
Quantifying Blowdown Disturbance in Overstory Retention Patches in Managed Nothofagus pumilio Forests with Variable Retention Harvesting

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
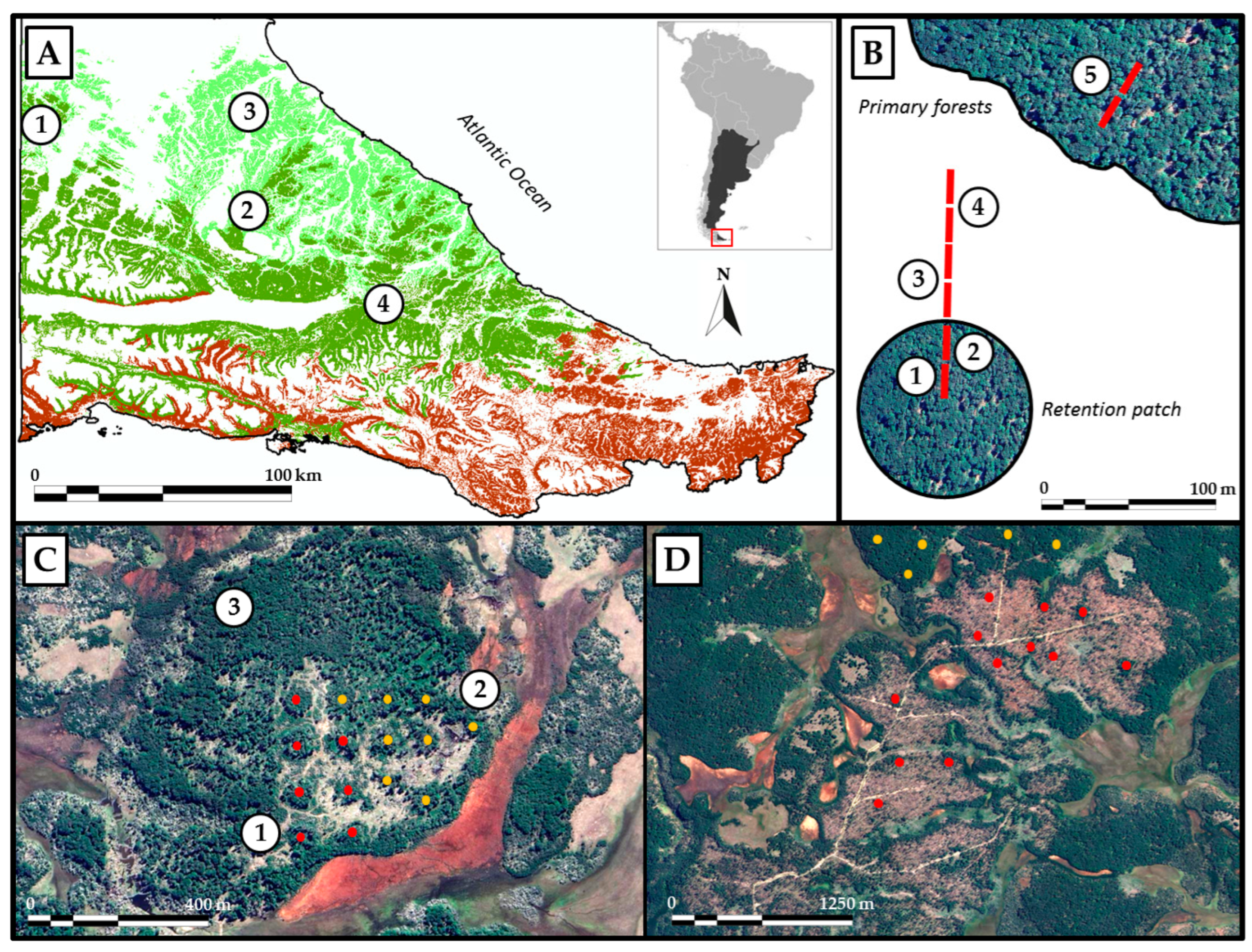
2.1. Sampling Design at San Justo Ranch
2.2. Sampling Design at Rivadavia Ranch
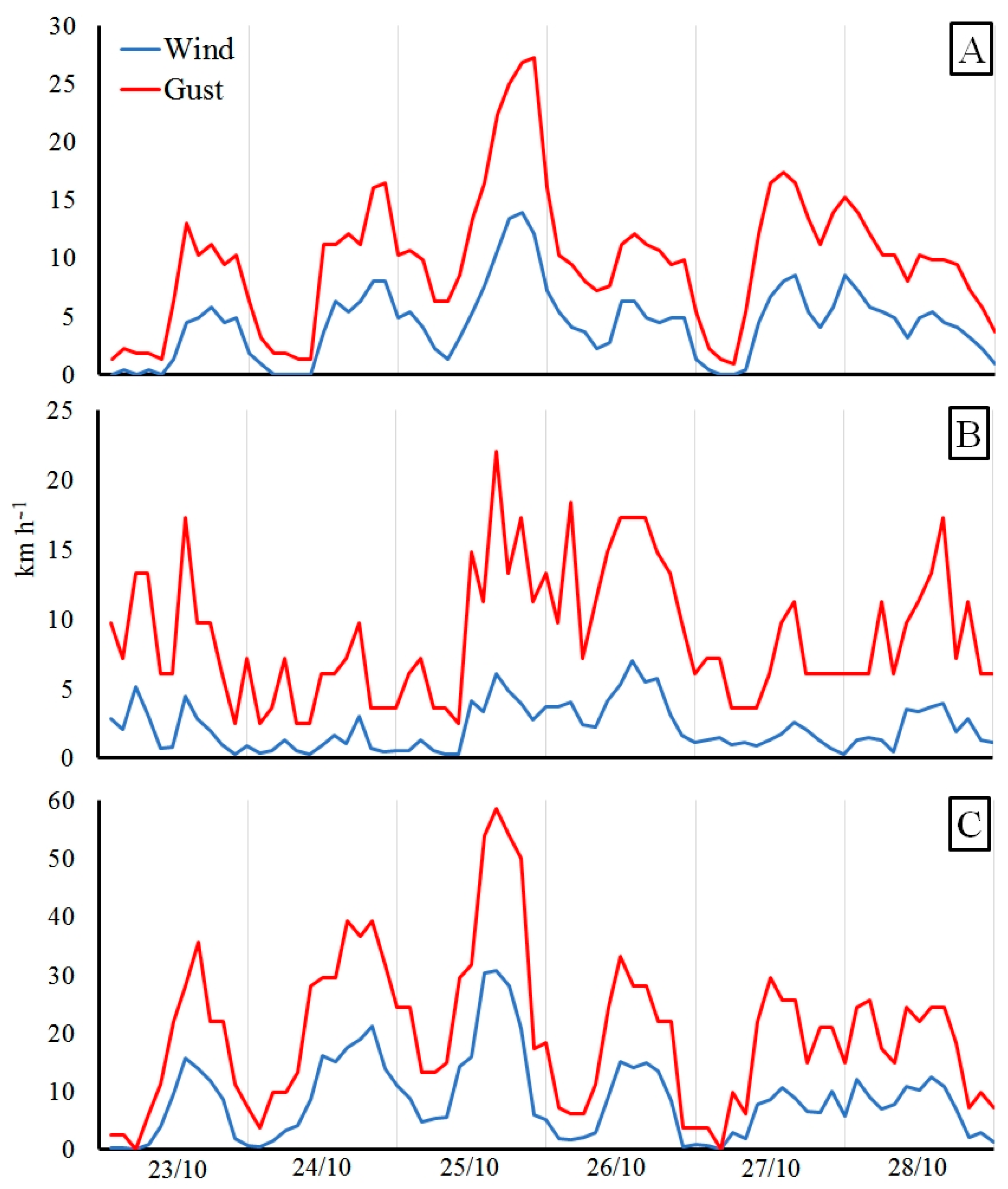
2.3. Characterization of the Windstorm Event
2.4. Statistical Analyses
3. Results
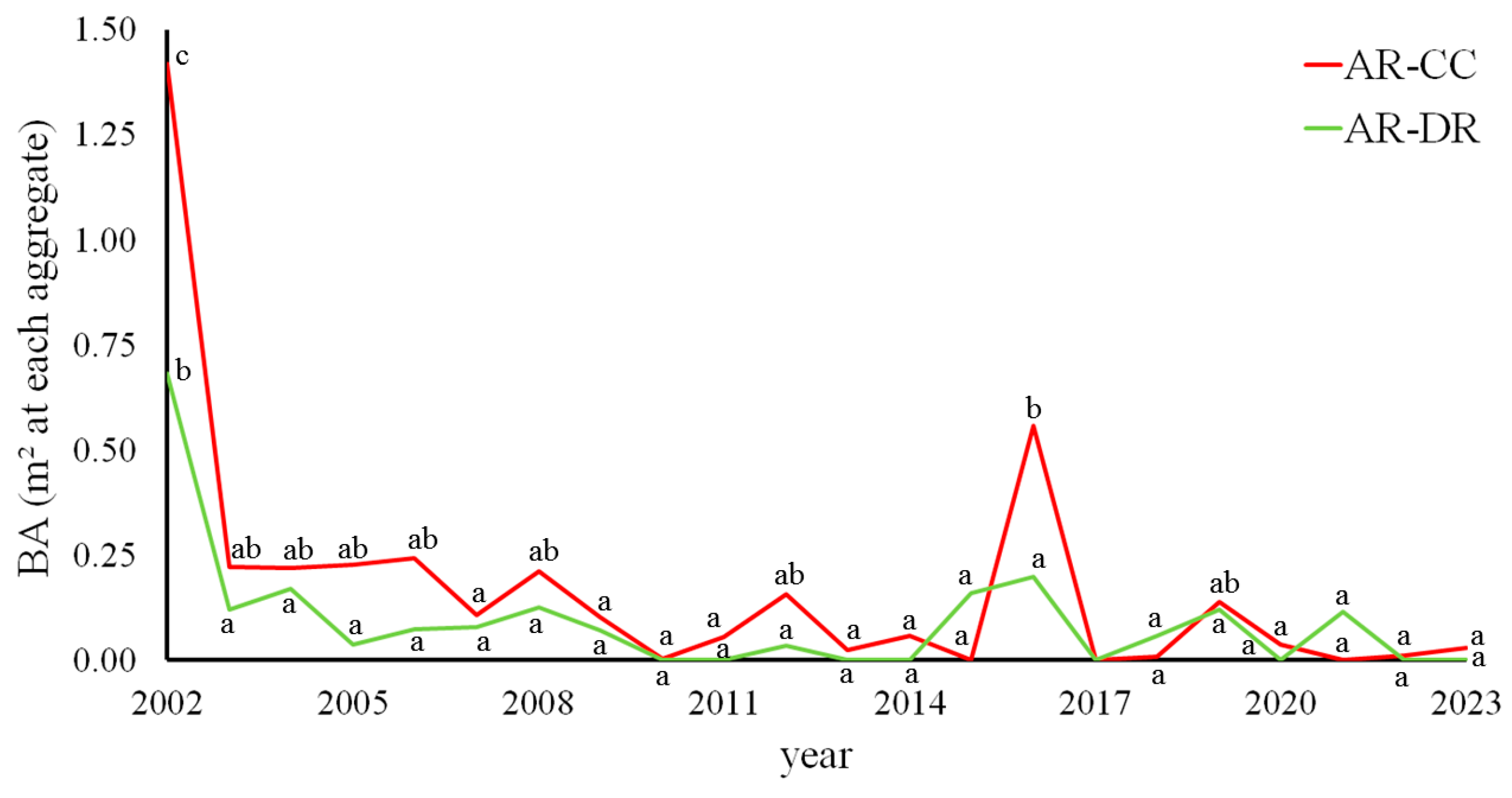
3.1. Long-Term Stability of Retention Patches in Variable Retention Harvesting at San Justo Ranch
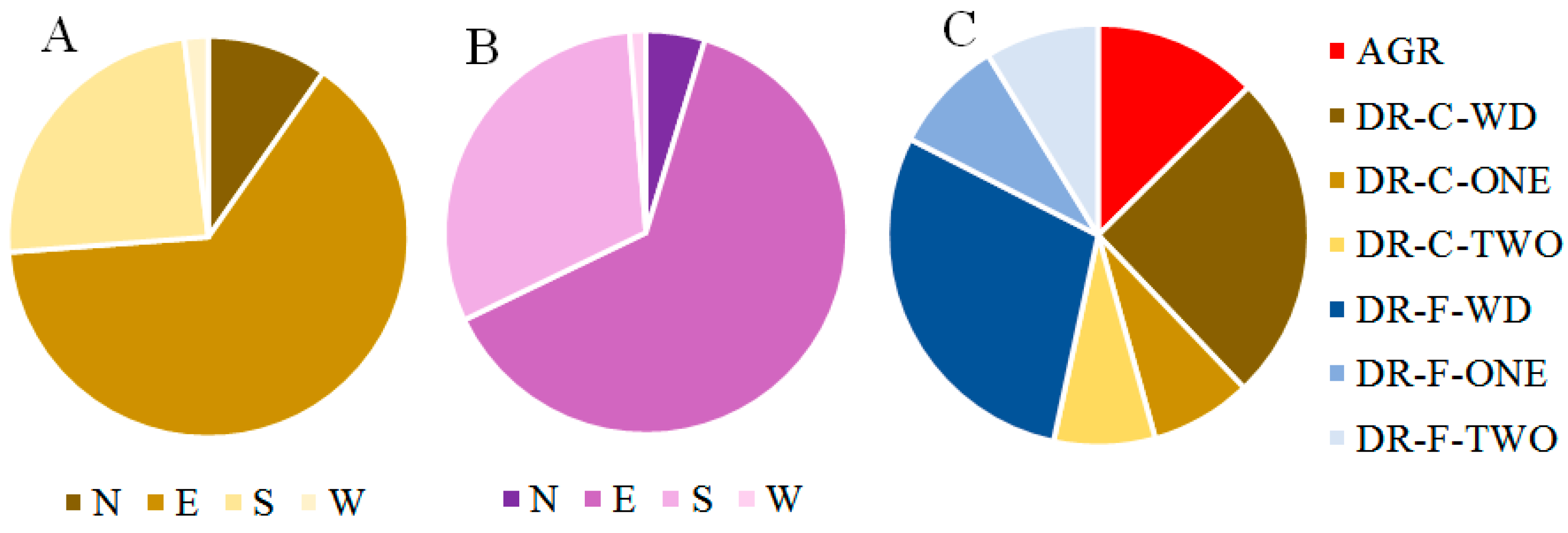
3.2. Remnant Overstory and Stand Conditions after the Windstorm Event at Rivadavia Ranch
3.3. Remnant Overstory after Harvesting Compared to Original and Impacted Forest Structure at Rivadavia Ranch
3.4. The Impact of the Windstorm and Harvesting over the Remnant Overstory at Rivadavia Ranch
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acronym | Variable | Unit | Reference |
|---|---|---|---|
| AR-CC | retention patches and clear-cuts | -- | [7] |
| AR-DR | retention patches and dispersed retention | -- | [7] |
| BA | basal area | m2 ha−1 | [40] |
| BS | bare soil | % | [41] |
| CC | canopy cover | % | [5] |
| DBH | diameter at breast height | cm | [40] |
| DEB | coarse-woody debris | % | [41] |
| DH | dominant height | m | [39] |
| DICO | dicot plants | % | [41] |
| DIF | transmitted diffuse solar radiation | % | [5] |
| DIR | transmitted direct solar radiation | % | [5] |
| DST | diameter of the stump at 30 cm height with bark | cm | [38] |
| INF | non-vascular plants | % | [41] |
| LAI | relative leaf area index | -- | [5] |
| MONO | monocot plants | % | [41] |
| REG | tree regeneration | % | [41] |
| TD | tree density | n ha−1 | [40] |
| TR | transmitted total solar radiation | % | [5] |
| TREE | overstory trees | % | [41] |
| TV | total over-bark volume | m3 ha−1 | [39,40] |
| VDEB | volume of coarse-woody debris | % | [37] |
| VRH | variable retention harvesting | -- | [7] |
| Year | F | p | Treatment | F | p |
|---|---|---|---|---|---|
| 2002 | 0.95 | 0.344 | AR-CC | 3.75 | <0.001 |
| 2003 | 0.67 | 0.422 | AR-DR | 2.37 | 0.001 |
| 2004 | 0.13 | 0.719 | |||
| 2005 | 1.31 | 0.267 | |||
| 2006 | 0.56 | 0.465 | |||
| 2007 | 0.05 | 0.820 | |||
| 2008 | 0.27 | 0.609 | |||
| 2009 | 0.09 | 0.767 | |||
| 2010 | 0.65 | 0.429 | |||
| 2011 | 1.77 | 0.199 | |||
| 2012 | 0.82 | 0.378 | |||
| 2013 | 1.44 | 0.246 | |||
| 2014 | 1.84 | 0.192 | |||
| 2015 | 3.52 | 0.077 | |||
| 2016 | 0.71 | 0.409 | |||
| 2017 | -- | -- | |||
| 2018 | 1.73 | 0.205 | |||
| 2019 | 0.02 | 0.896 | |||
| 2020 | 1.29 | 0.272 | |||
| 2021 | 2.38 | 0.140 | |||
| 2022 | 0.65 | 0.429 | |||
| 2023 | 1.13 | 0.301 |
| Treatment | Level | DBH-O | DBH-H | TD-O | TD-R | TD-D | TD-W | TD-H |
|---|---|---|---|---|---|---|---|---|
| PF | mean | 45.6 | -- | 388.0 | 89.9 | 7.4 | 2.7 | -- |
| SD | (9.5) | -- | (167.7) | (11.7) | (9.8) | (4.5) | -- | |
| A: Aspect | N | 56.8 b | 45.7 | 247 | 35.0 | 6.3 | 30.1 | 28.6 |
| E | 50.2 ab | 52.8 | 369 | 38.0 | 10.1 | 22.7 | 29.2 | |
