
Mapping Characteristics in Vaccinium uliginosum Populations Predicted Using Filtered Machine Learning Modeling

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
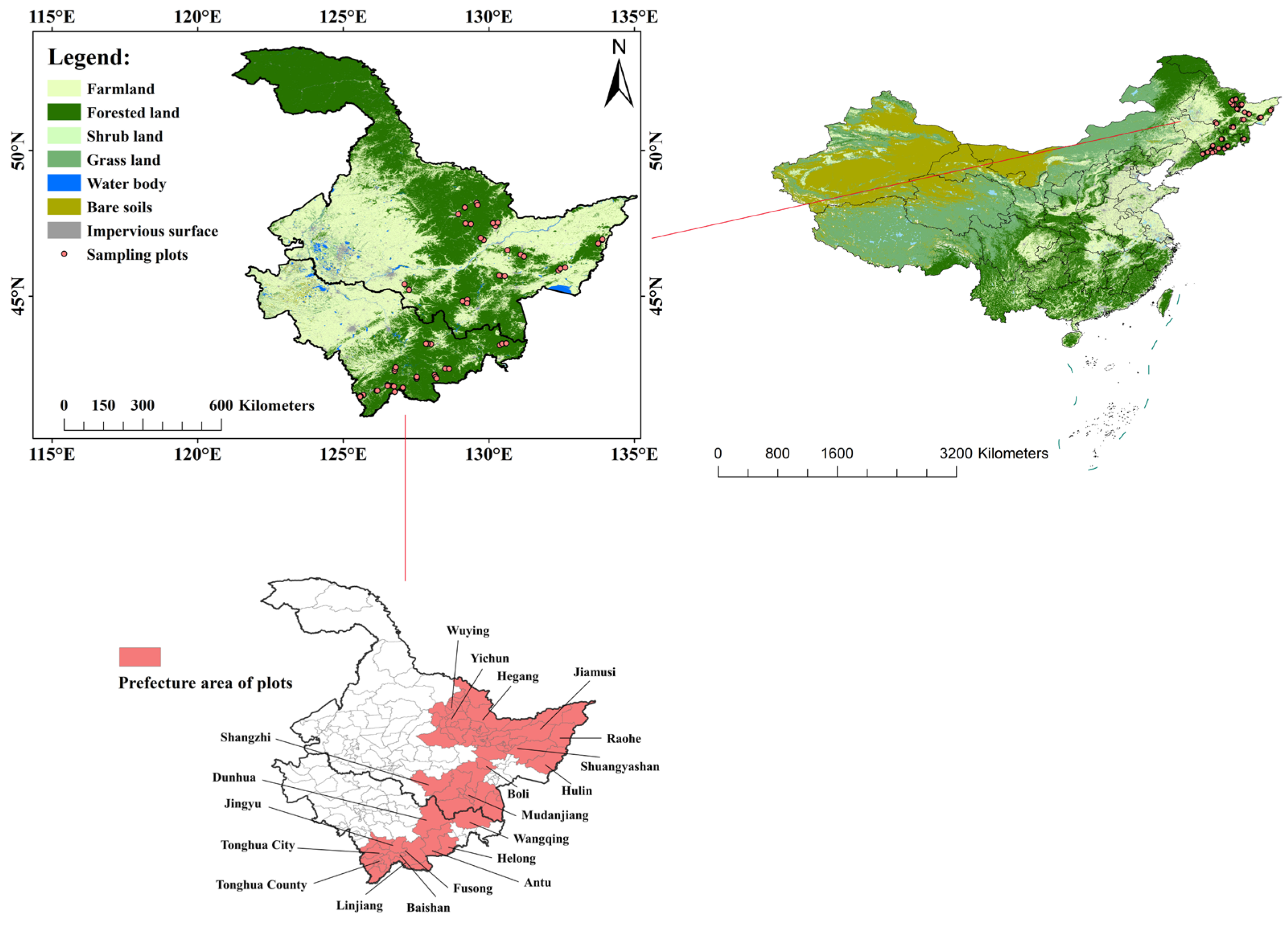
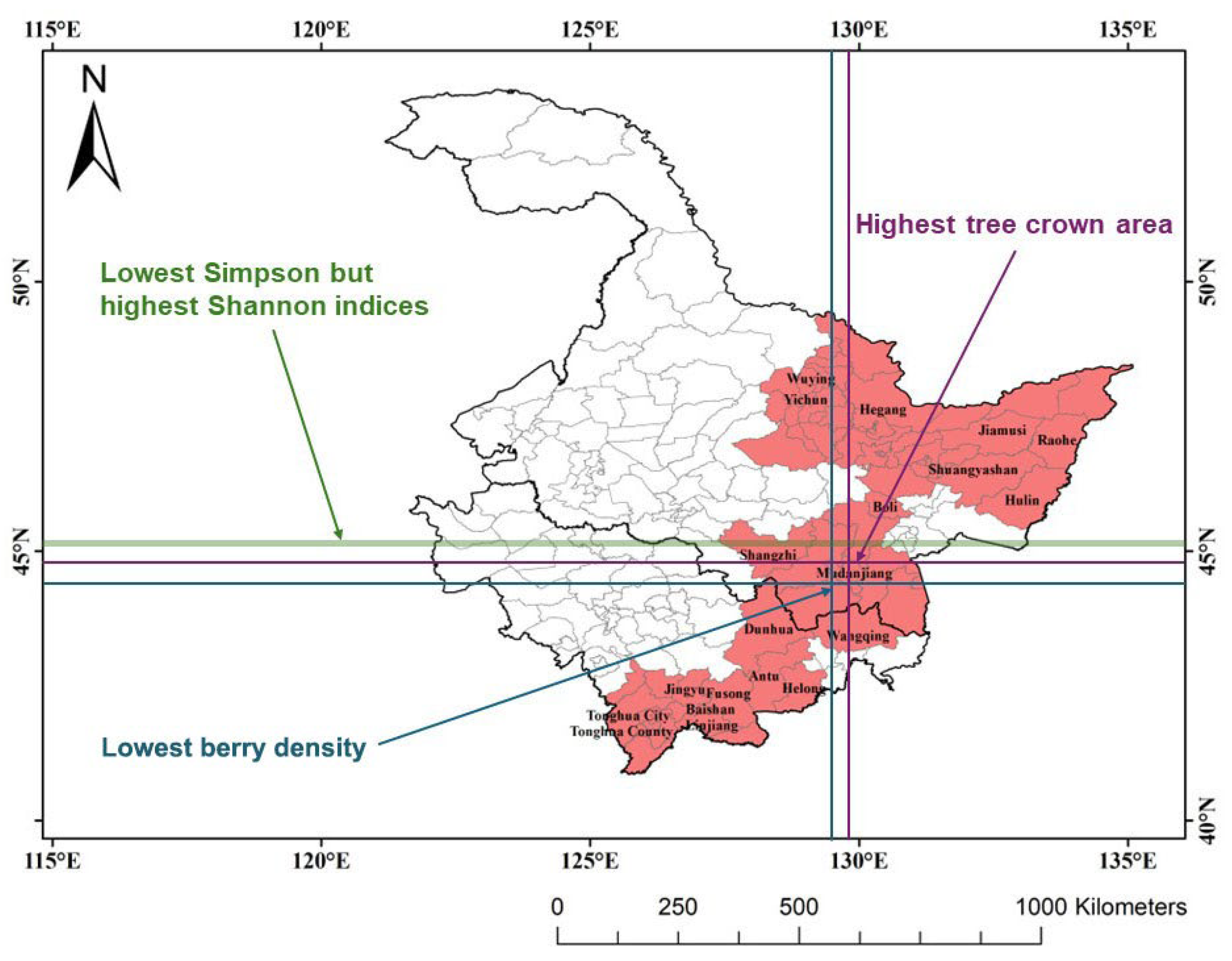
2.1. Study Area
2.2. Field Investigation
2.3. Variable Calculation
2.4. Statistical Analysis and Mapping Process
3. Results
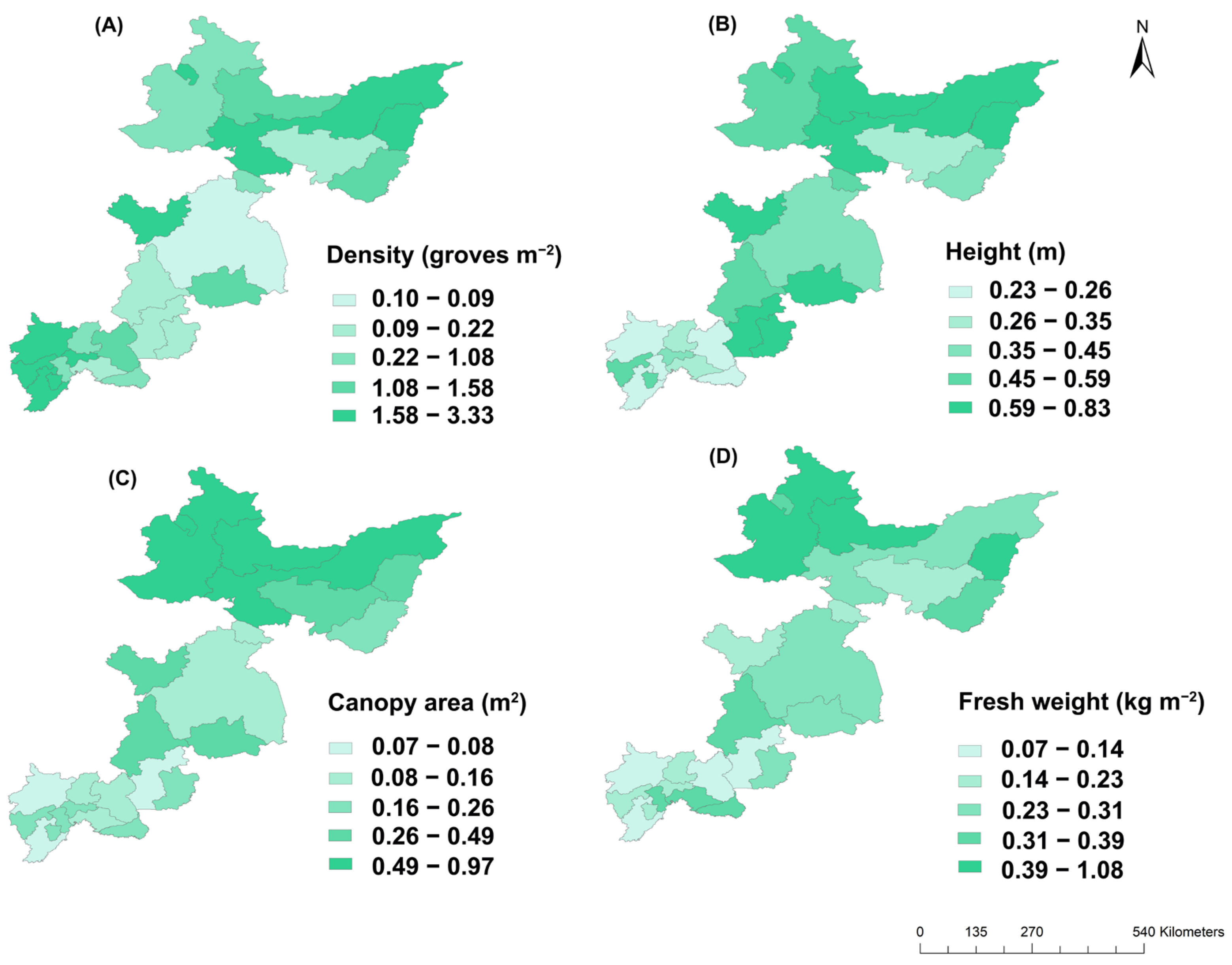
3.1. Spatial Distributions of Bog Bilberry Variables
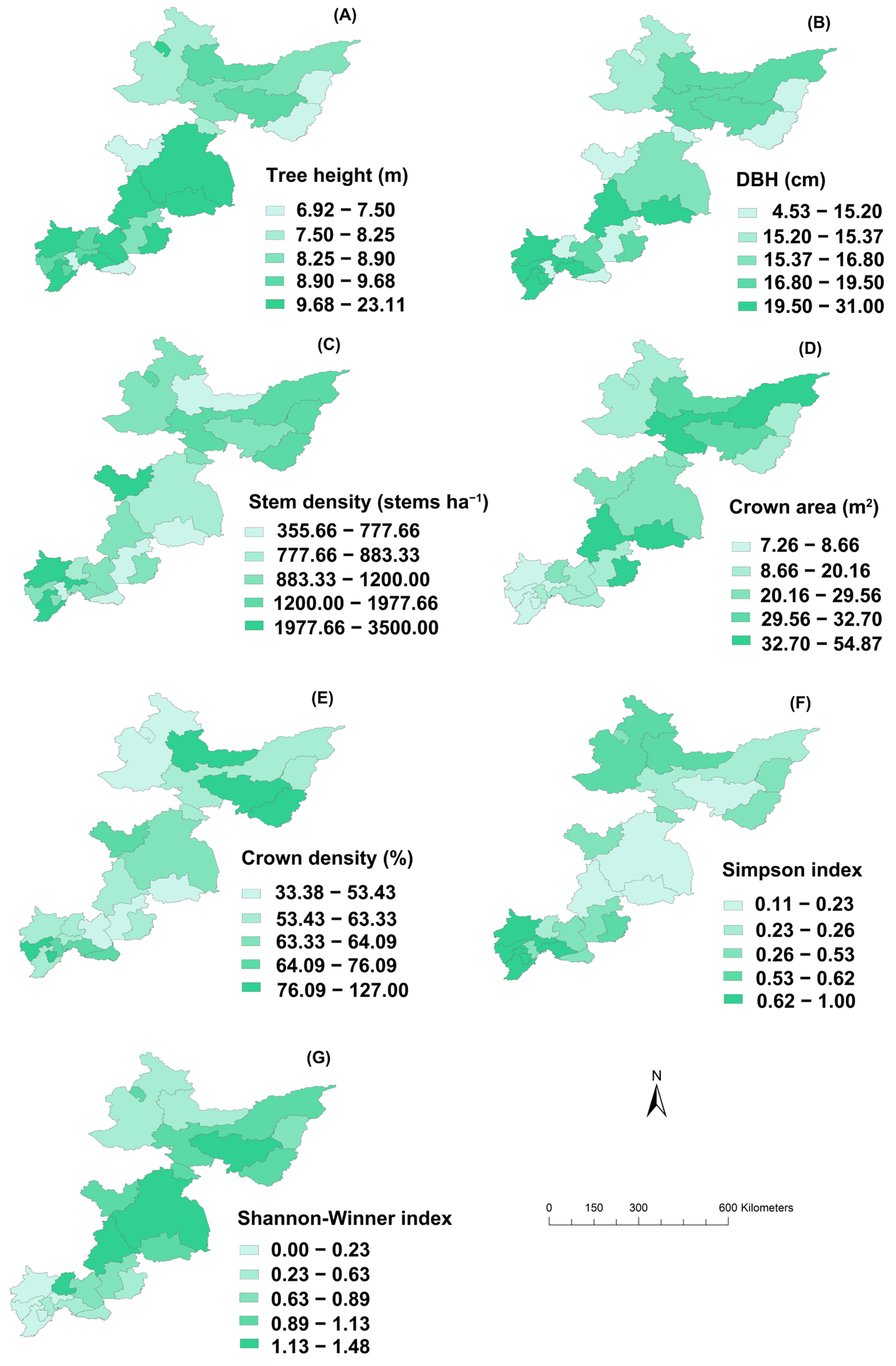
3.2. Spatial Distributions of Forest Stand Attributes
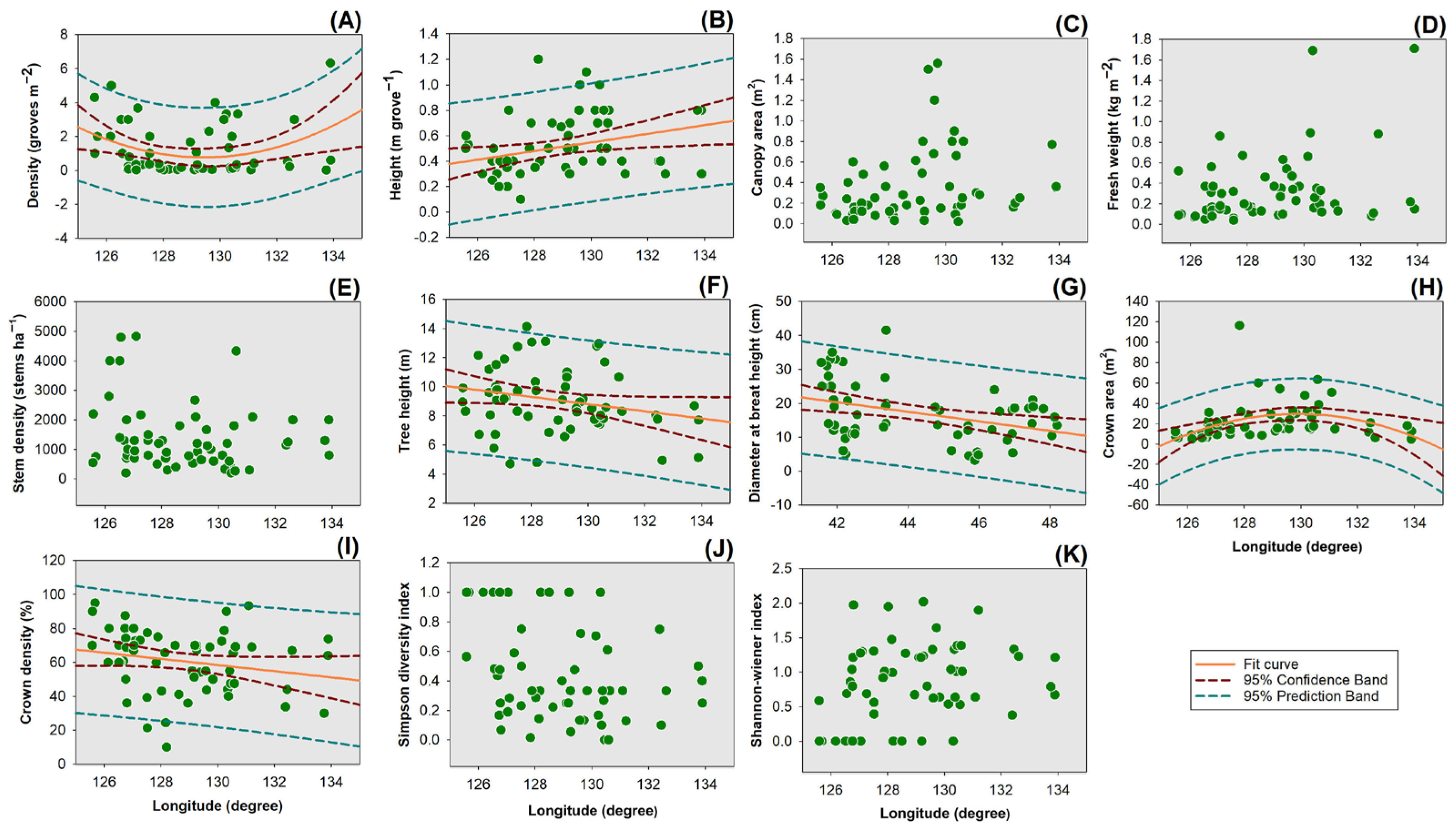
3.3. Relationships between Bog Bilberry Variables and Stand Latitude
3.4. Relationships between Tree Variables and Stand Longitude
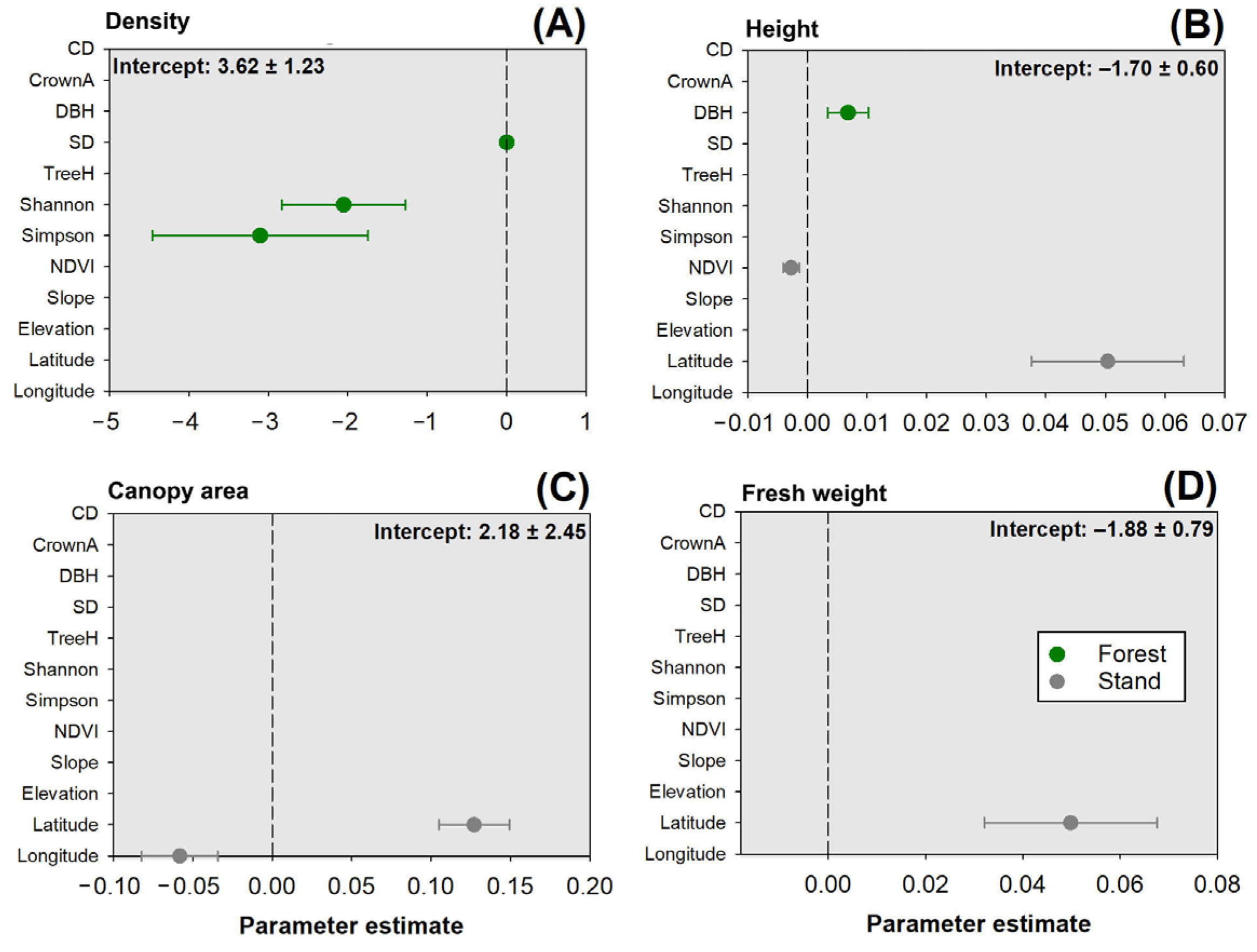
3.5. Multivariate Linear Regression Analysis
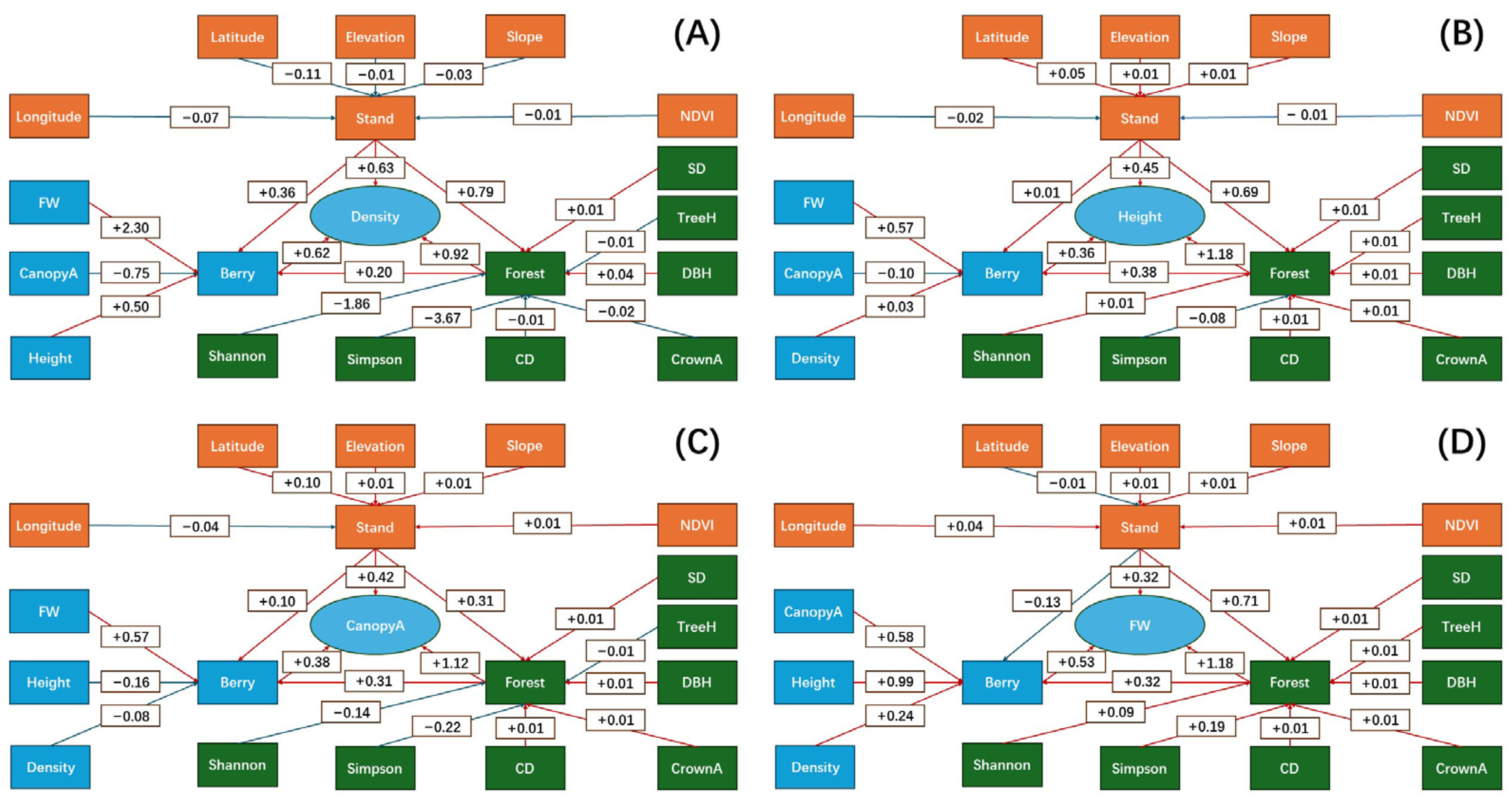
3.6. Structural Equation Model Analysis
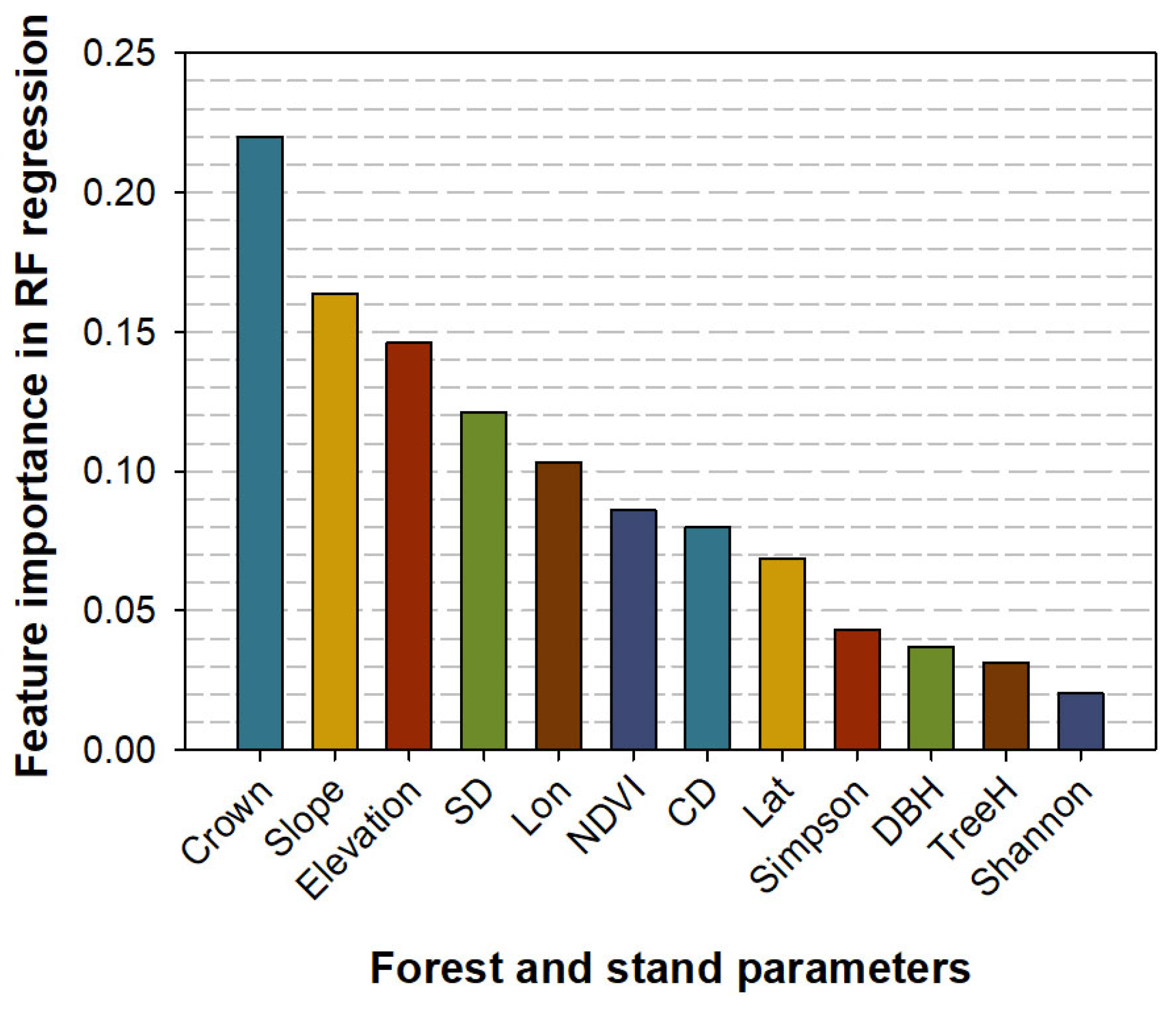
3.7. Machine Learning Model Regression
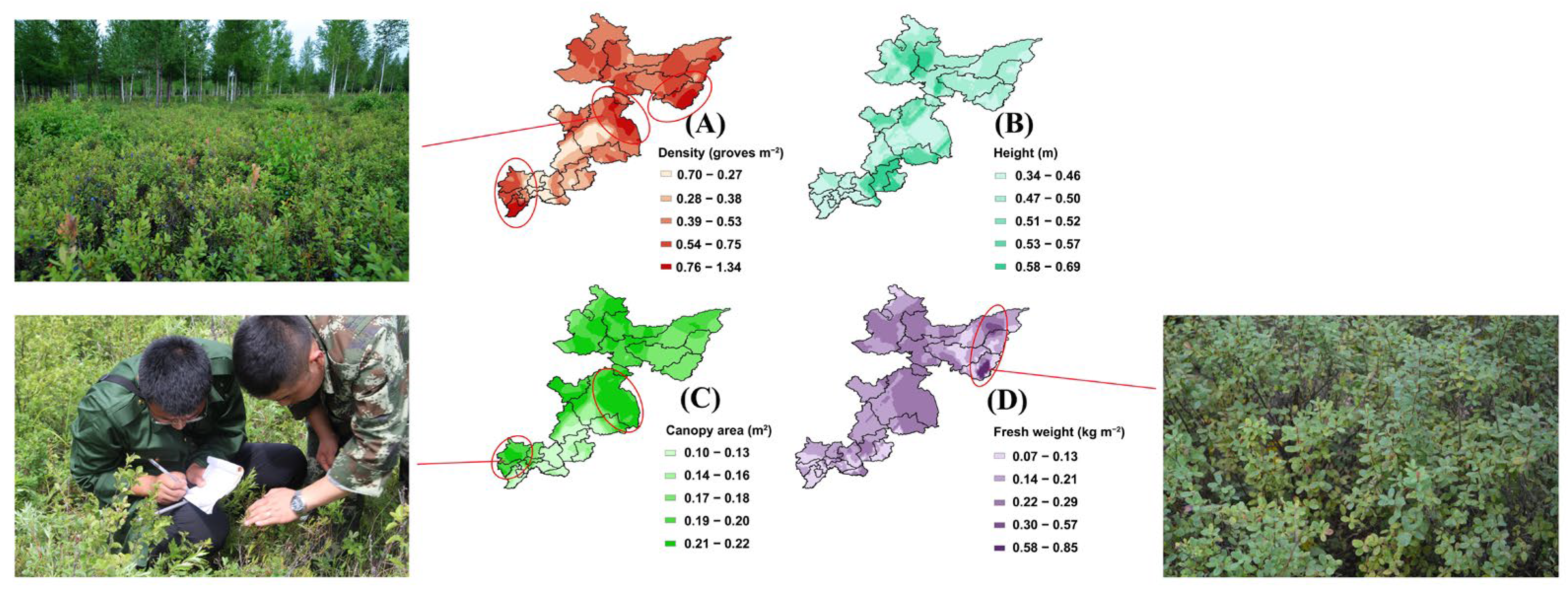
3.8. Spatial Distributions of Predicted Bog Bilberry Population Characteristics
4. Discussion
4.1. Geographical Distributions of Bog Bilberry Population Characteristics
4.2. Geographical Changes in Host Forest Structures
