
Impact of Increment Coring on Growth and Mortality across Various Size Classes of Khasi Pine (Pinus kesiya) in Northern Thailand


Abstract
:1. Introduction
2. Materials and Methods
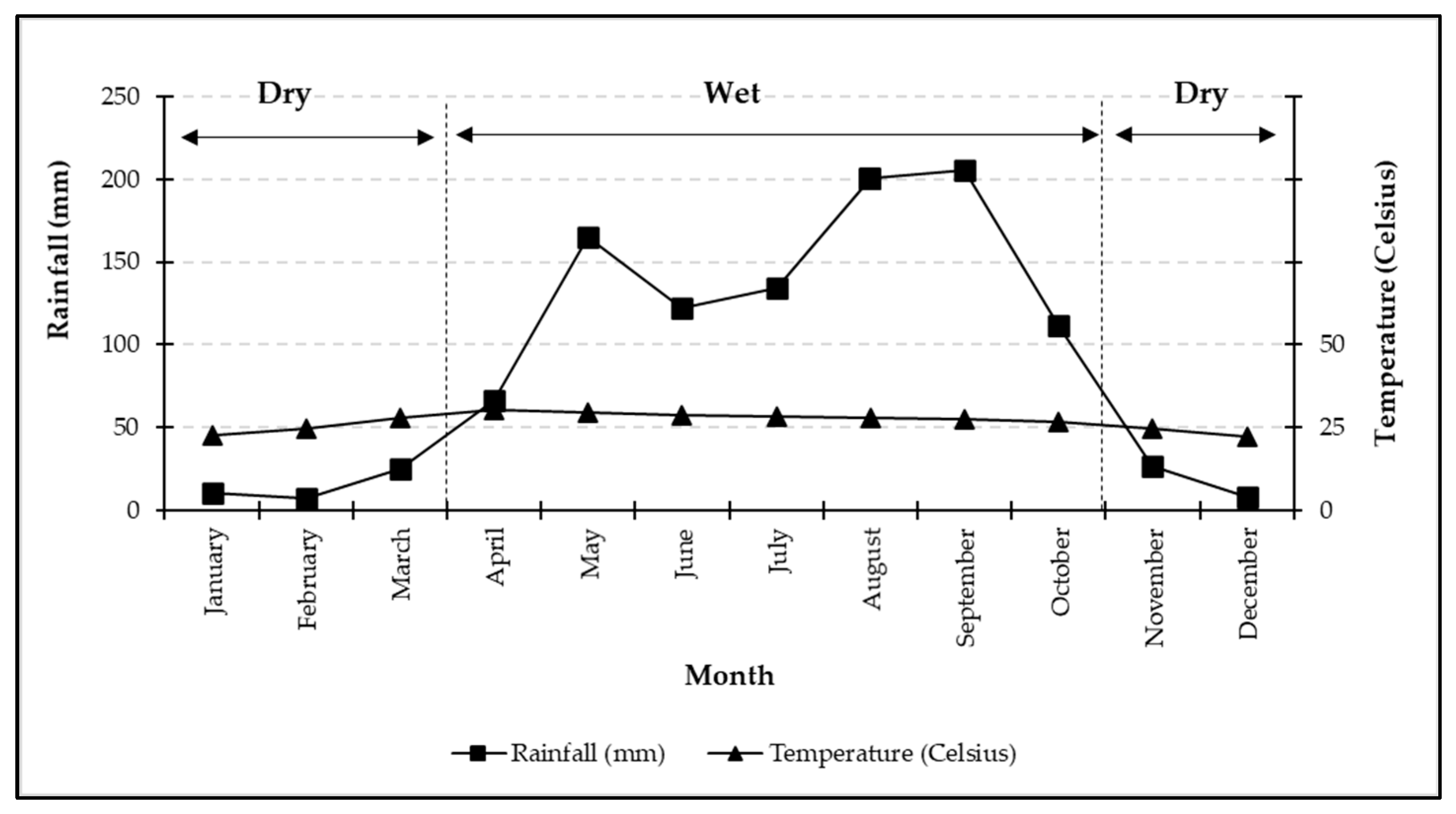
2.1. Study Site
2.2. Sample Selection and Periodic Growth Measurement
2.3. Periodic Growth Comparison
2.4. Analysis of Increment Coring Effects on Tree Growths
3. Results
3.1. Periodic Growth Data
