Abstract
Although the effects of invasive alien plants on natural ecosystems are well known, the effects of specific plant species can vary across habitat types and disturbance intensity. This study was carried out to analyze the effects of Lantana camara on associated vegetation and soil physico-chemical properties at invaded and non-invaded sites across three different habitat types (forest edge, fallow land, and roadside) in central Nepal. We sampled 50 pairs of 5 m × 5 m (for shrub species) and 1 m × 1 m (for herbs species) plots at invaded and non-invaded sites in each habitat and recorded community variables for each species within the sampling plots for both wet (monsoon) and dry (pre-monsoon) seasons. Further, we collected soil samples from each quadrat and determined the soil physico-chemical properties. We recorded 137 species of flowering plants (119 from non-invaded and 97 from invaded plots) and classified them in accordance with life form/habit. In invaded sites, we found a significant decline in species diversity as indicated by the Simpson and Shannon diversity indices. Specifically, L. camara reduced the species richness, Simpson index, and Shannon diversity index by 36.84%, 11.84%, and 40.21%, respectively. Soil nutrients such as total nitrogen, soil organic carbon, and available phosphorus were significantly higher in invaded sites than non-invaded ones except for available potassium and soil pH. This study provided evidence that Lantana L. camara has a substantial impact on the understory plant community assemblage and the physico-chemical properties of soil. The results suggest that the protection of native plant community requires management of L. camara by implementing appropriate measures.
1. Introduction
Invasive alien plant species (IAPS) are the established alien species that intimidate native ecosystems and biodiversity [1]. They cause a significant threat to native ecosystem function and structure of plant communities, and are one of the major causes of biodiversity loss, that bring about changes in vegetation structure and degradation of environment [2,3]. In the changing climate, the introduction and aggressiveness of IAPS is increasing, particularly on different land uses. According to recent reports, IAPS have massive socioeconomic and environmental consequences around the world [4]. Invasive plant species affect soil properties [5], hydrology [6] and fire regimes [7] at the ecosystem level. They are known to reduce native species diversity and alter composition at the community level [8], though such impacts can be species- or site-specific. Conversely, there is also evidence of facilitation and co-occurrence of invasive and native plants because of positive interactions between plant species with similar habitat requirements and phonologies [9]. The mixed responses demonstrate that invasive plant species can have both positive and negative effects on ecosystems. As a result, more research is needed to investigate the impacts of IAPS to develop effective control strategies.
Biological invasion has been identified as one of the emerging threats to biodiversity and ecosystem services in Nepal [10], as well as one of the major underlying causes of habitat degradation in Nepal, alongside unsustainable harvesting practices, environmental pollution, overgrazing, and infrastructure development [11]. The number of IAPS in Nepal has increased over time and the extent of climatically suitable areas for most of Nepal’s IAPS is likely to expand and shift upslope in the future under climate change scenarios [12]. A recent review by [13] has revealed that there are 30 species of invasive plants, and their number has been increasing at an alarming rate. Despite the significant efforts by researchers to generate new knowledge, little is known about the biology and ecology of individual IAPS [14], their diversity [15], distribution [16], and impacts [17,18]. Therefore, ecological investigations in different regions are needed to either specify the status or impact of individual IAPS on richness and diversity of native plant communities. The problems of invasive species are widespread in both developed and developing countries, but the impacts could be greater in developing countries like Nepal. The Indian subcontinent has been experiencing altered precipitation patterns and intensified monsoon seasonality in the recent decades [19], and this can play a crucial role in regulating plant productivity [20], thereby enhancing plant invasion in the region.
We studied L. camara as it is considered one of the world’s worst invasive plant species [21]. It is the major invader of forests, pastures, and wetlands [22]. Lanata camara invasion in natural ecosystems has been linked to widespread loss of native plant species diversity due to native species recruitment restrictions and altered ecosystem structure and function [23]. However, there is few quantitative evidence to support these claims and more data on the patterns of species loss associated with L. camara invasion is urgently needed to inform management priorities. High reproductive potential, phenotypic plasticity, resilience to grazing pressure, allelopathy, and fire resistance are the factors that cause L. camara to become a successful invader [24]. Various studies have found that L. camara to be an invasive species in various regions around the world [25,26,27,28,29,30,31]. Despite the large number of studies on the effects of invasive species, few have investigated the effects of invasive species on species diversity and composition as well as multiple soil properties across habitat types [32]. It is crucial to investigate the effects on both species composition and soil properties because soil and plants influence each other and changes in vegetation due to invasive species would eventually result in changes in soil properties and these changes in soil properties would result in additional changes in vegetation [33]. Despite the weed’s widespread occurrence in Nepal quantitative data on the impacts and scale of the problems are inadequate to inform management decisions. Therefore, data from different regions are needed both to specify their status and to analyze their impacts on diversity and richness of native plants.
In this study we assessed the impact of Lantana camara on understorey community composition, diversity and soil physico-chemical properties in central Nepal. We hypothesize that invasion by L. camara alters the community assemblage, biotic interaction among co-existing species and reduces plant diversity and soil physico-chemical properties. To test this hypothesis, we sought to (1) assess the impact of L. camara on species richness and diversity of understorey plant species, and (2) analyze the soil physico-chemical properties at the invaded and non-invaded sites.
2. Materials and Methods
2.1. Study Area
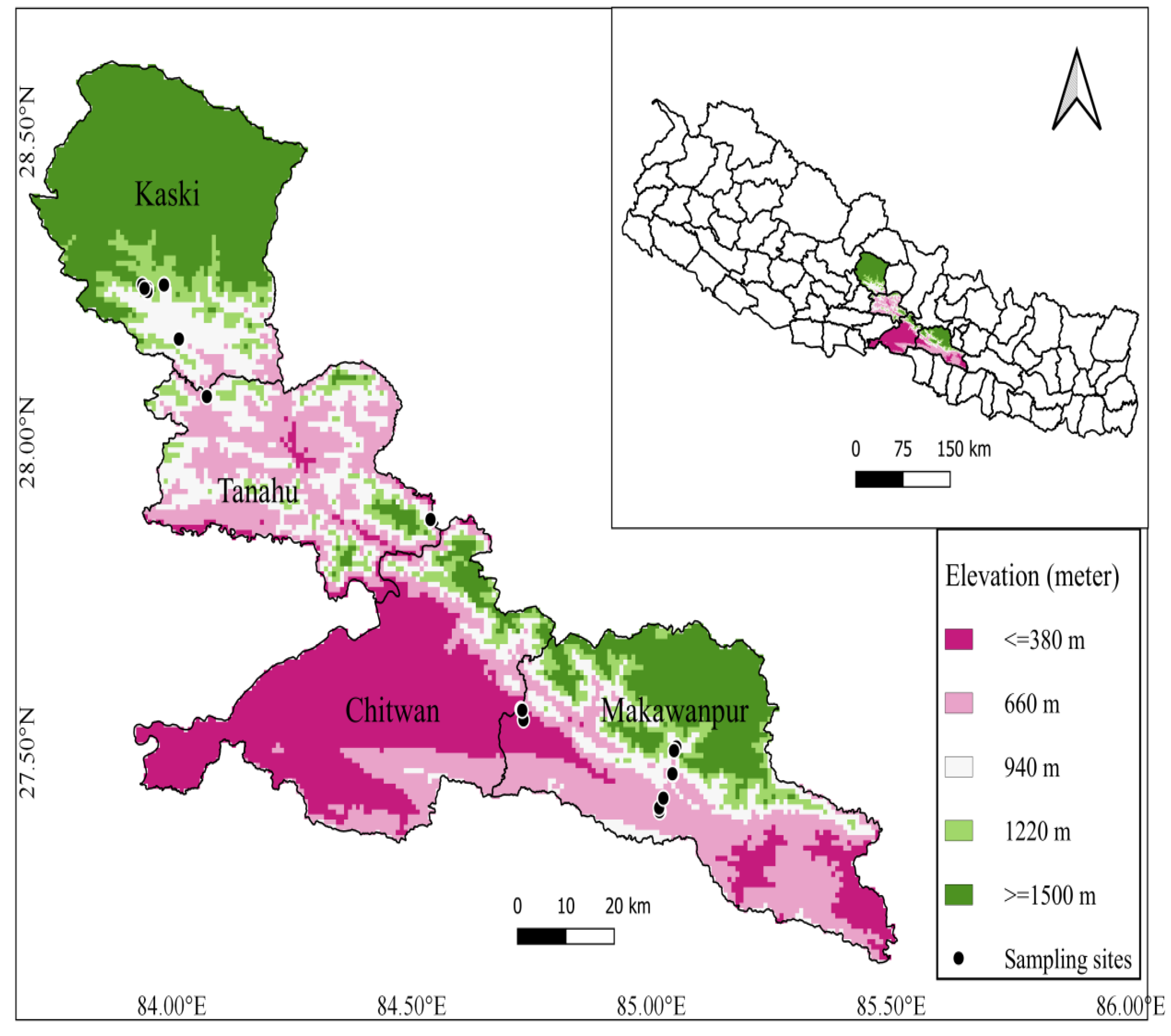
Study area lies in tropical and subtropical zones within four districts of central Nepal (Makawanpur, Chitwan, Tanahun, and Kaski). The elevation range of our study sites was 153 m (Chitwan) to 1490 m (Kaski) (Figure 1). The climate of the study area is a typical monsoon climate with four distinct seasons (monsoon, post-monsoon, winter, and pre-monsoon) marked by distinct dry and wet seasons, creating a seasonal variation in plant community assemblage. The average minimum and maximum temperature of study area ranges from 16 °C to 31 °C and average annual rainfall ranges from 2000 mm to 3800 mm. Forest of Shorea robusta Gaertn. extended throughout the flat land and it is replaced by riverine forest along river and streamside in tropical regions. Schima-Castanopsis, Pinus, and Alnus forests are the dominant vegetation of the subtropical regions in the study area.