4.3. Spatial Distribution of Population Characteristics Predicted by the Machine Learning Model
4.4. Limitations of the Present Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Latin American and Caribbean Forestry Commission—33rd Session; LACFC Secretariat; FAO: Rome, Italy, 2023. [Google Scholar]
- United Nations. The Sustainable Development Goals Report 2022; United Nations: New York, NY, USA, 2022. [Google Scholar]
- Duan, Y.D.; Guo, B.T.; Zhang, L.; Li, J.X.; Li, S.; Zhao, W.B.; Yang, G.; Zhou, S.; Zhou, C.; Song, P.; et al. Interactive climate-soil forces shape the spatial distribution of foliar N:P stoichiometry in Vaccinium uliginosum planted in agroforests of Northeast China. Front. Ecol. Evol. 2022, 10, 1065680. [Google Scholar] [CrossRef]
- Piras, F.; Santoro, A. Land use changes in globally important cultural forests. The case of two traditionally managed forests for non-wood forest products (NWFPs) in China and Japan. Biodivers. Conserv. 2023. [Google Scholar] [CrossRef]
- Wei, H.X.; Chen, G.S.; Chen, X.; Zhao, H.T. Geographical distribution of Aralia elata characteristics correlated with topography and forest structure in Heilongjiang and Jilin Provinces, Northeast China. J. For. Res. 2021, 32, 1115–1125. [Google Scholar] [CrossRef]
