
The Expanding Thread of Ungulate Browsing—A Review of Forest Ecosystem Effects and Management Approaches in Europe
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results

{kind=link}
{kind=link}
| No. | Country | Ungulate Species | Browsing Proportion (%) | Ungulate Density (ind./100 ha) | Reference |
|---|---|---|---|---|---|
| Europe | |||||
| Eastern Europe | |||||
| 1 | Czech Republic | Roe deer, red deer, Fallow deer | 5–85 | 0.8–56 | [32,33,34,35,36,37,38,39,40] |
| 2 | Poland | Roe deer, red deer, Chamois, Moose | 12.8–58 | 0.5–10.4 | [41,42,43,44,45,46,47,48] |
| Bison | n.m. * | 0.7 | [41,44,45,46] | ||
| Moose | n.m. * | 0.31–0.5 | [28] | ||
| 3 | Romania | Red deer, Roe deer | 3–88 | 1.8–8.6 | [25,49,50,51,52,53,54,55,56] |
| 4 | Russia | Moose | 79.5–80 | 1–5 | [57] |
| 5 | Slovakia | Roe deer, red deer | n.m. * | 21 | [58,59] |
| Northern Europe | |||||
| 6 | Denmark | Roe deer | 49–57 | 0.8–9.8 | [60] |
| 7 | Finland | Roe deer | 51 | 4–4.5 | [61] |
| Moose | 8–75 | 0.4–3 | [61,62,63,64,65,66] | ||
| 8 | Latvia | Red deer, roe deer | 10–12 | n.m.* | [67] |
| 11 | Sweden | Roe deer, red deer, fallow deer | 5–85 | 1–20 (32.7) | [68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85] |
| Moose | 4–84 | 0.45–2.4 | [68,69,72,76,77,78,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102] | ||
| 12 | UK | Red deer, roe deer, fallow deer, Muntjac deer | 5–81 (96) | 4–60 (75) | [103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125] |
| Southern Europe | |||||
| 13 | Italy | Roe deer, red deer, fallow deer, chamois, | 7–61 | 4.5–25 | [126,127,128,129,130,131,132,133] |
| 14 | Slovenia | Red deer, roe deer | 8.2 | 1–12 | [134,135] |
| 16 | Portugal | Red deer | 10 | 10 | [136] |
| Western Europe | |||||
| 17 | Austria | Roe deer, red deer, chamois | 5–67 | 3.6–25 (46) | [137,138,139,140,141,142,143,144] |
| 18 | France | Roe deer, red deer, chamois | 7–100 | 10–15 | [145,146,147,148,149,150,151,152,153] |
| 19 | Germany | Roe deer, red deer, fallow deer, Chamois, | 5–79 | 5.3–20 | [154,155,156,157,158,159,160,161,162,163,164,165,166,167] |
| 20 | Netherlands | Red deer, roe deer | 6–85 | 4–14 | [168,169] |
| 21 | Switzerland | Roe deer, red deer, chamois | 3.3–67.5 (92.5) | 10–27.5 | [30,170,171,172,173,174,175,176] |
3.1. Ungulate Browsing in Europe
3.1.1. Ungulate Browsing in Eastern Europe
3.1.2. Ungulate Browsing in Northern Europe
3.1.3. Ungulate Browsing in Southern Europe
3.1.4. Ungulate Browsing in Western Europe
3.2. Browsing Trends in Europe
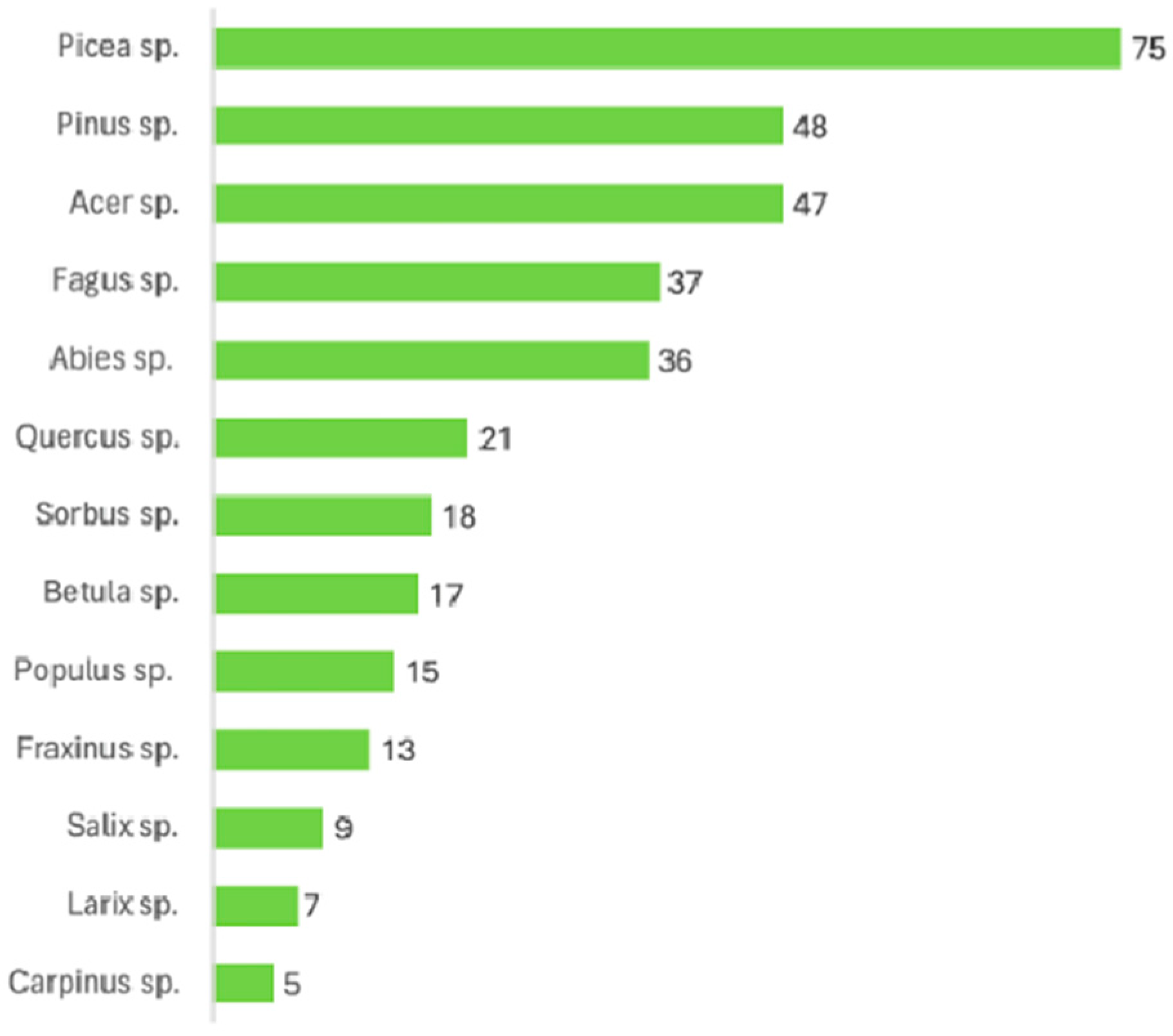
3.3. Frequency of Tree Species in the Literature Reviewed
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sinclair, A.; Fryxell, J.; Caughley, G. Wildlife Ecology, Conservation, and Management, 2nd ed.; Blackwell: Oxford, UK, 2006. [Google Scholar]
- Mahoney, S.P.; Jackson, J.J. Enshrining hunting as a foundation for conservation—The North American Model. Int. J. Environ. Stud. 2013, 70, 448–459. [Google Scholar] [CrossRef]
- Fernandez-Gil, A.; Cadete da Rocha Pereira, D.; Di Silvestre, I.; Dias Ferreira Pinto, S.M. Large Carnivore Management Plans of Protection: Best Practices in EU Member States; European Union: Brussels, Belgium, 2018. [Google Scholar]
- Breitenmoser, U. Large predators in the alps: The fall and rise of man’s competitors. Biol. Conserv. 1998, 83, 279–289. [Google Scholar] [CrossRef]
- Kratochvil, J. History of the distribution of the lynx in Europe, 4th ed.; Acta Scientiarum Naturalium Academiae Scientiarum Bohemicae—Brno; Czechoslovak Academy of Sciences: Prague, Czech Republic, 1968; Volume 2, pp. 1–50. [Google Scholar]
- Jakubiec, Z. Handbuch der Saugetiere Europas; AULA-Verlag: Wiesbaden, Germany, 1993; pp. 254–300. [Google Scholar]
- Zimen, E. Der Wolf—Mythos und Verhalten; Meyster: Munchen, Germany, 1978. [Google Scholar]
- Chapron, G.; Kaczensky, P.; Linnell, J.D.C.; Von Arx, M.; Huber, D.; Andrén, H.; López-Bao, J.V.; Adamec, M.; Álvares, F.; Anders, O.; et al. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 2014, 346, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Côté, S.D.; Rooney, T.P.; Tremblay, J.-P.; Dussault, C.; Waller, D.M. Ecological Impacts of Deer Overabundance. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 113–147. [Google Scholar] [CrossRef]
- Ripple, W.J.; Beschta, R.L. Large Predators Limit Herbivore Densities in Northern Forest Ecosystems. Eur. J. Wildl. Res. Springer, Heidelberg 2012, 58, 733–742. [Google Scholar] [CrossRef]
- Ripple, W.J.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Berger, J.; Berger, J.; Berger, J.; Berger, J.; et al. Status and ecological effects of the world’s largest carnivores. Science 2014, 342, 1241484. [Google Scholar] [CrossRef] [PubMed]
- Anthony, R.G.; Fisher, A.R. Wildlife Damage in Orchards: A Need for Better. Wildl. Soc. Bull. 1973, 5, 107–112. [Google Scholar]
- Apollonio, M.; Andersen, R.; Putman, R. European Ungulates and Their Management in the 21st Century; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Putman, R.; Apollonio, M.; Andersen, R. Ungulate Management in Europe: Problems and Practices; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Putman, R.; Apollonio, M.; Andersen, R.; Reimoser, F. Impacts of wild ungulate on vegetation: Cost and benefits. In Ungulate Management in Europe: Problems and Practices; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Szemethy, L.; Mátrai, K.; Bíró, Z.; Katona, K. Seasonal home range shift of red deer in a forest-agriculture area in southern Hungary. Acta Theriol. 2003, 48, 547–556. [Google Scholar] [CrossRef]
- Linnell, J.D.C.; Kaczensky, P.; Wotschikowsky, U.; Lescureux, N.; Boitani, L. Framing the relationship between people and nature in the context of European conservation. Conserv. Biol. 2015, 29, 978–985. [Google Scholar] [CrossRef]
- Putman, J.; Staines, B.W.; Putman, R.; Putman, R.J. Supplementary winter feeding of wild red deer Cervus elaphus in Europe and North America: Justifications, feeding practice and effectiveness. Mammal Rev. 2004, 34, 285–306. [Google Scholar] [CrossRef]
- Katona, K.; Gál-Bélteki, A.; Terhes, A.; Bartucz, K.; Szemethy, L. How important is supplementary feed in the winter diet of red deer? A test in Hungary. Wildl. Biol. 2014, 20, 326–334. [Google Scholar] [CrossRef]
- Janis, C. An Evolutionary History of Browsing and Grazing Ungulates. In The Ecology of Browsing and Grazing; Gordon, I., Prins, H., Eds.; Springer: Berlin, Germany, 2008; pp. 21–42. [Google Scholar]
- Skarpe, C.; Hester, A. Plant Traits, Browsing and Grazing Herbivores, and Vegetation Dynamics. In The Ecology of Browsing and Grazing; Gordon, I., Prins, H., Eds.; Springer: Berlin, Germany, 2008. [Google Scholar]
- Potash, A.D.; Conner, L.M.; McCleery, R.A. Vertical and horizontal vegetation cover synergistically shape prey behaviour. Anim. Behav. 2019, 152, 39–44. [Google Scholar] [CrossRef]