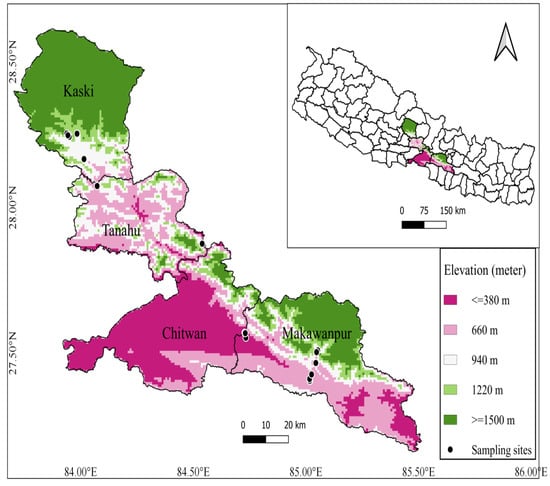
Figure 1.
Map of study area showing the sampling plots location.
2.2. Field Sampling
Three different habitat types were chosen for sampling, i.e., forest-edge, fallow-land, and roadside, as such microhabitats are highly vulnerable to plant invasions and the vegetation of these sites were dominated by L. camara in the study area. We defined forest-edge as the transitional zone between a forested area and an adjacent landscape such as grassland, agricultural land, or urban area, and fallow-land as the agricultural land left unplanted for a substantial time and roadside as the adjacent barren land to the road. These habitat types were chosen as they appeared to be highly invaded sites by L. camara while conducting a reconnaissance survey prior to this study. Plot locations for each habitat were chosen arbitrarily in topographically uniforms areas with 50 m distance between invaded and non-invaded sites. The ecological sampling was carried out from 28th September to 2nd October 2020 for the wet season (monsoon) and from 16th to 20th April 2021 for the dry season (pre-monsoon) to analyze if impacts vary seasonally. A field survey was conducted at two plot types: invaded and non-invaded. We carried out paired sampling of 5 m × 5 m (for shrub species) and 1 m × 1 m (for herbs species) for each habitat [34,35]. A total of 25 pairs of plots were sampled during the monsoon and 25 pairs during pre-monsoon. Plant species were counted, and their coverage was recorded in six cover classes, i.e., 1 (0%–5%), 2 (5%–25%), 3 (25%–50%), 4 (50%–75%) 5 (75%–95%), and 6 (95%–100%) [36] (Daubenmire, 1959), considering each plot as 100 percent based on a visual estimation method. Voucher specimens were collected except tree species (but saplings of trees were counted). Collected voucher specimens were identified at the National Herbarium and Plant Laboratories (KATH) by comparing them with identified specimens and consulting the literature [37]. The identified plants were categorized according to their life form (herbs, shrubs, and climbers).
2.3. Data Analysis
The species richness and Simpson and Shannon diversity indexes of vegetation were calculated. The species richness was defined as the number of species recorded in each sample plot.
Species diversity refers to the richness of species and their relative abundance in each region. To compare the species diversity between invaded and non-invaded sites, the Simpson and Shannon–Weiner diversity indexes were calculated.
The Shannon–Weiner Index was calculated by following equation [38]:
where Pi = ni/N, ni = average coverage of each individual species, N = total average coverage of all species.
Shannon Weiner Index (H) = −∑ Pi × ln Pi
Simpson Diversity Index:
where D = the Simpson diversity index, ni = the coverage of the ith species, and N = total coverage of all species.
D = 1 − ∑(ni − 1)/N (N − 1)
Statistical analysis was performed using the R 4.0.3 [39]. First, the normality of the data was tested. A significant value of the Shapiro–Wilk test was used to determine the normality of data (p ≥ 0.05) and the homogeneity of variance of the data was established with Leven’s test. The Simpson and Shannon diversity indexes, species richness, and soil physico-chemical properties in sites invaded and not invaded by L. camara during the pre-monsoon and monsoon seasons were analyzed by paired sampled t-test. Comparisons of the habitat types between invaded and non-invaded sites were performed using one-way ANOVA.
2.4. Soil Parameter Analysis
Soil samples were collected at a depth of 15 cm from four corners and a center, and they were mixed thoroughly to make single composite sample per plot. There were total of 100 samples (50 samples from the monsoon and 50 samples from the pre-monsoon season).
The soil samples were then left to air dry for two weeks in the shade in the lab. The soil samples were then ground and sieved through 2 mm wire mesh and a 0.5 mm mesh before being stored in airtight bags until laboratory analysis. The method outlined by [40] was used for determination of soil pH, the Walkey–Black method for soil organic carbon (SOC), and the micro-Kjeldahl method [41] for the determination of total nitrogen (TN). Available phosphorus and available potassium were estimated using a flame photometer and Olsen’s bicarbonate method, respectively, as described by [42,43].
3. Results
3.1. Species Composition and Diversity
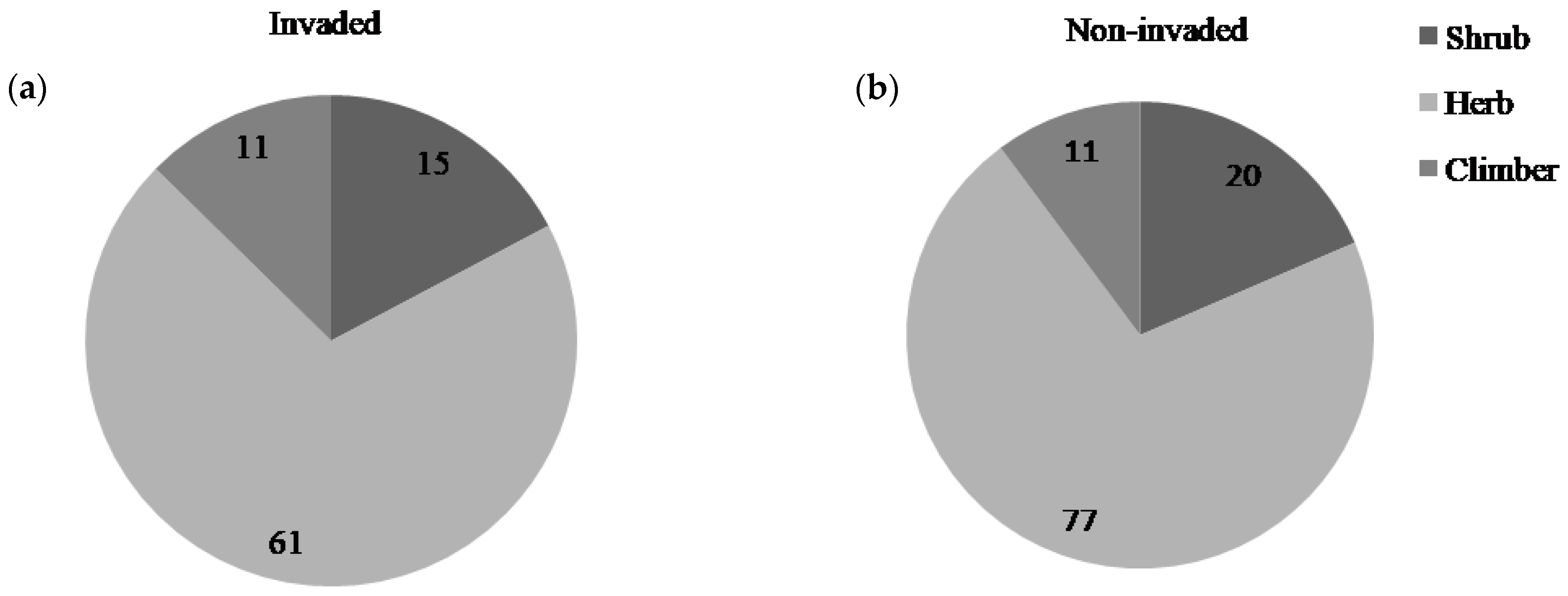
We found 137 species of flowering plants from 49 families, of which 119 were recorded in non-invaded sites and 97 in L. camara-invaded sites (Appendix A). Additionally, 79 plant species were present in both non-invaded and invaded sites. The number of herb species was high in non-invaded sites which decreased sharply compared to other life forms in invaded sites (Figure 2). Although all the growth forms (herb, shrub, and climber) showed changes in species composition, herbaceous species contributed the most to compositional changes. The plant family Poaceae was represented by the highest number of species (22), followed by Asteraceae (17) (Appendix A). Forty species were present only in non-invaded plots; and some of them were reported to have ethno-medicinal uses. Out of forty species, only eighteen species could successfully emerge from L. camara-invaded plots.

Figure 2.
Life form of plant species in (a) invaded and (b) non-invaded plots.