- Wei, H.X.; Chen, X.; Chen, G.S.; Zhao, H.T. Foliar nutrient and carbohydrate in Aralia elata can be modified by understory light quality in forests with different structures at Northeast China. Ann. For. Res. 2019, 62, 125–137. [Google Scholar] [CrossRef]
- Guo, S.L.; Wei, H.X.; Li, J.P.; Fan, R.F.; Xu, M.Y.; Chen, X.; Wang, Z.Y. Geographical Distribution and Environmental Correlates of Eleutherosides and Isofraxidin in Eleutherococcus senticosus from Natural Populations in Forests at Northeast China. Forests 2019, 10, 872. [Google Scholar] [CrossRef]
- Sheppard, J.P.; Chamberlain, J.; Agundez, D.; Bhattacharya, P.; Chirwa, P.W.; Gontcharov, A.; Sagona, W.C.J.; Shen, H.-L.; Tadesse, W.; Mutke, S. Sustainable Forest Management Beyond the Timber-Oriented Status Quo: Transitioning to Co-production of Timber and Non-wood Forest Products-a Global Perspective. Curr. For. Rep. 2020, 6, 26–40. [Google Scholar] [CrossRef]
- Spies, T.A.; Long, J.W.; Charnley, S.; Hessburg, P.F.; Marcot, B.G.; Reeves, G.H.; Lesmeister, D.B.; Reilly, M.J.; Cerveny, L.K.; Stine, P.A.; et al. Twenty-five years of the Northwest Forest Plan: What have we learned? Front. Ecol. Environ. 2019, 17, 511–520. [Google Scholar] [CrossRef]
- Ghanbari, S.; Vaezin, S.M.H.; Shamekhi, T.; Eastin, I.L.; Lovric, N.; Aghai, M.M. The Economic and Biological Benefits of Non-wood Forest Products to Local Communities in Iran. Econ. Bot. 2020, 74, 59–73. [Google Scholar] [CrossRef]