- Putman, R.J. Ungulates in temperate forest ecosystems: Perspectives and recommendations for future research. Ecol. Manag. 1996, 88, 205–214. [Google Scholar] [CrossRef]
- Gill, R.M.A. A Review of Damage by Mammals in North Temperate Forests: 3. Impact on Trees and Forests. For. Int. J. For. Res. 1992, 65, 363–388. [Google Scholar] [CrossRef]
- Radu, I. Damages caused by cervidae in the forests of North of Romania and prevention steps taken. Rev. Padur. 1989, 1, 26–31. [Google Scholar]
- Gill, R.M.A. A Review of Damage by Mammals in North Temperate Forests: 1. Deer. For. Int. J. For. Res. 1992, 65, 145–169. [Google Scholar] [CrossRef]
- Kupferschmid, A.D.; Greilsamer, R.; Brang, P.; Bugmann, H. Assessment of the Impact of Ungulate Browsing on Tree Regeneration. In Animal Nutrition—Annual Volume 2023; Ronquillo, M.G., Ed.; IntechOpen: Rijeka, Croatia, 2022; Chapter 9. [Google Scholar]
- Borkowska, A.; Konopko, A. The winter browse supply for moose in different forest site-types in the Biebrza Valley, Poland. Acta Theriol. 1994, 39, 67–71. [Google Scholar] [CrossRef]
- Nichols, R.V.; Cromsigt, J.P.G.M.; Spong, G. DNA left on browsed twigs uncovers bite-scale resource use patterns in European ungulates. Oecologia 2015, 178, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Kupferschmid, A.D.; Bugmann, H. Effect of microsites, logs and ungulate browsing on Picea abies regeneration in a mountain forest. Ecol. Manag. 2005, 210, 251–265. [Google Scholar] [CrossRef]
- United Nations. UNSD—Methodology. 14 June 2023. Available online: https://unstats.un.org/unsd/methodology/m49/ (accessed on 4 August 2023).
- Barančeková, M.; Krojerová-Prokešová, J.; Homolka, M. Impact of deer browsing on natural and artificial regeneration in floodplain forest Monitoring of European wildcat in the Western Carpathians View project Wild boar as an important factor in the development of forest ecosystems View project. Folia Zool. 2007, 56, 354–364. [Google Scholar]
- Bulušek, D.; Bílek, L. The role of shelterwood cutting and protection against game browsing for the regeneration of silver fir Sustainable forest management based on the harmonization of the individual components of forest ecosystems in the context of the ongoing climate change View project. Austrian J. For. Sci. 2015, 132, 81–102. [Google Scholar]
- Čermák, P.; Grundmann, P. Effects of browsing on the condition and development of regeneration of trees in the region of Rýchory (Krnap). Acta Univ. Agric. Silv. Mendel. Brun. 2006, 1, 7–14. [Google Scholar] [CrossRef]
- Cermak, P.; Glogar, J.; Jankovsky, L. Damage by deer barking and browsing and subsequent rots in Norway spruce stands of Forest Range Mořkov, Forest District Frenštát p. R. (the Beskids Protected Landscape Area). J. Sci. 2004, 50, 24–30. [Google Scholar] [CrossRef]
- Cermak, P.; Mikva, R. Effects of game on the condition and development of natural regeneration in the Vrapač National Nature Reserve (Litovelské Pomoraví). J. Sci. 2006, 52, 329–336. [Google Scholar] [CrossRef]
- Homolka, M.; Heroldová, M. Impact of large herbivores on mountain forest stands in the Beskydy Mountains. Ecol. Manag. 2003, 181, 119–129. [Google Scholar] [CrossRef]
- Kamler, J.; Homolka, M.; Barančeková, M.; Krojerová-Prokešová, J. Reduction of herbivore density as a tool for reduction of herbivore browsing on palatable tree species. Eur. J. Res. 2010, 129, 155–162. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Bílek, L.; Král, J.; Remeš, J.; Bulušek, D.; Králícek, I. Ungulate impact on natural regeneration in spruce-beech-fir stands in Černý důl nature reserve in the Orlické hory Mountains, case study from central sudetes. Forests 2014, 5, 2929–2946. [Google Scholar] [CrossRef]
- Fuchs, Z.; Vacek, Z.; Vacek, S.; Gallo, J. Effect of game browsing on natural regeneration of European beech (Fagus sylvatica L.) forests in the Krušné hory Mts. (Czech Republic and Germany). Cent. Eur. For. J. 2021, 67, 166–180. [Google Scholar] [CrossRef]
- Kuijper, D.P.J.; Jędrzejewska, B.; Brzeziecki, B.; Churski, M.; Jędrzejewski, W.; Żybura, H. Fluctuating ungulate density shapes tree recruitment in natural stands of the BiałOwieża Primeval Forest, Poland. J. Veg. Sci. 2010, 21, 1082–1098. [Google Scholar] [CrossRef]
- Borowski, Z.; Gil, W.; Bartoń, K.; Zajączkowski, G.; Łukaszewicz, J.; Tittenbrun, A.; Radliński, B. Density-related effect of red deer browsing on palatable and unpalatable tree species and forest regeneration dynamics. Ecol. Manag. 2021, 496, 119442. [Google Scholar] [CrossRef]
- Bodziarczyk, J.; Zwijacz-Kozica, T.; Zwijacz-Kozica, T.; Gazda, A.; Szewczyk, J.; Frączek, M.; Zięba, A.; Szwagrzyk, J. Species composition, elevation, and former management type affect browsing pressure on forest regeneration in the Tatra National Park. Leśne Pr. Badaw. 2017, 78, 238–247. [Google Scholar] [CrossRef]
- Kowalczyk, R.; Taberlet, P.; Coissac, E.; Valentini, A.; Miquel, C.; Kaminski, T.S.; Wójcik, J.M. Influence of management practices on large herbivore diet—Case of European bison in Białowieża Primeval Forest (Poland). Ecol. Manag. 2011, 261, 821–828. [Google Scholar] [CrossRef]
- Kuijper, D.P.J.; de Kleine, C.; Churski, M.; van Hooft, P.; Bubnicki, J.; Jedrzejewska, B. Landscape of fear in Europe: Wolves affect spatial patterns of ungulate browsing in Bialowieża Primeval Forest, Poland. Ecography 2013, 36, 1263–1275. [Google Scholar] [CrossRef]
- Kuijper, D.P.J.; Cromsigt, J.P.G.M.; Churski, M.; Adam, B.; Jędrzejewska, B.; Jędrzejewski, W. Do ungulates preferentially feed in forest gaps in European temperate forest. Ecol. Manag. 2009, 258, 1528–1535. [Google Scholar] [CrossRef]
- Orman, O.; Dobrowolska, D.; Szwagrzyk, J. 9/Gap regeneration patterns in Carpathian old-growth mixed beech forests—Interactive effects of spruce bark beetle canopy disturbance and deer herbivory. Ecol. Manag. 2018, 430, 451–459. [Google Scholar] [CrossRef]
- Szwagrzyk, J.; Gazda, A.; Muter, E.; Pielech, R.; Szewczyk, J.; Zięba, A.; Zwijacz-Kozica, T.; Wiertelorz, A.; Pachowicz, T.; Bodziarczyk, J. Effects of species and environmental factors on browsing frequency of young trees in mountain forests affected by natural disturbances. Ecol. Manag. 2020, 474, 118364. [Google Scholar] [CrossRef]
- Vlad, R. Scientific principles for the ecological reconstruction of the norway spruce stands affected by deer. Proc. Rom. Acad. Ser. B 2007, 2, 165–169. [Google Scholar]
- Vlad, R.; Popa, I. The probability of occurrence of deer damage in Norway spruce stands. Proc. Rom. Acad. Ser. B 2007, 1, 57–62. [Google Scholar]
- Vlad, R.; Cuciurean, C. Economic aspects of the deer damage in Norway spruce stand. Rev. Pădurilor 2009, 6, 25–32. [Google Scholar]
- Vlad, R.; Sidor, C.G. Amplitude of the deer damage in the norway spruce forest of the eastern carpathian mountains. Carpathian J. Earth Environ. Sci. 2011, 6, 207–214. [Google Scholar]