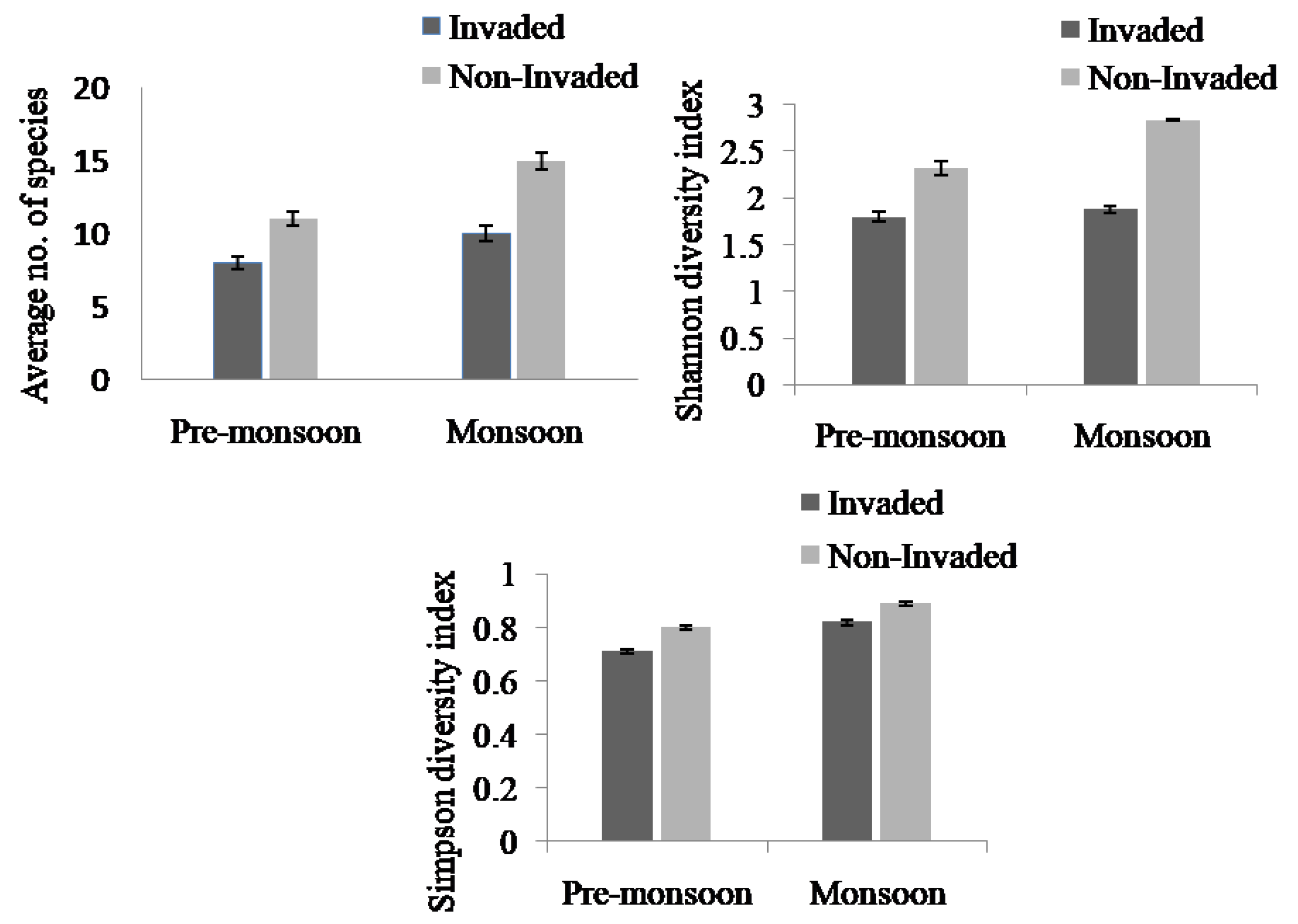
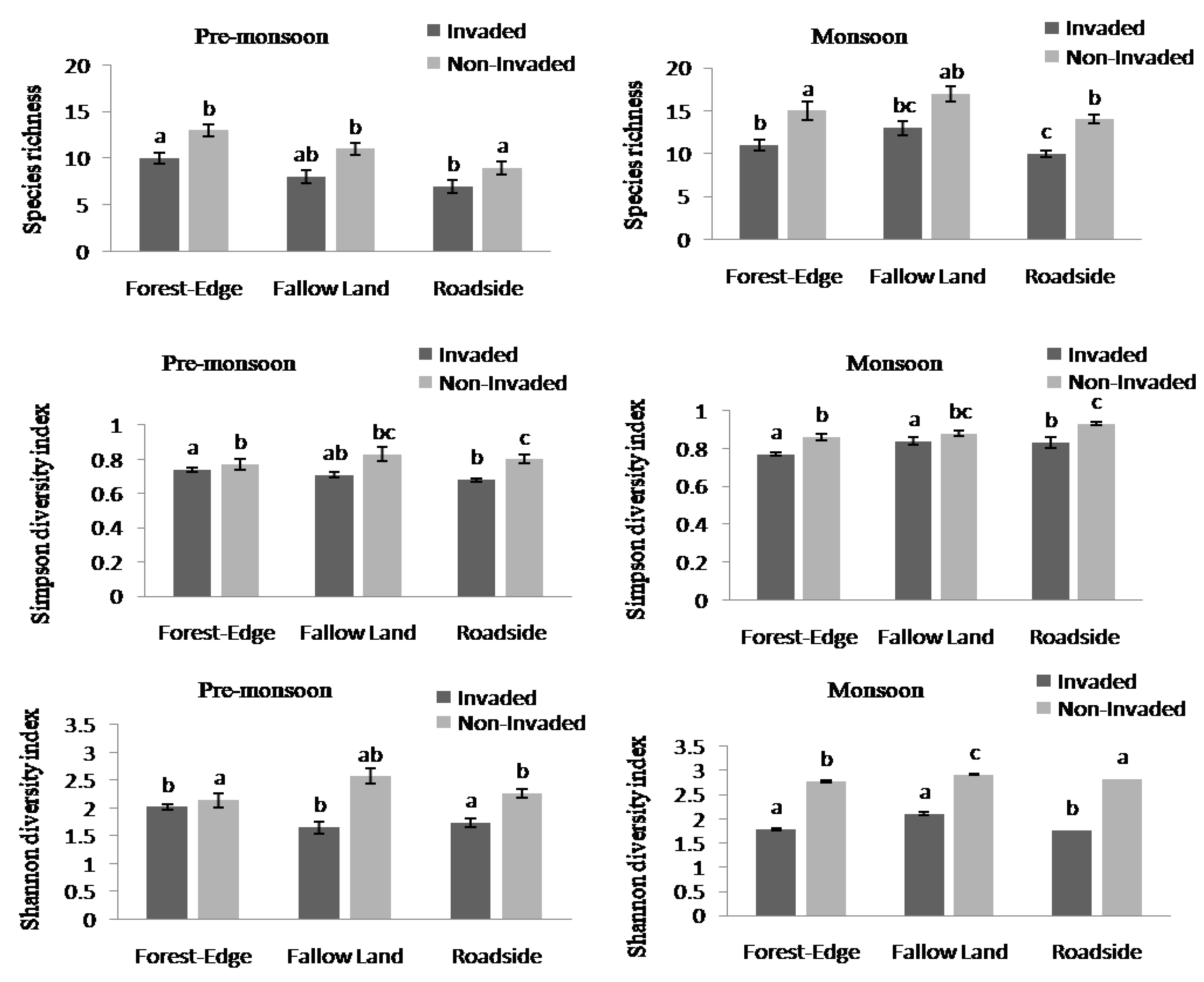
The species richness and diversity index between invaded and non-invaded sites differed significantly (Appendix B, Figure 2). The species richness and the Simpson and Shannon diversity indices of invaded sites decreased by 36.84%, 11.84%, and 40.21%, respectively, compared to the non-invaded sites. Non-invaded sites were composed of higher richness and diversity of plant species compared to invaded sites in both the pre-monsoon and monsoon seasons. There was a significant difference in species richness across the three habitat types (Figure 3, Appendix B). In the pre-monsoon season, the forest edge exhibited the highest species richness, followed by fallow land, and roadside; however, in the monsoon season, the fallow land exhibited the highest number of species followed by the forest edge and roadside.
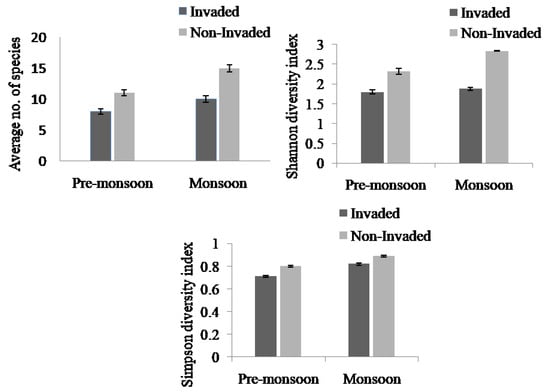
Figure 3.
Species richness, Simpson diversity index, and Shannon diversity index in Lantana camara-invaded and non-invaded sites (paired sample t-test). The values shown are means ± standard error.
