-glacial climate changes in the Carpathians. J. Biogeogr. 2011, 39, 258–271. [Google Scholar] [CrossRef]
- Feurdean, A.; Bhagwat, S.; Willis, K.; Birks, H.; Lischke, H.; Hickler, T. Tree migration-rates: Narrowing the gap between inferred post-glacial rates and projected rates. PLoS ONE 2013, 8, e71797. [Google Scholar] [CrossRef] [PubMed]
- Mátyás, C. What do feld trials tell about the future use of forest reproductive material? In Climate Change and Forest Genetic Diversity: Implications for Sustainable Forest Management in Europe; Koskela, J., Buck, A., Teissier du Cros, E., Eds.; Bioversity International: Rome, Italy, 2007; pp. 53–69. [Google Scholar]
- Iverson, L.; McKenzie, D. Tree-species range shifts in a changing climate: Detecting, modeling, assisting. Landsc. Ecol. 2013, 28, 879–889. [Google Scholar] [CrossRef]
- Iverson, L.; Schwartz, M.; Prasad, A. How fast and far might tree species migrate in the eastern United States due to climate change? Glob. Ecol. Biogeogr. 2004, 13, 209–219. [Google Scholar] [CrossRef]
- Hacket-Pain, A.; Bogdziewicz, M. Climate change and plant reproduction: Trends and drivers of mast seeding change. Philos. Trans. R. Soc. B 2021, 376, 20200379. [Google Scholar] [CrossRef] [PubMed]
- Chroust, L. Pěstování Lesa. Doplňkový Učební Text; Ústav Pěstování Lesa LDF MZLU: Brno, Czech Republic, 2001; Available online: https://rumex.mendelu.cz/uzpl/pestovani_v_heslech/pestov/pest_pojmy.html (accessed on 18 April 2024).
- Feurdean, A.; Willis, K.J. Long-term variability of Abies alba in NW Romania: Implications for its conservation management. Divers. Distrib. 2008, 14, 1004–1017. [Google Scholar] [CrossRef]
- Hlavová, Z. Technologie skladování a předosevní příprava pro jedli bělokorou a buk lesní používané v lesnickém závodě Týniště nad Orlicí. In Pěstování Sadebního Materiálu z Dlouhodobě Skladovaného Osiva Buku a Jedle. Proceedings of the Pěstování Sadebního Materiálu z Dlouhodobě Skladovaného Osiva Buku a Jedle, Hradec Králové, Czech Republic, 17 June 1999; AVE Centrum: Opava, Czech Republic, 1999; pp. 18–20. [Google Scholar]
- Chválová, K. Skúsenosti so spracováním, skladováním a predsejbovou prípravou buka a jedle na Slovensku. In Pěstování Sadebního Materiálu z Dlouhodobě Skladovaného Osiva Buku a Jedle. Proceedings of the Pěstování Sadebního Materiálu z Dlouhodobě Skladovaného Osiva Buku a Jedle, Hradec Králové, Czech Republic, 17 June 1999; AVE Centrum: Opava, Czech Republic, 1999; pp. 27–31. [Google Scholar]
- Teodosiu, M.; Botezatu, A.; Ciocîrlan, E.; Mihai, G. Variation of cones production in a silver fir (Abies alba Mill.) clonal seed orchard. Forests 2023, 14, 17. [Google Scholar] [CrossRef]
- de Andrés, E.G.; Camarero, J.J.; Martinéz, I.; Coll, L. Uncoupled spatiotemporal patterns of seed dispersal and regeneration in Pyrenean silver fir populations. For. Ecol. Manag. 2014, 319, 18–28. [Google Scholar] [CrossRef]
- Ruiz de la Torre, J. Flora Mayor; ICONA-OAPN: Madrid, Spain, 2006. [Google Scholar]
- Gradi, A. La conoscenza del contenuto d acqua degli strobili a dei semi faktore determinante per una razionale preparazione delle sementi di confere a per la loro conservazione. Monti Bosch 1963, 14, 195–208. [Google Scholar]
- Walter, V. Rozmnožování Okrasných Stromů a Keřů (Propagation of the Ornamental Trees and Shrubs); Brázda: Praha, Czech Republic, 1978; 312p. [Google Scholar]
- Leadem, C.L. Quick Tests for Tree Seed Viability; Land Management Report No. 18; BC Ministry of Forests, Research Branch: Victoria, BC, Canada, 1984; p. 45. [Google Scholar]
- Palátová, E. Zakládání Lesa I. Lesní Semenářství; MZLU: Brno, Czech Republic, 2008; p. 119. [Google Scholar]
- Řezníčková, J.; Bezděčková, L.; Procházková, Z. Cone collection and processing, storing, pre-sowing treatment and quality of European silver fir (Abies alba) seeds: A literature review. Rep. For. Res. 2010, 55, 180–186. [Google Scholar]