- Qu, H.C.; Xiang, R.; Obsie, E.Y.; Wei, D.W.; Drummond, F. Parameterization and Calibration of Wild Blueberry Machine Learning Models to Predict Fruit-Set in the Northeast China Bog Blueberry Agroecosystem. Agronomy 2021, 11, 1736. [Google Scholar] [CrossRef]
- Song, Y.Y.; Song, C.C.; Ren, J.S.; Tan, W.W.; Jin, S.F.; Jiang, L. Influence of nitrogen additions on litter decomposition, nutrient dynamics, and enzymatic activity of two plant species in a peatland in Northeast China. Sci. Total Environ. 2018, 625, 640–646. [Google Scholar] [CrossRef]
- Jiang, J.; Wei, J.; Yu, H.; He, S. The Developing Blueberry Industry in China. In The Developing Blueberry Industry in China; IntechOpen: London, UK, 2019. [Google Scholar]
- Duan, Y.; Fu, X.; Zhou, X.; Gao, D.; Zhang, L.; Wu, F. Removal of Dominant Species Impairs Nitrogen Utilization in Co-Existing Ledum palustre and Vaccinium uliginosum Communities Subjected to Five-Year Continuous Interruptions. Agronomy 2022, 12, 932. [Google Scholar] [CrossRef]
- Li, Q.; Qi, Y.; Wang, Q.; Wang, D. Prediction of the Potential Distribution of Vaccinium uliginosum in China Based on the Maxent Niche Model. Horticulturae 2022, 8, 1202. [Google Scholar] [CrossRef]