| S | 45.7 a | 47.8 | 353 | 40.5 | 8.5 | 27.0 | 24.0 | |
| W | 52.0 ab | 51.6 | 242 | 42.2 | 2.6 | 24.0 | 31.1 | |
| F | 3.92 | 1.72 | 3.41 | 0.37 | 1.47 | 0.51 | 0.38 | |
| (p) | (0.014) | (0.188) | (0.024) | (0.777) | (0.233) | (0.675) | (0.770) | |
| B: Distance | 0–25 | 42.9 a | -- | 490 b | 80.7 b | 15.7 | 3.5 a | 0.0 a |
| 25–50 | 51.4 ab | -- | 350 ab | 76.1 b | 10.9 | 12.9 ab | 0.0 a | |
| 50–75 | 54.7 b | 52.2 | 240 a | 19.8 a | 4.4 | 35.7 bc | 40.1 b | |
| 75–100 | 53.9 ab | 48.6 | 230 a | 22.7 a | 4.2 | 28.0 bc | 45.1 b | |
| 100–125 | 54.2 ab | 49.8 | 243 a | 15.1 a | 2.3 | 37.8 c | 44.8 b | |
| 125–150 | 49.9 ab | 47.4 | 263 a | 19.0 a | 3.9 | 37.7 c | 39.3 b | |
| F | 2.52 | 0.73 | 5.12 | 23.85 | 2.59 | 6.64 | 13.20 | |
| (p) | (0.042) | (0.541) | (0.001) | (<0.001) | (0.037) | (<0.001) | (<0.001) | |
| A × B | F | 1.96 | 1.49 | 2.11 | 1.14 | 1.28 | 1.38 | 1.33 |
| (p) | (0.040) | (0.204) | (0.026) | (0.348) | (0.251) | (0.195) | (0.223) |
| Treatment | Level | BA-O | BA-R | BA-D | BA-W | BA-H | TV-O | TV-R | TV-D | TV-W | TV-H |
|---|---|---|---|---|---|---|---|---|---|---|---|
| PF | mean | 67.9 | 94.6 | 2.4 | 2.9 | -- | 743.9 | 94.7 | 2.4 | 2.9 | -- |
| SD | (22.7) | (8.4) | (4.8) | (6.4) | -- | 243.9 | (8.4) | (4.7) | (6.5) | -- | |
| A: Aspect | N | 61.6 | 37.6 | 7.6 | 34.3 | 20.5 | 759.7 | 37.6 | 7.5 | 34.4 | 20.4 |
| E | 65.1 | 47.1 | 6.3 | 21.9 | 24.7 | 786.7 | 47.3 | 6.2 | 21.9 | 24.6 | |
| S | 57.0 | 44.0 | 6.6 | 26.8 | 22.6 | 671.5 | 44.0 | 6.6 | 26.8 | 22.5 | |
| W | 57.1 | 46.3 | 2.5 | 21.4 | 29.7 | 694.4 | 46.4 | 2.5 | 21.4 | 29.7 | |
| F | 0.63 | 0.51 | 0.85 | 1.17 | 0.62 | 0.74 | 0.52 | 0.84 | 1.17 | 0.63 | |
| (p) | (0.600) | (0.674) | (0.473) | (0.331) | (0.602) | (0.531) | (0.674) | (0.477) | (0.331) | (0.597) | |
| B: Distance | 0–25 | 70.0 | 86.0 b | 9.9 | 4.1 a | 0.0 a | 843.6 | 85.9 b | 9.9 | 4.1 a | 0.0 a |
| 25–50 | 64.1 | 75.9 b | 7.9 | 16.2 ab | 0.0 a | 770.1 | 75.9 b | 7.8 | 16.2 ab | 0.0 a | |
| 50–75 | 60.3 | 26.2 a | 7.8 | 32.2 ab | 33.9 b | 726.2 | 26.3 a | 7.8 | 32.1 ab | 33.7 b | |
| 75–100 | 56.3 | 31.7 a | 2.2 | 22.7 ab | 43.3 b | 688.8 | 31.9 a | 2.2 | 22.6 ab | 43.3 b | |
| 100–125 | 50.3 | 16.3 a | 1.9 | 42.5 b | 39.2 b | 606.1 | 16.4 a | 2.0 | 42.6 b | 39.1 b | |
| 125–150 | 60.3 | 26.5 a | 4.5 | 39.1 b | 29.9 b | 733.7 | 26.6 a | 4.5 | 39.3 b | 29.6 b | |
| F | 1.25 | 15.85 | 1.27 | 4.66 | 10.10 | 1.07 | 15.66 | 1.24 | 4.63 | 10.01 | |
| (p) | (0.299) | (<0.001) | (0.294) | (0.002) | (<0.001) | (0.388) | (<0.001) | (0.305) | (0.001) | (<0.001) | |
| A × B | F | 1.64 | 1.12 | 1.66 | 1.37 | 1.15 | 1.50 | 1.12 | 1.66 | 1.37 | 1.15 |
| (p) | (0.098) | (0.368) | (0.094) | (0.199) | (0.338) | (0.142) | (0.369) | (0.094) | (0.200) | (0.342) |
References
- Mitchell, S.J. Wind as a natural disturbance agent in forests: A synthesis. Forestry 2013, 86, 147–157. [Google Scholar] [CrossRef]
- Rebertus, A.J.; Veblen, T. Structure and tree-fall gap dynamics of old-growth Nothofagus forests in Tierra del Fuego, Argentina. J. Veg. Sci. 1993, 4, 641–654. [Google Scholar] [CrossRef]