3.2. Soil Physico-Chemical Characteristics
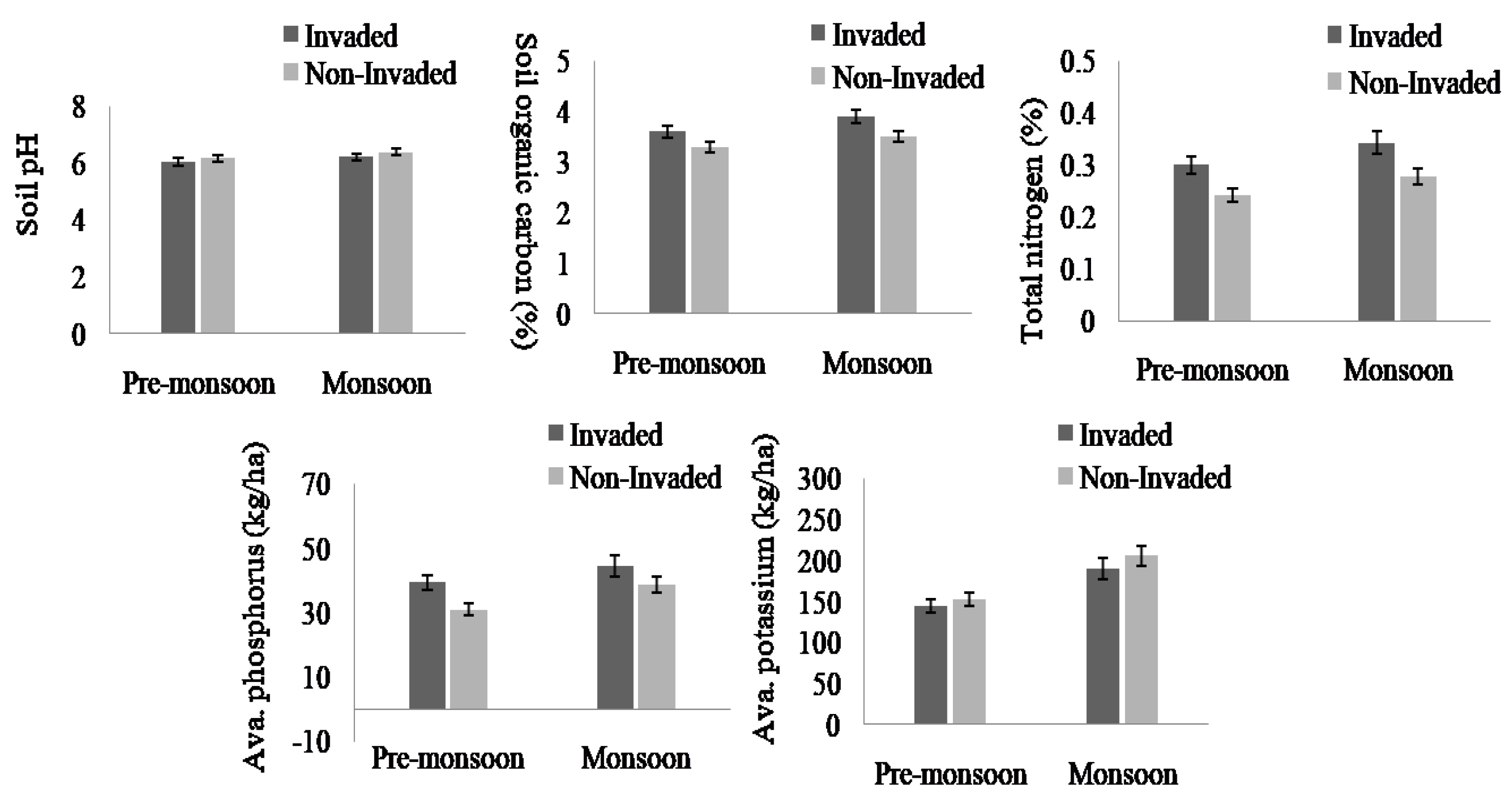
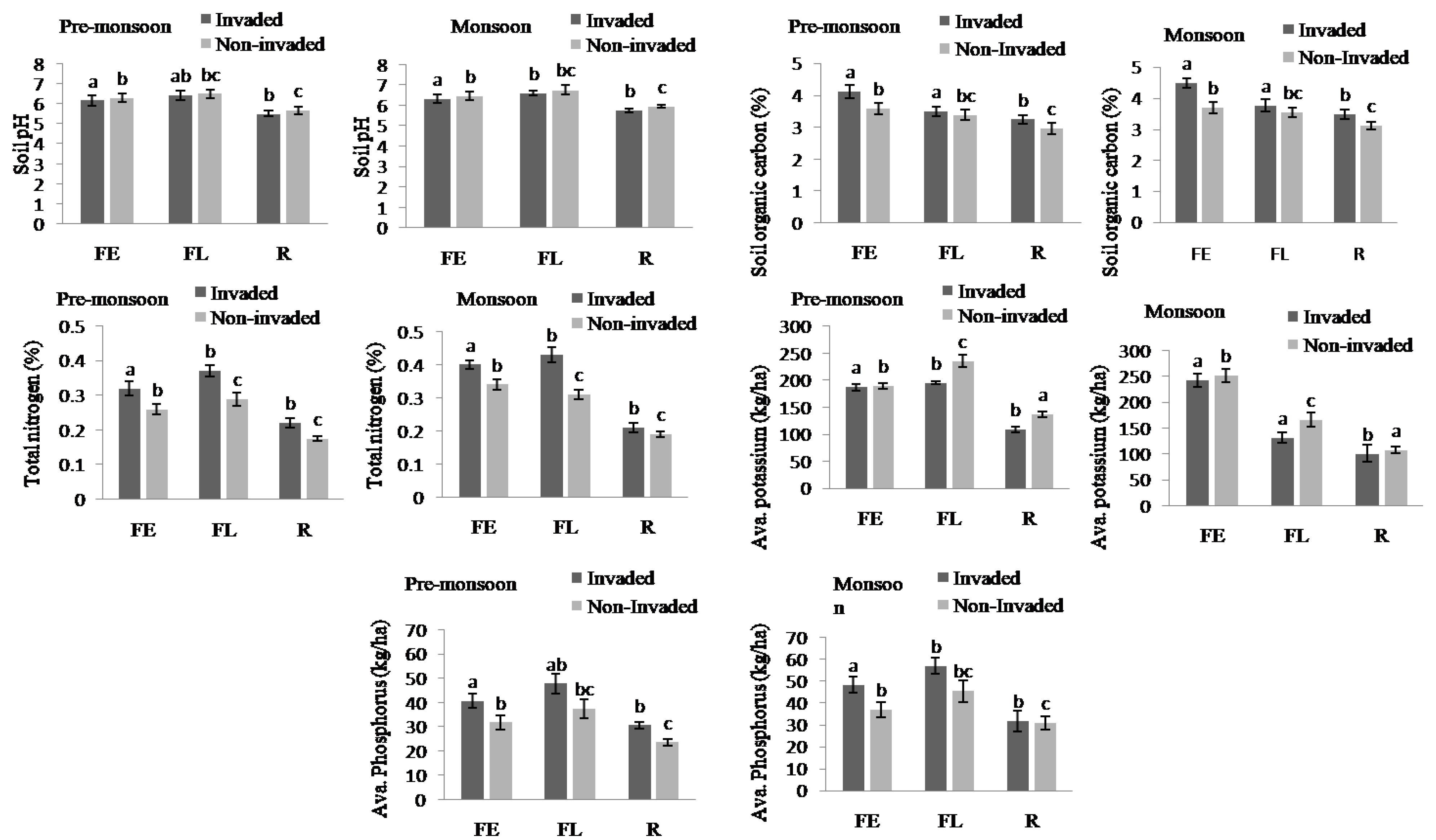
The soil nutrients were higher in invaded sites compared to non-invaded sites except available Potassium and soil pH in both the pre-monsoon and monsoon seasons (Figure 4, Appendix B). In total, the soil nutrients, like soil organic carbon, total nitrogen, and available phosphorus, of invaded sites increased by 11.61%, 28%, and 24.03%, respectively. Regarding the habitat type, total nitrogen, available phosphorus, available potassium, and soil pH were higher in fallow land followed by forest edge and roadside, while the total soil organic carbon was highest in forest edge and lowest in roadside (Figure 5 and Figure 6, Appendix B).
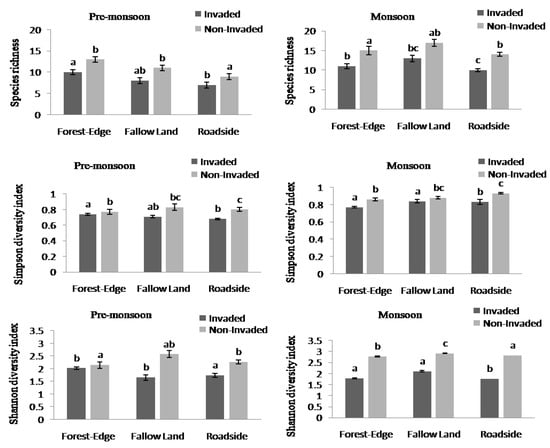
Figure 4.
Species richness, Simpson diversity index, and Shannon diversity index (one-way ANOVA test) in invaded and non-invaded sites across three habitat types. The values shown are means ± standard error. Means with different superscript letters are significantly different at p < 0.05.
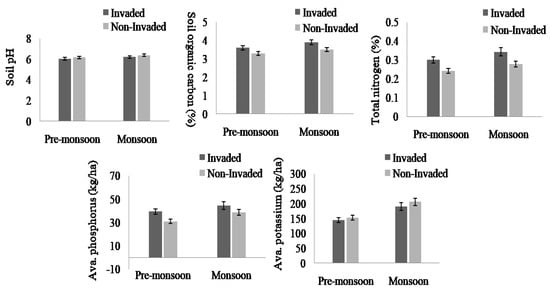
Figure 5.
Soil pH, total SOC, total N, available K, and available P in invaded and non-invaded plots as obtained from paired sample t-test results, the values shown are means (±standard error), p-values indicate significant different at p < 0.05.
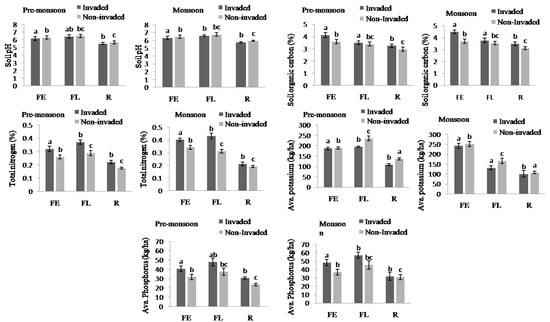
Figure 6.
Soil pH, total SOC, total N, available K, and available P at invaded and non-invaded sites of three habitat types. The values shown are means (±standard error). Means with different superscript letters are significantly different at p < 0.05.
4. Discussion
4.1. Species Composition and Diversity
Our results indicated that plant species decreased at invaded sites, including a sharp decrease in the number of herb species (Figure 2). The extensive cover produced by L. camara reduced the herbaceous species probably by restricting the amount and duration of light below its canopy. This could be because of the establishment of a photosynthetically inactive light regime at ground level [44,45]. Despite the levels of other resources, light limitation below a certain threshold can prevent species from surviving [46]. Species such as Tinospora sinensis, Sapium insigne, Calotropis gigantean, Solanum nigrum, Urena lobata, Woodfordia fruticosa, Anaphalis contorta, Buddleja asiatica, Callicarpa arborea, Cissampelos pareria, Elephantopus scaber, and Hydrocotyle sibthropioides were completely lost at invaded sites (Appendix A). Soil nutrients may also play a significant role in compositional difference in plant species between L. camara-invaded sites and non-invaded sites. It has been reported that the changes in litter and soil chemistry under the canopy of L. camara facilitate its invasion and displace the native species [47].
We reported that L. camara invasion is associated with a decrease in species diversity and richness. The ability of L. camara to adapt under various ecological conditions allows it to spread quickly, which causes a reduction in diversity and the development of its monocultures. The capacity of L. camara to establish rapidly, its tolerance to various environmental conditions and resistance to logging, burning, and climate change reduced the diversity and richness at the invaded sites [48,49]. Vegetation at the invaded sites had declined more significantly than that of non-invaded sites [50], and very few shade tolerant species stood out against L. camara invasion, that can grow taller than L. camara. Similar trends were seen in the riparian forests along the Nayar River, where invaded sites had a nearly 30% loss in species richness. Compared to non-invaded habitats, L. camara-invaded sites included significantly fewer plant species [22]. In regions where L. camara has invaded, the richness, diversity, structure, and composition of native plant species was reduced and degraded [51]. Our results indicated that the non-invaded sites comprised a higher richness and diversity of plant species compared to invaded sites in both the pre-monsoon and monsoon seasons, and this could be due to competitive exclusion [52] or altered habitat structure [53].
