
Total Polyphenol Content and Antioxidant Activity of Leaves and Fine Roots as Indicators of Drought Resistance in the Native Quercus robur and Alien Quercus rubra




Abstract
1. Introduction
- (1)
- The extent to which the additive action of low light intensity and progressive soil drying affects the polyphenol content and the oxidative capacity of leaves and fine roots;
- (2)
- Whether the drought intensity correlates with the polyphenol content and the antioxidant capacity;
- (3)
- The extent to which the two investigated species differ in their response.
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Extraction and Quantification of Total Polyphenols
2.3. Trolox-Equivalent Antioxidant Capacity (TEAC) Assay
2.4. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bohnert, H.J.; Nelson, D.E.; Jensen, R.G. Adaptations to environmental stresses. Plant Cell 1995, 7, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Król, A.; Amarowicz, R.; Weidner, S. Changes in the composition of phenolic compounds and antioxidant properties of grapevine roots and leaves (Vitis vinifera L.) under continuous of long-term drought stress. Acta Physiol. Plant. 2014, 36, 1491–1499. [Google Scholar] [CrossRef]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Kranner, I.; Beckett, R.P.; Wornik, S.; Zorn, M.; Pfeifhofer, H.W. Revival of a resurrection plant correlates with its antioxidant status. Plant J. 2002, 31, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Surveswaran, S.; Cai, Y.Z.; Corke, H.; Sun, M. Systematic evaluation of natural phenolic antioxidants from 133 Indian medicinal plants. Food Chem. 2007, 102, 938–953. [Google Scholar] [CrossRef]
- Krishnaiah, D.; Sarbatly, R.; Nithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233. [Google Scholar] [CrossRef]
- Agati, G.; Tattini, M. Multiple functional roles of flavonoids in photoprotection. New Phytol. 2010, 186, 786–793. [Google Scholar] [CrossRef]
- Motilva, M.-J.; Serra, A.; Macià, A. Analysis of food polyphenols by ultra high-performance liquid chromatography coupled to mass spectrometry: An overview. J. Chromatogr. A 2013, 1292, 66–82. [Google Scholar] [CrossRef]
- Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. J. Food Biochem. 2020, 44, e13394. [Google Scholar] [CrossRef]
- Lang, Y.; Gao, N.; Zang, Z.; Meng, X.; Lin, Y.; Yang, S.; Yang, Y.; Jin, Z.; Li, B. Classification and antioxidant assays of polyphenols: A review. J. Future Foods 2024, 4, 193–204. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P.; Stavroulaki, V.; Sumbele, S. “Carbon gain vs. water saving, growth vs. defence”: Two dilemmas with soluble phenolics as a joker. Plant Sci. 2014, 227, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Karabourniotis, G.; Liakopoulos, G. Phenolic compounds in plant cuticles: Physiological and ecological aspects. Adv. Plant Physiol. 2005, 8, 33–47. [Google Scholar]
- Galasso, G.; Conti, F.; Peruzzi, L.; Alessandrini, A.; Ardenghi, N.M.G.; Bacchetta, G.; Banfi, E.; Barberis, G.; Bernardo, L.; Bouvet, D.; et al. A second update to the checklist of the vascular flora alien to Italy. Plant Biosyst. 2024, 158, 297–340. [Google Scholar] [CrossRef]