- Amoroso, M.M.; Blazina, A.P. Disturbance history and dynamics of an old-growth Nothofagus forest in Southern Patagonia. Forests 2020, 11, e101. [Google Scholar] [CrossRef]
- Mitchell, S.J.; Ruel, J.C. Modeling windthrow at stand and landscape scales. In Simulation Modeling of Forest Landscape Disturbances; Perera, A., Sturtevant, B., Buse, L., Eds.; Springer: Cham, Switzerland, 2015; pp. 17–43. [Google Scholar] [CrossRef]
- Caldentey, J.; Mayer, H.; Ibarra, M.; Promis, A. The effects of a regeneration felling on photosynthetic photon flux density and regeneration growth in a Nothofagus pumilio forest. Eur. J. Forest Res. 2009, 128, 75–84. [Google Scholar] [CrossRef]
- Zubizarreta-Gerendiain, A.; Pukkala, T.; Peltola, H. Effects of wind damage on the optimal management of boreal forests under current and changing climatic conditions. Can. J. For. Res. 2017, 472, 246–256. [Google Scholar] [CrossRef]
- Martínez Pastur, G.; Rosas, Y.M.; Toro Manríquez, M.; Huertas Herrera, A.; Miller, J.; Cellini, J.M.; Barrera, M.D.; Peri, P.L.; Lencinas, M.V. Knowledge arising from long-term research of variable retention harvesting in Tierra del Fuego: Where do we go from here? Ecol. Process. 2019, 8, e24. [Google Scholar] [CrossRef]
- Soler, R.; Schindler, S.; Lencinas, M.V.; Peri, P.L.; Martínez Pastur, G. Retention forestry in southern Patagonia: Multiple environmental impacts and their temporal trends. Int. For. Rev. 2015, 17, 231–243. [Google Scholar] [CrossRef]
- Vergara, P.M.; Schlatter, R.P. Aggregate retention in two Tierra del Fuego Nothofagus forests: Short-term effects on bird abundance. For. Ecol. Manag. 2006, 225, 213–224. [Google Scholar] [CrossRef]
- Peri, P.L.; Rosas, Y.M.; López, D.; Lencinas, M.V.; Cavallero, L.; Martínez Pastur, G. Conceptual framework to define management strategies for silvopastoral systems in native forests. Ecol. Aust. 2022, 32, 749–766. [Google Scholar] [CrossRef]
- Rebertus, A.J.; Kitzberger, T.; Veblen, T.; Roovers, L.M. Blowdown history and landscape patterns in the Andes of Tierra del Fuego, Argentina. Ecology 1997, 78, 678–692. [Google Scholar] [CrossRef]
- Salas-Eljatib, C. An approach to quantify climate-productivity relationships: An example from a widespread Nothofagus forest. Ecol. Appl. 2021, 31, e2285. [Google Scholar] [CrossRef] [PubMed]
- Quine, C.P.; Gardiner, B.A.; Moore, J. Wind disturbance in forests: The process of wind created gaps, tree overturning, and stem breakage. In Plant Disturbance Ecology: The Process and the Response; Johnson, E.A., Miyanishi, K., Eds.; Academic Press: New York, NY, USA, 2020; pp. 117–184. [Google Scholar] [CrossRef]
- Coates, K.D.; Lilles, E.B.; Dhar, A.; Hall, E.C. Wind damage over 21 years across different levels of tree removal in natural-origin mixed forests of northwestern British Columbia. Can. J. For. Res. 2020, 50, 946–952. [Google Scholar] [CrossRef]
- Pukkala, T.; Laiho, O.; Lähde, E. Continuous cover management reduces wind damage. For. Ecol. Manag. 2016, 372, 120–127. [Google Scholar] [CrossRef]