- Radu, I. Some observations on deer damages in the woods of the Iacobeni forest district. Rev. Padur. 1964, 11, 640–643. [Google Scholar]
- Radu, I. The amount of damages caused by red deers in some spruce stands. Rev. Pădurilor 1971, 12, 629–631. [Google Scholar]
- Radu, I. The damage caused by deer in the forests from the north of Romania and the necessary preventive measures. Rev. Padur. 1989, 3, 151–154. [Google Scholar]
- Radu, I. Wolves and the ecological balance of forests in Bukovina. Rev. Padur. 1987, 1, 25–28. [Google Scholar]
- Abaturov, B.D.; Smirnov, K.A. Moose browsing impacts on reforestation in Central European USSR. Alces 1992, 3, 213. [Google Scholar]
- Konôpka, B.; Pajtík, J.; Shipley, L.A. Intensity of red deer browsing on young rowans differs between freshly-felled and standing individuals. Ecol. Manag. 2018, 429, 511–519. [Google Scholar] [CrossRef]
- Konôpka, B.; Šebeň, V.; Pajtík, J.; Shipley, L.A. Excluding large wild herbivores reduced norway spruce dominance and supported tree species richness in a young, naturally regenerated stand. Forests 2021, 12, 737. [Google Scholar] [CrossRef]
- Olesen, C.R.; Madsen, P. The impact of roe deer (Capreolus capreolus), seedbed, light and seed fall on natural beech (Fagus sylvatica) regeneration. Ecol. Manag. 2008, 255, 3962–3972. [Google Scholar] [CrossRef]
- Komonen, A.; Tuominen, L.; Purhonen, J.; Halme, P. Landscape structure influences browsing on a keystone tree species in conservation areas. Ecol. Manag. 2020, 457, 117724. [Google Scholar] [CrossRef]
- Bianchi, S.; Huuskonen, S.; Hynynen, J.; Oijala, T.; Siipilehto, J.; Saksa, T. Development of young mixed Norway spruce and Scots pine stands with juvenile stand management in Finland. Scand. J. Res. 2021, 36, 374–388. [Google Scholar] [CrossRef]
- Atte, K.; Komonen, A.; Paananen, E.; Esko, P.; Merja, E.; Elo, M.; Sauli, V.; Valkonen, S. Browsing hinders the regeneration of broadleaved trees in uneven-aged forest management in southern Finland. Scand. J. Res. 2020, 35, 134–138. [Google Scholar] [CrossRef]
- Heikkilä, R.; Hokkanen, P.; Kooiman, M.; Ayguney, N.; Bassoulet, C. The impact of moose browsing on tree species composition in Finland. ALCES 2003, 39, 203–213. [Google Scholar]
- Lyly, M.; Klemola, T.; Koivisto, E.; Huitu, O.; Oksanen, L.; Korpimäki, E. Varying impacts of cervid, hare and vole browsing on growth and survival of boreal tree seedlings. Oecologia 2014, 174, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Lyly, O.; Saksa, T. The effect of stand density on moose damage in young Pinus sylvestris stands. Scand. J. Res. 1992, 7, 393–403. [Google Scholar] [CrossRef]
- Šņepsts, G.; Bigac, Z.; Desaine, I.; Jansons, J.; Donis, J.; Strelnieks, K.; Adamovičs, A.; Krišāns, O. Characteristics of damages in Norway spruce stands. Res. Rural. Dev. 2018, 1, 65–71. [Google Scholar]
- Kaien, C. Deer Browsing and Impact on Forest Development. J. Sustain. For. 2006, 21, 53–64. [Google Scholar] [CrossRef]
- Bergquist, J.; Bergquist, J.; Örlander, G.; Nilsson, U. Deer browsing and slash removal affect field vegetation on south Swedish clearcuts. Ecol. Manag. 1999, 115, 171–182. [Google Scholar] [CrossRef]
- Bergquist, J.; Örlander, G. Browsing deterrent and phytotoxic effects of roe deer repellents on pinus sylvestris and picea abies seedlings. Scand. J. Res. 1996, 11, 145–152. [Google Scholar] [CrossRef]
- Bergquist, J.; Bergström, R.; Zakharenka, A. Responses of young Norway spruce (Picea abies) to winter browsing by roe deer (Capreolus capreolus): Effects on height growth and stem morphology. Scand. J. Res. 2003, 18, 368–376. [Google Scholar] [CrossRef]
- Bergquist, J.; Kullberg, Y.; Orlander, G. Effects of shelterwood and soil scarification on deer browsing on planted Norway spruce Picea abies L. (Karst) seedlings. Forestry 2001, 74, 359–367. [Google Scholar] [CrossRef]
- Bergquist, J.; Löf, M.; Örlander, G. Effects of roe deer browsing and site preparation on performance of planted broadleaved and conifer seedlings when using temporary fences. Scand. J. Res. 2009, 24, 308–317. [Google Scholar] [CrossRef]
- Bergquist, J.; Orlander, G.; Nilsson, U. Interactions among forestry regeneration treatments, plant vigour and browsing damage by deer. New 2003, 25, 25–40. [Google Scholar] [CrossRef]
- Bergquist, J.; Orlander, G.O. Browsing damage by roe deer on Norway spruce seedlings planted on clearcuts of different ages: 2. Effect of seedling vigour. Ecol. Manag. 1998, 105, 295–302. [Google Scholar] [CrossRef]
- Bergström, R.; Bergqvist, G. Frequencies and patterns of browsing by large herbivores on conifer seedlings. Scand. J. Res. 1997, 12, 288–294. [Google Scholar] [CrossRef]
- Bergström, R.; Bergqvist, G. Large herbivore browsing on conifer seedlings related to seedling morphology. Scand. J. Res. 1999, 14, 361–367. [Google Scholar] [CrossRef]
- de Chantal, M.; Granström, A. Aggregations of dead wood after wildfire act as browsing refugia for seedlings of Populus tremula and Salix caprea. Ecol. Manag. 2007, 250, 3–8. [Google Scholar] [CrossRef]
- Edenius, L.; Ericsson, G. Effects of ungulate browsing on recruitment of aspen and rowan: A demographic approach. Scand. J. Res. 2015, 30, 283–288. [Google Scholar] [CrossRef]
- Garrido, P.; Lindqvist, S.; Kjellander, P. Natural forage composition decreases deer browsing on Picea abies around supplemental feeding sites. Scand. J. Res. 2014, 29, 234–242. [Google Scholar]
- Jarnemo, A.; Minderman, J.; Bunnefeld, N.; Zidar, J.; Nsson, J.M.; Jarnemo, C.; Minderman, J.; Bunnefeld, N.; Zidar, J.; Månsson, J. Managing landscapes for multiple objectives: Alternative forage can reduce the conflict between deer and forestry. Ecosphere 2014, 5, 1–14. [Google Scholar] [CrossRef]
- Jensen, A.M.; Götmark, F.; Löf, M. Shrubs protect oak seedlings against ungulate browsing in temperate broadleaved forests of conservation interest: A field experiment. Ecol. Manag. 2012, 266, 187–193. [Google Scholar] [CrossRef]
- Kullberg, Y.; Bergström, R. Winter browsing by large herbivores on planted deciduous seedlings in southern Sweden. Scand. J. Res. 2001, 16, 371–378. [Google Scholar] [CrossRef]
- Nichols, R.V.; Spong, G. Ungulate browsing on conifers during summer as revealed by DNA. Scand. J. Res. 2014, 29, 650–652. [Google Scholar] [CrossRef]
- Pfeffer, S.E.; Singh, N.J.; Cromsigt, J.P.G.M.; Widemo, F. Summer and winter browsing affect conifer growth differently: An experimental study in a multi-species ungulate community. Ecol. Manag. 2021, 494, 119314. [Google Scholar] [CrossRef]
- Bergqvist, G.; Bergström, R.; Wallgren, M. Recent browsing damage by moose on scots pine, birch and aspen in young commercial forests-effects of forage availability, moose population density and site productivity. Silva Fenn. 2014, 48, 1077. [Google Scholar] [CrossRef]