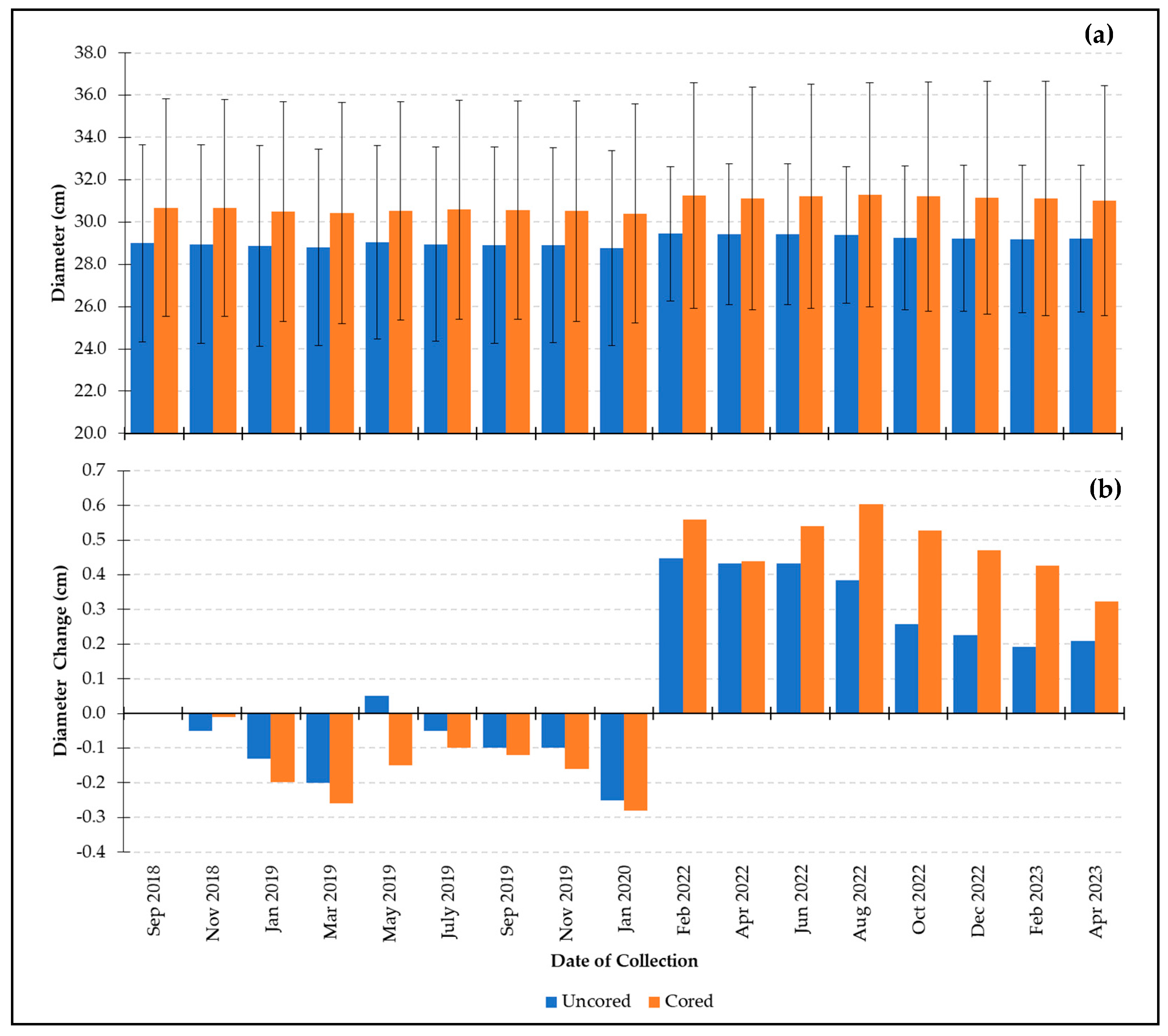
3.1.1. Small-Size Class
3.1.2. Medium-Size Class
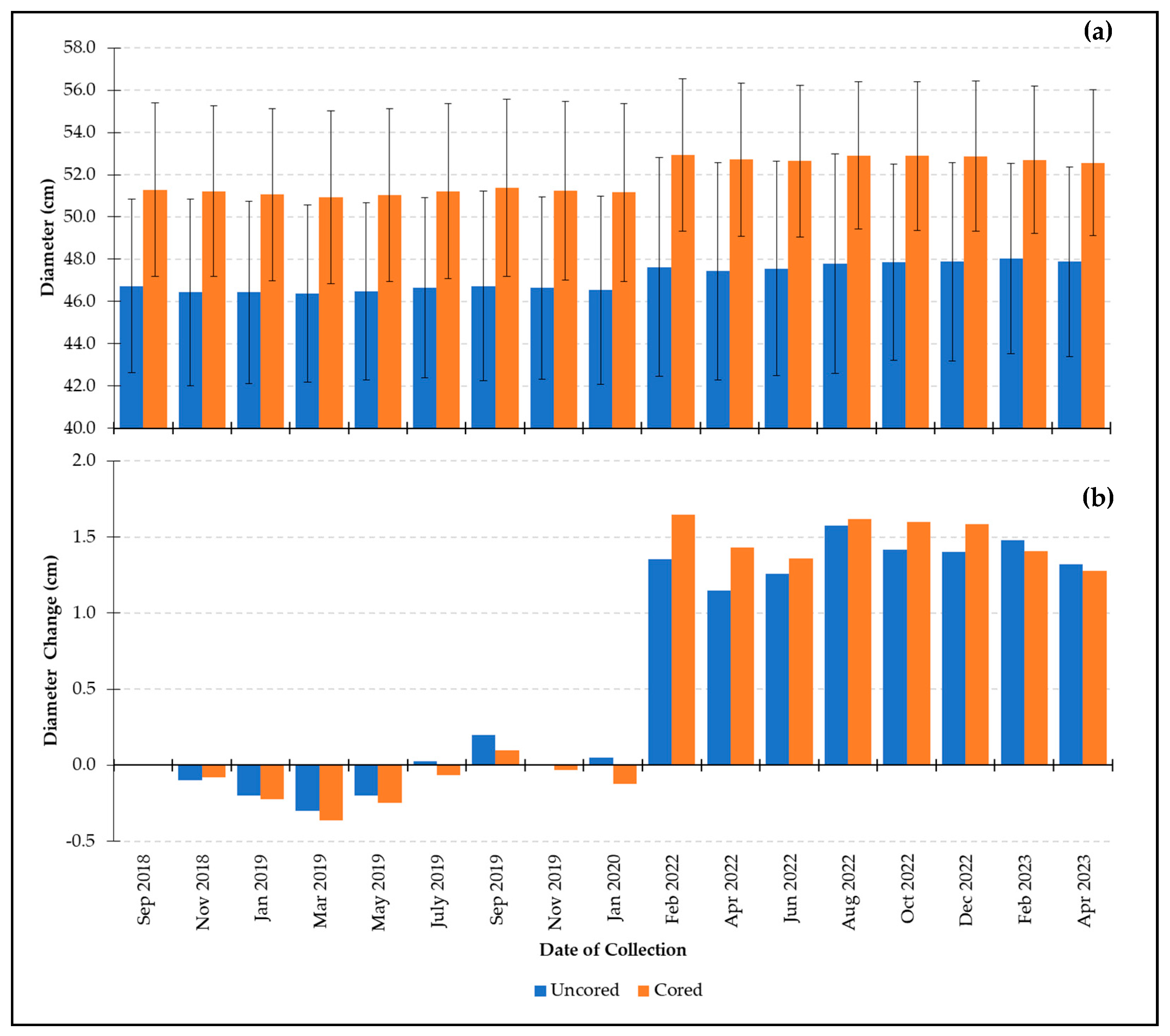
3.1.3. Large-Size Class
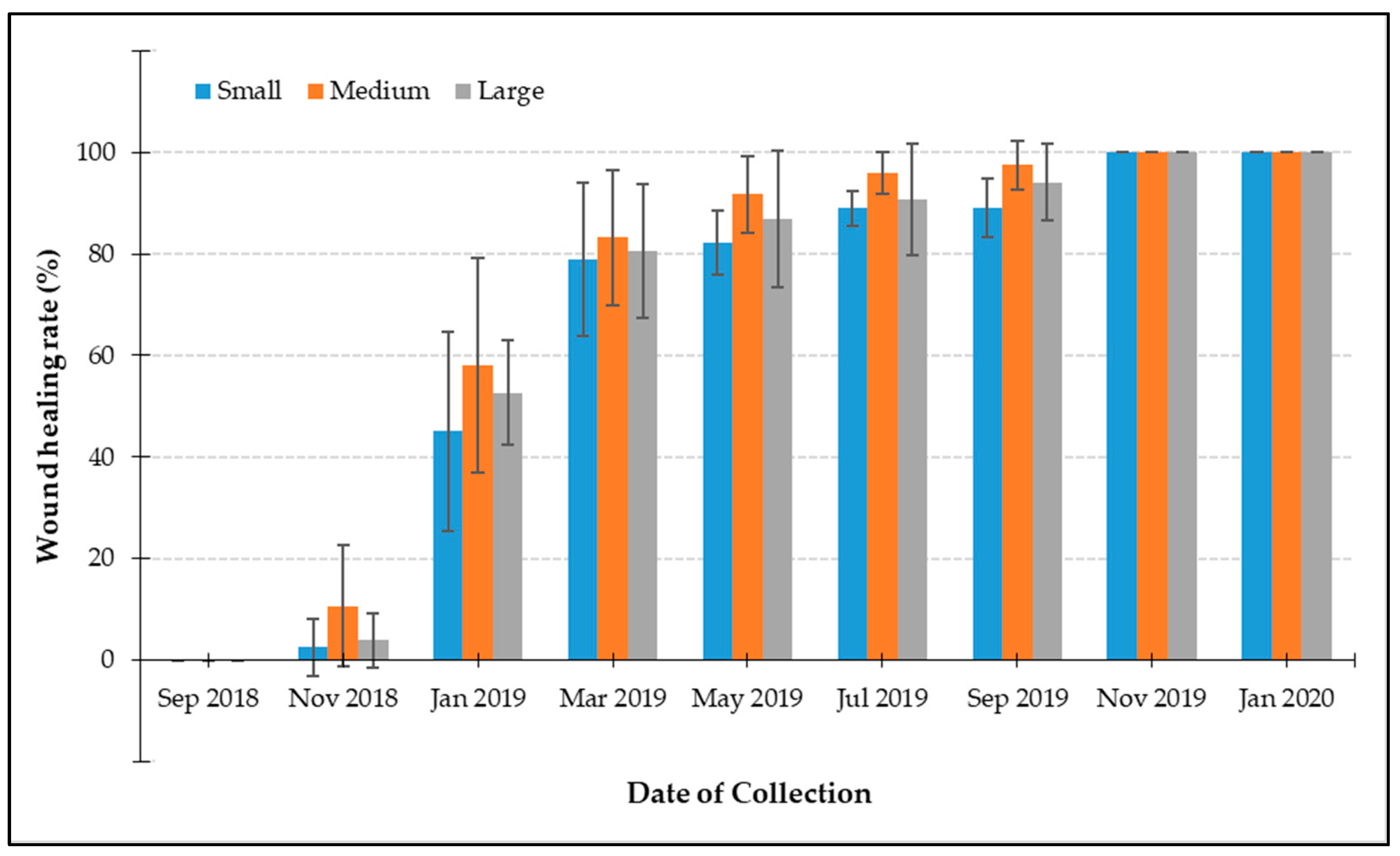
3.2. Wound Healing Rates from Increment Coring
3.3. Comparison of Periodic Growth and Wound Healing Rate
3.3.1. Periodic Growth
- Growth comparison of cored and uncored trees in the small-size class:
- Growth comparison of cored and uncored trees in the medium-size class:
- Growth comparison of cored and uncored trees in the large-size class:
- Growth comparison of cored and uncored trees across all size classes:
3.3.2. Wound Healing Rates
3.4. Data Interpretation for Increment Coring Effects on Tree Growth
4. Discussion
4.1. Periodic Growth Patterns and Seasonal Influence
4.2. Wound Healing Rate Following Increment Coring
4.3. Increment Coring Effects on Tree Growths
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ITTO. Criteria and Indicators for the Sustainable Management of Tropical Forests; ITTO Policy Development Series No. 21; International Tropical Timber Organization: Yokohama, Japan, 2016; Available online: https://www.itto.int/direct/topics/topics_pdf_download/topics_id=4872&no=1&disp=inline (accessed on 7 July 2024).