- Stanek, M.; Zubek, S.; Stefanowicz, A.M. Differences in phenolics produced by invasive Quercus rubra and native plant communities induced changes in soil microbial properties and enzymatic activity. For. Ecol. Manag. 2021, 482, 118901. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Francis Salifu, K.; Davis, A.S. Drought susceptibility and recovery of transplanted Quercus rubra seedlings in relation to root system morphology. Ann. For. Sci. 2009, 66, 504. [Google Scholar] [CrossRef]
- Sander, I. Quercus rubra L. In Silvics of North America: Volume 2. Hardwoods; USDA Agricultural Handbook 654; Burns, R.M., Honkala, B.H., Eds.; USDA: Washington, DC, USA, 1990; Volume 2, pp. 727–738. ISBN 654. [Google Scholar]
- Di Iorio, A.; Caspani, A.C.; Beatrice, P.; Montagnoli, A. Drought-related root morphological traits and non-structural carbohydrates in the seedlings of the alien Quercus rubra and the native Quercus robur: Possible implication for invasiveness. Front. For. Glob. Chang. 2024, 7, 1307340. [Google Scholar] [CrossRef]
- Nicolescu, V.-N.; Vor, T.; Mason, W.L.; Bastien, J.-C.; Brus, R.; Henin, J.-M.; Kupka, I.; Lavnyy, V.; La Porta, N.; Mohren, F.; et al. Ecology and management of northern red oak (Quercus rubra L. syn. Q. borealis F. Michx.) in Europe: A review. For. Int. J. For. Res. 2020, 93, 481–494. [Google Scholar] [CrossRef]
- Weidner, S.; Karolak, M.; Karamać, M.; Kosińska, A.; Amarowicz, R. Phenolic compounds and properties of antioxidants in grapevine roots (Vitis vinifera L.) under drought stress followed by recovery. Acta Soc. Bot. Pol. 2009, 78, 97–103. [Google Scholar] [CrossRef]
- Giertych, M.J.; Karolewski, P.; Oleksyn, J. Carbon allocation in seedlings of deciduous tree species depends on their shade tolerance. Acta Physiol. Plant. 2015, 37, 216. [Google Scholar] [CrossRef]
- Varela, M.C.; Arslan, I.; Reginato, M.A.; Cenzano, A.M.; Luna, M.V. Phenolic compounds as indicators of drought resistance in shrubs from Patagonian shrublands (Argentina). Plant Physiol. Biochem. 2016, 104, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Wood, K.E.A.; Kobe, R.K.; McCarthy-Neumann, S. Tree seedling shade tolerance arises from interactions with microbes and is mediated by functional traits. Front. Ecol. Evol. 2023, 11, 1224540. [Google Scholar] [CrossRef]
- Zagoskina, N.V.; Zubova, M.Y.; Nechaeva, T.L.; Kazantseva, V.V.; Goncharuk, E.A.; Katanskaya, V.M.; Baranova, E.N.; Aksenova, M.A. Polyphenols in Plants: Structure, Biosynthesis, Abiotic Stress Regulation, and Practical Applications (Review). Int. J. Mol. Sci. 2023, 24, 13874. [Google Scholar] [CrossRef] [PubMed]
- Munakata, R.; Larbat, R.; Duriot, L.; Olry, A.; Gavira, C.; Mignard, B.; Hehn, A.; Bourgaud, F. Polyphenols from Plant Roots. Recent Adv. Polyphen. Res. 2019, 6, 207–236. [Google Scholar] [CrossRef]
- Kecis, H.; Abdelouahab, Y.; Bagues, M.; Gali, L.; Mekircha, F.; Alloun, W.; Nagaz, K. Phenolic profile and bioactivity of the aerial part and roots of Mentha rotundifolia L. grown in two different localities in northeastern Algeria: A comparative study. Biocatal. Agric. Biotechnol. 2023, 47, 102581. [Google Scholar]
- Tattini, M.; Galardi, C.; Pinelli, P.; Massai, R.; Remorini, D.; Agati, G. Differential accumulation of flavonoids and hydroxycinnamates in leaves of Ligustrum vulgare under excess light and drought stress. New Phytol. 2004, 163, 547–561. [Google Scholar] [CrossRef]
- Zhang, T.J.; Zheng, J.; Yu, Z.C.; Huang, X.D.; Zhang, Q.L.; Tian, X.S.; Peng, C.L. Functional characteristics of phenolic compounds accumulated in young leaves of two subtropical forest tree species of different successional stages. Tree Physiol. 2018, 38, 1486–1501. [Google Scholar] [CrossRef]