Although all the habitat-types were susceptible to invasion in our study, the diversity of understorey plants species were high in fallow land and forest edge compared to roadside, which indicated that the roadside habitat was highly affected by L. camara invasion compared to forest edge and fallow land. The lowest invasion effect was noticed in forest edge with its high species richness. This result is consistent with those of [54], who revealed that invasive species could not get enough light under the canopy of tree, hence they cannot spread rapidly in dense forests. Forest trees and vines often provide shade for invasive plant seedlings and slow down their growth [55]. Habitats near the roads were found to be more susceptible to invasion as the invasive species are conspicuous along roadsides, and roads are more anthropogenically disturbed and provide suitable microhabitats for many IAPS [56]. Further, we observed the seasonal changes in species richness across different habitat type—forest edge, fallow land, and roadside—and these variations could be attributed to various ecological and environmental factors including changes in microclimate [57], the emergence of opportunistic species [58], and changes in water availability [59].
4.2. Soil Physico-Chemical Characteristics
Invaded sites were composed of more acidic (low pH value) soil compared to non-invaded sites (higher pH value) throughout both the pre-monsoon and monsoon seasons. This might be related to the effect of podsolization (base depletion) [60]. Soil organic carbon, nitrogen, and phosphorus contents were higher in the invaded sites compared to the non-invaded sites. The positive correlations of L. camara cover with soil nutrients, except soil pH and K (Appendix B), indicated that soil nutrient increases with increased L. camara cover. The higher nutrient content in invaded sites may be related to the higher above-ground biomass and net primary production as well as faster growth rates than those of the native species being displaced as reported by [26]. Lantana camara-invaded sites were associated with higher levels of organic carbon in the soil compared to the non-invaded sites [61]. The dense monoculture of L. camara altered the microenvironment (pH, light, and temperature) beneath it and prevented the establishment and germination (or both) of associated species as reported by [62]. The allelopathic compounds released by the root of L. camara prevent the development and establishment of other native plant species, which reflects the competition between them [18,63].
The high biomass (mainly leaf litter) resulting from the taller plant present in invaded sites and no consumption of L. camara by livestock could be two possible explanations for the observed significant increase in soil nutrient at the L. camara-invaded sites compared to the non-invaded sites. Due to the deep root system of L. camara, it can absorb and use nutrients more quickly in deeper soil layers [64], a process known as “nutrient uplift” [65], which could increase the amount of phosphorus that was accessible in the surface soil. The higher total soil organic carbon, total nitrogen, and available phosphorus could be due to the abundance of invasive species which contribute to nutrient-rich higher litterfall and biomass [66,67], which eventually increase the organic carbon and nutrient pool in the soil. In other statements, invasive species prefer nutrient-rich soil and inoculate positive plant soil feedback which encourages further invasion in such habitats [68]. However, the results of the present study showed that availability of potassium is lower in the invaded sites than the non-invaded sites. These outcomes support the observation of [69], who reported a low level of potassium in invaded sites. This might be because L. camara produces many seeds with a high flowering rate, which causes the highly invaded areas to have low potassium levels [70,71].
5. Conclusions
This study provides evidence that L. camara invasion poses a substantial impact to species richness, community assemblage, and species diversity, as well as the soil properties. The study revealed that native plant species, like Artemisia indica, Woodfordia fruticosa, Urena lobata, and Achyranthes aspera, were reduced by L. camara invasion. The soil nutrient levels in most of the L. camara-invaded sites was high and were potentially increased by its invasion, which could allow more invasive plants to enter the area and could affect the ecosystem functions. These findings are highly important to biodiversity/forest managers and farmers. Further success of the L. camara invasion depends on soil properties like pH, SOC, N, P and K. Hence, proper utilization of land resources, control of deforestation, and land degradation are highly recommended to contend L. camara invasion. Further studies on the comparative physiological traits of invasive and native species and soil microbial dynamics would help refine our knowledge on plant invasion dynamics. Long-term monitoring on permanently established plots is also needed to understand the long-term impact of invasive species on vegetation dynamics to frame appropriate policy and management strategies to control plant invasion and its impacts.
Author Contributions
Conceptualization, A.T. and P.K.J.; Validation, C.B.B.; Formal analysis, C.B.B.; Investigation, C.K.P., A.T. and B.B.S.; Data curation, C.K.P. and C.B.B.; Writing—original draft, C.K.P.; Writing—review & editing, A.T. and B.B.S.; Supervision, B.B.S. and P.K.J.; Funding acquisition, P.K.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the United States Agency for International Development (USAID) Bureau of Food Security under the Cooperative Agreement No. AID-OAA-L-15-00001 as part of the Feed the Future Innovation Lab for Integrated Pest Management.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
| Name of Plants | Family | Invaded | Non-Invaded |
| Acacia catechu (L.f.) Wild | Fabaceae | - | + |
| Achyranthes aspera L. | Amaranthaceae | + | + |
| Acmella sp. | Asteraceae | - | + |
| Ageratum conyzoides L. | Asteraceae | + | + |
| Ageratum houstonianum Mill. | Asteraceae | + | + |
| Ageratina adenophora (Spreng.) R. King and H. Rob. | Asteraceae | + | + |
| Alternanthera phyloxeroides (Mart.) Griseb | Amaranthaceae | - | + |
| Anaphalis contorta (D. Don) Hook. f. | Asteraceae | - | + |
| Artemisia indica Willd. | Asteraceae | + | + |
| Arundo sp. | Poaceae | + | + |
| Asparagus racemosus Willd. | Asparagaceae | + | - |
| Axonopus Compressus (Sw.) P. Beauv. | Poaceae | - | + |
| Azadirachta indica A. Juss. | Meliaceae | + | - |
| Bidens pilosa L. | Asteraceae | + | + |
| Boehmeria sp. | Urticaceae | + | + |
| Bothriochloa ischaemum (L.) Keng | Poaceae | + | + |
| Bothriochloa pertusa (L.) A. Camus | Poaceae | - | + |
| Buddleja asiatica Lour. | Schrophulariaceae | - | + |
| Callicarpa arborea Roxb. | Verbenaceae | - | + |
| Callicarpa macrophylla Vahl | Verbenaceae | + | - |
| Calotropis gigantea (L.) Dryand | Apocynaceae | + | - |
| Cannabis sativa L. | Cannabaceae | + | + |
| Cardiospermum halicacabum L. | Sapindaceae | - | + |
| Castanopsis indica (Roxb.) Miq. | Fagaceae | - | + |
| Chromolaena odorata (L.) R. M. King and H. Rob. | Asteraceae | + | + |
| Chrysopogon aciculatus (Retz.) Trin. | Poaceae | - | + |
| Cissampelos pareria L. | Menispermaceae | - | + |
| Clerodendrum viscosum Vent. | Lamiaceae | + | + |
| Coccinia grandis (L.) Voigt | Cucurbitaceae | + | - |
| Colebrookea oppositifolia sm. | Lamiaceae | + | + |
| Colocasia esculenta (L.) Schott | Araceae | + | + |
| Commelina benghalensis (L.) | Commelinaceae | + | + |
| Conyza bonariensis (L.) Cronquist | Asteraceae | - | + |
| Corchorus aestuans L. | Tiliaceae | + | + |
| Crassocephalum crepidiodes (Benth.) S. Moore | Asteraceae | + | + |
| Cucumis callosus (Rottb.) Cogn. | Cucurbitaceae | + | - |
| Curcuma aromatica Salisb. | Zingiberaceae | - | + |
| Cyanotis cristata (L.) D. Don | Commelinaceae | - | + |
| Cynodon dactylon (L.) Pers. | Poaceae | + | + |
| Cynoglossum lanceolatum Forssk. | Boraginaceae | - | + |
| Cyperus esculentus L. | Cyperaceae | + | + |
| Cyperus rotundus L. | Cyperaceae | + | + |
| Cyperus sp. | Cyperaceae | - | + |
| Dalbergia sisso DC. | Fabaceae | + | + |
| Desmodium heterocarpon (L.) DC. | Fabaceae | - | + |
| Desmodium triflorum (L.) DC. | Fabaceae | + | + |
| Digitaria ciliaris (Retz.) Koeler | Poaceae | + | + |
| Digitaria setigera Roth | Poaceae | + | - |
| Dioscorea bulbifera L. | Dioscoreaceae | + | + |
| Drepanostachyum intermedium (Munro) Keng f. | Poaceae | - | + |
| Drymaria diandra Blume | Caryophyllaceae | + | + |