- Magagnotti, N.; Picchi, G.; Spinelli, R. A versatile machine system for salvaging small-scale forest windthrow. Biosyst. Eng. 2013, 115, 381–388. [Google Scholar] [CrossRef]
- Franklin, J.F.; Berg, D.R.; Thornburgh, D.A.; Tappeiner, J.C. Alternative silvicultural approaches to timber harvesting: Variable retention harvest systems. In Creating a Forestry for the 21st Century; Kohm, K.A., Franklin, J.F., Eds.; Island Press: Washington, DC, USA, 1997; pp. 111–140. [Google Scholar]
- Halpern, C.B.; McKenzie, D. Disturbance and post-harvest ground conditions in a structural retention experiment. For. Ecol. Manag. 2001, 154, 215–225. [Google Scholar] [CrossRef]
- Costa, S.; Ibanez, L. Can wood storage be profitable? French experience after the windstorms in 1999. J. For. Econ. 2005, 11, 161–176. [Google Scholar] [CrossRef]
- Beese, W.J.; Rollerson, T.P.; Peters, C.M. Quantifying wind damage associated with variable retention harvesting in coastal British Columbia. For. Ecol. Manag. 2019, 443, 117–131. [Google Scholar] [CrossRef]
- Man, R.; Rice, M. Trembling aspen stand response 15 years after windthrow, salvage harvesting, and forest renewal. Forests 2022, 13, e843. [Google Scholar] [CrossRef]
- Rosenfeld, J.M.; Navarro Cerrillo, R.; Guzman Alvarez, J.R. Regeneration of Nothofagus pumilio (Poepp et. Endl.) Krasser forests after five years of seed tree cutting. J. Environ. Manag. 2006, 78, 44–51. [Google Scholar] [CrossRef]
- Rodríguez Souilla, J.; Cellini, J.M.; Roig, F.A.; Lencinas, M.V.; Chaves, J.E.; Aravena Acuña, M.C.; Peri, P.L.; Martínez Pastur, G. Variable retention harvesting and climate variations influence over natural regeneration dynamics in Nothofagus pumilio forests of Southern Patagonia. For. Ecol. Manag. 2023, 544, e121221. [Google Scholar] [CrossRef]
- Mattera, M.G.; Pastorino, M.J.; Lantschner, M.V.; Marchelli, P.; Soliani, C. Genetic diversity and population structure in Nothofagus pumilio, a foundation species of Patagonian forests: Defining priority conservation areas and management. Sci. Rep. 2020, 10, e19231. [Google Scholar] [CrossRef]
- Marchelli, P.; Gallo, L.A. Annual and geographic variation in seed traits of Argentinean populations of southern beech Nothofagus nervosa (Phil.) Dim. et Mil. For. Ecol. Manag. 1999, 121, 239–250. [Google Scholar] [CrossRef]
- Ataíde, G.M.; Castro, R.V.; Correia, A.C.; dos Reis, G.G.; Reis, M.G.; Rosado, A.M. Interação árvores e ventos: Aspectos ecofisiológicos e silviculturais. Cienc. Florest. 2015, 25, 523–536. [Google Scholar] [CrossRef]
- Konôpka, B.; Zach, P.; Kulfan, J. Wind: An important ecological factor and destructive agent in forests. Forestry 2016, 62, 123–130. [Google Scholar] [CrossRef]
- Gardiner, B. Wind damage to forests and trees: A review with an emphasis on planted and managed forests. J. For. Res. 2021, 26, 248–266. [Google Scholar] [CrossRef]
- Finnigan, J.J.; Brunet, Y. Turbulent airflow in forest on flat and hilly terrain. In Wind and Trees; Coutts, M.P., Grace, J., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 3–40. [Google Scholar]