- MCPFE. General Declaration and Resolutions Adopted. In Proceedings of the Second Ministerial Conference on the Protection of Forest in Europe, Helsinki, Finland, 16–17 June 1993. [Google Scholar]
- Rice, R.E.; Sugal, C.A.; Ratay, S.M.; Fonseca, G.A. Sustainable Forest Management: A Review of Conventional Wisdom; Advances in Applied Biodiversity Science No. 3; CABS/Conservation International: Washington, DC, USA, 2001; pp. 1–29. [Google Scholar]
- Dobbertin, M.; Neumann, M.; Schroeck, H.W. Tree growth measurements in long-term forest monitoring in Europe. In Developments in Environmental Science; Ferretti, M., Fischer, R., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2013; Volume 12, pp. 183–204. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK, 1976; 567p. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Arizona Press: Tucson, AZ, USA, 1996; 73p. [Google Scholar]
- Worbes, M.; Schöngart, J. Measures for sustainable forest management in the tropics—A tree-ring based case study on tree growth and forest dynamics in a Central Amazonian lowland moist forest. PLoS ONE 2019, 14, e0219770. [Google Scholar] [CrossRef]
- Spiecker, H. Tree rings and forest management in Europe. Dendrochronologia 2002, 20, 191–202. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Zuidema, P.A. The use of tree rings in tropical forest management: Projecting timber yields of four Bolivian tree species. For. Ecol. Manag. 2006, 226, 256–267. [Google Scholar] [CrossRef]
- Sano, M.; Buckley, B.M.; Sweda, T. Tree-ring based hydroclimate reconstruction over northern Vietnam from Fokienia hodginsi: Eighteenth century mega-drought and tropical Pacific influence. Clim. Dyn. 2009, 33, 331–340. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Wang, H.; Guo, J.; Zhang, E.; Wang, J.; Li, X.; Wei, H.; Zhou, C. Tree-ring-based drought reconstruction in northern north China over the past century. Atmosphere 2022, 13, 482. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, H.; Chen, F.; Wang, S.; Hu, M.; Hadad, M.; Roig, F. Drought reconstruction since 1796 CE based on tree-ring widths in the upper Heilongjiang (Amur) River basin in Northeast Asia and its linkage to Pacific Ocean climate variability. Clim. Past. 2023, 19, 2079–2092. [Google Scholar] [CrossRef]
- Asad, F.; Adil, M.; Zhu, H.; Büntgen, U.; Hamayun, M.; Fahad, A.; Ali, S. A tree ring-based spring temperature reconstruction for the Hindu Kush region in northern Pakistan. Trees For. People 2024, 16, 100541. [Google Scholar] [CrossRef]
- Lumyai, P.; Duangsathaporn, K. Climate reconstruction on the growth of teak in Umphang Wildlife Sanctuary, Thailand. Environ. Nat. Resour. J. 2018, 16, 21–30. [Google Scholar]
- Temchai, T.; Saengswang, M.; Jaikaew, P.; Deekaew, P.; Thammalangka, P.; Hengswang, D.; Wanmanee, S.; Jitra, N.; Thongsuk, P.; Thongkerd, T. The relative between phenology and climates factor of plants in Pine-Dry dipterocarp forest, Suphanburi province, Thailand. J. Thail. Natl. Parks Res. 2018, 2, 122–136. [Google Scholar]
- Wunder, J.; Reineking, B.; Hillgarter, F.W.; Bigler, C.; Bugmann, H. Long-term effects of increment coring on Norway spruce mortality. Can. J. For. Res. 2011, 41, 2326–2336. [Google Scholar] [CrossRef]
- Wunder, J.; Manusch, C.; Queloz, V.; Brang, P.; Ringwald, V.; Bugmann, H. Does increment coring enhance tree decay? New insights from tomography assessments. Can. J. For. Res. 2013, 43, 711–718. [Google Scholar] [CrossRef]
- Portier, J.; Shackleton, R.T.; Klesse, S.; Ferretti, M.; Flury, R.; Hobi, M.L.; Stillhard, J.; von Arx, G.; Rohner, B.; Thürig, E. No evidence that coring affects tree growth or mortality in three common European temperate forest tree species. Eur. J. For. Res. 2024, 143, 129–139. [Google Scholar] [CrossRef]
- Neo, L.; Chong, K.Y.; Koh, C.Y.; Tan, S.Y.; Loh, J.W.; Lim, R.C.J.; Seah, W.W.; Tan, H.T.W. Short-term external effects of increment coring on some tropical trees. J. Trop. For. Sci. 2017, 29, 519–529. [Google Scholar]
- Thai Meteorological Department. Available online: https://www.tmd.go.th/service/tmdData (accessed on 10 April 2024).
