Abstract
Moso bamboo (Phyllostachys edulis (Carrière) J. Houzeau), which is native to China, is considered to be an invasive species due to its powerful asexual reproductive capabilities that allow it to rapidly spread into neighboring ecosystems and replace existing plant communities. In the absence of human intervention, it remains poorly understood how indigenous moso bamboo forests naturally expand into surrounding areas over the long term, and whether these patterns vary with environmental changes. Using multi-year forest resource inventory data, we extracted moso bamboo patches that emerged from 2010 to 2020 and proposed a bamboo expansion index to calculate the average rate of patch expansion during this period. Using the first global 30 m land-cover dynamic monitoring product with a fine classification system, we assessed the expansion speeds of moso bamboo into various areas, particularly forests with different canopy closures and categories. Using parameter-optimized geographic detectors, we explored the significance of multi-factors in the expansion process. The results indicate that the average expansion rate of moso bamboo forests in China is 1.36 m/y, with evergreen broadleaved forests being the primary area for invasion. Moso bamboo expands faster into open forest types (0.15 < canopy closure < 0.4), shrublands, and grasslands. The importance of factors influencing the expansion rate is ranked as follows: temperature > chemical properties of soil > light > physical properties of soil > moisture > atmosphere > terrain. When considering interactions, the primary factors contributing to expansion rates include various climate factors and the combined effect of climate factors and soil factors. Our work underscores the importance of improving the quality and density of native vegetation, such as evergreen broadleaved forests. Effective management strategies, including systematic monitoring of environmental variables, as well as targeted interventions like bamboo removal and soil moisture control, are essential for mitigating the invasion of moso bamboo.
1. Introduction
Biological invasions, particularly those involving plants, pose ongoing and future threats to the functionality of terrestrial ecosystems. This type of invasion is primarily driven by non-native species [1,2] that gradually become dominant due to the absence of competitors. Consequences of these invasions vary widely, impacting water resources and nutrient cycling, and causing changes in the ecosystem structure and functionality [3,4]. However, native species can also invade, as an abundance within their original ranges can similarly alter the structure and dynamics of native communities [5,6]. In many regions of the world, the invasive behavior of native bamboo hinders the natural succession of secondary forests, leading to losses in biodiversity and production opportunities [7,8,9]. While the impacts are similar, the effects of native species expansion are far less recognized than those of non-native plants.
The invasion of a species is intricately connected to the vulnerability of local ecosystems [10]. Extensive deforestation and afforestation efforts since the 1950s have augmented the vulnerability of China’s native forest ecosystems [11]. The decline in forest quantity and quality has resulted in the degradation of forest ecosystems, habitat fragmentation, and a loss of biodiversity [12]. As a native species in the subtropics, moso bamboo (Phyllostachys edulis (Carrière) J. Houzeau) has increasingly been recognized as an indigenous invasive species in China over the past few decades, gradually occupying larger land areas [13]. With its rapid growth and strong cloning abilities, moso bamboo can outcompete most tree species and extensively spread into adjacent forests, resulting in the dominance of this single species within forest ecosystems [14]. The assessment of the ecological effects caused by moso bamboo expansion is currently a focal point of concern. The invasion of bamboo can significantly alter forest structures [9,15,16,17,18], reduce species diversity [7,19,20], and change soil organic carbon levels and microbial communities [21,22,23,24]. These impacts adversely affect natural succession and ecosystem functions in both native and novel habitats. If the expansion of moso bamboo continues, it could pose a significant threat to the ecological stability of surrounding areas. Therefore, it is imperative to assess the extent of moso bamboo expansion to fully understand the threats it poses and to implement effective control measures.
Previous studies have utilized the expansion distance and area of individual bamboo plants to quantify the extent of expansion [25]. Landscape indices can quantify areas and fragmentation of bamboo patches [26], yet they fail to depict its expansion level. Additionally, while many studies focus on specific reserves, numerous bamboo species have been introduced from Asia into non-protected areas across Europe, America, and Latin America [27], intensifying global concerns about bamboo invasions [28]. Parameters for evaluating the extent of large-scale moso bamboo expansion at the watershed, regional, and global scales are currently lacking.
Investigations into bamboo distribution, expansion, or invasion suggest a substantial increase in the area covered by bamboo forests due to climate change [29,30,31]. Moso bamboo exhibits a pronounced trend of spreading toward higher latitudes under various climate change conditions [26,32]. The characteristics of forests at the invasion interfaces play a crucial role in the bamboo’s expansion into adjacent communities [33]. Moso bamboo tends to proliferate in structurally simple forests [25], and its expansion may also be influenced by the slope of neighboring forests [34]. Wind and snowfall can accelerate the cloning rates of bamboo, and some studies suggest that soil properties also influence its expansion [35,36]. It can be surmised that bamboo expansion rates may be associated with various factors; however, conclusive evidence regarding the contributions and mechanisms of these factors to moso bamboo expansion is still lacking. Investigating the impacts of multiple factors on the expansion of bamboo forests, particularly across various habitat types, holds significant scientific significance for ecological security monitoring of moso bamboo expansion.
This study was conducted in mainland China, where moso bamboo has emerged as a significant concern for native plant species and biodiversity conservation. Several key issues need to be addressed: (1) obtaining long-standing moso bamboo patches and calculating their expansion rates, (2) examining variations in bamboo expansion into different habitat types, especially in various forests and under different degrees of canopy closure, and (3) identifying the factors contributing to the differential expansion speed. To address these issues, we aim to define an expansion index suitable for national and even broader-scale measurements to assess the average patch expansion rate of moso bamboo. By using this index, we will be able to assess differences in bamboo invasion across various habitats, identify the key factors influencing these expansion differences, and explore the combined effects of climate, soil, and topography on bamboo forest spread. Our research lays the groundwork for a theoretical and practical exploration into moso bamboo invasion across various habitat types, particularly in different forest environments. This is crucial for a better understanding of the mechanisms driving the widespread natural growth of moso bamboo into surrounding areas.
2. Materials and Methods
2.1. Study Area
China is located in the eastern part of Asia, with high topography in the west and low topography in the east, in a stepped pattern, and a complex and diverse climate, including tropical, subtropical, warm temperate, middle temperate, and cold temperate climates zones from south to north. China represents a vital reservoir of global bamboo diversity, boasting an unmatched richness of species, including 857 species across 39 genera, comprising 51% of the world’s total bamboo classification [37]. Notably, moso bamboo stands out as the predominant species, making up over 70% of China’s bamboo forests, with approximately 85% of global moso bamboo distribution within China’s borders [26]. Furthermore, the historical lineage of Chinese moso bamboo stretches back over 1600 years to the Western Jin Dynasty, highlighting its pivotal role as a native Chinese plant. China’s diverse climate and intricate mountainous topography predominantly harbor moso bamboo forests, which primarily thrive in the subtropical regions of central and southern China.
2.2. Additional Patches of Moso Bamboo
The moso bamboo forest stand data utilized in our study were obtained from the National Forestry and Grassland Science Data Center (NFGSDC, https://www.forestdata.cn/sso/login.html, accessed on 1 August 2022) in China. These data were collected by the National Forestry and Grassland Administration of China using the Technical Specifications for Forest Management Planning and Design Inventory (GBT 26424-2010) [38]. This document describes the standards, codes, methods, and definitions for field inventory that are consistent and uniform across China. The inventory aimed to comprehensively survey the distribution, quantity, quality, and ownership of forests in China, presenting an accurate and objective overview of the national forest resources. Plots served as the basic unit of investigation, defined as areas with similar internal forestry characteristics that differ from neighboring regions, labeled as “dominant tree species” codes. The boundaries and attributes within each plot were gathered through field surveys. The outcomes of this inventory formed the forest resources inventory database in NFGSDC, including vector boundaries and specific stand characteristics. Each database record corresponds to the content of each plot. The original coordinate system of the vector data is the China Geodetic Coordinate System 2000. Utilizing ArcGIS Pro 3.01, the geographic coordinate system was transformed to WGS_1984. To facilitate subsequent distance calculations, a projection transformation was applied to the data, adopting the Krasovsky_1940_Albers projection coordinate system.
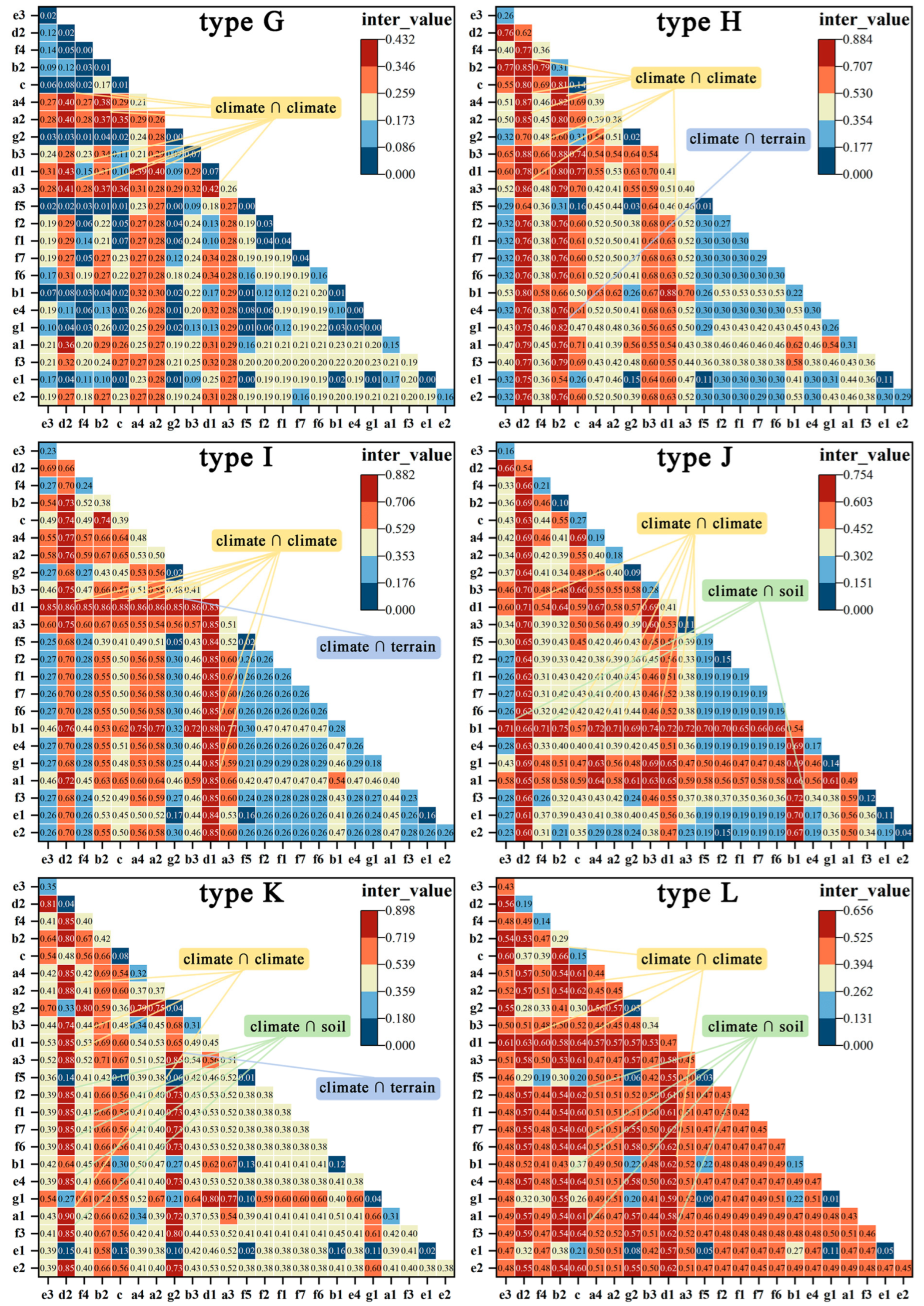
The data encompass 2010, 2015, and 2020, with no data from Hong Kong, Macao, and Taiwan. Using the code “66000” (the survey code for Phyllostachys edulis (Carrière) J. Houzeau) for dominant tree species identification, vector patches of moso bamboo were extracted for each year. We conducted a three-year data intersection analysis, excluding moso bamboo forests that remained as separate patches in any individual year. Utilizing the symmetrical difference tool by ArcGIS Pro 3.01, we extracted the added moso bamboo patches during the periods of 2010–2015 and 2015–2020, respectively. The growth span of a moso bamboo rhizome is approximately 4–5 m annually [39]. To mitigate potential human-induced planting, patches with a significant growth distance in each stage were eliminated, resulting in 2,756,739 newly added stable moso bamboo forest patches (Figure 1).