- Gardiner, B.A.; Stacey, G.R.; Belcher, R.E.; Wood, C.J. Field and wind-tunnel assessment of the implications of respacing and thinning on tree stability. Forestry 1997, 70, 233–252. [Google Scholar] [CrossRef]
- Hannah, P.; Palutikof, J.P.; Quine, C.P. Predicting wind speeds for forest areas in complex terrain. In Wind and Trees; Coutts, M.P., Grace, J., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 113–132. [Google Scholar]
- Zhu, J.; Liu, Z.; Li, X.; Matsuzaki, T.; Gonda, Y. Review: Effects of wind on trees. J. For. Res. 2004, 15, 153–160. [Google Scholar] [CrossRef]
- Gardiner, B.; Berry, P.; Moulia, B. Review: Wind impacts on plant growth, mechanics and damage. Plant Sci. 2016, 245, 94–118. [Google Scholar] [CrossRef] [PubMed]
- Romagnoli, F.; Cadei, A.; Costa, M.; Marangon, D.; Pellegrini, G.; Nardi, D.; Masiero, M.; Secco, L.; Grigolato, S.; Lingua, E.; et al. Windstorm impacts on European forest-related systems: An interdisciplinary perspective. For. Ecol. Manag. 2023, 541, e121048. [Google Scholar] [CrossRef]
- Peri, P.L.; Lencinas, M.V.; Bousson, J.; Lasagno, R.; Soler, R.; Bahamonde, H.A.; Martínez Pastur, G. Biodiversity and ecological long-term plots in southern Patagonia to support sustainable land management: The case of PEBANPA network. J. Nat. Conserv. 2016, 34, 51–64. [Google Scholar] [CrossRef]
- Martínez Pastur, G.; Lencinas, M.V.; Cellini, J.M.; Peri, P.L.; Soler, R. Timber management with variable retention in Nothofagus pumilio forests of Southern Patagonia. For. Ecol. Manag. 2009, 258, 436–443. [Google Scholar] [CrossRef]
- Martínez Pastur, G.; Rosas, Y.M.; Chaves, J.E.; Cellini, J.M.; Barrera, M.D.; Favoretti, S.; Lencinas, M.V.; Peri, P.L. Changes in forest structure values along the natural cycle and different management strategies in Nothofagus antarctica forests. For. Ecol. Manag. 2021, 486, e118973. [Google Scholar] [CrossRef]
- Fajardo, A.; Moreno-Meynard, P.; Soto, D. Forest stand dynamics of a short-stature tree species: Ecological knowledge for sustainable forest management. J. Appl. Ecol. 2024, 61, 1500–1507. [Google Scholar] [CrossRef]
- Martínez Pastur, G.; Peri, P.L.; Vukasovic, R.; Vaccaro, S.; Piriz Carrillo, V. Site index equation for Nothofagus pumilio Patagonian forest. Phyton 1997, 6, 55–60. [Google Scholar]
- Martínez Pastur, G.; Lencinas, M.V.; Cellini, J.M.; Diaz, B.; Peri, P.L.; Vukasovic, R. Herramientas disponibles para la construcción de un modelo de producción para la lenga (Nothofagus pumilio) bajo manejo en un gradiente de calidad de sitio. Bosque 2002, 23, 69–80. [Google Scholar] [CrossRef]
- Levy, E.G.; Madden, E.A. The point method of pasture analyses. N. Z. J. Agric. 1933, 46, 267–379. [Google Scholar]
- Bauhus, J.; Forrester, D.I.; Gardiner, B.; Jactel, H.; Vallejo, R.; Pretzsch, H. Ecological stability of mixed-species forests. In Mixed-Species Forests; Pretzsch, H., Forrester, D., Bauhus, J., Eds.; Springer: Berlin, Germany, 2017. [Google Scholar] [CrossRef]
- Vergani, C.; Giadrossich, F.; Buckley, P.; Conedera, M.; Pividori, M.; Salbitano, F.; Rauch, H.; Lovreglio, R.; Schwarz, M. Root reinforcement dynamics of European coppice woodlands and their effect on shallow landslides: A review. Earth-Sci. Rev. 2017, 167, 88–102. [Google Scholar] [CrossRef]