- Maxwell, J.F.; Elliott, S.; Palee, P.; Anusarnsunthorn, V. The vegetation of Doi Khuntan National Park, Lamphun-Lampang provinces, Thailand. Nat. Hist. Bull. Siam Soc. 1995, 43, 185–205. [Google Scholar]
- Field, A.P. Discovering Statistics Using SPSS, 3rd ed.; SAGE Publications: Dubai, United Arab Emirates, 2009; pp. 457–505. [Google Scholar]
- Bathke, A.C.; Schabenberger, O.; Tobias, R.D.; Madden, L.V. Greenhouse-Geisser adjustment and the ANOVA-type statistic: Cousins or twins? Am. Stat. 2009, 63, 239–246. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Intern. 2004, 11, 36–42. [Google Scholar]
- Pumijumnong, N.; Wanyaphet, T. Seasonal cambial activity and tree-ring formation of Pinus merkusii and Pinus kesiya in Northern Thailand in dependence on climate. For. Ecol. Manag. 2006, 226, 279–289. [Google Scholar] [CrossRef]
- Inthawong, S.; Pumijumnong, N.; Muangsong, C.; Buajan, S.; Cai, B.; Chatwatthana, R.; Chareonwong, U.; Phewphan, U. Growth response of Thai pine (Pinus latteri) to climate drivers in Tak province of northwestern Thailand. Forests 2024, 15, 345. [Google Scholar] [CrossRef]
- D’Arrigo, R.; Barbetti, M.; Watanasak, M.; Buckley, B.; Krusic, P.; Boonchirdchoo, S.; Sarutanon, S. Progress in dendroclimatic studies of Mountain pine in Northern Thailand. IAWA J. 1997, 18, 433–444. [Google Scholar] [CrossRef]
- Buckley, B.; Barbetti, M.; Watanasak, M.; D’Arrigo, R.; Boonchirdchoo, S.; Suratanoon, S. Dendrochronological investigation in Thailand. IAWA J. 1995, 16, 393–409. [Google Scholar] [CrossRef]
- Pumijumnong, N.; Eckstein, D. Reconstruction of pre-monsoon weather conditions in northwestern Thailand from the tree-ring widths of Pinus merkusii and Pinus kesiya. Trees 2011, 25, 125–132. [Google Scholar] [CrossRef]
- Kumar, K.; Shah, S.K.; Roy, A.; Kant, S. Dendroclimatology of teak indicates prevailing climatic conditions of tropical moist forests in India. Ecol. Indic. 2021, 129, 107888. [Google Scholar] [CrossRef]
- Ng’andwe, P.; Chungu, D.; Tailoka, F.; Bwembya, M. Assessing cross-datable distinct annual growth rings in non-native Pinus kesiya Royle ex Gordon in Zambia. Dendrochronologia 2021, 67, 125835. [Google Scholar] [CrossRef]
- Singh, N.D.; Venugopal, N. Cambial activity and annual rhythm of xylem production of Pinus kesiya Royle ex. Gordon (Pinaceae) in relation to phenology and climatic factors growing in sub-tropical wet forest of North East India. Flora Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 198–204. [Google Scholar]
- Chaudhary, V.; Bhattacharyya, A. Suitability of Pinus kesiya in Shillong, Meghalaya, for tree-ring analyses. Curr. Sci. 2002, 83, 1010–1015. [Google Scholar]
- Anderson-Teixeira, K.J.; Herrmann, V.; Rollinson, C.R.; Gonzalez, B.; Gonzalez-Akre, E.B.; Pederson, N.; Alexander, M.R.; Allen, C.D.; Alfaro-Sánchez, R.; Awada, T.; et al. Joint effects of climate, tree size, and year on annual tree growth derived from tree-ring records of ten globally distributed forests. Glob. Chang. Biol. 2022, 28, 245–266. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Antia, J.P.; González de Andrés, E.; Gazol, A.; Camarero, J.J.; Linares, J.C. Tree-level climate sensitivity reveals size effects and impending growth decline in silver fir affected by dieback. Forests 2024, 15, 999. [Google Scholar] [CrossRef]