| Elephantopus scaber L. | Asteraceae | - | + |
| Eulaliopsis binata (Retz.) C. E. Hubb | Poaceae | + | + |
| Euphorbia hirta L. | Euphorbiaceae | + | + |
| Evolvulus nummularius (L.) L. | Convolvulaceae | + | + |
| Ficus hispida L. | Moraceae | + | + |
| Ficus semicordata Buch.-Ham. ex J. E. Sm. | Moraceae | + | - |
| Fimbristylis dichotoma (L.) Vahl. | Cyperaceae | + | + |
| Flemingia macrophylla (Willd.) Merr. | Fabaceae | + | - |
| Galinsoga perviflora Cav. | Asteraceae | + | + |
| Gnaphalium affine D. Don. | Asteraceae | - | + |
| Gonostegia pentandra (Roxb.) Miq. | Urticaceae | + | + |
| Holarrhena pubescens (Buch.-Ham.) Wall. ex G. Don | Apocynaceae | + | + |
| Hydrocotyle sibthorpioides Lam. | Apiaceae | - | + |
| Hyptis suaveolens (L.) Poit. | Lamiaceae | + | + |
| Imperata cylindrica (L.) P. Beauv. | Poaceae | + | + |
| Ipomoea nil (L.) Roth | Convolvulaceae | - | + |
| Ipomoea purpurea (L.) Roth | Convolvulaceae | + | - |
| Ipomoea quamoclit L. | Convolvulaceae | + | + |
| Justicia adhatoda L. | Acanthaceae | + | + |
| Justicia simplex D. Don. | Acanthaceae | + | + |
| Kyllinga brevifolia Rottb. | Cyperaceae | + | + |
| Lantana camara L. | Verbenaceae | + | + |
| Leea crispa Royen ex L. | Leeaceae | + | + |
| Leucaena leucocephala (Lam.) de Wit | Fabaceae | + | - |
| Leucas cephaloates (Roth) Spreng. | Lamiaceae | - | + |
| Lindernia crustacea (L.) F. Muell. | Schrophulariaceae | - | + |
| Maesa chisia Buch.-Ham. ex D. Don | Myrsinaceae | - | + |
| Melastoma malabathricum L. | Melastomataceae | + | + |
| Mikania micrantha Kunth. | Asteraceae | + | + |
| Momordica charantia L. | Cucurbitaceae | + | + |
| Mimosa pudica L. | Fabaceae | + | + |
| Mimosa rubicaulis Lam. | Fabaceae | - | + |
| Morus alba L. | Moraceae | + | + |
| Murraya koenigii (L.) Spreng | Rutaceae | + | + |
| Oplismenus hirtellus (L.) P.Beauv. | Poaceae | + | + |
| Oxalis corniculata L. | Oxalidaceae | + | + |
| Parthenium hysterophorus L. | Asteraceae | + | + |
| Paspalidium flavidum (Retz.) A. Camus | Poaceae | + | + |
| Paspalum scrobiculatum L. | Poaceae | + | + |
| Paspalum sp. | Poaceae | - | + |
| Persicaria capitata (Buch.-Ham. ex D. Don) H. Gross | Polygonaceae | + | + |
| Persicaria perfoliata (L.) H. Gross | Polygonaceae | - | + |
| Phragmites karka (Retz.) Trin. ex Steud | Poaceae | + | + |
| Phyllanthus urinaria L. | Euphorbiaceae | + | + |
| Phyllanthus virgatus G. Forst | Euphorbiaceae | + | + |
| Physalis angulata L. | Solanaceae | + | - |
| Pinus roxburghii Sarg. | Pinaceae | + | - |
| Piper longum L. | Piperaceae | + | - |
| Plumbago zeylanica L. | Plumbaginaceae | - | + |
| Pogonatherum paniceum (Lam.) Hack. | Poaceae | - | + |
| Polygonum persicaria L. | Polygonaceae | + | + |
| Polygonum sp. | Polygonaceae | + | + |
| Prunus persica (L.) Batsch. | Rosaceae | - | + |
| Reinwardtia indica Dumort. | Linaceae | - | + |
| Ricinus communis L. | Euphorbiaceae | - | + |
| Rubus ellipticus Sm. | Rosaceae | + | - |
| Rubus sp. | Rosaceae | + | + |
| Saccharum spontaneum L. | Poaceae | + | + |
| Salvia sp. | Lamiaceae | + | + |
| Sapium insigne (Royle) Benth. ex. Hook. f. | Euphorbiaceae | + | - |
| Saraca asoca (Roxb.) Willd. | Fabaceae | - | + |
| Schima wallichii (DC.) Korth. | Theaceae | - | + |
| Senna occidentalis (L.) Roxb. | Fabaceae | + | + |
| Senna tora (L.) Roxb. | Fabaceae | + | + |
| Setaria glauca (L.) P. Beauv. | Poaceae | + | + |
| Setaria pallidifusca (Schumach.) Stapf and C. E. Hubb. | Poaceae | + | + |
| Shorea robusta Gaertn. | Dipterocarpaceae | + | + |
| Sida acuta (L. fil.) Borss. Waalk. | Malvaceae | + | + |
| Sida cordifolia L. | Malvaceae | + | + |
| Sida rhombifolia L. | Malvaceae | + | + |
| Solanum aculeatissimum Jacq. | Solanaceae | + | + |
| Solanum nigrum L. | Solanaceae | + | - |
| Spermacoce alata Aubl. | Rubiaceae | + | + |
| Stephania glandulifera Miers | Menispermaceae | + | + |
| Sambucus sp. | Sambucaceae | + | - |
| Syzygium cumini (L.) Skeels | Myrtaceae | - | + |
| Tetrastigma serrulatum (Roxb.) Planch. | Vitaceae | + | + |
| Thysanolaena maxima (Roxb.) Kuntze | Poaceae | + | + |
| Tinospora sinensis (Lour.) Merr. | Menispermaceae | - | + |
| Tridax procumbens L. | Asteraceae | - | + |
| Triumfetta pilosa Roth. | Malvaceae | + | + |
| Urena lobata L. | Malvaceae | + | + |
| Urtica dioca L. | Urticaceae | - | + |
| Vetiveria sp. | Poaceae | + | + |
| Woodfordia fruticosa (L.) Kurz. | Lythraceae | + | + |
| Xanthium strumarium L. | Asteraceae | + | + |
Appendix B

Table A1.
Results of paired sample t-test comparing species richness and diversity between Lantana camara-invaded and non-invaded plots. Abbreviation: 1-D, Simpson diversity index; H’, Shannon diversity index; LI, Lantana camara-invaded; NI, non-invaded; p-value indicate significant different at p < 0.05.
Table A1.
Results of paired sample t-test comparing species richness and diversity between Lantana camara-invaded and non-invaded plots. Abbreviation: 1-D, Simpson diversity index; H’, Shannon diversity index; LI, Lantana camara-invaded; NI, non-invaded; p-value indicate significant different at p < 0.05.
| Season | Simpson Index (1-D) | Shannon Index (H’) | Species Richness | |||
|---|---|---|---|---|---|---|
| LI | NI | LI | NI | LI | NI | |
| Pre-monsoon | 0.72 ± 0.008 | 0.80 ± 0.008 | 1.80 ± 0.547 | 2.32 ± 0.737 | 8 ± 0.469 | 11 ± 0.476 |
| p-value | <0.001 | <0.001 | <0.001 | |||
| Monsoon | 0.81 ± 0.010 | 0.89 ± 0.009 | 1.88 ± 0.035 | 2.84 ± 0.013 | 11 ± 0.404 | 15 ± 0.564 |
| p-value | <0.001 | <0.001 | <0.001 | |||
Table A2.
Results of one-way ANOVA comparing species richness and diversity between Lantana camara-invaded and non-invaded plots of each habitat and plot types. The values shown are means (±standard error). Means with different superscript letters are significantly different at p < 0.05. Abbreviation: 1-D, Simpson diversity index; H’, Shannon diversity index; LI, Lantana camara-invaded; NI, non-invaded.
Table A2.
Results of one-way ANOVA comparing species richness and diversity between Lantana camara-invaded and non-invaded plots of each habitat and plot types. The values shown are means (±standard error). Means with different superscript letters are significantly different at p < 0.05. Abbreviation: 1-D, Simpson diversity index; H’, Shannon diversity index; LI, Lantana camara-invaded; NI, non-invaded.
| Season | Habitat | Shannon Index (H’) | Simpson Index (1-D) | Species Richness | |||
|---|---|---|---|---|---|---|---|
| LI | NI | LI | NI | LI | NI | ||
| Pre-monsoon | Forest-edge | 2.02 ± 0.05 b | 2.14 ± 0.13 a | 0.74 ± 0.01 a | 0.77 ± 0.03 b | 10 ± 0.55 a | 13 ± 0.65 b |
| Fallow land | 1.65 ± 0.10 b | 2.57 ± 0.14 ab | 0.71 ± 0.02 ab | 0.83 ± 0.04 bc | 8 ± 0.75 ab | 11 ± 0.59 b | |
| Roadside | 1.73 ± 0.07 a | 2.26 ± 0.071 b | 0.68 ± 0.01 b | 0.80 ± 0.02 c | 7 ± 0.7 b | 9 ± 0.7 a | |
| p-value | 0.011 | 0.048 | 0.007 | 0.008 | 0.009 | 0.003 | |
| F-value | 5.57 | 3.49 | 6.30 | 5.97 | 5.93 | 7.86 | |
| Monsoon | Forest-edge | 1.78 ± 0.02 a | 2.78 ± 0.02 b | 0.77 ± 0.01 a | 0.86 ± 0.016 b | 11 ± 0.59 b | 15 ± 1.07 a |
| Fallow land | 2.11 ± 0.03 a | 2.92 ± 0.008 c | 0.84 ± 0.02 a | 0.88 ± 0.016 bc | 13 ± 0.82 bc | 17 ± 0.90 ab | |
| Roadside | 1.76 ± 0.01 b | 2.82 ± 0.009 a | 0.83 ± 0.03 b | 0.93 ± 0.009 c | 10 ± 0.38 c | 14 ± 0.52 b | |
| p-value | <0.001 | <0.001 | 0.015 | 0.005 | 0.019 | 0.015 | |
| F-value | 76.72 | 55.33 | 5.13 | 6.82 | 4.76 | 5.10 | |
Table A3.
Paired sample t-test results of soil attributes of invaded and non-invaded plots. The values shown are means (±Standard error), p-value indicate significant different at p < 0.05, ns = not significant. Abbreviation: SOC, Soil organic carbon; N, Nitrogen; K, Potassium; P, Phosphorus; LI, Lantana camara invaded; NI, non-invaded.
Table A3.
Paired sample t-test results of soil attributes of invaded and non-invaded plots. The values shown are means (±Standard error), p-value indicate significant different at p < 0.05, ns = not significant. Abbreviation: SOC, Soil organic carbon; N, Nitrogen; K, Potassium; P, Phosphorus; LI, Lantana camara invaded; NI, non-invaded.
| Season | pH | SOC % | N % | K | P | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| LI | NI | LI | NI | LI | NI | LI | NI | LI | NI | |
| Pre-monsoon | 6.02 ± 0.139 | 6.15 ± 0.132 | 3.60 ± 0.119 | 3.29 ± 0.107 | 0.29 ± 0.016 | 0.24 ± 0.013 | 161.64 ± 8.55 | 185.52 ± 9.34 | 39.30 ± 2.21 | 30.62 ± 1.96 |
| p-value | ns | 0.04 | <0.001 | <0.001 | <0.001 | |||||
| Monsoon | 6.20 ± 0.113 | 6.38 ± 0.119 | 3.89 ± 0.128 | 3.43 ± 0.104 | 0.34 ± 0.022 | 0.27 ± 0.015 | 155.92 ± 14.45 | 172.78 ± 13.62 | 45.33 ± 3.13 | 37.61 ± 2.45 |
| p-value | ns | 0.003 | <0.001 | 0.026 | 0.05 | |||||
Table A4.