- Tumaneng-Diete, T.; Ferguson, I.S.; MacLaren, D. Log export restrictions and trade policies in the Philippines: Bane or blessing to sustainable forest management? For. Policy Econ. 2005, 7, 187–198. [Google Scholar] [CrossRef]
- Liu, K.; Liang, Y.; He, H.S.; Wang, W.J.; Huang, C.; Zong, S.W.; Wang, L.; Xiao, J.; Du, H. Long-Term Impacts of China’s New Commercial Harvest Exclusion Policy on Ecosystem Services and Biodiversity in the Temperate Forests of Northeast China. Sustainability 2018, 10, 1071. [Google Scholar] [CrossRef]
- Zhao, Z.F.; Guo, Y.L.; Zhu, F.X.; Jiang, Y. Prediction of the impact of climate change on fast-growing timber trees in China. For. Ecol. Manag. 2021, 501, 119653. [Google Scholar] [CrossRef]
- Geng, Y.D.; Sun, S.B.; Yeo-Chang, Y. Impact of Forest Logging Ban on the Welfare of Local Communities in Northeast China. Forests 2021, 12, 3. [Google Scholar] [CrossRef]
- Alldredge, M.W.; Peek, J.M.; Wall, W.A. Shrub community development and annual productivity trends over a 100-year period on an industrial forest of Northern Idaho. For. Ecol. Manag. 2001, 152, 259–273. [Google Scholar] [CrossRef]