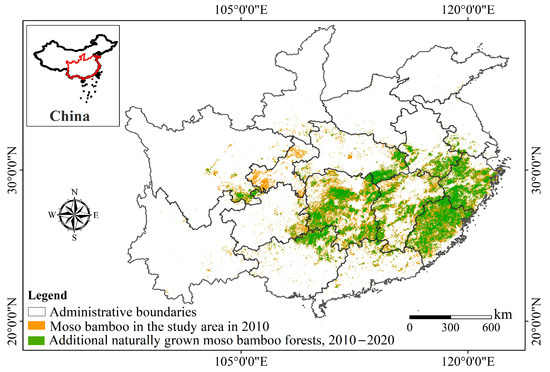
Figure 1.
Distribution of moso bamboo in the study area in 2010 and additional naturally grown moso bamboo forests from 2010 to 2020.
2.3. Expansion Index and Mechanism
Native moso bamboo expansion or invasion has been confirmed in most protected areas or multiple sample plots in experimental studies. To measure the expansion of moso bamboo forest patches across a larger area, we sought to define an expansion index to evaluate the average expansion level of these patches. In a planar view, each newly formed moso bamboo patch can be considered an irregular figure. The average height of such an irregular figure is calculated by dividing its total area by its base [40]. For moso bamboo patches, this base is defined as the contact boundary between the years before and after the bamboo expansion, and the average height reflects the extent of the new patch’s expansion. Additionally, since an added patch may expand from various directions and positions, the base of the expanding patch should consider the total length of all contact edges in the calculation process. Accordingly, we developed a formula for deriving the Moso Bamboo Expansion Index (MBEI), which represents the annual average expansion rate of each bamboo patch from the xth to the yth year.
where, MBEI is the expansion index of the newly added moso bamboo patch i, measured in meters per year.
x and y are the years before and after the expansion of the patch, respectively.
ΔAi is the area of the added patch i, measured in square meters.
Lij is the length of the adjacent edge jth of the added patch i (in year y) bordering on all moso bamboo patches in the previous stage (year x), measured in meters. n is the number of adjacent edges of patch i.
The mechanism and procedure of the MBEI are presented in Figure 2.
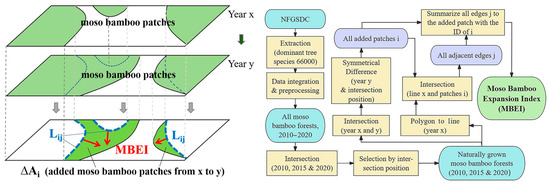
Figure 2.
Mechanism and procedure of Moso Bamboo Expansion Index.
2.4. Database on Meteorological, Soil, and Terrain Factors
We selected climate, soil, and terrain factors for analysis. Meteorological data included ten indicators from four aspects: temperature (a), moisture (b), light (c), and atmosphere (d) (Table 1). These indicators were sourced from the Geographical Data Sharing Infrastructure-Global Resources Data Cloud, the National Science & Technology Infrastructure of China, and the National Tibetan Plateau Data Center.

Table 1.
Data sources and codes for meteorological factors.
Soil conditions are represented by eleven indicators from e1 to f7, divided into physical and chemical properties (Table 2). Data were obtained from the FAO 2023, Harmonized World Soil Database version 2.0, and SIS China: Soil Information System of China. The data for elevation (g1) and slope (g2) were obtained from Alos palsar. The original coordinate system of the environmental variables is the WGS 1984 geodetic coordinate system. All variables underwent a projection transformation to the Krasovsky 1940 Albers coordinate system.
Table 2.
Data sources and codes for soil and terrain factors.
2.5. Habitat Classification
Global surface cover products serve as the primary source of fine-grained forest-type data. Presently, mainly surface cover products encompass MOD12Q1 (2001–2019), Globe Land Cover Map (2005–2006/2009), CCI_LC (1992–2020), Globeland30 (2000, 2010, 2020), GLC_FCS30 (1985–2020), ESA10 (2020), FROMGLC10 (2017), and Esri10 (2020). Among them, Globeland30 and GLC_FCS30 are closest to the Ninth National Forest Resources Inventory in China [41]. GLC_FCS30, released by the Aerospace Information Research Institute of the Chinese Academy of Sciences, is the global 30 m fine-grained land-cover classification product from 1985 to 2020. It is worth noting that, unlike the other products that primarily differentiate forest categories (deciduous or evergreen, coniferous or broadleaf), the first global 30 m land-cover dynamic monitoring product with a fine classification system (GLC_FCS30) distinguishes various types and canopy closure conditions of forests. In GLC_FCS30, the canopy closure of a forest (fc) reflects its stand density and is categorized into closed forests (fc > 0.4) and open forests (0.15 < fc < 0.4). By GLC-FCS30 classification, we categorized all ground types where added patches were grown. Based on these, moso bamboo expanded into 12 different habitat categories, as shown in Table 3.
Table 3.
Habitat system classification of natural expansion areas of moso bamboo within the study area based on GLC_FCS30.
2.6. Geo-Detector Based on Optimization Parameters
We used the latest phase (2015–2020) for factor detection. We employed a Geo-Detector to detect the spatial heterogeneity of moso bamboo expansion rates and to explore the impact of environmental factors on the differences in bamboo expansion rates [42,43,44,45]. The independent variables and dependent variables in the factor detection process need to correspond one-to-one. Firstly, we converted the MBEI of each patch into vector point features, and then assigned the values of the environmental factor raster to the vector points one-to-one. Further, in the detection process, environmental variables need to be transformed into categorical variables, and different discretization methods and spatial statistical units will affect the calculation results. We used a parameter-based optimized geographic detector model [46]. By applying classification methods such as equal intervals, natural breakpoints, quantiles, geometric intervals, and standard deviations, with the number of breaks set to 5–10 categories, we identified all combinations of parameter classifications with the maximum q-values as the optimal classification results. The degree of influence in factor detection is measured by q-values (Equation (2)), and the presence of interaction between two factors, as well as the strength, direction, linearity, or non-linearity of the interaction, are determined by the inter_value (Table 4). The calculation process adopts a t-test with a significance level of 0.05.
where, q measures the explanatory extent of a single factor; h represents the stratification (Strata) of the dependent or independent variable; and Nh and σh denote the number of units and variance for strata h, respectively. The q value ∈ [0, 1], where higher values indicate stronger explanatory power of the independent variable on the dependent variable.
Table 4.
Types of interaction for geodetectors.
3. Results
3.1. Moso Bamboo Expansion Index in Habitat
The total area of natural expansion of moso bamboo in China from 2010 to 2020 was 1578.8 km2. Closed evergreen broadleaved forests (C), closed evergreen needle-leaved forests (E), closed deciduous broadleaved forests (G), and agricultural land (A) were the primary invaded types, accounting for 1019.57, 230.30, 158.6, and 157.87 km2, respectively. Their invasion proportions reached 99.22% of the total expansion area, with closed evergreen broadleaved forests (C) being the predominant invaded type, covering 68.3%. Meanwhile, despite their comparatively smaller areas, in open forests, invasion areas exhibited a sequence of evergreen broadleaved forest (B) > evergreen needle-leaved forest (D) > deciduous broadleaved forest (F) (Table 5).
Table 5.
The number, area, proportion and MBEI value of panels of moso bamboo expanding into different habitats from 2010 to 2020.