One way ANOVA results of soil attributes of invaded and non-invaded plots of each habitat and plot types. The values shown are means (±standard error). Means with different superscript letters are significantly different at p < 0.05. Abbreviations: SOC, soil organic carbon; N, nitrogen; K, Potassium; P, Phosphorus; Lantana camera-invaded; NI, non-invaded.
Table A4.
One way ANOVA results of soil attributes of invaded and non-invaded plots of each habitat and plot types. The values shown are means (±standard error). Means with different superscript letters are significantly different at p < 0.05. Abbreviations: SOC, soil organic carbon; N, nitrogen; K, Potassium; P, Phosphorus; Lantana camera-invaded; NI, non-invaded.
| Season | Habitat | pH | SOC % | N % | K | P | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| LI | N | LI | NI | LI | NI | LI | NI | LI | NI | ||
| Pre-monsoon | Forest-edge | 6.18 ± 0.24 a | 6.31 ± 0.21 b | 4.12 ± 0.20 a | 3.58 ± 0.17 b | 0.32 ± 0.02 a | 0.26 ± 0.02 b | 187.33 ± 6.21 a | 189.53 ± 5.64 b | 40.60 ± 2.93 a | 31.76 ± 2.92 b |
| Fallow land | 6.44 ± 0.22 ab | 6.51 ± 0.20 bc | 3.49 ± 0.14 a | 3.38 ± 0.16 bc | 0.37 ± 0.02 b | 0.29 ± 0.01 c | 195.29 ± 2.45 b | 235.92 ± 11.38 c | 47.81 ± 4.27 ab | 37.46 ± 3.88 bc | |
| Roadside | 5.51 ± 0.14 b | 5.69 ± 0.18 c | 3.24 ± 0.14 b | 2.96 ± 0.16 c | 0.22 ± 0.01 b | 0.17 ± 0.008 c | 109.04 ± 5.65 b | 137.16 ± 5.09 a | 30.58 ± 1.40 b | 23.5 ± 1.46 c | |
| p-value | 0.012 | 0.023 | 0.004 | 0.043 | <0.001 | <0.001 | <0.001 | <0.001 | 0.002 | 0.007 | |
| F-value | 5.50 | 4.50 | 7.23 | 3.64 | 18.86 | 16.74 | 89.64 | 41.84 | 8.53 | 6.22 | |
| Monsoon | Forest-edge | 6.31 ± 0.21 a | 6.47 ± 0.19 b | 4.49 ± 0.15 a | 3.54 ± 0.18 b | 0.40 ± 0.02 a | 0.34 ± 0.01 b | 242.45 ± 12.23 a | 252.08 ± 12.58 b | 48.47 ± 3.52 a | 37.0.7 ± 3.44 b |
| Fallow land | 6.60 ± 0.11 b | 6.76 ± 0.23 bc | 3.76 ± 0.19 a | 3.70 ± 0.14 bc | 0.43 ± 0.02 b | 0.31 ± 0.01 c | 131.15 ± 10.03 a | 166.26 ± 13.92 c | 57.16 ± 3.72 b | 45.58 ± 5.04 bc | |
| Roadside | 5.75 ± 0.09 b | 5.95 ± 0.07 c | 3.47 ± 0.15 b | 3.11 ± 0.13 c | 0.21 ± 0.01 b | 0.19 ± 0.009 c | 101.03 ± 16.43 b | 108.11 ± 6.80 a | 31.77 ± 4.63 b | 30.99 ± 2.94 c | |
| p-value | 0.002 | 0.011 | <0.001 | 0.034 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.042 | |
| F-value | 8.67 | 5.51 | 9.73 | 3.96 | 40.80 | 42.57 | 30.21 | 42.11 | 10.43 | 3.66 | |
References
- CBD. Alien species that threaten ecosystems, habits or species. In Convention on Biological Diversity; Report on Consultations Regarding International Standards; FAO: Rome, Italy, 2008. [Google Scholar]
- Downey, P.O.; Rirchardson, D.M. Alien plant invasions and native plant extinctions: A six-threshold framework. AoB Plants 2016, 8, plw047. [Google Scholar] [CrossRef] [PubMed]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinction. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef] [PubMed]
- IPBES. Summary for Policymakers of the Thematic Assessment Report on Invasive Alien Species and Their Control of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Roy, H.E., Pauchard, A., Stoett, P., Renard Truong, T., Bacher, S., Galil, B.S., Hulme, P.E., Ikeda, T., Sankaran, K.V., McGeoch, M.A., et al., Eds.; IPBES Secretariat: Bonn, Germany, 2023. [Google Scholar] [CrossRef]
- Gibbons, S.M.; Lekberg, Y.; Mummey, D.L.; Sangwan, N.; Ramsey, P.W.; Gilbert, J.A. Invasive plants rapidly reshape soil properties in a grassland ecosystem. MSystems 2017, 2, e00178-16. [Google Scholar] [CrossRef] [PubMed]
- Le Maitre, D.C.; Richardson, D.M.; Chapman, R.A. Alien plant invasions in South Africa: Driving forces and the human dimension: Working for water. S. Afr. J. Sci. 2004, 100, 103–112. [Google Scholar]
- Brooks, M.L.; D’antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; DiTomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of invasive alien plants on fire regimes. BioScience 2004, 54, 677–688. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Rodriguez, L.F. Can invasive species facilitate native species? Evidence of how, when, and why these impacts occur. Biol. Invasions 2006, 8, 927–939. [Google Scholar] [CrossRef]
- Shrestha, B.B. Management of invasive alien plants in Nepal: Current practices and future prospects. In Tropical Ecosystems: Structure, Functions and Challenges in the Face of Global Change; Springer: Berlin/Heidelberg, Germany, 2019; pp. 45–68. [Google Scholar]
- Chaudhary, R.P.; Uprety, Y.; Rimal, S.K. Deforestation in Nepal: Causes, consequences and responses. Biol. Environ. Hazards Risks Disasters 2016, 12, 335–372. [Google Scholar]
- Shrestha, U.B.; Shrestha, B.B. Climate change amplifies plant invasion hotspots in Nepal. Divers. Distrib. 2019, 25, 1599–1612. [Google Scholar] [CrossRef]
- Shrestha, B.B.; Shrestha, K.K. Invasions of alien plant species in Nepal: Patterns and process. Invasive Alien Species Obs. Issues Around World 2021, 2, 168–183. [Google Scholar] [CrossRef]
- Maharjan, S.; Joshi, S.; Shrestha, B.B.; Devkota, A.; Jha, P.K. Life History Traits and Invasion Success of Parthenium hysterophorus L. in Kathmandu Valley, Nepal. Nepal J. Sci. Technol. 2014, 15, 31–38. [Google Scholar] [CrossRef]
- Bhattarai, K.R.; Maren, I.E.; Subedi, S.C. Biodiversity and invasibility: Distribution patterns of invasive plant species in the Himalayas, Nepal. J. Mt. Sci. 2014, 11, 688–696. [Google Scholar] [CrossRef]
- Shrestha, B.B.; Pokhrel, K.; Paudel, N.; Poudel, S.; Shabbir, A.; Adkins, S.W. Distribution of Parthenium hysterophorus and one of its biological control agents (Coleoptera: Zygogramma bicolorata) in Nepal. Weed Res. 2019, 59, 467–478. [Google Scholar] [CrossRef]
- Bhatta, S.; Joshi, L.R.; Shrestha, B.B. Distribution and impact of invasive alien plant species in Bardia National Park, western Nepal. Environ. Conserv. 2020, 47, 197–205. [Google Scholar] [CrossRef]
- Thapa, L.B.; Kaewchumnong, K.; Sinkkonen, A.; Sridith, K. “Soaked in rainwater” effect of Ageratina adenophora on seedling growth and development of native tree species in Nepal. Flora 2020, 263, 151554. [Google Scholar] [CrossRef]
- Roxy, M.K.; Ritika, K.; Terray, P.; Murtugudde, R.; Ashok, K.; Goswami, B.N. Drying of Indian subcontinent by rapid Indian Ocean warming and a weakening land-sea thermal gradient. Nat. Commun. 2015, 6, 7423. [Google Scholar] [CrossRef]
- Post, A.K.; Knapp, A.K. The importance of extreme rainfall events and their timing in a semi-arid grassland. J. Ecol. 2020, 108, 2431–2443. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species A Selection from the Global Invasive Species Database; The Invasive Species Specialist Group (ISSG): Rome Italy, 2000. [Google Scholar]
- Dobhal, P.K.; Kohli, R.K.; Batish, D.R. Evaluation of impact of Lantana camara L. invasion, on four major woody shrubs, along Nayar river of Pauri Garhwal in Uttarakhand Himalaya. Int. J. Biodivers. Conserv. 2010, 2, 166–172. [Google Scholar]
- Gooden, B.; French, K.; Turner, P.J.; Downey, P.O. Impact threshold for an alien plant invader, Lantana camara L., on native plant communities. Biol. Conserv. 2009, 142, 2631–2641. [Google Scholar] [CrossRef]
- Bhakat, R.K.; Maiti, P.P. Invasiveness and allelopathy as a threat to biodiversity. In Proceedings of the International Seminar on Multidisciplinary Approaches in Angiosperm Systematics, Kalyani, India, January 2012; pp. 748–751. [Google Scholar]
- Sharma, G.P.; Raghubanshi, A.S. Lantana invasion alters soil nitrogen pools and processes in the tropical dry deciduous forest of India. Appl. Soil Ecol. 2009, 42, 134–140. [Google Scholar] [CrossRef]
- Fan, L.; Chen, Y.; Yuan, J.; Yang, Z. The effect of Lantana camara Linn. invasion on soil chemical and microbiological properties and plant biomass accumulation in southern China. Geoderma 2010, 154, 370–378. [Google Scholar] [CrossRef]
- Osunkoya, O.O.; Perrett, C. Lantana camara L. (Verbenaceae) invasion effects on soil physicochemical properties. Biol. Fertil. Soils 2011, 47, 349–355. [Google Scholar] [CrossRef]
- Simba, Y.R.; Kamweya, A.M.; Mwangi, P.N.; Ochora, J.M. Impact of the invasive shrub, Lantana camara L. on soil properties in Nairobi National Park, Kenya. Int. J. Biol. Divers. Conserv. 2013, 5, 803–809. [Google Scholar]
- Ruwanza, S.; Shackleton, C.M. Effects of the invasive shrub Lantana camara on soil properties in the Eastern Cape, South Africa. Weed Biol. Manag. 2016, 16, 67–79. [Google Scholar] [CrossRef]
- Mahla, N.; Mlambo, D. Influence of two co-occurring invasive plant species on resident woody species and surface soil properties in Chipinge Safari Area, Zimbabwe. Trop. Ecol. 2019, 60, 129–139. [Google Scholar] [CrossRef]
- Ruwanza, S. Effects of Lantana camara invasion on vegetation diversity and composition in the Vhembe Biosphere Reserve, Limpopo Province of South Africa. Sci. Afr. 2020, 10, e00610. [Google Scholar] [CrossRef]
- Chacón, N.; Herrera, I.; Flores, S.; González, J.A.; Nassar, J.M. Chemical, physical, and biochemical soil properties and plant roots as affected by native and exotic plants in Neotropical arid zones. Biol. Fertil. Soils 2009, 45, 321–328. [Google Scholar] [CrossRef]
- Vitousek, P.M. Biological invasions and ecosystem processes: Towards an integration of population biology and ecosystem studies. Oikos 1990, 57, 7–13. [Google Scholar] [CrossRef]
- Kershaw, K.A. Quantitative and Dynamic Plant Ecology, 2nd ed.; Arnold, E., Krebs, C.J., Eds.; Ecological Methodology (No. QH541. 15. S72. K74 1999); Harper & Row: New York, NY, USA, 1973. [Google Scholar]
- Misra, R. Ecology Workbook; Oxford and IBH Publishing Company: Calcutta, India, 1968. [Google Scholar]
- Daubenmire, R. A canopy-coverage method of vegetation analysis. Northwest Sci. 1959, 33, 43–64. [Google Scholar]
- KATH (National Herbarium & Plant Laboratories). Flora of Nepal. 2019. Available online: https://kath.gov.np/Flora_of_Nepal (accessed on 15 April 2019).