- Ferrio, J.P.; Shestakova, T.A.; del Castillo, J.; Voltas, J. Oak competition dominates interspecific interactions in growth and water-use efficiency in a mixed pine-oak Mediterranean forest. Forests 2021, 12, e1093. [Google Scholar] [CrossRef]
- Pretzsch, H. Facilitation and competition reduction in tree species mixtures in Central Europe: Consequences for growth modeling and forest management. Ecol. Model. 2022, 464, e109812. [Google Scholar] [CrossRef]
- Serrano-León, H.; Nitschke, R.; Scherer-Lorenzen, M.; Forrester, D. Intra-specific leaf trait variability of F. sylvatica, Q. petraea and P. abies in response to inter-specific competition and implications for forest functioning. Tree Physiol. 2022, 42, 253–272. [Google Scholar] [CrossRef]
- Kern, C.; Waskiewicz, J.; Frelich, L.; Muñoz Delgado, B.; Kenefic, L.; Clark, K.; Kabrick, J. Understanding compositional stability in mixedwood forests of eastern North America. Can. J. For. Res. 2021, 51, 897–909. [Google Scholar] [CrossRef]
- Maher Hasselquist, E.; Kuglerová, L.; Sjögren, J.; Hjältén, J.; Ring, E.; Sponseller, R.; Andersson, E.; Lundström, J.; Mancheva, I.; Nordin, A.; et al. Moving towards multi-layered, mixed-species forests in riparian buffers will enhance their long-term function in boreal landscapes. For. Ecol. Manag. 2021, 493, e119254. [Google Scholar] [CrossRef]
- Churchill, D.; Larson, A.; Dahlgreen, M.; Franklin, J.F.; Hessburg, P.; Lutz, J. Restoring forest resilience: From reference spatial patterns to silvicultural prescriptions and monitoring. For. Ecol. Manag. 2013, 291, 442–457. [Google Scholar] [CrossRef]
- Halpin, C.R.; Lorimer, C.G. Trajectories and resilience of stand structure in response to variable disturbance severities in northern hardwoods. For. Ecol. Manag. 2016, 365, 69–82. [Google Scholar] [CrossRef]
- Pollmann, W.; Veblen, T.T. Nothofagus regeneration dynamics in south-central chile: A test of a general model. Ecol. Mon. 2004, 74, 615–634. [Google Scholar] [CrossRef]
- Rodríguez Souilla, J.; Lencinas, M.V.; Cellini, J.M.; Chaves, J.E.; Roig, F.A.; Peri, P.L.; Martínez Pastur, G. Seed fall and leaf litter relationships in Nothofagus pumilio forests: Changes according to retention levels and years after harvesting. Trees 2023, 37, 583–597. [Google Scholar] [CrossRef]
- Curzon, M.T.; Kern, C.; Baker, S.; Palik, B.; D’Amato, A. Retention forestry influences understory diversity and functional identity. Ecol. Appl. 2020, 30, e2097. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, R.C.; Jenkins, M.; Saunders, M. Accumulation and connectivity of coarse woody debris in partial harvest and unmanaged relict forests. PLoS ONE 2014, 9, e113323. [Google Scholar] [CrossRef] [PubMed]
- Hämäläinen, A.; Hujo, M.; Heikkala, O.; Junninen, K.; Kouki, J. Retention tree characteristics have major influence on the post-harvest tree mortality and availability of coarse woody debris in clear-cut areas. For. Ecol. Manag. 2016, 369, 66–73. [Google Scholar] [CrossRef]