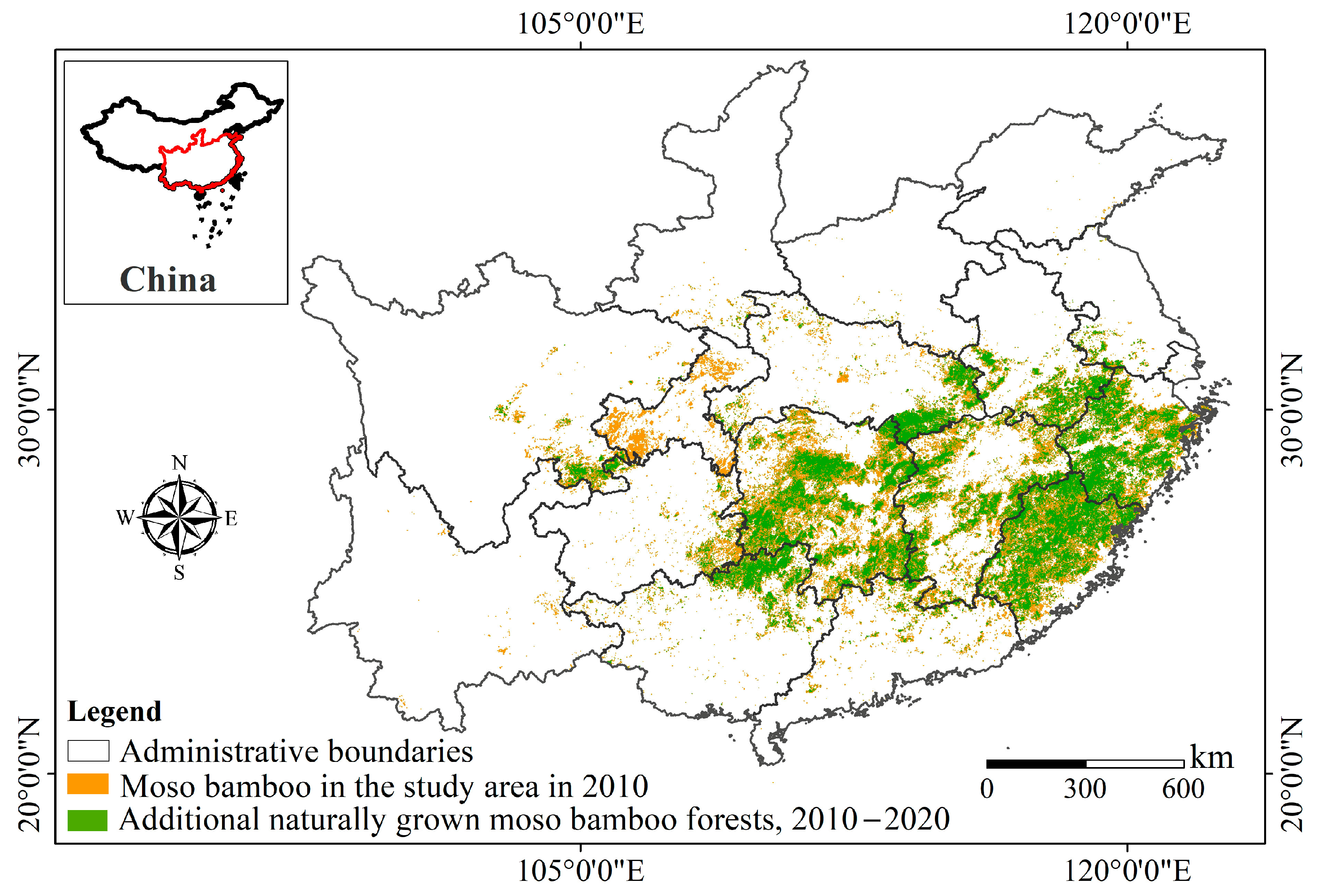
From 2010 to 2020, the average natural expansion rate of moso bamboo was 1.36 m/year, with rates of 1.25 m/year and 1.47 m/year for the earlier and later periods, respectively (Table 5). When considering different habitat types, the MBEI values for closed evergreen broadleaved forests (C) and agricultural land (A) consistently ranked the lowest. The invasion speed into shrublands (H, I, evergreen or not) remained the highest, with average speeds exceeding 2 m/year, at 2.01 m/year and 2.04 m/year, respectively. When considering different canopy closure conditions, the MBEI in closed forests was lower than in open forests. In terms of the mean and median MBEI values for the two stages, evergreen broadleaved forests C (fc > 0.4) were lower than evergreen broadleaved forests B (0.15 < fc < 0.4), evergreen needle-leaved forests E (fc > 0.4) were lower than D (0.15 < fc < 0.4), and deciduous broadleaved forests G (fc > 0.4) were lower than F (0.15 < fc < 0.4) (Table 5 and Figure 3).
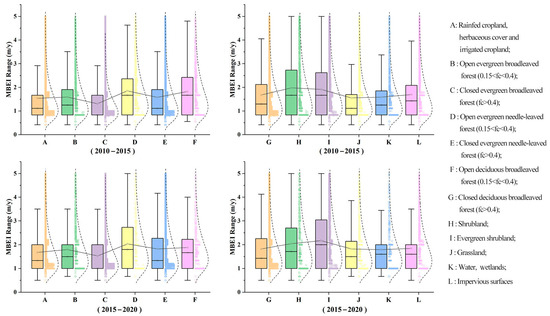
Figure 3.
Sample normal distribution curves and box plots of MBEI for mosaic bamboo expansion into different habitats from 2010 to 2020.
3.2. Factor Detector for MBEI
Most climate, terrain, and soil factors greatly affect the expansion rate of moso bamboo. Overall, the q-values of climate indicators surpass those of soil factors, which in turn exceed those of terrain indicators. The contribution of factors to the MBEI is as follows: temperature (a) > chemical properties of soil (f) > light (d) > physical properties of soil (e) > moisture (b) > atmosphere (c) > terrain (g) (Figure 4, right). Among them, the contribution rates of three temperature indicators, including ≥10 °C accumulated temperature (a1), annual mean temperature (a2), and annual mean minimum temperature (a3), are all higher than 0.5 (Figure 4, left).
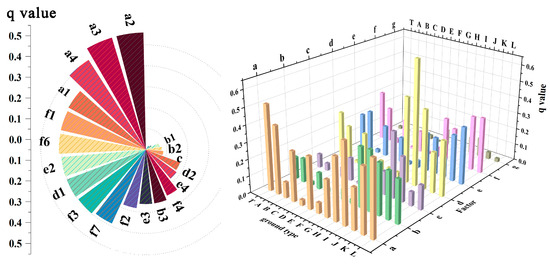
Figure 4.
On the left, the circular bar chart illustrates the contribution values (q values) of individual factors for the bamboo area. The calculation process adopts a t-test with a significance level of 0.05, The p-values for all factors are < 0.05. Indicators with very low contribution rates are not labeled in the figure, as they are sorted from highest to lowest contribution rate.(a1: ≥10 °C accumulated temperature; a2: annual mean temperature; a3: annual mean minimum temperature; a4: annual mean maximum temperature; b1: annual precipitation; b2: relative humidity; b3: potential mean evapotranspiration; c: near-surface wind speed; d1: surface solar radiation; d2: sunshine duration; e1: dominant soil unit; e2: texture class; e3: reference bulk density; e4: AWC for rootable soil depth; f1: organic carbon content; f2: total nitrogen content; f3: total phosphorus content; f4: total potassium content; f5: pH in water; f6: case saturation as the percentage of CECsoil; f7: cation exchange capacity of the soil; g1: elevation; and g2: slope). On the right, the 3D histogram shows the average contribution rate q values of the seven major factors for the expansion of bamboo into different habitats. (T: all expansion areas of moso bamboo; A: rainfed cropland, herbaceous cover, and irrigated cropland; B: open evergreen broadleaved forest (0.15 < fc < 0.4); C: closed evergreen broadleaved forest (fc > 0.4); D: open evergreen needle-leaved forest (0.15 < fc < 0.4); E: closed evergreen needle-leaved forest (fc > 0.4); F: open deciduous broadleaved forest (0.15 < fc < 0.4); G: closed deciduous broadleaved forest (fc > 0.4); H: shrubland; I: evergreen shrubland; J: grassland; K: water, wetlands; L: impervious surfaces. a: temperature; b: moisture; c: atmosphere; d: light; e: physical properties of soil; f: chemical properties of soil; and g: terrain).
When comparing different specific categories, temperature (a) remained the most significant factor affecting the expansion rate of moso bamboo into various types, including types A, B, C, E, G, and K, which are primarily agricultural land, arboreal forests, and wetlands (Figure 4, right). Light (d) emerged as the most influential factor affecting moso bamboo expansion in shrublands and grasslands (H, I, and J) (Figure 4, right, and Figure 5). The highest explanatory factors for categories D and F were atmosphere(c) and terrain(g). Additionally, when considering different degrees of canopy closure, environmental factors generally played a more significant role in closed forests than in various types of open forests (B, D, and F) (Figure 5).
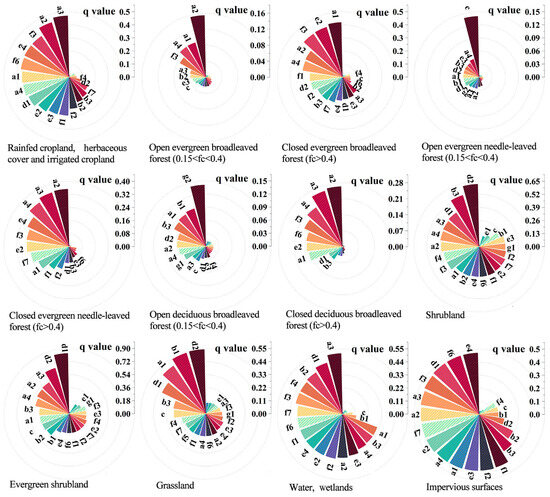
Figure 5.
The circular bar chart shows the q values of the driving factors for the expansion of bamboo into 12 different habitats. The calculation process adopts a t-test with a significance level of 0.05. Except for f5 in type open evergreen broadleaved forests (0.15 < fc < 0.4) and e2 in type open evergreen needle-leaved forests (0.15 < fc < 0.4), the p-values of others are <0.05. Indicators with very low contribution rates are not labeled in the figure, as they are sorted from highest to lowest contribution rate. (a1: ≥10 °C accumulated temperature; a2: annual mean temperature; a3: annual mean minimum temperature; a4: annual mean maximum temperature; b1: annual precipitation; b2: relative humidity; b3: potential mean evapotranspiration; c: near-surface wind speed; d1: surface solar radiation; d2: sunshine duration; e1: dominant soil unit; e2: texture class; e3: reference bulk density; e4: AWC for rootable soil depth; f1: organic carbon content; f2: total nitrogen content; f3: total phosphorus content; f4: total potassium content; f5: pH in water; f6: base saturation as the percentage of CECsoil; f7: cation exchange capacity of the soil; g1: elevation; and g2: slope).
3.3. Interaction Detector for MBEI
Despite varying degrees, interactions between all factors were more influential than any individual factor. The types of interactions among factors include Enhance_bi or Enhance_nonlinear. Considering the combined influence of these factors on the disparity in bamboo expansion rates, we have summarized the primary interacting factors and categories across different types of land.
Overall, in the entire newly added areas, there are 18 sets of interactions with importance higher than 0.617 (Figure 6). Among them, 17 sets involve the mutual interactions among climate factors, labeled as climate ∩ climate. The combination of surface solar radiation (d1) ∩ sunshine duration (d2) (0.771), and annual precipitation (b1) ∩ surface solar radiation (d1) (0.765) stands out as the two most important interaction pairs.
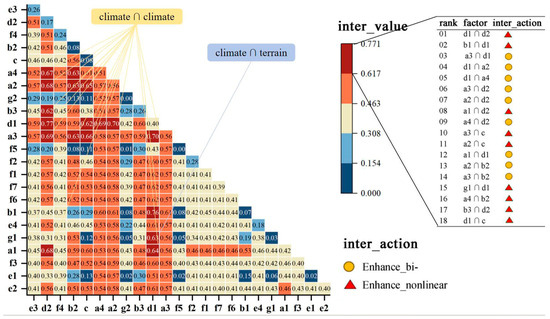
Figure 6.
A magnitude and heat map of interaction values between 23 drivers across the expansion region of moso bamboo. (a1: ≥10 °C accumulated temperature; a2: annual mean temperature; a3: annual mean minimum temperature; a4: annual mean maximum temperature; b1: annual precipitation; b2: relative humidity; b3: potential mean evapotranspiration; c: near-surface wind speed; d1: surface solar radiation; d2: sunshine duration; e1: dominant soil unit; e2: texture class; e3: reference bulk density; e4: AWC for rootable soil depth; f1: organic carbon content; f2: total nitrogen content; f3: total phosphorus content; f4: total potassium content; f5: pH in water; f6: base saturation as the percentage of CECsoil; f7: cation exchange capacity of the soil; g1: elevation; and g2: slope).