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004; 256p. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Gupta, P.K. Methods in Environmental Analysis: Water, Soil and Air, 1st ed.; Updesh Purohit for Agrobios, India Jodhpur Agro House: Jodhpur, India, 2004; pp. 47–48. [Google Scholar]
- Black, C.A.; Evans, D.D.; White, J.L. Methods of soil analysis: Chemical and microbiological properties. Physical and mineralogical properties including statistics of measurement and sampling. Science 1965, 151, 982–983. [Google Scholar]
- Olsen, S.R.; Sommers, L.E. Phosphorous. In Methods of Soil Analysis, Part 2, 2nd ed.; Page, A.L., Ed.; Chemical and Microbiological Properties, ASA-SSSa, Inc.: Madison, WI, USA, 1982. [Google Scholar]
- NARC. Methods of Soil Sample Collection and Analysis; Soil Science Division, Council of National Agriculture Research Center: Khumaltar, Nepal, 2013; 167p.
- Fetcher, N.; Strain, B.R.; Oberbauer, S.F. Effects of light regime on the growth, leaf morphology, and water relations of seedlings of two species of tropical trees. Oecologia 1983, 58, 314–319. [Google Scholar] [CrossRef]
- Turton, S.M.; Duff, G.A. Light environments and floristic composition across an open forest-rainforest boundary in northeastern Queensland. Aust. J. Ecol. 1992, 17, 415–423. [Google Scholar] [CrossRef]
- Tilman, D. Resource Competition and Community Structure; Princeton University Press: Princeton, NJ, USA, 1982; Volume 28, pp. 1043–1045. [Google Scholar] [CrossRef]
- Sharma, G.P.; Raghubanshi, A.S. Tree population structure, regeneration and expected future composition at different levels of L. camara L. invasion in the Vindhyan tropical dry deciduous forest of India. Lyonia 2006, 11, 25–37. [Google Scholar]
- Taylor SKumar, L.; Reid, N.; Kriticos, D.J. Climate change and the potential distribution of an invasive shrub, Lantana camara L. PLoS ONE 2012, 7, e35565. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, Y.; Peng, S.; Zobel, K. Climate warming may facilitate invasion of the exotic shrub Lantana camara. PLoS ONE 2014, 9, e105500. [Google Scholar] [CrossRef]
- Dobhal, P.K.; Kohli, R.K.; Batish, D.R. Impact of Lantana camara L. invasion on riparian vegetation of Nayar region in Garhwal Himalayas (Uttarakhand, India). J. Ecol. Nat. Environ. 2011, 3, 11–22. [Google Scholar]
- Aravindhan, V.; Rajendran, A. Impact of invasive species Lantana camara (L.) on the vegetation of Velliangiri Hills, the Southern Western Ghats, India. Glob. J. Environ. Res. 2014, 8, 35–40. [Google Scholar]
- Vila, M.; Weiner, J. Are invasive plant species better competitors than native plant species?—Evidence from pair-wise experiments. Oikos 2004, 105, 229–238. [Google Scholar] [CrossRef]
- Levine, J.M.; D’Antonio, C.M. Elton revisited: A review of evidence linking diversity and invasibility. Oikos 1999, 87, 15–26. [Google Scholar] [CrossRef]
- Rouw, A.D. Chromolaena odorata in the farming systems of South-West Côte d’Ivoire. In Proceedings of the Distribution, Ecology and Management of Chromolaena odorata, 1996; ORSTOM, ICRAF and University of Guam, Mangilao, GUAM: Guam, Micronesia, 1996; pp. 76–87. [Google Scholar]
- Tjitrosemito, S. The management of Chromolaena odorata. In Proceedings of the Third International Chromolaena Workshop on Distribution, Ecology and Management of Chromolaena odorata, 1996; Robinson, H., Ed.; ORSTOM, ICRAF and University of Guam, Mangilao, GUAM: Guam, Micronesia, 1996; pp. 135–142. [Google Scholar]
- Adhikari, A.; Subedi, A.; Tiwari, A.; Shrestha, B.B. Impacts of road on plant invasions in the Middle Mountain region of central Nepal. J. Mt. Sci. 2024, 21, 619–632. [Google Scholar] [CrossRef]
- Starfinger, U.; Kowarik, I.; Rode, M.; Schepker, H. From desirable ornamental plant to pest to accepted addition to the Flora?—The perception of an alien tree species through the centuries. Biol. Invasions 2003, 5, 323–335. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Burton, P.J. Edge influence on forest structure and composition in fragmented landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- Guo, Q.; Brown, J.H. Temporal fluctuations and experimental effects in desert plant communities. Oecologia 1996, 107, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Osunkoya, O.O.; Perrett, C.; Fernando, C. Population viability analysis models for Lantana camara L. (Verbenaceae): A weed of national significance. In Proceedings of the 17th Australasian Weeds Conference, Christchurch, New Zealand, 26–30 September 2010; Zydenbos, S.M., Ed.; New Zealand Plant Protection Society: Auckland, New Zealand, 2010; pp. 91–94. [Google Scholar]
- Sharma, G.P.; Raghubanshi, A.S. Effect of Lantana camara L. cover on plant species depletion in the Vindhyan tropical dry deciduous forest of India. Appl. Ecol. Environ. Res. 2007, 5, 109–121. [Google Scholar] [CrossRef]
- Fu, D.; Wu, X.; Huang, N.; Duan, C. Effects of the invasive herb Ageratina adenophora on understory plant communities and tree seedling growth in Pinus yunnanensis forests in Yunnan, China. J. For. Res. 2018, 23, 112–119. [Google Scholar] [CrossRef]
- Castillo, J.M.; Leira-Doce, P.; Carrión-Tacuri, J.; Munoz-Guacho, E.; Arroyo-Solís, A.; Curado, G.; Tye, A. Contrasting strategies to cope with drought by invasive and endemic species of Lantana in Galapagos. Biodivers. Conserv. 2007, 16, 2123–2136. [Google Scholar] [CrossRef]
- Dassonville, N.; Vanderhoeven, S.; Gruber, W.; Meerts, P. Invasion by Fallopia japonica increases topsoil mineral nutrient concentrations. Ecoscience 2008, 14, 230–240. [Google Scholar] [CrossRef]
- Kumar, M.; Verma, A.K.; Garkoti, S.C. Lantana camara and Ageratina adenophora invasion alter the understory species composition and diversity of chir pine forest in central Himalaya, India. Acta Oecologica 2020, 109, 103–115. [Google Scholar] [CrossRef]
- Kumar, M.; Garkoti, S.C. Functional traits, growth patterns, and litter dynamics of invasive alien and co-occurring native shrub species of chir pine forest in the central Himalaya, India. Plant Ecol. 2021, 222, 723–735. [Google Scholar] [CrossRef]
- Vitti, S.; Pellegrini, E.; Casolo, V.; Trotta, G.; Boscutti, F. Contrasting responses of native and alien plant species to soil properties shed new light on the invasion of dune systems. J. Plant Ecol. 2020, 13, 667–675. [Google Scholar] [CrossRef]
- Chatanga, P. Impacts of the Alien Species L. camara L. on Vegetation in Northern Gonarezhou National Park, Zimbabwe. Master’s Thesis, University of Zimbabwe, Harare, Zimbabwe, 2007; pp. 1–84. [Google Scholar]
- Thomas, S.E.; Ellison, C.A. A century of classical biological control of Lantana camara: Can pathogens make a significant difference. In Proceedings of the X International Symposium on Biological Control of Weeds, Bozeman, MT, USA, 4–14 July 1999; Montana State University: Bozeman, MT, USA, 2000; Volume 4, pp. 97–104. [Google Scholar]
- ISSG. One Hundred of the World Worst Invasive Allien Species. A Selection from the Global Invasive Database; ISSG: Auckland, New Zealand, 2006. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).