- Messer, H.; Hanau, W. Der Wassergehalt des Forstsaatgutes als Grundlage der Ernte-, Veredelungs- und Aufbewahrungs- massnahmen. Forst Holz. 1959, 9, 226–229. [Google Scholar]
- Musil, I.; Hamerník, J. Lesnická Dendrologie 1: Jehličnaté Dřeviny: Přehled Nahosemenných (i Výtrusných) Dřevin; Česká Zemědělská Univerzita: Praha, Czech Republic, 2002; pp. 79–98, 177. [Google Scholar]
- Kantor, P.E.A. Zakládání Lesů; Státní Zemědělské Nakladatelství: Praha, Czech Republic, 1965; p. 490. [Google Scholar]
- Dušek, V.; Kotyza, F. Moderní Lesní Školkařství; Státní Zemědělské Nakladatelství: Praha, Czech Republic, 1970; p. 480. [Google Scholar]
- Kupka, I. Reaction of Silver fir (Abies alba Mill.) plantation to fertilization. J. For. Sci. 2005, 51, 95–100. [Google Scholar] [CrossRef]
- Pesendorfer, M.B.; Bogdziewicz, M.; Borowski, Z.; Kantorowicz, W.; Espelta, J.M.; Fernández-Martínez, M. Investigating the relationship between climate, stand age, and temporal trends in masting behavior of European forest trees. Glob. Chang. Biol. 2020, 26, 1654–1667. [Google Scholar] [CrossRef] [PubMed]
- Stejskalová, J.; Kupka, I. Forest vegetation zones influence on seed quality of silver fir (Abies alba Mill.). In Proceedigs of Central European Silviculture—12th International Conference. Proceedings of Central European Silviculture 2011, Opočno, Czech Republic, 28–29 June 2011; Kacálek, D., Jurásek, A., Novák, J., Slodičák, M., Eds.; Forestry and Game Management Research Institute, Strnady—Opocno Research Station: Opočno, Czech Republic, 2011; pp. 235–242. [Google Scholar]
- Bezděčkova, L.; Řezníčková, J. Effect of pre-sowing treatment on the germination and emergence of silver fir seeds. Rep. For. Res. 2012, 57, 249–256. [Google Scholar]
- Skořepa, H. Jedle bělokorá v našich lesích. Živa 2006, 3, 105–110. [Google Scholar]
- Boncaldo, E.; Bruno, G.; Tommasi, F.; Mastropasqua, L. Germinability and fungal occurrence in seeds of Abies alba Mill. populations in southern Italy. Plant Biosyst. 2010, 144, 740–745. [Google Scholar] [CrossRef]
- Gradečki-Poštenjak, M.; Ćelepirović, N. The influence of crown defoliation on the variability of some physiological and morphological properties of silver fir (Abies alba) seeds in the seed zone of Dinaric beech-fir forests in Croatia. Period. Biol. 2016, 117, 479–492. [Google Scholar] [CrossRef]
- Kerr, G.; Stokes, V.; Peace, A.; Jinks, R. Effects of provenance on the survival, growth and stem form of European silver fir (Abies alba Mill.) in Britain. Eur. J. For. Res. 2015, 134, 349–363. [Google Scholar] [CrossRef]
- Gunia, S.; Łukaszewicz, J.; Szeligowski, H. The first Polish provenance experiments with silver fir Abies alba Mill. For. Res. Pap. 2019, 80, 201–212. [Google Scholar] [CrossRef]
- Stępniewska, H.; Rębisz, P. Mycorrhizae of fir (Abies alba Mill.) seedlings growing on sawdust-peat substrates of different periods of utilization. Sylwan 2004, 8, 34–42. [Google Scholar]
- Jaworski, A. Charakterystyka Hodowlana Drzew Leśnych; Gutenberg: Kraków, Poland, 1995; p. 237. [Google Scholar]
- Čater, M.; Diaci, J. Scientific support for close-to-nature forestry. In Forests and Forestry in Slovenia; Čater, M., Železnik, P., Eds.; Studia Forestalia Slovenica: Ljubljana, Slovenia, 2021; pp. 55–58. [Google Scholar]
- Čater, M.; Adamič, P.; Dařenová, E. Response of beech and fir to different light intensities along the Carpathian and Dinaric Mountains. Front. Plant Sci. 2024, 15, 1380275. [Google Scholar] [CrossRef] [PubMed]
- Čater, M.; Levanič, T. Beech and silver fir’s response along the Balkan’s latitudinal gradient. Sci. Rep. 2019, 9, 16269. [Google Scholar] [CrossRef] [PubMed]
- Adamič, P.C.; Levanic, T.; Hanzu, M.; Čater, M. Growth response of European beech (Fagus sylvatica L.) and silver fir (Abies alba Mill.) to climate factors along the Carpathian massive. Forests 2023, 14, 1318. [Google Scholar] [CrossRef]