Additionally, factors with a higher inter_value in interaction detectors are also of a higher q_value in single-factor testing. However, some factors with weaker power in single-factor testing may gain significant importance when interacting with others. In single-factor testing, the importance of terrain to MBEI is lower than other variables, yet its interaction with surface solar radiation (d1) also reaches 0.631 (Figure 6).
There are slight differences in interactions among different types. Among the 130 dominant interactions across 12 types and the T category, interactions between climate factors account for the most significant contribution, totaling 90 groups (69%). The interaction between d1 ∩ d2 is a crucial interacting factor for nine land types, providing the highest inter_value for types A, C, and G. The interaction between b1 ∩ d1 is significant for seven types, providing the highest value for types E and I. Interactions between climate and soil are a secondary category, totaling 29 groups, representing 22%. Among various types, the two interactions with the highest explanatory power are the total nitrogen content (f2) ∩ d2 and the total phosphorus content (f3) ∩ d2. Furthermore, there are nine groups where climate interacts with terrain factors. However, interactions between soil and terrain, as well as elevation and slope, are represented by only one group (Appendix A Table A1, Figure A1 and Figure A2).
4. Discussion
4.1. Different Rates of Expansion into Various Habitats
From 2010 to 2020, the total area of natural expansion of moso bamboo in China amounted to 1578.81 km2, with an average rate of 1.36 m/y. In surveys of unmanaged forests in Japan, the expansion length was 2.1 m/y [47]. Moso bamboo is an introduced species invading broadleaf forests in Japan [34,48]. However, in China, moso bamboo is native, and the surrounding forest and ecosystem structure remains relatively stable. Moreover, rapid growth induced by human activities was excluded. Nevertheless, moso bamboo persists in its invasion into other native regions.
When canopy closure is disregarded, the expansion rate of moso bamboo in evergreen broadleaved forests consistently exceeds that in evergreen needle-leaved forests and deciduous broadleaved forests. As the primary forest type in subtropical regions, evergreen broadleaved forests exhibit a geographical distribution similar to moso bamboo, making them more susceptible to invasion compared to other forest types [49]. The expansion rates in our study exceed the 1.04 m and 1.28 m observed in natural Cunninghamia lanceolata (Lamb.) Hook. forests and evergreen broadleaved forests in Tianmushan [12]. The potential reason for this difference may lie in the scale, as previous studies are based on individual protected areas. However, large-scale forest surveys cover all areas, thus offering more representative results. It also implies that outside the protected areas, native moso bamboo is encroaching upon adjacent forests at faster rates, without human intervention. Therefore, it is necessary to mitigate the native invasion of moso bamboo by adopting appropriate management strategies, such as selective cutting.
The expansion rate of open forests (0.15 < fc < 0.4), grasslands, and shrublands is higher than that of closed forests (fc > 0.4) and agricultural land. Moso bamboo is light-demanding and shade-tolerant, but dense canopy cover reduces bamboo photosynthesis [50,51]. Similar to many other bamboo species, moso bamboo tends to grow new culms in locations such as forest gaps and areas with sparse tree canopies, utilizing available sunlight to sustain its growth. Practices aimed at increasing bamboo yield by tree-felling [33] also indirectly contribute to the negative impact of high canopy density on bamboo expansion. Species diversity forms the basis for community structure and function, with rich biodiversity enhancing a community’s resistance to invasion and mitigating the effects of external environmental changes [52]. Forests with high canopy closure often establish dominant forest communities [53], maintaining higher stability and stronger resistance to invasion [33], resulting in slower rates of moso bamboo invasion. Therefore, enhancing the quality of native forests surrounding moso bamboo stands is crucial for mitigating invasion.
It is noteworthy that, unlike measurements based on individual plants, the expansion index in this study is calculated from the perspective of newly added patches. The expansion process of bamboo from the bamboo community into the surrounding broadleaf forest in the Qiyunshan National Nature Reserve in Jiangxi Province also follows a patch-based expansion process [54]. Bamboo expansion into surrounding areas manifests as patch expansion both at the macro remote sensing scale and at the ground community scale. Since patches are fundamental units in landscape ecology, this index is also valuable for studying the invasion of other plant species.
4.2. Significance of Climate, Soil, and Topography on Expansion Rate
The expansion of moso bamboo is affected by various factors [12]. We found that, when explaining expansion rates, temperature is the most crucial climatic factor, surpassing light, moisture, and wind speed. Since moso bamboo growth is primarily limited by thermal conditions, the most significant factors are precipitation and temperature [55]. The average temperature in the newly added bamboo areas ranges from 6.21 to 25.51 °C, with precipitation ranging from 671.6 to 2488.4 mm, which is lower than those recorded in earlier studies, showing average temperatures of 15 to 25 °C and annual precipitations of 1200 to 2500 mm required for bamboo growth [32]. This discrepancy may reflect an enhanced adaptability of moso bamboo to temperature and moisture during the expansion process.
We found that light intensity stands as the primary factor for invasion into shrubland and grassland. Most bamboo species favor sunlight because light intensity significantly affects bamboo tiller numbers, height, basal diameter, and metabolism [56]. As light intensity decreases, the branching number and height of bamboo decrease to varying degrees [57]. During expansion into grasslands and shrublands, the area and intensity of radiation are higher than in forests with tall and dense canopies [58,59,60]. However, beyond a saturation point, excessive light exposure may induce photoinhibition, thereby restraining expansion [61]. Additionally, wind speed also exerts a significant influence. Wind speed stands out as the most critical factor affecting open evergreen broadleaved forests (with maximum wind speeds of 2.1 m/s). Ventilation maintains stable oxygen and carbon dioxide levels [62]. Bamboo exhibits remarkable wind resilience. Wind disturbances may expedite bamboo cloning rates [63].
When considering the soil factors, chemical properties exceed physical properties. There is an emphasis on how invasion alters soil properties and microbial environments [64,65], yet the role of soil properties in moso bamboo growth receives little attention. We found that soil organic carbon content is the most significant soil factor, followed by base saturation as a percentage of CEC soil. Soil organic carbon is critical for maintaining soil quality [66], and base saturation influences the soil’s nutrient and water adsorption capacity [67], affecting moso bamboo growth and development. The number of new shoots in bamboo forests correlates positively with soil fertility [35]. Total nitrogen and organic matter decrease with soil depth [68], becoming limiting factors to growth to some extent. The influence of soil texture is reflected in the varying nutrient content [69] among sandy, loamy, silty, and clay soils [70], potentially affecting the growth rate of bamboo. Soil porosity exhibits a quadratic relationship with bamboo diameter and height [71]. Pore size affects soil moisture conditions, potentially influencing the growth rate of bamboo. Additionally, Dong emphasized that water saturation may be detrimental to the growth of bamboo rhizomes [35].
According to the results, terrain exhibits the highest significance for open deciduous broadleaved forests. The average slope of the expansion area is 5.4 degrees, with an average elevation of 545.9 m. The terrain is also one of the limiting factors for the growth of moso bamboo forests in Japan [72,73,74,75]. Suzuki and Nakagoshi emphasized that in Japan, mixed forests of bamboo and Pinus are limited in distribution in areas with slopes less than 4 degrees [34].
4.3. Enhancement Interaction of Environmental Factors for Expansion Rate
When considering interactions, the combined effects of climatic factors are paramount. Our study demonstrates that the interaction between sunlight ∩ precipitation and sunlight ∩ temperature is crucial to invasion into multiple area types. Takano et al. [75] confirmed the influence of minimum temperature on the distribution of two bamboo species in central and northern Japan, identifying the annual minimum temperature as a limiting factor, while rainfall showed no significant impact. It might be associated with Japan’s maritime climate, surrounded by the sea on all sides. However, China’s distribution spans from the subtropical to the temperate zones, from the southeastern maritime regions to the northwestern deserts. This extensive geographical span results in a decrease in rainfall from the southeast to the northwest [55,76]. Linger [77] also emphasized that precipitation is the most significant factor driving plant distribution, yet tree growth is attributed to reduced cloud cover and increased solar radiation [78,79]. In regions with frequent rainfall, cloud cover increases and radiation decreases, ultimately impacting photosynthesis. Additionally, rising temperatures lead to increased evaporation, which increases atmospheric water vapor and reduces radiation levels [80].
Climate factors can influence soil conditions [81], with the most significant impact from sunlight. Bamboo possesses a high adaptability to low-light conditions [82]. Under weak light, bamboo alters its biomass allocation strategy, reducing the root-to-shoot ratio, increasing nutrient uptake by the roots, and exhibiting growth tendencies compared to other species. The rapid growth of moso bamboo roots also determines its competitive advantage in resource and water absorption [83]. Additionally, rainfall is closely related to the early growth rate of vegetation [78]. Rainfall directly compensates for soil moisture during the period of bamboo shoot growth. However, it can also reduce soil nutrients, limiting bamboo’s nutrient uptake. In some regions, higher moisture levels may potentially decrease soil oxygen content [77]. Temperature can affect bamboo root absorption of mineral elements [84]. Furthermore, altitude and slope, by influencing conditions of sunlight and temperature, restrict the vertical geographical distribution of bamboo [34,85], thereby affecting its growth.
The complexity of environmental factors, especially climate, needs more long-term monitoring, ideally with the establishment of relevant databases. Additionally, suitable management practices, such as selective harvesting, establishing buffer zones, regulating moisture during the growing season, and restricting root spread, can effectively mitigate bamboo expansion and maintain ecosystem balance.
4.4. Limitation and Future Direction
Our innovative utilization of the fluctuation in indices to distinguish degrees of expansion will contribute to a more comprehensive exploration of bamboo invasion research. However, the index based on the patches may differ from measurements based on individual bamboo plants in plot studies. Nonetheless, this lays a new theoretical and methodological foundation for studying species in adjacent ecosystems. Despite the research extending beyond protected areas, many bamboo species were introduced from Asia to Europe, the United States, and Canada. Future expansion studies should encompass broader regions and larger spatial scales, potentially integrating high-resolution remote sensing imagery if feasible.
We emphasize the correlation between expansion rates and driving factors. However, most studies only focus on determining factors for moso bamboo distribution, overlooking the multiple aspects of expansion rates. The focus of our research is on understanding the relationships between bamboo forest expansion and various factors like climate and soil at a larger scale. Soil data are less precise compared to vector data and may not adequately capture small-scale environmental changes, a known challenge in large-scale research. We must acknowledge that a deeper understanding of the physiological mechanisms driving expansion requires more field studies, potentially involving the consideration of other factors such as microbial and vegetation interactions. Due to unpredictability, we excluded the significant impacts of potential anthropogenic disturbances. Furthermore, under the influence of climate change, there is a trend toward high latitude expansion of bamboo forests. Hence, it is crucial to consider evaluating edge effects, monitoring anthropogenic disturbances, and predicting future developments in future research.