- Edenius, L. Browsing by Moose on Scots Pine in Relation to Plant Resource Availability. Ecology 1993, 8, 2261–2269. [Google Scholar] [CrossRef]
- Ericsson, G.; Edenius, L.; Sundström, D. Factors affecting browsing by moose (Alces alces L.) on European aspen (Populus tremula L.) in a managed boreal landscape. Ecoscience 2001, 8, 344–349. [Google Scholar] [CrossRef]
- Ball, J.P.; Danell, K.; Sunesson, P. Response of a herbivore community to increased food quality and quantity: An experiment with nitrogen fertilizer in a boreal forest. J. Appl. Ecol. 2000, 37, 247–255. [Google Scholar] [CrossRef]
- Edenius, L.; Danell, K.; Bergström, R. Impact of herbivory and competition on compensatory growth in woody plants: Winter browsing by moose on Scots pine. Oikos 1993, 66, 286–292. [Google Scholar] [CrossRef]
- Andrén, H.; Angelstam, P. Moose Browsing on Scots Pine in Relation to Stand Size and Distance to Forest Edge. J. Appl. Ecol. 1993, 30, 133–142. [Google Scholar] [CrossRef]
- Danell, K.; Edenius, L.; Lundberg, P. Herbivory and Tree Stand Composition: Moose Patch Use in Winter. Ecology 1991, 72, 1350–1357. [Google Scholar] [CrossRef]
- Faber’, W.E.; Pehrson, A. Foraging on Norway spruce and its potential association with a wasting syndrome in moose in Sweden. ALCES 2000, 36, 17–34. [Google Scholar]
- Franklin, C.M.A.; Harper, K.A. Moose browsing, understorey structure and plant species composition across spruce budworm-induced forest edges. J. Veg. Sci. 2016, 27, 524–534. [Google Scholar] [CrossRef]
- Franklin, O.; Krasovskiy, A.; Kraxner, F.; Platov, A.; Schepaschenko, D.; Leduc, S.; Mattsson, B. Moose or spruce: A systems analysis model for managing conflicts between moose and forestry in Sweden. bioRxiv 2020. [Google Scholar] [CrossRef]
- Löf, M.; Bergquist, J.; Brunet, J.; Karlsson, M.; Torkel, N. Conversion of Norway spruce stands to broadleaved woodland-regeneration systems, fencing and performance of planted seedlings. Ecol. Bull. 2010, 53, 165–174. [Google Scholar]
- van Beeck Calkoen, S.T.S.; Kuijper, D.P.J.; Sand, H.; Singh, N.J.; van Wieren, S.E.; van Beeck Calkoen, J.P.G.M.C.S.T.S.; Singh, N.J.; Cromsigt, J.P.G.M.; Kuijper, S.P.A.-D.J.; van Wieren, S.E.-S. Does wolf presence reduce moose browsing intensity in young forest plantations? Ecography 2018, 41, 1776–1787. [Google Scholar] [CrossRef]
- Shipley, L.; Blomquist, S.; Danell, K. Diet choices made by free-ranging moose in northern Sweden in relation to plant distribution, chemistry, and morphology. Can. J. Zool. 1998, 76, 1722–1733. [Google Scholar] [CrossRef]
- Danell, K.; Niemelä, P.; Niemelä, P.; Varvikko, T.; Vuorisalo, T. Moose browsing on Scots pine along a gradient of plant productivity. Ecology 1991, 72, 1624–1633. [Google Scholar] [CrossRef]
- Hörnberg, S. The relationship between moose (Alces alces) browsing utilisation and the occurrence of different forage species in Sweden. Ecol. Manag. 2001, 149, 91–102. [Google Scholar] [CrossRef]
- van Beest, F.M.; Gundersen, H.; Mathisen, K.M.; Milner, J.M.; Skarpe, C. Long-term browsing impact around diversionary feeding stations for moose in Southern Norway. Ecol. Manag. 2010, 259, 1900–1911. [Google Scholar] [CrossRef]
- Hörnberg, S. Changes in population density of moose (Alces alces) and damage to forests in Sweden. Ecol. Manag. 2001, 149, 141–151. [Google Scholar] [CrossRef]
- Harmer, R. The effect of plant competition and simulated summer browsing by deer on tree regeneration. J. Appl. Ecol. 2002, 38, 1094–1103. [Google Scholar] [CrossRef]
- Joys, A.C.; Fuller, R.J.; Dolman, P.M. Influences of deer browsing, coppice history, and standard trees on the growth and development of vegetation structure in coppiced woods in lowland England. Ecol. Manag. 2004, 202, 23–37. [Google Scholar] [CrossRef]
- Baines, D.; Sage, R.B.; Sage, R.B.; Baines, M.M. The implications of red deer grazing to ground vegetation and invertebrate communities of scottish native pinewoods. J. Appl. Ecol. 1994, 31, 776–783. [Google Scholar] [CrossRef]
- Duncan, A.J.; Hartley, S.E.; Iason, G.R. The effect of monoterpene concentrations in Sitka spruce (Picea sitchensis) on the browsing behaviour of red deer (Cervus elaphus). Can. J. Zool. 1994, 72, 1715–1720. [Google Scholar] [CrossRef]
- Duncan, A.J.; Hartley, S.E.; Iason, G.R. The effect of previous browsing damage on the morphology and chemical composition of Sitka spruce Picea sitchensis saplings and on their subsequent susceptibility to browsing by red deer. Ecol. Manag. 1998, 103, 57–67. [Google Scholar] [CrossRef]
- Gill, R.M.A.; Fuller, R.J. The effects of deer browsing on woodland structure and songbirds in lowland Britain. Ibis 2007, 149, 119–127. [Google Scholar] [CrossRef]
- Harrison, K.A.; Bardgett, R.D. Browsing by red deer negatively impacts on soil nitrogen availability in regenerating native forest. Soil. Biol. Biochem. 2004, 36, 115–126. [Google Scholar] [CrossRef]
- Hester, A.J.; Millard, P.; Millard, P.; Baillie, G.J.; Wendl, R.er. How does timing of browsing affect above- and below-ground growth of Betula pendula, Pinus sylvestris and Sorbus aucuparia? Oikos 2004, 105, 536–550. [Google Scholar] [CrossRef]
- Kay, S. Factors affecting severity of deer browsing damage within coppiced woodlands in the south of England. Biol. Conserv. 1993, 63, 21–222. [Google Scholar] [CrossRef]
- Moore, N.P.; Hart, J.D.; Kelly, P.F.; Langton, S.D. Browsing by fallow deer (Dama dama) in young broadleaved plantations: Seasonality, and the effects of previous browsing and bud eruption. Forestry 2000, 73, 437–445. [Google Scholar] [CrossRef]
- Moore, N.P.; Hart, J.D.; Langton, S.D.; Langton, S.D. Factors influencing browsing by fallow deer Dama dama in young broad-leaved plantations. Biol. Conserv. 1999, 87, 255–260. [Google Scholar] [CrossRef]
- Page, L.M.; Cameron, A.D. Regeneration dynamics of Sitka spruce in artificially created forest gaps. Ecol. Manag. 2006, 221, 260–266. [Google Scholar] [CrossRef]
- Palmer, S.C.F.; Truscott, A.M. Browsing by deer on naturally regenerating Scots pine (Pinus sylvestris L.) and its effects on sapling growth. Ecol. Manag. 2003, 182, 31–47. [Google Scholar] [CrossRef]
- Scott, D.F.; Scott, D.; Welch, D.W.; Thurlow, M.; Elston, D.A. Regeneration of Pinus sylvestris in a natural pinewood in NE Scotland following reduction in grazing by Cervus elaphus. Ecol. Manag. 2000, 130, 199–211. [Google Scholar] [CrossRef]
- Mayhead, G.J.; Jenkins, A.A.R. Growth of Young Sitka Spruce (Picea sitchensis (Bong.) Can.) and the Effect of Simulated Browsing, Staking and Treeshelters. Forestry 1992, 65, 453–462. [Google Scholar] [CrossRef]
- Miller, G.R.; Kinnaird, J.W.; Cummins, R.P. Liability of saplings to browsing on a red deer range in the Scottish highlands. Source J. Appl. Ecol. 1982, 19, 941–951. [Google Scholar] [CrossRef]