- Bona, A.; Brzezinski, D.; Jadwiszczak, K.A. Genetic Diversity and Fine-Scale Spatial Genetic Structure of the Endangered Shrub Birch (Betula humilis Schrk.) Populations in Protected and Unprotected Areas. Diversity 2022, 14, 684. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiang, W.; Gou, M.; Chen, L.; Lei, P.; Xiao, W.; Deng, X.; Zeng, L.; Li, J.; Zhang, T.; et al. Stability in subtropical forests: The role of tree species diversity, stand structure, environmental and socio-economic conditions. Glob. Ecol. Biogeogr. 2021, 30, 500–513. [Google Scholar] [CrossRef]
- Guo, Z.W.; Wang, X.P.; Fan, D.Y. Ecosystem functioning and stability are mainly driven by stand structural attributes and biodiversity, respectively, in a tropical forest in Southwestern China. For. Ecol. Manag. 2021, 481, 118696. [Google Scholar] [CrossRef]
- Wang, T.; Dong, L.B.; Liu, Z.G. Stand structure is more important for forest productivity stability than tree, understory plant and soil biota species diversity. Front. For. Glob. Change 2024, 7, 1354508. [Google Scholar] [CrossRef]
- Woziwoda, B.; Dyderski, M.K.; Jagodzinski, A.M. Effects of land use change and Quercus rubra introduction on Vaccinium myrtillus performance in Pinus sylvestris forests. For. Ecol. Manag. 2019, 440, 1–11. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT® 14.3 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2017. [Google Scholar]