5. Conclusions
We examined the natural expansion of Chinese moso bamboo forests over a decade, finding an average rate of 1.36 m per year. Compared to various ground types, closed evergreen broadleaf forests (fc > 0.4) exhibited the largest expansion area at 1.42 m per year, surpassing expansion rates measured in other protected areas. The MBEI is calculated based on newly established patches rather than individual plants, making it applicable to macro and community-scale studies of bamboo invasion in other regions of the world. Patches serve as fundamental components in landscape ecology; thus, this method can also provide insights into the expansion of other invasive plant species. The results provide robust evidence for the native invasion of moso bamboo, underscoring the necessity for attention to bamboo expansion and effective control measures on a broader scale. The expansion area in evergreen broadleaf forests far exceeded that in deciduous broadleaf forests and evergreen coniferous forests. Considering canopy closure, moso bamboo expands faster into open forest types (canopy closure less than 0.4), shrublands, and grasslands.
We found that climate, topography, and soil significantly influence the expansion rate of bamboo forests. The primary interaction of expansion rates is among climate factors (surface solar radiation and sunshine duration, and annual precipitation and surface solar radiation) and the combined climate and soil (total nitrogen content and sunshine duration, and total phosphorus content and surface solar radiation). We emphasize that the quality and density of native vegetation, such as native evergreen broad-leaved forests, should be protected. Conducting timely monitoring of environmental variables is essential. Additionally, enforcing appropriate control measures like cutting bamboo and blocking soil moisture acquisition is crucial for controlling bamboo expansion.
Author Contributions
Conceptualization, J.W. and Y.Z.; methodology, J.W. and D.L.; software, J.W.; validation, J.W., D.L. and Z.L.; formal analysis, J.W.; resources, Y.Z.; data curation, S.Z.; writing—original draft preparation, J.W.; writing—review and editing, Y.Z., J.D., Z.C. and S.Z.; visualization, Z.L.; supervision, Z.C. and J.D.; project administration, S.Z and Y.Z; funding acquisition, S.Z and Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the “Hunan Natural Science Foundation, grant number 2023JJ31017”, the “Special Funding for Basic Education Development from Hunan Provincial Department of Finance, grant number 2022No.69”, the “Key Projects of Hunan Provincial Department of Education, grant number 23A0232”, and “the Ministry of Science and Technology of The People’s Republic of China”, grants 2019YFD1100400.
Data Availability Statement
Dataset available on request from the authors.
Acknowledgments
We are particularly grateful to the National Forestry and Grassland Administration for providing the national forest inventory data.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
In Table A1, Figure A1 and Figure A2, a1: ≥10 °C accumulated temperature; a2: annual mean temperature; a3: annual mean minimum temperature; a4: annual mean maximum temperature; b1: annual precipitation; b2: relative humidity; b3: potential mean evapotranspiration; c: near-surface wind speed; d1: surface solar radiation; d2: sunshine duration; e1: dominant soil unit; e2: texture class; e3: reference bulk density; e4: AWC for rootable soil depth; f1: organic carbon content; f2: total nitrogen content; f3: total phosphorus content; f4: total potassium content; f5: pH in water; f6: base saturation as the percentage of CECsoil; f7: cation exchange capacity of the soil; g1: elevation; and g2: slope.
Table A1.
The interaction values and interaction types of the top 10 factors for the expansion of bamboo into 12 different habitats. The colors are differentiated as follows: orange for climate ∩ climate, green for climate ∩ soil, blue for climate ∩ terrain, yellow for soil ∩ topography, and purple for terrain ∩ terrain.  denotes enhance_nonlinear,
denotes enhance_nonlinear,  denotes enhance_bi. (A: rainfed cropland, herbaceous cover, and irrigated cropland; B: open evergreen broadleaved forest (0.15 < fc < 0.4); C: closed evergreen broadleaved forest (fc > 0.4); D: open evergreen needle-leaved forest (0.15 < fc < 0.4); E: closed evergreen needle-leaved forest (fc > 0.4); F: open deciduous broadleaved forest (0.15 < fc < 0.4); G: closed deciduous broadleaved forest (fc > 0.4); H: shrubland; I: evergreen shrubland; J: grassland; K: water, wetlands; and L: impervious surfaces.).
denotes enhance_bi. (A: rainfed cropland, herbaceous cover, and irrigated cropland; B: open evergreen broadleaved forest (0.15 < fc < 0.4); C: closed evergreen broadleaved forest (fc > 0.4); D: open evergreen needle-leaved forest (0.15 < fc < 0.4); E: closed evergreen needle-leaved forest (fc > 0.4); F: open deciduous broadleaved forest (0.15 < fc < 0.4); G: closed deciduous broadleaved forest (fc > 0.4); H: shrubland; I: evergreen shrubland; J: grassland; K: water, wetlands; and L: impervious surfaces.).
denotes enhance_nonlinear, denotes enhance_bi. (A: rainfed cropland, herbaceous cover, and irrigated cropland; B: open evergreen broadleaved forest (0.15 < fc < 0.4); C: closed evergreen broadleaved forest (fc > 0.4); D: open evergreen needle-leaved forest (0.15 < fc < 0.4); E: closed evergreen needle-leaved forest (fc > 0.4); F: open deciduous broadleaved forest (0.15 < fc < 0.4); G: closed deciduous broadleaved forest (fc > 0.4); H: shrubland; I: evergreen shrubland; J: grassland; K: water, wetlands; and L: impervious surfaces.).
Table A1.
The interaction values and interaction types of the top 10 factors for the expansion of bamboo into 12 different habitats. The colors are differentiated as follows: orange for climate ∩ climate, green for climate ∩ soil, blue for climate ∩ terrain, yellow for soil ∩ topography, and purple for terrain ∩ terrain. denotes enhance_nonlinear, denotes enhance_bi. (A: rainfed cropland, herbaceous cover, and irrigated cropland; B: open evergreen broadleaved forest (0.15 < fc < 0.4); C: closed evergreen broadleaved forest (fc > 0.4); D: open evergreen needle-leaved forest (0.15 < fc < 0.4); E: closed evergreen needle-leaved forest (fc > 0.4); F: open deciduous broadleaved forest (0.15 < fc < 0.4); G: closed deciduous broadleaved forest (fc > 0.4); H: shrubland; I: evergreen shrubland; J: grassland; K: water, wetlands; and L: impervious surfaces.).
denotes enhance_nonlinear, denotes enhance_bi. (A: rainfed cropland, herbaceous cover, and irrigated cropland; B: open evergreen broadleaved forest (0.15 < fc < 0.4); C: closed evergreen broadleaved forest (fc > 0.4); D: open evergreen needle-leaved forest (0.15 < fc < 0.4); E: closed evergreen needle-leaved forest (fc > 0.4); F: open deciduous broadleaved forest (0.15 < fc < 0.4); G: closed deciduous broadleaved forest (fc > 0.4); H: shrubland; I: evergreen shrubland; J: grassland; K: water, wetlands; and L: impervious surfaces.).| ID | Rank | Factor | Inter_ Value | Inter- Action | ID | Rank | Factor | Inter_ Value | Inter- Action | ID | Rank | Factor | Inter_ Value | Inter- Action |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | 1 | d1∩d2 | 0.779 | | B | 1 | c∩b2 | 0.822 | | C | 1 | d1∩d2 | 0.775 | |
| 2 | a3∩d2 | 0.707 | | 2 | a2∩b2 | 0.593 | | 2 | a4∩d2 | 0.744 | | |||
| 3 | a2∩d2 | 0.705 | | 3 | a4∩b2 | 0.486 | | 3 | a2∩d2 | 0.741 | | |||
| 4 | a1∩d2 | 0.678 | | 4 | a1∩b2 | 0.472 | | 4 | a1∩d2 | 0.682 | | |||
| 5 | b1∩d1 | 0.677 | | 5 | b3∩b2 | 0.447 | | 5 | f1∩d2 | 0.614 | | |||
| 6 | a3∩d1 | 0.664 | | 6 | a2∩c | 0.427 | | 6 | f2∩d2 | 0.611 | | |||
| 7 | d1∩a2 | 0.649 | | 7 | f7∩b2 | 0.426 | | 7 | e2∩d2 | 0.611 | | |||
| 8 | a1∩d1 | 0.649 | | 8 | f1∩b2 | 0.426 | | 8 | a3∩d2 | 0.603 | | |||
| 9 | a4∩d2 | 0.648 | | 9 | f2∩b2 | 0.426 | | 9 | f3∩d2 | 0.602 | | |||
| 10 | f3∩d2 | 0.629 | | 10 | f6∩b2 | 0.426 | | 10 | d1∩a4 | 0.601 | | |||
| D | 1 | b1∩e3 | 0.373 | | E | 1 | b1∩d1 | 0.492 | | F | 1 | b1∩g2 | 0.482 | |
| 2 | a3∩c | 0.354 | | 2 | d1∩d2 | 0.489 | | 2 | a1∩g2 | 0.476 | | |||
| 3 | d1∩c | 0.351 | | 3 | a3∩d2 | 0.468 | | 3 | b3∩g2 | 0.468 | | |||
| 4 | a2∩c | 0.331 | | 4 | a2∩d2 | 0.453 | | 4 | a1∩d2 | 0.455 | | |||
| 5 | d1∩e3 | 0.322 | | 5 | a4∩d2 | 0.453 | | 5 | g1∩a3 | 0.447 | | |||
| 6 | a1∩c | 0.313 | | 6 | a1∩d2 | 0.434 | | 6 | g2∩a4 | 0.444 | | |||
| 7 | a3∩e3 | 0.307 | | 7 | a3∩d1 | 0.432 | | 7 | g2∩f4 | 0.444 | | |||
| 8 | e4∩a3 | 0.291 | | 8 | b1∩a3 | 0.424 | | 8 | g1∩g2 | 0.437 | | |||
| 9 | f1∩a3 | 0.291 | | 9 | d1∩a2 | 0.416 | | 9 | a3∩g2 | 0.433 | | |||
| 10 | f2∩a3 | 0.288 | | 10 | b1∩a2 | 0.414 | | 10 | a3∩f4 | 0.429 | | |||
| G | 1 | d1∩d2 | 0.431 | | H | 1 | b3∩d2 | 0.883 | | I | 1 | b1∩d1 | 0.881 | |
| 2 | a3∩di | 0.418 | | 2 | b3∩b2 | 0.880 | | 2 | d1∩c | 0.876 | | |||
| 3 | a3∩d2 | 0.412 | | 3 | b1∩d1 | 0.876 | | 3 | d1∩b2 | 0.864 | | |||
| 4 | a4∩d2 | 0.404 | | 4 | a4∩d2 | 0.870 | | 4 | d1∩a4 | 0.859 | | |||
| 5 | d1∩a2 | 0.404 | | 5 | a3∩d2 | 0.861 | | 5 | d1∩a2 | 0.858 | | |||
| 6 | a2∩d2 | 0.399 | | 6 | a2∩d2 | 0.851 | | 6 | d1∩d2 | 0.857 | | |||
| 7 | d1∩a4 | 0.391 | | 7 | b2∩d2 | 0.850 | | 7 | d1∩b3 | 0.856 | | |||
| 8 | a4∩b2 | 0.38 | | 8 | g1∩b2 | 0.821 | | 8 | a1∩d1 | 0.855 | | |||
| 9 | a2∩b2 | 0.371 | | 9 | a4∩b2 | 0.815 | | 9 | a3∩d1 | 0.853 | | |||
| 10 | a3∩b2 | 0.367 | | 10 | c∩b2 | 0.806 | | 10 | d1∩g2 | 0.850 | | |||
| J | 1 | b1∩b2 | 0.750 | | K | 1 | a1∩d2 | 0.898 | | L | 1 | c∩b2 | 0.656 | |
| 2 | b1∩b3 | 0.735 | | 2 | a3∩d2 | 0.879 | | 2 | e4∩c | 0.643 | | |||
| 3 | b1∩d1 | 0.719 | | 3 | a2∩d2 | 0.877 | | 3 | f6∩c | 0.643 | | |||
| 4 | b1∩a4 | 0.718 | | 4 | a3∩g2 | 0.853 | | 4 | d1∩c | 0.641 | | |||
| 5 | f3∩b1 | 0.718 | | 5 | e4∩d2 | 0.851 | | 5 | f3∩c | 0.636 | | |||
| 6 | b1∩a3 | 0.717 | | 6 | d1∩d2 | 0.851 | | 6 | d1∩d2 | 0.628 | | |||
| 7 | b1∩e3 | 0.715 | | 7 | f2∩d2 | 0.851 | | 7 | f3∩d1 | 0.624 | | |||
| 8 | b1∩f4 | 0.713 | | 8 | f7∩d2 | 0.851 | | 8 | a2∩c | 0.622 | | |||
| 9 | d1∩d2 | 0.712 | | 9 | f3∩d2 | 0.851 | | 9 | b1∩d1 | 0.618 | | |||
| 10 | b1∩a2 | 0.711 | | 10 | f6∩d2 | 0.851 | | 10 | f2∩c | 0.615 | |
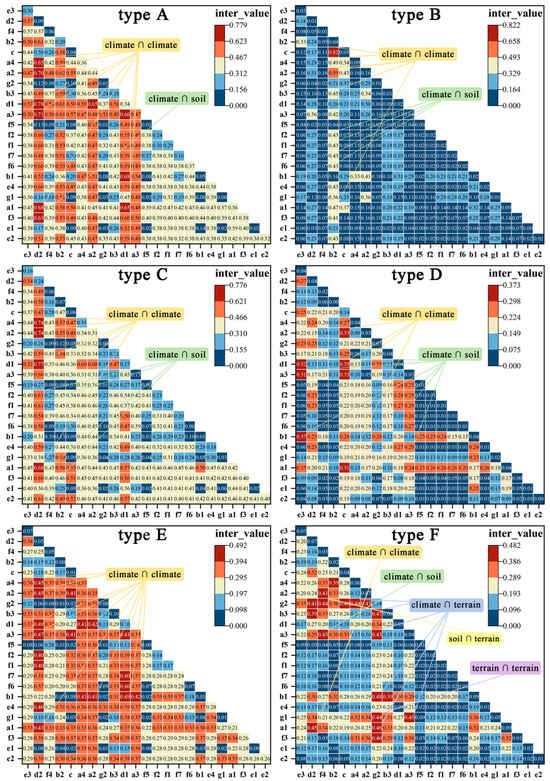
Figure A1.