- Woodall, C.W.; Nagel, L.M. Downed woody fuel loading dynamics of a large-scale blowdown in northern Minnesota, USA. For. Ecol. Manag. 2007, 247, 194–199. [Google Scholar] [CrossRef]
- Yang, B.; Shaw, R.H.; Paw, U.K. Wind loading on trees across a forest edge: A large eddy simulation. Agric. For. Meteor. 2006, 141, 133–146. [Google Scholar] [CrossRef]
- Wohl, E. Logjam fluctuations during the decade after a major blowdown along a mountain stream in the US Southern Rockies. Earth Sur. Proc. Landf. 2022, 47, 699–705. [Google Scholar] [CrossRef]
- Bosley-Smith, C.; Fraver, S.; D’Amato, A.W.; Rogers, N.; Tabak, N.; Wason, J. The natural ‘exclosure effect’ and tree regeneration following post-windstorm salvage logging. J. Appl. Ecol. 2024, 61, 260–270. [Google Scholar] [CrossRef]
- Woods, K.; Kern, C. Intermediate disturbances drive long-term fluctuation in old-growth forest biomass: An 84-yr temperate forest record. Ecosphere 2022, 13, e03871. [Google Scholar] [CrossRef]
- Brando, P.; Silvério, D.; Maracahipes, L.; Benzi, R.; Paolucci, L.; Maracahipes-Santos, L.; Rattis, L.; Macedo, M.; Balch, J. Legacies of multiple disturbances on fruit and seed patterns in Amazonia: Implications for forest functional traits. Ecosphere 2024, 15, e4780. [Google Scholar] [CrossRef]
- Dahir, S.E.; Lorimer, C. Variation in canopy gap formation among developmental stages of northern hardwood stands. Can. J. For. Res. 1996, 26, 1875–1892. [Google Scholar] [CrossRef]
- Ziegler, S. Disturbance regimes of hemlock-dominated old-growth forests in northern New York, USA. Can. J. For. Res. 2002, 32, 2106–2115. [Google Scholar] [CrossRef]
- Franklin, J.F.; Donato, D. Variable retention harvesting in the Douglas-fir region. Ecol. Process. 2020, 9, e8. [Google Scholar] [CrossRef]
- Gustafsson, L.; Bauhus, J.; Asbeck, T.; Augustynczik, A.; Basile, M.; Frey, J.; Gutzat, F.; Hanewinkel, M.; Helbach, J.; Jonker, M.; et al. Retention as an integrated biodiversity conservation approach for continuous-cover forestry in Europe. Ambio 2020, 49, 85–97. [Google Scholar] [CrossRef]
- Gustafsson, L.; Hannerz, M.; Koivula, M.; Shorohova, E.; Vanha-Majamaa, I.; Weslien, J. Research on retention forestry in Northern Europe. Ecol. Process. 2020, 9, e3. [Google Scholar] [CrossRef]
- Marshall, V.G. Impacts of forest harvesting on biological processes in northern forest soils. For. Ecol. Manag. 2000, 133, 43–60. [Google Scholar] [CrossRef]
- Raj Kizha, A.; Nahor, E.; Coogen, N.; Louis, L.; George, A. Residual stand damage under different harvesting methods and mitigation strategies. Sustainability 2021, 13, e7641. [Google Scholar] [CrossRef]


| Treatment | Level | DH | CC | LAI | DIR | DIF | TR |
|---|---|---|---|---|---|---|---|
| PF | Mean | 21.4 | 90.4 | 2.65 | 14.3 | 12.2 | 12.5 |
| SD | (1.0) | (4.8) | (0.53) | (6.4) | (5.9) | (5.8) | |
| A: Aspect | N | 23.7 | 55.6 a | 0.79 a | 64.6 c | 56.2 c | 57.4 c |
| E | 23.1 | 63.1 b | 1.08 b | 49.6 b | 46.7 ab | 47.1 ab | |
| S | 23.1 | 64.6 b | 1.13 b | 35.7 a | 442 a | 42.9 a | |
| W | 22.7 | 57.1 a | 0.82 a | 49.9 b | 52.7 bc | 52.3 bc | |
| F | 1.21 | 11.54 | 9.33 | 11.12 | 9.16 | 10.38 | |