- Topic, V.; Butorac, L.; Jelic, G.; Peric, S.; Rosavec, R. Biomass of hop hornbeam (Ostrya carpinifolia Scop.) shrub on Velebit. Period. Biol. 2008, 110, 151–156. [Google Scholar]
- Dangeti, P. Statistics for Machine Learning; Packt Publishing Ltd.: Birmingham, UK, 2017. [Google Scholar]
- Ganglo, J.C. Ecological niche model transferability of the white star apple (Chrysophyllum albidum G. Don) in the context of climate and global changes. Sci. Rep. 2023, 13, 2430. [Google Scholar] [CrossRef]
- Wang, X.; Ouyang, S.; Sun, O.J.; Fang, J. Forest biomass patterns across northeast China are strongly shaped by forest height. For. Ecol. Manag. 2013, 293, 149–160. [Google Scholar] [CrossRef]
- Dong, L.B.; Wei, H.Y.; Liu, Z.G. Optimizing Forest Spatial Structure with Neighborhood-Based Indices: Four Case Studies from Northeast China. Forests 2020, 11, 413. [Google Scholar] [CrossRef]
- Wang, Y.G.; Sun, X.Y.; Li, S.Y.; Wei, B. Lignin and Cellulose Contents in Chinese Red Pine (Pinus tabuliformis Carr.) Plantations Varied in Stand Structure, Soil Property, and Regional Climate. Forests 2024, 15, 240. [Google Scholar] [CrossRef]
- NASA EarthData. NASA EarthData. Available online: https://search.earthdata.nasa.gov/search/?ac=true&m=0.0703125!0!2!1!0!0%2C2 (accessed on 3 July 2024).