The interaction values and heatmap of the 23 driving factors during the expansion of bamboo into habitats A: (rainfed cropland, herbaceous cover, and irrigated cropland), B: open evergreen broadleaved forest (0.15 < fc < 0.4)), C: (closed evergreen broadleaved forest (fc > 0.4)), D: (open evergreen needle-leaved forest (0.15 < fc < 0.4)), E: (closed evergreen needle-leaved forest (fc > 0.4)), and F: (open deciduous broadleaved forest (0.15 < fc < 0.4).
Figure A1.
The interaction values and heatmap of the 23 driving factors during the expansion of bamboo into habitats A: (rainfed cropland, herbaceous cover, and irrigated cropland), B: open evergreen broadleaved forest (0.15 < fc < 0.4)), C: (closed evergreen broadleaved forest (fc > 0.4)), D: (open evergreen needle-leaved forest (0.15 < fc < 0.4)), E: (closed evergreen needle-leaved forest (fc > 0.4)), and F: (open deciduous broadleaved forest (0.15 < fc < 0.4).
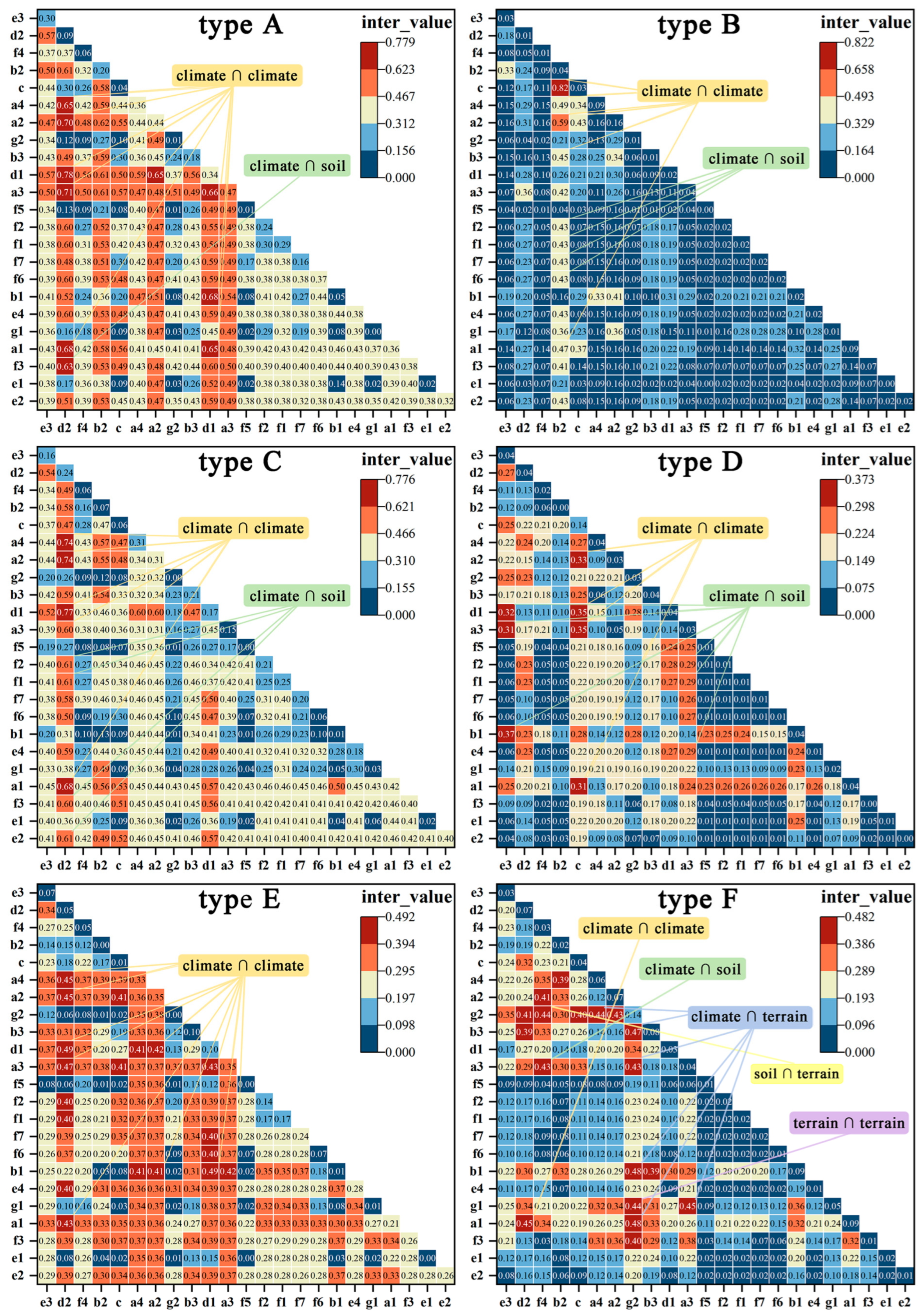
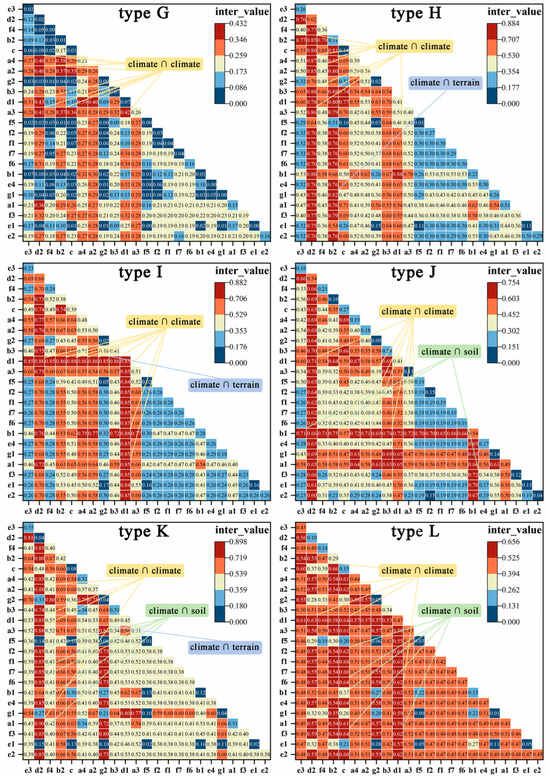
Figure A2.
The interaction values and heatmap of the 23 driving factors during the expansion of bamboo into habitats G: (closed deciduous broadleaved forest (fc > 0.4)), H: (shrubland), I: (evergreen shrubland), J: (grassland), K: (water, wetlands), and L: (impervious surfaces).
Figure A2.
The interaction values and heatmap of the 23 driving factors during the expansion of bamboo into habitats G: (closed deciduous broadleaved forest (fc > 0.4)), H: (shrubland), I: (evergreen shrubland), J: (grassland), K: (water, wetlands), and L: (impervious surfaces).