- Scott, D.; Scott, D.; Welch, D.W.; Elston, D.A. Long-term effects of leader browsing by deer on the growth of Sitka spruce (Picea sitchensis). Forestry 2009, 82, 387–401. [Google Scholar] [CrossRef]
- Welch, D.W.; Staines, B.W.; Scott, D.; Scott, D.; French, D.D.; Catt, D.C. Leader Browsing by Red and Roe Deer on Young Sitka Spruce Trees in Western Scotland I. Damage Rates and the Influence of Habitat Factors. Forestry 1991, 1, 61–82. [Google Scholar] [CrossRef]
- Staines, B.W.; Welch, D. Habitat selection and impact of red (Cervus elaphus L.) and roe (Capreolus capreolus L.) deer in a Sitka spruce plantation. Proc. R. Soc. Edinb. 1984, 82, 303–319. [Google Scholar] [CrossRef]
- de Jong, C.B.; Gill, R.M.A.; van Wieren, S.E.; Burlton, F.W.E. Diet selection by roe deer Capreolus capreolus in Kielder Forest in relation to plant cover. Ecol. Manag. 1995, 79, 91–97. [Google Scholar] [CrossRef]
- Cooke, A.S.; Farrell, L. Impact of muntjac deer (Muntiacus reevesi) at Monks Wood National Nature Reserve, Cambridgeshire, eastern England. Forestry 2001, 74, 241–250. [Google Scholar] [CrossRef]
- Cooke, A.S.; Lakhani, K.H. Damage to coppice regrowth by muntjac deer (Muntiacus reevesi) and protection with electric fencing. Biol. Conserv. 1996, 75, 231–238. [Google Scholar] [CrossRef]
- Cooke, A. Survival and regrowth performance of coppiced ash (Fraxinus excelsior) in relation to browsing damage by muntjac deer (Muntiacus reevesi). Q. J. For. 1998, 92, 286–290. [Google Scholar]
- Bottero, A.; Garbarino, M.; Long, J.N.; Motta, R. The interacting ecological effects of large-scale disturbances and salvage logging on montane spruce forest regeneration in the western European Alps. Ecol. Manag. 2013, 292, 19–28. [Google Scholar] [CrossRef]
- D’Aprile, D.; Vacchiano, G.; Vacchiano, G.; Meloni, F.; Garbarino, M.; Motta, R.; Ducoli, V.; Partel, P. Effects of Twenty Years of Ungulate Browsing on Forest Regeneration at Paneveggio Reserve, Italy. Forests 2020, 11, 612. [Google Scholar] [CrossRef]
- Motta, R. Impact of wild ungulates on forest regeneration and tree composition of mountain forests in the western Italian Alps. Ecol. Manag. 1996, 88, 93–98. [Google Scholar] [CrossRef]
- Motta, R. Ungulate impact on rowan (Sorbus aucuparia L.) and Norway spruce (Picea abies (L.) Karst.) height structure in mountain forests in the eastern Italian Alps. Ecol. Manag. 2003, 181, 139–150. [Google Scholar] [CrossRef]
- Motta, R. Wild ungulate browsing, natural regeneration and silviculture in the italian alps. J. Sustain. For. 1998, 8, 35–53. [Google Scholar] [CrossRef]
- Cutini, A.; Chianucci, F.; Apollonio, M. Wild ungulates and forests in Europe: Insights from long term studies in Central Italy. In Proceedings of the Second International Congress of Silviculture, Designing the Future of the Forestry Sector, Florence, Italy, 26–29 November 2014; Italian Academy of Forestry Sciences: Florence, Italy, 2015; Volume 1, pp. 509–517. [Google Scholar]
- Lovari, S.; Cuccus, P.; Murgia, A.; Murgia, C.; Soi, F.; Plantamura, G. Space use, habitat selection and browsing effects of red deer in Sardinia. Ital. J. Zool. 2007, 74, 179–189. [Google Scholar] [CrossRef]
- Freschi, P.; Fascetti, S.; Riga, F.; Rizzardini, G.; Musto, M.; Cosentino, C. Feeding preferences of the Italian roe deer (Capreolus capreolus italicus festa, 1925) in a coastal mediterranean environment. Animals 2021, 11, 308. [Google Scholar] [CrossRef]
- Klopcic, M.; Jerina, K.; Bončina, A. Long-term changes of structure and tree species composition in Dinaric uneven-aged forests: Are red deer an important factor? Eur. J. Res. 2010, 192, 277–288. [Google Scholar] [CrossRef]
- Rozman, A.; Diaci, J.; Krese, A.; Fidej, G.; Rozenbergar, D. Forest regeneration dynamics following bark beetle outbreak in Norway spruce stands: Influence of meso-relief, forest edge distance and deer browsing. Ecol. Manag. 2015, 353, 196–207. [Google Scholar] [CrossRef]
- Monzón, A.; da Silva, S.V.; Manso, F.T. Integrating the deer (Cervus elaphus) in the Portuguese forests: Impacts and new challenges for forest certification. Ecol. Manag. 2012, 267, 1–6. [Google Scholar] [CrossRef]
- Meier, M.; Stöhr, D.; Walde, J.; Tasser, E. Influence of ungulates on the vegetation composition and diversity of mixed deciduous and coniferous mountain forest in Austria. Eur. J. Wildl. Res. 2017, 63, 29. [Google Scholar] [CrossRef]
- Pröll, G.; Darabant, A.; Gratzer, G.; Katzensteiner, K. Unfavourable microsites, competing vegetation and browsing restrict post-disturbance tree regeneration on extreme sites in the Northern Calcareous Alps. Eur. J. Res. 2015, 134, 293–308. [Google Scholar] [CrossRef]
- Rüegg, D.; Nigg, H. Mehrstufige Verjüngungskontrollen und Grenzwerte für die Verbissintensität | Comparitive regeneration control and limiting value of browsing damage intensity. Schweiz. Z. Forstwes. 2003, 154, 314–321. [Google Scholar] [CrossRef]
- Reimoser, F.; Gossow, H. Impact of ungulates on forest vegetation and its dependence on the silvicultural system. For. Ecol. Manag. 1996, 88, 107–126. [Google Scholar] [CrossRef]
- Partl, E.; Szinovatz, V.; Reimoser, F.; Schweiger-Adler, J. Forest restoration and browsing impact by roe deer. Ecol. Manag. 2002, 159, 87–100. [Google Scholar] [CrossRef]
- Griesberger, P.; Kunz, F.; Reimoser, F.; Hackländer, K.; Obermair, L. Spatial Distribution of Hunting and Its Potential Effect on Browsing Impact of Roe Deer (Capreolus capreolus) on Forest Vegetation. Diversity 2023, 15, 613. [Google Scholar] [CrossRef]
- Nopp-Mayr, U.; Reimoser, S.; Reimoser, F.; Sachser, F.; Obermair, L.; Gratzer, G. Analyzing long-term impacts of ungulate herbivory on forest-recruitment dynamics at community and species level contrasting tree densities versus maximum heights. Sci. Rep. 2020, 10, 20274. [Google Scholar] [CrossRef]
- Reimoser, F.; Reimoser, S.; Zsak, K. Long-term impact of wild ungulates on natural forest regeneration in the Donau-Auen National Park, Austria. Acta ZooBot Austria 2022, 158, 97–127. [Google Scholar]
- Bernard, M.; Gamelon, M.; Boulanger, V.; Dupouey, J.-L.; Laurent, L.; Laurent, L.; Montpied, P.; Montpied, P.; Morin, X.; Picard, J.F.; et al. Deer browsing promotes Norway spruce at the expense of silver fir in the forest regeneration phase. Ecol. Manag. 2017, 400, 269–277. [Google Scholar] [CrossRef]
- Chevrier, T.; Saïd, S.; Widmer, O.; Hamard, J.P.; Saint-Andrieux, C.; Gaillard, J.M. The oak browsing index correlates linearly with roe deer density: A new indicator for deer management? Eur. J. Wildl. Res. 2012, 58, 17–22. [Google Scholar] [CrossRef]
- Drexhage, M.; Colin, F. Effects of browsing on shoots and roots of naturally regenerated sessile oak seedlings. Ann. Sci. 2003, 60, 173–178. [Google Scholar] [CrossRef]