- Onaindia, M.; Dominguez, I.; Albizu, I.; Garbisu, C.; Amezaga, I. Vegetation diversity and vertical structure as indicators of forest disturbance. For. Ecol. Manag. 2004, 195, 341–354. [Google Scholar] [CrossRef]
- Sharma, A.; Cory, B.; McKeithen, J.; Frazier, J. Structural diversity of the longleaf pine ecosystem. For. Ecol. Manag. 2020, 462, 117987. [Google Scholar] [CrossRef]
- Ha, T.N.; Lubo-Robles, D.; Marfurt, K.J.; Wallet, B.C. An in-depth analysis of logarithmic data transformation and per-class normalization in machine learning: Application to unsupervised classification of a turbidite system in the Canterbury Basin, New Zealand, and supervised classification of salt in the Eugene Island minibasin, Gulf of Mexico. Interpretation 2021, 9, T685–T710. [Google Scholar] [CrossRef]
- Branco, P.; Torgo, L.; Ribeiro, R.P. SMOGN: A Pre-processing Approach for Imbalanced Regression. In Proceedings of the First International Workshop on Learning with Imbalanced Domains: Theory and Applications, Skopje, Macedonia, 22 September 2017; pp. 36–50. [Google Scholar]
- Kudo, G.; Suzuki, S. Warming effects on growth, production, and vegetation structure of alpine shrubs: A five-year experiment in northern Japan. Oecologia 2003, 135, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.S.; Hu, Y.K.; Wang, J.; Liu, D.D.; Lin, Y.H.; Liu, G.; Zhang, Y.H.; Zhou, Z.Q. Diversity of Understory Communities in Boreal Forests: Influences of Forest Type, Latitude, and Spatial Scale. Forests 2019, 10, 1003. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, H.B.; Zhong, S.; Liu, X.; Li, T.; Zong, C.W. Variations in Sugar and Organic Acid Content of Fruit Harvested from Different Vaccinium uliginosum Populations in the Changbai Mountains of China. J. Am. Soc. Hortic. Sci. 2019, 144, 420–428. [Google Scholar] [CrossRef]
- Woods, C.L.; Cardelús, C.L.; DeWalt, S.J. Microhabitat associations of vascular epiphytes in a wet tropical forest canopy. J. Ecol. 2015, 103, 421–430. [Google Scholar] [CrossRef]
- Wang, G.; Gertner, G.; Xiao, X.; Wente, S.; Anderson, A.B. Appropriate plot size and spatial resolution for mapping multiple vegetation types. Photogramm. Eng. Remote Sens. 2001, 67, 575–584. [Google Scholar]
- Jones, M.O.; Allred, B.W.; Naugle, D.E.; Maestas, J.D.; Donnelly, P.; Metz, L.J.; Karl, J.; Smith, R.; Bestelmeyer, B.; Boyd, C.; et al. Innovation in rangeland monitoring: Annual, 30 m, plant functional type percent cover maps for U.S. rangelands, 1984–2017. Ecosphere 2018, 9, e02430. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dependent Variables | Fit Curve Model | R2 | p | b | a | y0 |
|---|---|---|---|---|---|---|
| Bog bilberry | ||||||
| Density | Polynomial quadratic 1 | 0.3509 | 0.0133 | −12.75 | 0.14 | 282.85 |
| Height | Linear 2 | 0.2126 | 0.0039 | - | 0.05 | −1.61 |
| Canopy area | Linear | 0.3505 | <0.0001 | - | 0.09 | −3.52 |
| Fresh weight | Linear | 0.3452 | 0.0203 | - | 0.05 | −1.88 |
| Tree | ||||||
| Height | Linear | 0.0820 | 0.0002 | - | −0.27 | 21.25 |
| DBH | Linear | 0.1408 | 0.0002 | - | −1.41 | 79.37 |
| Crown area | Polynomial quadratic | 0.3526 | 0.0134 | 156.49 | −1.74 | −3488.23 |
| Simpson index | Polynomial quadratic | 0.2813 | 0.0001 | −4.14 | 0.05 | 93.40 |
| Shannon index | Polynomial quadratic | 0.2655 | 0.0002 | 7.10 | −0.08 | −158.63 |
| Dependent Variables | Fit Curve Model | R2 | p | b | a | y0 |
|---|---|---|---|---|---|---|
| Bog bilberry | ||||||
| Density | Polynomial quadratic 1 | 0.3204 | 0.0133 | −23.46 | 0.09 | 1518.87 |
| Height | Linear 2 | 0.3040 | 0.0359 | - | 0.03 | −3.86 |
| Tree | ||||||
| Height | Linear | 0.2410 | 0.0182 | - | −0.25 | 41.10 |
| DBH | Linear | 0.4056 | 0.0006 | - | −1.64 | 227.76 |
| Crown area | Polynomial quadratic | 0.4069 | 0.0024 | 346.52 | −1.33 | −22,475.46 |
| Crown density | Linear | 0.2121 | 0.0419 | - | −1.82 | 295.02 |
| Berry Variables | RMSE 1 | MSE 2 | MAE 3 | R2 4 |
|---|---|---|---|---|
| Density | 0.045322 | 0.002054 | 0.02321 | 0.585016 |
| Height | 0.014539 | 0.000211 | 0.007961 | 0.730527 |
| Canopy | 0.012955 | 0.000168 | 0.006967 | 0.786526 |
| Fresh weight | 0.017353 | 0.000301 | 0.007202 | 0.636299 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, Y.; Wei, X.; Wang, N.; Zang, D.; Zhao, W.; Yang, Y.; Wang, X.; Xu, Y.; Zhang, X.; Liu, C. Mapping Characteristics in Vaccinium uliginosum Populations Predicted Using Filtered Machine Learning Modeling. Forests 2024, 15, 1252. https://doi.org/10.3390/f15071252
Duan Y, Wei X, Wang N, Zang D, Zhao W, Yang Y, Wang X, Xu Y, Zhang X, Liu C. Mapping Characteristics in Vaccinium uliginosum Populations Predicted Using Filtered Machine Learning Modeling. Forests. 2024; 15(7):1252. https://doi.org/10.3390/f15071252
Chicago/Turabian StyleDuan, Yadong, Xin Wei, Ning Wang, Dandan Zang, Wenbo Zhao, Yuchun Yang, Xingdong Wang, Yige Xu, Xiaoyan Zhang, and Cheng Liu. 2024. "Mapping Characteristics in Vaccinium uliginosum Populations Predicted Using Filtered Machine Learning Modeling" Forests 15, no. 7: 1252. https://doi.org/10.3390/f15071252