References
- MacDougall, A.S.; Gilbert, B.; Levine, J.M. Plant invasions and the niche. J. Ecol. 2009, 97, 609–615. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Walker, L.R.; Whiteaker, L.D.; Mueller-Dombois, D.; Matson, P.A. Biological invasion by myrica faya alters ecosystem development in Hawaii. Science 1987, 238, 802–804. [Google Scholar] [CrossRef]
- Foxcroft, L.C.; Pickett, S.T.; Cadenasso, M.L. Expanding the conceptual frameworks of plant invasion ecology. Perspect. Plant. Ecol. 2011, 13, 89–100. [Google Scholar] [CrossRef]
- Simberloff, D.; Von, H.B. Positive interactions of nonindigenous species: Invasional meltdown? Biol. Invasions. 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Valéry, L.; Fritz, H.; Lefeuvre, J.C.; Simberloff, D. In search of a real definition of the biological invasion phenomenon itself. Biol. Invasions 2008, 10, 1345–1351. [Google Scholar] [CrossRef]
- Valéry, L.; Fritz, H.; Lefeuvre, J.C.; Simberloff, D. Invasive species can also be native. Trends Ecol. Evol. 2009, 24, 585. [Google Scholar] [CrossRef]
- Lacerda, A.E.B. Understanding the long-term impact of bamboos on secondary forests: A case for bamboo management in southern brazi. Diversity 2021, 13, 567. [Google Scholar] [CrossRef]
- Greig, C.; Robertson, C.; Lacerda, A.E.B. Spectral-temporal modelling of bamboo-dominated forest succession in the Atlantic Forest of Southern Brazil. Ecol. Model. 2018, 384, 316–332. [Google Scholar] [CrossRef]
- Montti, L.; Villagra, M.; Campanello, P.I.; Gatti, M.G.; Goldstein, G. Functional traits enhance invasiveness of bamboos over co-occurring tree saplings in the semideciduous Atlantic Forest. Acta Oecologica 2014, 54, 36–44. [Google Scholar] [CrossRef]
- Ibáñez, I.; Petri, L.; Barnett, D.T.; Beaury, E.M.; Blumenthal, D.M.; Corbin, J.D.; Diez, J.; Dukes, J.S.; Early, R.; Pearse, I.S.; et al. Combining local, landscape, and regional geographies to assess plant community vulnerability to invasion impact. Ecol. Appl. 2023, 33, e2821. [Google Scholar] [CrossRef]
- Dai, L.; Wang, Y.; Su, D.; Zhou, L.; Yu, D.; Lewis, B.J.; Qi, L. Major forest types and the evolution of sustainable forestry in China. Environ. Manag. 2011, 48, 1066–1078. [Google Scholar] [CrossRef] [PubMed]
- Wenhua, L. Degradation and restoration of forest ecosystems in China. For. Ecol. Manag. 2004, 201, 33–41. [Google Scholar] [CrossRef]
- Bai, S.; Wang, Y.; Conant, R.; Zhou, G.; Xu, Y.; Wang, N.; Fang, F.; Chen, J. Can native clonal moso bamboo encroach on adjacent natural forest without human intervention? Sci. Rep. 2016, 6, 31504. [Google Scholar] [CrossRef]
- Lima, R.A.; Rother, D.C.; Muler, A.E.; Lepsch, I.F.; Rodrigues, R.R. Bamboo overabundance alters forest structure and dynamics in the Atlantic Forest hotspot. Biol. Conserv. 2012, 147, 32–39. [Google Scholar] [CrossRef]
- Long, K.; Yin, R.; Kardol, P.; Wei, Q.; Li, Y.; Huang, J. Bamboo invasion alters Collembola community composition varying with life-forms. Pest Manag. Sci. 2023, 79, 2517–2526. [Google Scholar] [CrossRef]
- Song, Q.N.; Lu, H.; Liu, J.; Yang, J.; Yang, G.Y.; Yang, Q.P. Accessing the impacts of bamboo expansion on NPP and N cycling in evergreen broadleaved forest in subtropical China. Sci. Rep. 2017, 7, 40383. [Google Scholar] [CrossRef]
- Liu, C.; Zhou, Y.; Qin, H.; Liang, C.; Shao, S.; Fuhrmann, J.J.; Chen, J.; Xu, Q. Moso bamboo invasion has contrasting effects on soil bacterial and fungal abundances, co-occurrence networks and their associations with enzyme activities in three broadleaved forests across subtropical China. For. Ecol. Manag. 2021, 498, 119549. [Google Scholar] [CrossRef]
- Tong, R.; Zhou, B.; Jiang, L.; Cao, Y.; Ge, X.; Yang, Z. Influence of Moso bamboo invasion on forest plants and soil: A review. Acta Ecol. Sinica. 2019, 39, 3808–3815. [Google Scholar] [CrossRef]
- Wei, Q.; Yin, R.; Huang, J.; Vogler, A.P.; Li, Y.; Miao, X.; Kardol, P. The diversity of soil mesofauna declines after bamboo invasion in subtropical China. Sci. Total Environ. 2021, 789, 147982. [Google Scholar] [CrossRef]
- Fang, H.; Liu, Y.; Bai, J.; Li, A.; Deng, W.; Bai, T.; Liu, X.; Lai, M.; Feng, Y.; Zhang, J.; et al. Impact of Moso Bamboo (Phyllostachys edulis) expansion into Japanese cedar plantations on soil fungal and bacterial community compositions. Forests 2022, 13, 1190. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, W.; Bai, S.H.; Xu, Z.; Yun, Z.; Zhang, W. Linking Phyllostachys edulis (moso bamboo) growth with soil nitrogen cycling and microbial community of plant-soil system: Effects of plant age and niche differentiation. Ind. Crops. Prod. 2022, 177, 114520. [Google Scholar] [CrossRef]
- Li, S.; Xie, D.; Ge, X.; Dong, W.; Luan, J. Altered diversity and functioning of soil and root-associated microbiomes by an invasive native plant. Plant. Soil. 2022, 473, 235–249. [Google Scholar] [CrossRef]
- Meng, M.; Lin, J.; Guo, X.; Liu, X.; Wu, J.; Zhao, Y.; Zhang, J. Impacts of forest conversion on soil bacterial community composition and diversity in subtropical forests. Catena 2019, 175, 167–173. [Google Scholar] [CrossRef]
- Jin, W.; Tu, J.; Wu, Q.; Peng, L.; Xing, J.; Liang, C.; Shao, S.; Chen, J.; Xu, Q.; Qin, H. Moso bamboo expansion decreased soil heterotrophic respiration but increased arbuscular mycorrhizal mycelial respiration in a subtropical broadleaved forest. For. Ecosyst. 2023, 10, 100116. [Google Scholar] [CrossRef]
- Okutomi, K.; Shinoda, S.; Fukuda, H. Causal analysis of the invasion of broad-leaved forest by bamboo in Japan. J. Veg. Sci. 1996, 7, 723–728. [Google Scholar] [CrossRef]
- Yu, L.; Wei, J.; Li, D.; Zhong, Y.; Zhang, Z. Explaining Landscape Levels and Drivers of Chinese Moso Bamboo Forests Based on the Plus Model. Forests 2023, 14, 397. [Google Scholar] [CrossRef]
- Lieurance, D.; Cooper, A.; Young, A.L.; Gordon, D.R.; Flory, S.L. Running bamboo species pose a greater invasion risk than clumping bamboo species in the continental United States. J. Nat. Conserv. 2018, 43, 39–45. [Google Scholar] [CrossRef]
- Canavan, S.; Richardson, D.M.; Visser, V.; Roux, J.J.L.; Vorontsova, M.S.; Wilson, J.R. The global distribution of bamboos: Assessing correlates of introduction and invasion. Aob. Plants. 2017, 9, plw078. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y.; Zhang, Y.; Booth, T.; He, X. Changes of carbon stocks in bamboo stands in China during 100 years. For. Ecol. Manag. 2009, 258, 1489–1496. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y.; Zhang, X.; Guo, Y. Carbon stock changes in bamboo stands in China over the last 50 years. Acta Ecol. Sinica. 2008, 28, 5218–5227. [Google Scholar] [CrossRef]
- Du, H.; Mao, F.; Li, X.; Zhou, G.; Xu, X.; Han, N.; Sun, S.; Gao, G.; Gui, L.; Li, Y.; et al. Mapping global bamboo forest distribution using multisource remote sensing data. IEEE J. Stars. 2018, 11, 1458–1471. [Google Scholar] [CrossRef]
- Zhang, Y. A study of the effects of climatic fluctuation on Chinese fir and bamboo ecological environment in subtropical regions in China. Q. J. Appl. Meteorol. 1995, 6, 75–82. [Google Scholar]
- Ge, Z.; Huang, S.; Ouyang, M.; Luan, F.; Fang, X.; Yang, Q.; Liu, J.; Song, Q. Stand Characteristics Rather than Soil Properties Contribute More to the Expansion of Moso Bamboo (Phyllostachys edulis) into Its Neighboring Forests in Subtropical Region. Forests 2022, 13, 2159. [Google Scholar] [CrossRef]
- Suzuki, S.; Nakagoshi, N. Expansion of bamboo forests caused by reduced bamboo-shoot harvest under different natural and artificial conditions. Ecol. Res. 2008, 23, 641–647. [Google Scholar] [CrossRef]
- Dong, C. Study on growth effect on new stands of Phyllostachys heterocycla cv. Pubescens through rhizome expanding and growing. J. Bamboo Res. 2003, 22, 30–33. [Google Scholar]
- Cao, Y.; Zhou, B.; Wang, X.; Wang, G. The photosynthetic characteristics of Moso bamboo (Phyllostachys pubescens) for different canopy leaves. Adv. Mater. Res. 2013, 726, 4274–4279. [Google Scholar] [CrossRef]
- National Forestry and Grassland Administration National Park Administration. Chinese bamboo: The Most Abundant Resources and the Most Ambitious Industry. 8 November 2022. Available online: https://www.forestry.gov.cn/c/www/ggscy/32007.jhtml (accessed on 10 August 2024).
- GBT 26424-2010; Technical Regulations for Inventory for Forest Management Planning and Design. National Technical Committee on Forest Resources Standardization: Beijing, China, 2010.