- Szymura, T.H. Silver fir sapling bank in seminatural stand: Individuals architecture and vitality. For. Ecol. Manag. 2005, 212, 101–108. [Google Scholar] [CrossRef]
- Kantor, P. Obnova jedle bělokoré. In Pěstování a Umělá Obnova Jedle Bělokoré. Proceedings of the Pěstování a Umělá Obnova Jedle Bělokoré, Chudobín u Litovele, Czech Republic, 28 August 2001; Kotrla, K., Kyslík, P., Eds.; Česká lesnická společnost: Praha, Czech Republic, 2001; pp. 5–13. [Google Scholar]
- Čater, M. Response and mortality of beech, fir, spruce and sycamore to rapid light exposure after large-scale disturbance. For. Ecol. Manag. 2021, 498, 119554. [Google Scholar] [CrossRef]
- Backman, G. Wachstum und Organische Zeit; J. A. Barth: Leipzig, Germany, 1943. [Google Scholar]
- Bezačinský, H. Problém Odumierania Jedle na Slovensku z Pestovateľského Hľadiska; VŠLD: Zvolen, Slovakia, 1962; pp. 87–102. [Google Scholar]
- Vinš, B. Příspěvek k výzkumu proměnlivosti jedle (Abies alba Mill.). Rozpravy Čs. Akademie věd řada mat. a přír. věd 1966, 76, 1–82. [Google Scholar]
- Zakopal, V. Studie u nás vytvořených tvarů výběrného lesa. Lesnictví 1959, 5, 995–1012. [Google Scholar]
- Robakowski, P.; Bielinis, E. Needle age dependence of photosynthesis along a light gradient within an Abies alba crown. Acta Physiol. Plant. 2017, 39, 83. [Google Scholar] [CrossRef]
- Jarzyna, K. Climatic hazards for native tree species in Poland with special regards to silver fir (Abies alba Mill.) and European beech (Fagus sylvatica L.). Theor. Appl. Climatol. 2021, 144, 581–591. [Google Scholar] [CrossRef]
- Piedallu, C.; Dallery, D.; Bresson, C.; Legay, M.; Gégout, J.C.; Pierrat, R. Spatial vulnerability assessment of silver fir and Norway spruce dieback driven by climate warming. Landsc. Ecol. 2023, 38, 341–361. [Google Scholar] [CrossRef]
- Zakopal, V. Struktura porostů jako základní příčina hynutí jedle bělokoré (Abies alba Mill.) v našich lesích. Lesnictví 1978, 24, 731–754. [Google Scholar]
- Sokol, A. Jedľové porasty. In Pěstění lesů III; Polanský, D., Jurča, J., Zachar, D., Bezačinský, H., Korpel, Š., Zlatník, A., Pelíšek, J., Sokol, A., Čížek, J., Eds.; SZN: Praha, Czech Republic, 1956; pp. 439–446. [Google Scholar]
- Kadlus, Z. Obnova jedle v hospodářských porostech smrku a borovice na stanovištích jedlových doubrav. Práce VÚLHM 1970, 39, 79–102. [Google Scholar]
- Průša, E. Die Böhmischen und Mährischen Urwälder, Ihre Struktur und Ökologie; Academia: Praha, Czech Republic, 1985; p. 578. [Google Scholar]
- Vrška, T.; Adam, D.; Hort, L.; Kolář, T.; Janík, D. European beech (Fagus sylvatica L.) and silver fir (Abies alba Mill.) rotation in the Carpathians—A developmental cycle or a linear trend induced by man? For. Ecol. Manag. 2009, 258, 347–356. [Google Scholar] [CrossRef]
- Klopčič, M.; Bončina, A. A century long dynamics of silver fir population in mixed silver fir-European beech forests. Zb. Gozdarstva Lesar. 2012, 97, 43–54. [Google Scholar]
- Polách, R.; Špulák, O. Prosperity of Silver fir planted under preparatory stands of pioneer broadleaves of different śtocking and age. Rep. For. Res. 2022, 64, 269–277. [Google Scholar]
- Bončina, A. History, current status and future prospects of uneven-aged forest management in the Dinaric region: An overview. Forestry 2011, 84, 467–478. [Google Scholar] [CrossRef]
- Kerr, G. The management of silver fir forests: De Liocourt (1898) revisited. Forestry 2014, 87, 29–38. [Google Scholar] [CrossRef]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.-J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü. Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: Past stress history, stress interactions, tolerance and acclimation. For. Ecol. Manag. 2010, 260, 1623–1639. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).