- Eckstein, D.; Dujesiefken, D. Long-term effects in trees due to increment borings. Dendrochronologia 1998, 16–17, 205–211. [Google Scholar]
- Shigo, A.L. Compartmentalization: A conceptual framework for understanding how trees grow and defend themselves. Ann. Rev. Phytopathol. 1984, 22, 189–214. [Google Scholar] [CrossRef]
- Blanchette, R.A.; Bigges, A.R. Defense Mechanisms of Woody Plants Against Fungi, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1992; 478p. [Google Scholar]
- Shigo, A.L. Compartmentalization of decay in trees. Sci. Am. 1985, 152, 96–103. [Google Scholar] [CrossRef]
- Dujesiefken, D.; Rhaesa, A.; Eckstein, D.; Stobbe, H. Tree wound reactions of differently treated boreholes. J. Arboric. 1999, 25, 113–123. [Google Scholar] [CrossRef]
- Grissino-Mayer, H.D. A manual and tutorial for the proper use of an increment borer. Tree-Ring Res. 2003, 59, 63–79. [Google Scholar]
- van Mantgem, P.J.; Stephenson, N.L. Does coring contribute to tree mortality? Can. J. For. Res. 2004, 34, 2394–2398. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Size Class | Period | Greenhouse–Geisser Correction | Time Effect | Time × Group Interaction |
|---|---|---|---|---|
| Small | September 2018– January 2020 | Not Violated | F(7, 56) = 7.248, p = 0.00 (Significant) | F(7, 56) = 1.129, p = 0.359 (Insignificant) |
| February 2022– April 2023 | Not Violated | F(7, 35) = 2.249, p = 0.053 (Insignificant) | F(7, 35) = 0.581, p = 0.766 (Insignificant) | |
| Medium | September 2018– January 2020 | Violated, Correction Applied | F(3.011, 36.129) = 9.121, p = 0.00 (Significant) | F(3.011, 36.129) = 0.743, p = 0.534 (Insignificant) |
| February 2022– April 2023 | Violated, Correction Applied | F(1.474, 14.736) = 1.420, p = 0.266 (Insignificant) | F(1.474, 14.736) = 1.381, p = 0.274 (Insignificant) | |
| Large | September 2018– January 2020 | Violated, Correction Applied | F(3.938, 55.129) = 13.557, p = 0.00 (Significant) | F(3.938, 55.129) = 0.444, p = 0.774 (Insignificant) |
| February 2022– April 2023 | Violated, Correction Applied | F(1.582, 18.981) = 5.952, p = 0.014 (Significant) | F(1.582, 18.981) = 1.811, p = 0.194 (Insignificant) | |
| All Sizes | September 2018– January 2020 | Violated, Correction Applied | F(3.577, 135.916) = 19.920, p = 0.00 (Significant) | F(3.577, 135.916) = 0.830, p = 0.497 (Insignificant) |
| February 2022– April 2023 | Violated, Correction Applied | F(1.673, 51.862) = 5.044, p = 0.014 (Significant) | F(1.673, 51.862) = 0.366, p = 0.657 (Insignificant) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palakit, K.; Pumijumnong, N. Impact of Increment Coring on Growth and Mortality across Various Size Classes of Khasi Pine (Pinus kesiya) in Northern Thailand. Forests 2024, 15, 1444. https://doi.org/10.3390/f15081444
Palakit K, Pumijumnong N. Impact of Increment Coring on Growth and Mortality across Various Size Classes of Khasi Pine (Pinus kesiya) in Northern Thailand. Forests. 2024; 15(8):1444. https://doi.org/10.3390/f15081444
Chicago/Turabian StylePalakit, Kritsadapan, and Nathsuda Pumijumnong. 2024. "Impact of Increment Coring on Growth and Mortality across Various Size Classes of Khasi Pine (Pinus kesiya) in Northern Thailand" Forests 15, no. 8: 1444. https://doi.org/10.3390/f15081444