- Zhou, B.; Fu, M. Review on bamboo’s underground rhizome-root system research. For. Res. 2004, 17, 8. [Google Scholar] [CrossRef]
- Courant, R.; John, F. The Fundamental Ideas of the Integral and Differential Calculus. In Introduction to Calculus and Analysis; Springer: New York, NY, USA, 1989; Volume I, p. 141. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.; Zou, Y.; Yang, Y. Assessment of Forest Type Data in Global Land Cover Products over China. Remote Sens. Tech. Appl. 2023, 38, 341–352. [Google Scholar]
- Wang, J.F.; Li, X.H.; Christakos, G.; Liao, Y.L.; Zhang, T.; Gu, X.; Zheng, X.Y. Geographical detectors-based health risk assessment and its application in the neural tube defects study of the Heshun Region, China. Int. J. Geogr. Inf. Sci. 2010, 24, 107–127. [Google Scholar] [CrossRef]
- Chen, J.; Wu, T.; Zou, D.; Liu, L.; Wu, X.; Gong, W.; Zhu, X.; Li, R.; Hao, J.; Hu, G.; et al. Magnitudes and patterns of large-scale permafrost ground deformation revealed by Sentinel-1 InSAR on the central Qinghai-Tibet Plateau. Remote Sens. Environ. 2022, 268, 112778. [Google Scholar] [CrossRef]
- Hou, M.; Deng, Y.; Yao, S. Coordinated relationship between urbanization and grain production in China: Degree measurement, spatial differentiation and its factors detection. J. Clean. Prod. 2022, 331, 129957. [Google Scholar] [CrossRef]
- Yang, M.; Shi, L.; Liu, B. Risks assessment and driving forces of urban environmental accident. J. Clean. Prod. 2022, 340, 130710. [Google Scholar] [CrossRef]
- Song, Y.; Wang, J.; Ge, Y.; Xu, C. An optimal parameters-based geographical detector model enhances geographic characteristics of explanatory variables for spatial heterogeneity analysis: Cases with different types of spatial data. GISci. Remote Sens. 2020, 57, 593–610. [Google Scholar] [CrossRef]
- Isagi, Y.; Torii, A. Range Expansion and Its Mechanisms in a Naturalized Bamboo Species, Phyllostachys pubescens, in Japan. J. Sustain. For. 1997, 6, 127–141. [Google Scholar] [CrossRef]
- Hierro, J.L.; Maron, J.L.; Callaway, R.M. A biogeographical approach to plant invasions: The importance of studying exotics in their introduced and native range. J. Ecol. 2004, 93, 5–15. [Google Scholar] [CrossRef]
- Davis, M.; Grime, J.P.; Thompson, K. Fluctuating resources in plant communities: A general theory of invasibility. J. Ecol. 2005, 88, 528–534. [Google Scholar] [CrossRef]
- Gao, P.; Chen, S.; Guo, Z.; Yang, Q. Comparison of photosynthetic light response models of Sophora flavescens and Catsia tora under Phyllostachys edulis seedlings. For. Res. 2018, 31, 156–163. [Google Scholar] [CrossRef]
- Ying, W.; Jin, J.; Jiang, H.; Zhang, X.; Lu, X.; Chen, X.; Zhang, J. Satellite-based detection of bamboo expansion over the past 30 years in Mount Tianmushan, China. Int. J. Remote Sens. 2016, 37, 2908–2922. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, Y.; Yan Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Lou, Y.; Fan, Y.; Dai, Q.; Wang, Z.; Weipeng, K.U.; Zhao, M. Relationship between vertical structure and overall species diversity in an evergreen deciduous broad-leaved forest community of tianmu mountain natural reserve. Acta Ecol. Sinica. 2021, 41, 8568–8577. [Google Scholar] [CrossRef]
- Zeng, X.; Luo, H.; Lu, J.; Zhu, X.; He, Y.; Gong, C.; Ren, Z.; Huang, D.; Song, Q.; Yang, Q. The Process of Patchy Expansion for Bamboo (Phyllostachys edulis) at the Bamboo–Broadleaf Forest Interface: Spreading and Filling in Order. Forests 2024, 15, 438. [Google Scholar] [CrossRef]
- Li, X.; Mao, F.; Du, H.; Zhou, G.; Xing, L.; Liu, T.; Han, N.; Liu, Y.; Zhu, D.; Zheng, J.; et al. Spatiotemporal evolution and impacts of climate change on bamboo distribution in China. J. Environ. Manag. 2019, 248, 109265. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, S.; Li, Y.; Xie, S. Research Review in the Effects of Climate Factors on Bamboo Growth. J. Bamboo Res. 2011, 30, 9–12+17. [Google Scholar]
- Long, C.; Li, Y.; Zhu, X.; Mu, B. Characteristics of solar radiation energy in communities at different successional stages in karst forests. Guizhou Meteorol. 1998, 5, 22–25. (In Chinese) [Google Scholar]
- Yang, Z.; Yue, C.; Wang, K. Analysis on Causes of Photosynthetic Midday-depress in Camellia sinensis Leaves. J. Zhejiang. For. Sci. Tech. 2022, 3, 74–76+83. [Google Scholar] [CrossRef]
- He, Q.; Ye, H.; Li, P.; Tong, X.; Hua, X. Effect of Light Intensity on Growth of Potted Ornamental Bamboo. J. Zhejiang Forest Sci. Tech. 2007, 2, 51–53. [Google Scholar] [CrossRef]
- Liu, S.; Zhou, G.; Bai, S. Light intensity changes on Cunninghamia lanceolata in mixed stands with different concentrations of Phyllostachys pubescens. J. Zhejiang A. For. Univ. 2011, 28, 550–554. [Google Scholar]
- Wang, Y.; Wang, K.; Zhang, Y.; Qiao, Y. Lack of gas exchange for CO in B. bractis2Response to concentration and light intensity. J. Northwest For. Uni. 2008, 1+6–9+13. [Google Scholar] [CrossRef]
- Ojo, A.R.; Sadiku, N.A. Slenderness coefficient and growth characteristics of Africa giant bamboo: Bambusa vulgaris Schrad. ex JC Wendl. Adv. Bamboo Sci. 2023, 2, 100017. [Google Scholar] [CrossRef]
- Gagnon, P.R.; Platt, W.J.; Moser, E.B. Response of a native bamboo [Arundinaria gigantea (Walt.) Muhl.] in a wind-disturbed forest. For. Ecol. Manag. 2007, 241, 288–294. [Google Scholar] [CrossRef]
- Luo, W.; Zhang, Q.; Wang, P.; Luo, J.; She, C.; Guo, X.; Yuan, J.; Sun, Y.; Guo, R.; Li, Z.; et al. Unveiling the impacts moso bamboo invasion on litter and soil properties: A meta-analysis. Sci. Total Environ. 2024, 909, 168532. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, G.; Wu, Z.; Wen, X.; Zhong, H.; Zhong, Z.; Yang, C.; Bian, F.; Gai, X. Responses of soil nutrients and microbial communities to intercropping medicinal plants in moso bamboo plantations in subtropical China. Environ. Sci. Pollut. Res. 2020, 27, 2301–2310. [Google Scholar] [CrossRef]
- Chu, H.; Ni, H.; Su, W.; Fan, S.; Long, Y.; Sun, Y. Enhanced Nitrogen Fertilizer Input Alters Soil Carbon Dynamics in Moso Bamboo Forests, Impacting Particulate Organic and Mineral-Associated Carbon Pools. Forests 2023, 14, 2460. [Google Scholar] [CrossRef]
- Mao, X.; Zheng, J.; Yu, W.; Guo, X.; Xu, K.; Zhao, R.; Xioa, L.; Wang, M.; Jiang, Y.; Zhang, S.; et al. Climate-induced shifts in composition and protection regulate temperature sensitivity of carbon decomposition through soil profile. Soil Biol. Biochem. 2022, 172, 108743. [Google Scholar] [CrossRef]
- Ding, Z.; Wang, L.; Liu, X.; Xu, X. Productivity of Phyllostachys pubescens stands and soil nutrient in Huoshan of Anhui. Non Wood For. Res. 2011, 29, 72–76. [Google Scholar] [CrossRef]
- Li, C.; Shi, Y.; Zhou, G.; Zhou, Y.; Xu, L.; Tong, L.; Liu, X. Effects of different management approaches on soil carbon dynamics in Moso bamboo forest ecosystems. Catena 2018, 169, 59–68. [Google Scholar] [CrossRef]
- Li, T.; Dong, W.; Zheng, J.; Liu, J.; Li, B.; Zhong, F. A Study on Site Condition of Phyllostachys nigra Stand of Technology Utilization. World Bamboo Ratt. 2007, 4, 17–21. [Google Scholar]
- Zhang, H.; Zhang, T.; Deng, D.; Zhuang, W. The Research Progress on Interactions Between Soil and the Ecological Effect of Bamboo. J. Sichuan For. Sci. Tech. 2015, 36, 17–22. [Google Scholar] [CrossRef]
- Torii, A. Estimation of range expansion rate of bamboo stands using aerial photographs—Case study on Mt. Hachiman, Shiga Prefecture, and Mt. Otoko, Kyoto Prefecture, Japan. Jpn. J. Ecol. 1998, 48, 37–47. (In Japanese) [Google Scholar]
- Ohno, T. A study of the patterns of bamboo forest expansion in Kishiwada city, Osaka. J. Jpn Inst. Landsc. 2002, 65, 603–608. (In Japanese) [Google Scholar] [CrossRef]
- Swanson, F.J.; Kratz, T.K.; Caine, N.; Woodmansee, R.G. Landform effects on ecosystem patterns and processes. BioScience 1988, 38, 92–98. [Google Scholar] [CrossRef]
- Takano, K.T.; Hibino, K.; Numata, A.; Oguro, M.; Aiba, M.; Shiogama, H.; Takayabu, I.; Nakashizuka, T. Detecting latitudinal and altitudinal expansion of invasive bamboo Phyllostachys edulis and Phyllostachys bambusoides (poaceae) in japan to project potential habitats under 1.5 °C–4.0 °C global warming. Ecol. Evol. 2017, 7, 9848–9859. [Google Scholar] [CrossRef]
- Shi, P.; Preisler, H.K.; Quinn, B.K.; Zhao, J.; Huang, W.; Röll, A.; Cheng, X.; Li, H.; Hölscher, D. Precipitation is the most crucial factor determining the distribution of moso bamboo in mainland china. Glob. Ecol. Conserv. 2020, 22, e00924. [Google Scholar] [CrossRef]
- Linger, E.; Hogan, J.A.; Cao, M.; Zhang, W.F.; Yang, X.F.; Hu, Y.H. Precipitation influences on the net primary productivity of a tropical seasonal rainforest in Southwest China: A 9-year case study. For. Ecol. Manag. 2020, 467, 118153. [Google Scholar] [CrossRef]
- Zhou, X.; Geng, X.; Yin, G.; Hänninen, H.; Hao, F.; Zhang, X.; Fu, Y.H. Legacy effect of spring phenology on vegetation growth in temperate China. Agr. For. Meteorol. 2020, 281, 107845. [Google Scholar] [CrossRef]
- Zeng, X.; Hu, Z.; Chen, A.; Yuan, W.; Hou, G.; Han, D.; Liang, M.; Di, K.; Cao, R.; Luo, D. The global decline in the sensitivity of vegetation productivity to precipitation from 2001 to 2018. Glob. Change Biol. 2022, 28, 6823–6833. [Google Scholar] [CrossRef] [PubMed]
- Fernández-de-Uña, L.; McDowell, N.G.; Canellas, I.; Gea-Izquierdo, G. Disentangling the effect of competition, co2 and climate on intrinsic water-use efficiency and tree growth. J. Ecol. 2016, 104, 678–690. [Google Scholar] [CrossRef]
- Wu, X.P.; Liu, S.; Luan, J.; Wang, Y.; Gao, X.; Chen, C. Nitrogen addition alleviates drought effects on water status and growth of Moso bamboo (Phllostachys edulis). For. Ecol. Manag. 2023, 530, 120768. [Google Scholar] [CrossRef]
- Fuke, P.; Kumar, M.; Sawarkar, A.D.; Pandey, A.; Singh, L. Role of microbial diversity to influence the growth and environmental remediation capacity of bamboo: A review. Ind. Crop. Prod. 2021, 167, 113567. [Google Scholar] [CrossRef]
- Yang, S.J.; Sun, M.; Zhang, Y.J.; Cochard, H.; Cao, K.F. Strong leaf morphological, anatomical, and physiological responses of a subtropical woody bamboo (Sinarundinaria nitida) to contrasting light environments. Plant Ecol. 2014, 215, 97–109. [Google Scholar] [CrossRef]
- Xue, J.; Wang, Z.; Lv, X. A review of the interaction between tree roots and soil environment. J. Nanjing For. Univ. (Nat. Sci.). 2022, 3, 79–84. (In Chinese) [Google Scholar]
- Li, Y.; Yang, Q.; Guo, Z.; Chen, S.; Hu, J. Damage characteristics of Phyllostachys edulis stands under continuous high temperature and drought. For. Res. 2015, 28, 646–653. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).