- Boulanger, V.; Baltzinger, C.; Saïd, S.; Ballon, P.; Picard, J.F.; Dupouey, J.L. Decreasing deer browsing pressure influenced understory vegetation dynamics over 30 years. Ann. Sci. 2015, 72, 367–378. [Google Scholar] [CrossRef]
- Boulanger, V.; Baltzinger, C.; Saïd, S.; Ballon, P.; Picard, J.F.; Dupouey, J.L. Ranking temperate woody species along a gradient of browsing by deer. Ecol. Manag. 2009, 258, 1397–1406. [Google Scholar] [CrossRef]
- Mårell, A.; Hamard, J.P.; Pérot, T.; Perret, S.; Korboulewsky, N. The effect of deer browsing and understory light availability on stump mortality and sprout growth capacity in sessile oak. Ecol. Manag. 2018, 430, 134–142. [Google Scholar] [CrossRef]
- Pépin, D.; Renaud, P.C.; Boscardin, Y.; Goulard, M.; Mallet, C.; Anglard, F.; Ballon, P. Relative impact of browsing by red deer on mixed coniferous and broad-leaved seedlings—An enclosure-based experiment. Ecol. Manag. 2006, 222, 302–313. [Google Scholar] [CrossRef]
- Unkule, M.; Piedallu, C.; Balandier, P.; Courbaud, B. Climate and ungulate browsing impair regeneration dynamics in spruce-fir-beech forests in the French Alps. Ann. Sci. 2022, 79, 11. [Google Scholar] [CrossRef]
- Heuze, P.; Schnitzler, A.; Klein, F. Is browsing the major factor of silver fir decline in the Vosges Mountains of France? Ecol. Manag. 2005, 217, 219–228. [Google Scholar] [CrossRef]
- Tschöpe, O.; Wallschläger, D.; Burkart, M.; Tielbörger, K. Managing open habitats by wild ungulate browsing and grazing: A case-study in North-Eastern Germany. Appl. Veg. Sci. 2011, 14, 200–209. [Google Scholar] [CrossRef]
- Amrner, C. Impact of ungulates on structure and dynamics of natural regeneration of mixed mountain forests in the Bavarian Alps. Ecol. Manag. 1996, 88, 43–53. [Google Scholar] [CrossRef]
- Bobrowski, M.; Gillich, B.; Stolter, C. Modelling browsing of deer on beech and birch in northern Germany. Ecol. Manag. 2015, 358, 212–221. [Google Scholar] [CrossRef]
- Bödeker, K.; Ammer, C.; Knoke, T.; Heurich, M. Determining Statistically Robust Changes in Ungulate Browsing Pressure as a Basis for Adaptive Wildlife Management. Forests 2021, 12, 1030. [Google Scholar] [CrossRef]
- van Beeck Calkoen, S.T.S.; Deis, M.H.; Oeser, J.; Kuijper, D.P.J.; Heurich, M. Humans rather than Eurasian lynx (Lynx lynx) shape ungulate browsing patterns in a temperate forest. Ecosphere 2022, 13, e3931. [Google Scholar]
- Heinrichs, S.; Winterhoff, W.; Schmidt, W. Vegetation dynamics of beech forests on limestone in central Germany over half a century—Effects of climate change, forest management, eutrophication or game browsing? Biodivers. Ecol. 2012, 4, 49–61. [Google Scholar] [CrossRef]
- Heinze, E.; Boch, S.; Fischer, M.; Hessenmöller, D.; Klenk, B.; Müller, J.; Prati, D.; Schulze, E.D.; Seele, C.; Socher, S.; et al. Habitat use of large ungulates in northeastern Germany in relation to forest management. Ecol. Manag. 2011, 261, 288–296. [Google Scholar] [CrossRef]
- Heuze, P.; Schnitzler, A.; Klein, F. Consequences of increased deer browsing winter on silver fir and spruce regeneration in the Southern Vosges mountains: Implications for forest management. Ann. Sci. 2005, 62, 175–181. [Google Scholar] [CrossRef]
- Hothorn, T.; Müller, J. Large-scale reduction of ungulate browsing by managed sport hunting. Ecol. Manag. 2010, 260, 1416–1423. [Google Scholar] [CrossRef]
- Möst, L.; Hothorn, T.; Müller, J.; Heurich, M. Creating a landscape of management: Unintended effects on the variation of browsing pressure in a national park. Ecol. Manag. 2015, 338, 46–56. [Google Scholar] [CrossRef]
- Schäfer, D.; Prati, D.; Schall, P.; Ammer, C.; Fischer, M. Exclusion of large herbivores affects understorey shrub vegetation more than herb vegetation across 147 forest sites in three German regions. PLoS ONE 2019, 13, e3931. [Google Scholar] [CrossRef]
- Thom, D.; Ammer, C.; Annighöfer, P.; Aszalós, R.; Dittrich, S.; Hagge, J.; Keeton, W.S.; Kovacs, B.; Krautkrämer, O.; Müller, J.; et al. Regeneration in European beech forests after drought: The effects of microclimate, deadwood and browsing. Eur. J. Res. 2022, 142, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Winter, M.B.; Baier, R.; Ammer, C. Regeneration dynamics and resilience of unmanaged mountain forests in the Northern Limestone Alps following bark beetle-induced spruce dieback. Eur. J. Res. 2015, 134, 949–968. [Google Scholar] [CrossRef]
- Schulze, E.D.; Finér, L.; Bouriaud, O.; Wäldchen, J.; Eisenhauer, N.; Walentowski, H.; Seele, C.; Heinze, E.; Heinze, E.; Pruschitzki, U.; et al. Ungulate browsing causes species loss in deciduous forests independent of community dynamics and silvicultural management in Central and Southeastern Europe. Ann. Res. 2014, 57, 267–288. [Google Scholar] [CrossRef]
- Van Hees, A.F.M.; Kuiters, A.T.; Slim, P.A. Growth and development of silver birch, pedunculate oak and beech affected by deer browsing. Ecol. Manag. 1996, 88, 55–63. [Google Scholar] [CrossRef]
- Kuiters, A.T.; Kuiters, A.T.; Slim, P.A. Regeneration of mixed deciduous forest in a Dutch forest-heathland, following a reduction of ungulate densities. Biol. Conserv. 2002, 105, 65–74. [Google Scholar] [CrossRef]
- Kupferschmid, A.D.; Zimmermann, S.; Zimmermann, S.; Bugmann, H. Browsing regime and growth response of naturally regenerated Abies alba saplings along light gradients. Ecol. Manag. 2013, 310, 393–404. [Google Scholar] [CrossRef]
- Cunningham, C.; Zimmermann, N.E.; Stoeckli, V.; Bugmann, H. Growth of Norway spruce (Picea abies L.) saplings in subalpine forests in Switzerland: Does spring climate matter? Ecol. Manag. 2006, 228, 19–32. [Google Scholar] [CrossRef]
- Moser, B.; Schütz, M.; Hindenlang, K.E. Importance of alternative food resources for browsing by roe deer on deciduous trees: The role of food availability and species quality. Ecol. Manag. 2006, 226, 248–255. [Google Scholar] [CrossRef]
- Senn, J.; Suter, W. Ungulate browsing on silver fir (Abies alba) in the Swiss Alps: Beliefs in search of supporting data. Ecol. Manag. 2003, 181, 151–164. [Google Scholar] [CrossRef]
- Kupferschmid, A.D.; Bütikofer, L.; Hothorn, T.; Schwyzer, A.; Brang, P. Ungulate species and abundance as well as environmental factors determine the probability of terminal shoot browsing on temperate forest trees. Forests 2020, 11, 764. [Google Scholar] [CrossRef]
- Kupferschmid, A.D. Selective browsing behaviour of ungulates influences the growth of Abies alba differently depending on forest type. Ecol. Manag. 2018, 429, 317–326. [Google Scholar] [CrossRef]
- Häsler, H.; Senn, J. Ungulate browsing on European silver fir Abies alba: The role of occasions, food shortage and diet preferences. Wildl. Biol. 2012, 18, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Plumb-Gryn, G.; Kowalczyk, R.; Hernandez-Blanco, J. Bison Bonasus. 2020, IUCN Assesement, 2020, 22 Pages. Available online: https://www.iucnredlist.org/species/2814/45156279 (accessed on 23 July 2024).