| (p) | (0.318) | (<0.001) | (<0.001) | (<0.001) | (<0.001) | (<0.001) | |
| B: Distance | 0–25 | 23.1 | 81.6 d | 1.98 d | 22.1 a | 22.1 a | 22.1 a |
| 25–50 | 23.1 | 71.6 c | 1.34 c | 33.2 ab | 36.0 b | 35.6 b | |
| 50–75 | 23.1 | 61.7 b | 0.89 b | 46.8 bc | 48.7 c | 48.4 c | |
| 75–100 | 23.1 | 51.7 a | 0.60 ab | 62.9 cd | 60.3 d | 60.7 d | |
| 100–125 | 23.1 | 46.9 a | 0.46 a | 69.2 d | 66.2 d | 66.6 d | |
| 125–150 | 23.1 | 47.1 a | 0.46 a | 65.7 d | 66.3 d | 66.2 d | |
| F | 0.01 | 79.86 | 75.06 | 19.69 | 65.72 | 57.27 | |
| (p) | (0.999) | (<0.001) | (<0.001) | (<0.001) | (<0.001) | (<0.001) | |
| A × B | F | 0.01 | 0.67 | 0.72 | 0.77 | 0.60 | 0.65 |
| (p) | (0.999) | (0.803) | (0.748) | (0.702) | (0.863) | (0.820) |
| Treatment | Level | BS | TREE | DICO | REG | MONO | INF | DEB | VDEB |
|---|---|---|---|---|---|---|---|---|---|
| PF | Mean | 56.8 | 2.0 | 7.6 | 5.2 | 1.6 | 3.6 | 16.4 | 365.1 |
| SD | (12.9) | (2.1) | (7.4) | (10.0) | (2.1) | (3.0) | (9.5) | (242.4) | |
| A: Aspect | N | 42.8 | 1.8 | 11.5 | 5.1 | 2.9 | 2.9 | 27.3 | 585.9 |
| E | 45.3 | 1.3 | 14.2 | 2.4 | 2.8 | 0.8 | 29.6 | 561.1 | |
| S | 49.7 | 1.3 | 10.0 | 3.5 | 3.8 | 1.1 | 27.3 | 567.4 | |
| W | 38.7 | 2.2 | 18.2 | 3.8 | 4.2 | 1.7 | 29.6 | 644.0 | |
| F | 2.11 | 0.49 | 1.90 | 0.43 | 1.03 | 1.72 | 0.18 | 0.21 | |
| (p) | (0.111) | (0.692) | (0.142) | (0.733) | (0.388) | (0.175) | (0.908) | (0.885) | |
| B: Distance | 0–25 | 59.0 b | 3.0 | 7.3 | 3.3 | 3.3 | 2.7 | 15.6 a | 357.1 |
| 25–50 | 42.3 a | 1.3 | 12.0 | 3.3 | 3.3 | 1.0 | 26.3 ab | 604.2 | |
| 50–75 | 40.3 a | 1.7 | 15.3 | 5.3 | 4.0 | 1.7 | 30.0 ab | 631.5 | |
| 75–100 | 44.0 ab | 2.0 | 15.0 | 3.0 | 2.3 | 0.6 | 31.7 b | 689.6 | |
| 100–125 | 39.0 a | 0.3 | 13.0 | 3.3 | 3.3 | 1.3 | 34.0 b | 591.4 | |
| 125–150 | 40.3 a | 1.7 | 18.3 | 4.0 | 2.0 | 2.7 | 33.0 b | 663.9 | |
| F | 3.64 | 1.36 | 1.36 | 0.17 | 0.23 | 1.01 | 3.41 | 1.43 | |
| (p) | (0.007) | (0.256) | (0.256) | (0.971) | (0.946) | (0.422) | (0.010) | (0.229) | |
| A × B | F | 1.98 | 1.29 | 0.58 | 0.88 | 0.83 | 0.60 | 1.52 | 1.26 |
| (p) | (0.037) | (0.246) | (0.873) | (0.594) | (0.642) | (0.857) | (0.134) | (0.263) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez Pastur, G.; Rodríguez-Souilla, J.; Bottan, L.; Favoretti, S.; Cellini, J.M. Quantifying Blowdown Disturbance in Overstory Retention Patches in Managed Nothofagus pumilio Forests with Variable Retention Harvesting. Forests 2024, 15, 1432. https://doi.org/10.3390/f15081432
Martínez Pastur G, Rodríguez-Souilla J, Bottan L, Favoretti S, Cellini JM. Quantifying Blowdown Disturbance in Overstory Retention Patches in Managed Nothofagus pumilio Forests with Variable Retention Harvesting. Forests. 2024; 15(8):1432. https://doi.org/10.3390/f15081432
Chicago/Turabian StyleMartínez Pastur, Guillermo, Julián Rodríguez-Souilla, Lucía Bottan, Santiago Favoretti, and Juan M. Cellini. 2024. "Quantifying Blowdown Disturbance in Overstory Retention Patches in Managed Nothofagus pumilio Forests with Variable Retention Harvesting" Forests 15, no. 8: 1432. https://doi.org/10.3390/f15081432