- Krasińska, M.; Krasiński, Z.A. European Bison the Nature Monograph, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Caboń-Raczyńska, K.; Krasińska, M.; Krasiński, Z.; Wójcik, J. Rhythm of daily activity and behavior of European Bison in the Białowieza Forest in the period without snow cover. Acta Theriol. 1987, 32, 335–372. [Google Scholar] [CrossRef]
- Borowski, S.; Kossak, S. The natural food preferences of the European bison in seasons free of snow cover. Acta Theriol. 1972, 17, 151–169. [Google Scholar] [CrossRef]
- Tudoran, G.M.; Zotta, M. Adapting the planning and management of Norway spruce forests in mountain areas of Romania to environmental conditions including climate change. Sci. Total Environ. 2020, 698, 133761. [Google Scholar] [CrossRef] [PubMed]
- Turnock, D. The romanian carpathians under communism: The changing significance of a mountain region. Rev. Hist. Geogr. Toponomast. 2006, 1, 157–206. [Google Scholar]
- Savin, Y.V.; Zaripov, Y.V.; Belov, L.A.; Zalesova, E.S.; Shubin, D.A. Effect of Elk and Roe on Forest Cultures of Pine and Spruce Conservation; Ural State Forestry Engineering University: Ekaterinburg, Russia, 2017. [Google Scholar]
- Belov, L.A. The Influence of Roe Deer on Natural and Artificial Reforestation of the Dzhabyk-Karagai Pine Forest; Ministry of Natural Resources of the Russian Federation: Moscow, Russia, 2008. (In Russian) [Google Scholar]
- Balčiauskas, L.; Kawata, Y. Red Deer in Lithuania: History, Status and Management. Sustainability 2022, 14, 14091. [Google Scholar] [CrossRef]
- Padaiga, V. The counting of winter pellet groups of cervines as the method of assessment of their browsing pressure and population structure. Balt. 1998, 4, 36–41. [Google Scholar]
- Mysterud, A.; Østbye, E. Roe deer (Capreolus capreolus) browsing pressure affects yew (Taxus baccata) recruitment within nature reserves in Norway. Biol. Conserv. 2004, 120, 545–548. [Google Scholar] [CrossRef]
- Kolstad, A.L.; Austrheim, G.; Solberg, E.J.; De Vriendt, L.; Speed, J.D.M. Pervasive moose browsing in boreal forests alters successional trajectories by severely suppressing keystone species. Ecosphere 2018, 9. [Google Scholar]
- Klopčič, M.; Mina, M.; Bugmann, H.; Bončina, A. The prospects of silver fir (Abies alba Mill.) and Norway spruce (Picea abies (L.) Karst) in mixed mountain forests under various management strategies, climate change and high browsing pressure. Eur. J. Res. 2017, 136, 1071–1090. [Google Scholar] [CrossRef]
- Sofletea, N.; Curtu, L. Dendrologie; Editura Universitatii Transilvania: Brasov, Romania, 2007. [Google Scholar]
- Muñoz-Reinoso, J.C. Effects of deer browsing in a Mediterranean coastal juniper stand. Forestry 2017, 90, 304–311. [Google Scholar] [CrossRef]
- Reimoser, F.; Armstrong, H.; Suchant, R. Measuring forest damage of ungulates: What should be considered. Ecol. Manag. 1999, 120, 47–58. [Google Scholar] [CrossRef]
- Vospernik, S.; Reimoser, S.; Reimoser, S. Modelling changes in roe deer habitat in response to forest management. Ecol. Manag. 2008, 255, 530–545. [Google Scholar] [CrossRef]
- Reimoser, F. Nature Conservation Steering the impacts of ungulates on temperate forests. J. Nat. Conserv. 2003, 10, 243–252. [Google Scholar] [CrossRef]
- Bergeron, D.H.; Bergeron, D.; Pekins, P.J.; Jones, H.; Leak, W.B. Moose browsing and forest regeneration: A case in Northern New Hampshire. Alces 2011, 47, 39–51. [Google Scholar]
- Weigand, J.F.; Haynes, R.W.; Tiedemann, A.R.; Riggs, R.A.; Quigley, T.M. Economic assessment of ungulate herbivory in commercial forests of eastern Oregon and Washington, USA. For. Ecol. Manag. Elsevier, Amsterdam 1993, 61, 137–155. [Google Scholar] [CrossRef]
- Diaci, J. Regeneration dynamics in a Norway spruce plantation on a silver fir-beech forest site in the Slovenian Alps. For. Ecol. Manag., Elsevier, Amsterdam 2001, 161, 27–38. [Google Scholar] [CrossRef]
- Simard, M.A.; Dussault, C.; Huot, J.; Côté, S.D. Is hunting an effective tool to control overabundant deer? A test using an experimental approach. J. Wildl. Manag. 2013, 77, 254–269. [Google Scholar] [CrossRef]
- Boer, A.H. Hunting: A product or a tool for wildlife managers? Alces 1991, 27, 74–78. [Google Scholar]
- Harmer, R. Survival and new shoot production by artificially browsed seedlings of ash, beech, oak and sycamore grown under different levels of shade. Ecol. Manag. 1999, 116, 39–50. [Google Scholar] [CrossRef]
- Barton, O.; Gresham, A.; Healey, J.R.; Cordes, L.S.; Shannon, G. The effects of population management on wild ungulates: A systematic map of evidence for UK species. PLoS ONE 2022, 17, e0267385. [Google Scholar] [CrossRef] [PubMed]
- Valente, A.M.; Acevedo, P.; Figueiredo, A.M.; Fonseca, C.; Torres, R.T. Overabundant wild ungulate populations in Europe: Management with consideration of socio-ecological consequences. Mamm. Rev. 2020, 50, 353–366. [Google Scholar] [CrossRef]
- Ramirez, J.I.; Jansen, P.A.; Poorter, L. Effects of wild ungulates on the regeneration, structure and functioning of temperate forests: A semi-quantitative review. Ecol. Manag. 2018, 424, 406–419. [Google Scholar] [CrossRef]
- Coulson, T. The Science of Overabundance: Deer Ecology and Population Management. Biodivers. Conserv. 1999, 8, 1719–1721. [Google Scholar] [CrossRef]
- Healy, W.M.; de Calesta, D.S.; Stout, S.L. A research perspective on white-tailed deer overabundance in the northeastern United States. Wildl. Soc. Bull. 1997, 25, 259–263. [Google Scholar]
- Clasen, C.; Griess, V.C.; Knoke, T. Financial consequences of losing admixed tree species: A new approach to value increased financial risks by ungulate browsing. Policy Econ. 2011, 13, 503–511. [Google Scholar] [CrossRef]
- Kittredge, D.B.; Ashton, P.M.S. Impact of Deer Browsing on Regeneration in Mixed Stands in Southern New England. North. J. Appl. For. 1995, 12, 115–120. [Google Scholar] [CrossRef]
- DeGraaf, R.M.; Healy, W.M.; Brooks, R.T. Effects of thinning and deer browsing on breeding birds in New England oak woodlands. Ecol. Manag. 1991, 41, 179–191. [Google Scholar] [CrossRef]
- Castleberry, S.B.; Ford, W.M.; Miller, K.V.; Smith, W.P. Influences of herbivory and canopy opening size on forest regeneration in a southern bottomland hardwood forest. Ecol. Manag. 2000, 131, 57–64. [Google Scholar] [CrossRef]
- Cretaz, A.L.; Kelty, M.J. Development of tree regeneration in fern-dominated forest understories after reduction of deer browsing. Restor. Ecol. 2002, 10, 416–426. [Google Scholar] [CrossRef]
- Rea, R.V. Impacts of moose (Alces alces) browsing on paper birch (Betula papyrifera) morphology and potential timber quality. Silva Fenn. 2011, 45, 227–236. [Google Scholar] [CrossRef]
- Beguin, J.; Pothier, D.; Prévost, M. Can the impact of deer browsing on tree regeneration be mitigated by shelterwood cutting and strip clearcutting. Ecol. Manag. 2009, 257, 38–45. [Google Scholar] [CrossRef]
- Chollet, S.; Padié, S.; Stockton, S.; Allombert, S.; Gaston, A.J.; Martin, J.L. Positive plant and bird diversity response to experimental deer population reduction after decades of uncontrolled browsing. Divers. Distrib. 2016, 22, 274–287. [Google Scholar] [CrossRef]
- Cook-Patton, S.C.; LaForgia, M.; Parker, J.D. Positive interactions between herbivores and plant diversity shape forest regeneration. Proc. R. Soc. B Biol. Sci. 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Pekin, B.K.; Wisdom, M.J.; Endress, B.A.; Naylor, B.J.; Parks, C.G. Ungulate browsing maintains shrub diversity in the absence of episodic disturbance in seasonally-arid conifer forest. PLoS ONE 2014, 9, e86288. [Google Scholar] [CrossRef] [PubMed]
- Burņeviča, N.; Ozoliņš, J.; Gaitnieks, T. Vertebrate herbivore browsing and impact on forest production. For. Microbiol. 2022, 3, 251–262. [Google Scholar]
- Grisez, T. Slash Helps Protect Seedlings From Deer Browsing. J. For. 1960, 58, 385–387. [Google Scholar]
- Redick, C.H.; Jacobs, D.F. Mitigation of deer herbivory in temperate hardwood forest regeneration: A meta-analysis of research literature. Forests 2020, 11, 1220. [Google Scholar] [CrossRef]
- Van Lerberghe, P. Protecting Trees from Wildlife Damage-Mesh Tree Guards; CNPF-IDF: Paris, France, 2014. [Google Scholar]
- Berger, J.; Stacey, P.B.; Bellis, L.; Johnson, M.P. A mammalian predator-prey imbalance: Grizzly bear and wolf extinction affect avian neotropical migrants. Ecol. Issues Conserv. 2001, 11, 947–960. [Google Scholar]
- Arnett, E.B.; Southwick, R. Economic and social benefits of hunting in North America. Int. J. Environ. Stud. 2015, 72, 734–745. [Google Scholar] [CrossRef]
- Cotta, V. Economia Vânatului și Salmonicultura; Editura Agricola de Stat: Bucharest, Romania, 1956. [Google Scholar]
- Warren, R.J.; Warnell, D.B. Overview of fertility control in urban deer management. In Proceedings of the 2000 Annual Conference of the Society for Theriogenology, San Antonio, TX, USA, 30 November–2 December 2000; pp. 237–246. [Google Scholar]
- Ripple, W.J.; Larsen, E.J.; Renkin, R.A.; Smith, D.W. Trophic cascades among wolves, elk and aspen on Yellowstone National Park’s northern range. Biol. Conserv. 2001, 102, 227–234. [Google Scholar] [CrossRef]
- Fortin, D.; Beyer, H.L.; Boyce, M.S.; Smith, D.W.; Duchesne, T.; Mao, J.S. Wolves influence elk movements: Behaviour shapes a trophic cascade in Yellowstone National Park. Ecology 2005, 86, 1320–1330. [Google Scholar] [CrossRef]
- Brown, J.S.; Laundré, J.W.; Gurung, M. The Ecology of Fear: Optimal Foraging, Game Theory, and Trophic Interactions. J. Mammal. 1999, 80, 385–399. [Google Scholar] [CrossRef]
- Ripple, W.J.; Beschta, R.L. Wolf reintroduction, predation risk, and cottonwood recovery in Yellowstone National Park. Ecol. Manag. 2003, 184, 299–313. [Google Scholar] [CrossRef]
- Theuerkauf, J.; Rouys, S. Habitat selection by ungulates in relation to predation risk by wolves and humans in the Białowieża Forest, Poland. Ecol. Manag. 2008, 256, 1325–1332. [Google Scholar] [CrossRef]
- Je, W.; Je, B.; Okarma, H.; Schmidt, K.; Zub, K.; Musiani, M. Prey selection and predation by wolves in Białowieża Primeval Forest, Poland. J. Mammal. 2000, 81, 197–212. [Google Scholar]
- Sand, H.; Wikenros, C.; Wabakken, P.; Liberg, O. Cross-continental differences in patterns of predation: Will naive moose in Scandinavia ever learn? Proc. R. Soc. B Biol. Sci. 2006, 273, 1421–1427. [Google Scholar] [CrossRef]
- Gervasi, V.; Sand, H.; Zimmermann, B.; Mattisson, J.; Wabakken, P.; Linnell, J.D.C. Decomposing risk: Landscape structure and wolf behavior generate different predation patterns in two sympatric ungulates. Ecol. Appl. 2013, 23, 1722–1734. [Google Scholar] [CrossRef]
- McCullough, D.R.; Andersen, R.; Duncan, P.; Linnell, J.D.C. The European Roe Deer: The Biology of Success. J. Wildl. Manag. 2000, 64, 608. [Google Scholar] [CrossRef]
- Linnel, D.C.; Aanes, R.; Swenson, S. Translocation of carnivores as a method for managing problem animals: A review. Biodivers. Conserv. 1997, 6, 1245–1257. [Google Scholar] [CrossRef]
- Franchini, M.; Corazzin, M.; Bovolenta, S.; Filacorda, S. The Return of Large Carnivores and Extensive Farming Systems: A Review of Stakeholders’ Perception at an EU Level. Animals 2021, 11, 1735. [Google Scholar] [CrossRef] [PubMed]
- Putman, R.; Langbein, J.; Green, P.; Watson, P. Identifying threshold densities for wild deer in the UK above which negative impacts may occur. Mammal. Rev. 2011, 41, 175–196. [Google Scholar] [CrossRef]

| Europe | |||||||
|---|---|---|---|---|---|---|---|
| Species | Roe Deer | Red Deer | Moose | Chamois | Fallow Deer | European Bison | Muntjac Deer |
| N | 95 | 93 | 32 | 19 | 12 | 4 | 3 |
| Country | Ungulate Species | Problems | Causes | Protective Measures | Large Carnivores | Reference |
|---|---|---|---|---|---|---|
| Czech Republic | Red deer, Roe deer | Development of commercial forests | △ | *** | / | [32,33,34,35,36,37,38,39,40] |
| Poland | Red deer, Roe deer, Chamois, Moose | Development of protection forests | △ | ¦ | +++ | [41,42,43,44,45,46,47,48] |
| Romania | Red deer, Roe deer | Development of commercial forests | △, Monocultures on large areas | ** | ++ | [25,50,52,53,54,55,56] |
| Russia | Moose | Unwanted changes in tree composition | △ | ¦ | + | [57] |
| Roe deer | Development of commercial forests | △ | ¦ | + | [184,185] | |
| Slovakia | Red deer | Favoring Picea abies L., Exclusion of broadleaved species | △ | * | / | [58,59] |
| United Kingdom | Red deer, Roe deer | Risk of plantation failure | △ | * | / | [104,105,106,107,108,109,110,111,114,115,116,117,118,119,120,122] |
| Fallow deer | Risk of plantation failure | △ | * | / | [112,113] | |
| Muntjac deer | Reduction of vegetative regeneration | △, Exotic species | * | / | [123,124,125] | |
| Sweden | Moose | Development of commercial forests, Exclusion of broadleaved species | △ | ** | + | [68,69,72,76,77,78,86,88,90,91,92,93,94,95,96,97,98,99,101] |
| Roe deer | Development of commercial forests, Exclusion of broadleaved species | △ | *** | / | [69,70,71,72,73,74,75,76,77,78,79,80] | |
| Fallow deer | Tree survival severely reduced | △, Hunting reserve | ¦ | / | [80] | |
| Finland | Moose | Development of commercial forests | △, Low-density planting scheme | ** | + | [61,62,63,64,65,66] |
| Denmark | Roe deer | Diminished growth in height | △ | ** | / | [60] |
| Latvia | Red deer, Roe deer | Development of commercial forests | △ | * | / | [67] |
| Lithuania | Red deer, Moose | Development of commercial forests | Harsh winters | * | / | [187] |
| Norway | Roe deer | Conservation of a protected species (Taxus baccata L.) | △ | ¦ | / | [188] |
| Italy | Red deer, Roe deer, chamois | Unwanted changes in tree composition, Reduction of broadleaved species and Abies alba Mill. | △ | ¦ | / | [126,127,128,129,130] |
| Slovenia | + | [134,135] | ||||
| Austria | Red deer, Roe deer, Chamois | Development of commercial forests, High financial losses | △ | **** | / | [137,138,139,140,141,142,143,144] |
| France | Red deer, Roe deer | Development of commercial forests | △ | ** | / | [145,146,147,148,149,150,151,152,153] |
| Germany | Red deer, Roe deer | Development of commercial forests High financial losses | △ | *** | / | [154,155,156,157,158,159,160,161,162,163,164,165,166,167] |
| Netherlands | Red deer, roe deer | Suppression of broadleaves | △ | * | / | [168,169] |
| Switzerland | Red deer, roe deer, chamois | Unwanted changes in tree composition, Reduction of broadleaved species and Abies alba Mill. | △ | * | / | [30,170,171,172,173,174,175] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hardalau, D.; Codrean, C.; Iordache, D.; Fedorca, M.; Ionescu, O. The Expanding Thread of Ungulate Browsing—A Review of Forest Ecosystem Effects and Management Approaches in Europe. Forests 2024, 15, 1311. https://doi.org/10.3390/f15081311
Hardalau D, Codrean C, Iordache D, Fedorca M, Ionescu O. The Expanding Thread of Ungulate Browsing—A Review of Forest Ecosystem Effects and Management Approaches in Europe. Forests. 2024; 15(8):1311. https://doi.org/10.3390/f15081311
Chicago/Turabian StyleHardalau, Darius, Codrin Codrean, Daniel Iordache, Mihai Fedorca, and Ovidiu Ionescu. 2024. "The Expanding Thread of Ungulate Browsing—A Review of Forest Ecosystem Effects and Management Approaches in Europe" Forests 15, no. 8: 1311. https://doi.org/10.3390/f15081311