- Yu, Z.C.; Zheng, X.T.; Lin, W.; Cai, M.L.; Zhang, Q.L.; Peng, C.L. Different photoprotection strategies for mid- and late-successional dominant tree species in a high-light environment in summer. Environ. Exp. Bot. 2020, 171, 103927. [Google Scholar] [CrossRef]
- Yu, Z.C.; Lin, W.; Zheng, X.T.; Cai, M.L.; Zhang, T.J.; Luo, Y.N.; Peng, C.L. Interpretation of the difference in shade tolerance of two subtropical forest tree species of different successional stages at the transcriptome and physiological levels. Tree Physiol. 2021, 41, 1669–1684. [Google Scholar] [CrossRef]
- Lassouane, N.; Aïd, F.; Quinet, M.; Lutts, S. Phenolic acids and flavonoids classes in Acacia arabica (Lam) Willd. seedling during water stress and subsequent re-hydration. Plant Soil 2024, 496, 449–471. [Google Scholar] [CrossRef]
- Quettier-Deleu, C.; Gressier, B.; Vasseur, J.; Thierry, D.; Claude, B.; Luyckx, M.; Cazin, M.; Cazin, J.-C.; Bailleul, F.; Trotin, F. Phenolic compounds and antioxidant activities of buckwheat (Fagopyrum esculentum Moench) hulls and flour. J. Ethnopharmacol. 2000, 72, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Vanhakylä, S.; Salminen, J.-P. Mass Spectrometric Fingerprint Mapping Reveals Species-Specific Differences in Plant Polyphenols and Related Bioactivities. Molecules 2023, 28, 6388. [Google Scholar] [CrossRef] [PubMed]
- Barber, N.A.; Fahey, R.T. Consequences of phenology variation and oxidative defenses in Quercus. Chemoecology 2015, 25, 261–270. [Google Scholar] [CrossRef]
- Salminen, J.P.; Karonen, M. Chemical ecology of tannins and other phenolics: We need a change in approach. Funct. Ecol. 2011, 25, 325–338. [Google Scholar] [CrossRef]
- Ossipov, V.; Salminen, J.P.; Ossipova, S.; Haukioja, E.; Pihlaja, K. Gallic acid and hydrolysable tannins are formed in birch leaves from an intermediate compound of the shikimate pathway. Biochem. Syst. Ecol. 2003, 31, 3–16. [Google Scholar] [CrossRef]
- Riipi, M.; Ossipov, V.; Lempa, K.; Haukioja, E.; Koricheva, J.; Ossipova, S.; Pihlaja, K. Seasonal changes in birch leaf chemistry: Are there trade-offs between leaf growth and accumulation of phenolics? Oecologia 2002, 130, 380–390. [Google Scholar] [CrossRef]
- Jaafar, H.Z.E.; Ibrahim, M.H.; Fakri, N.F.M. Impact of soil field water capacity on secondary metabolites, phenylalanine ammonia-lyase (PAL), malondialdehyde (MDA) and photosynthetic responses of Malaysian Kacip Fatimah (Labisia pumila Benth). Molecules 2012, 17, 7305–7322. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.E. Reduced photoinhibition under low irradiance enhanced kacip fatimah (Labisia pumila Benth) secondary metabolites, phenyl alanine lyase and antioxidant activity. Int. J. Mol. Sci. 2012, 13, 5290–5306. [Google Scholar] [CrossRef]
- Muller, R.N.; Kalisz, P.J.; Luken, J.O. Fine root production of astringent phenolics. Oecologia 1989, 79, 563–565. [Google Scholar] [CrossRef]
- Chung, I.M.; Kim, J.J.; Lim, J.D.; Yu, C.Y.; Kim, S.H.; Hahn, S.J. Comparison of resveratrol, SOD activity, phenolic compounds and free amino acids in Rehmannia glutinosa under temperature and water stress. Environ. Exp. Bot. 2006, 56, 44–53. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Janas, K.M.; Cvikrová, M.; Palagiewicz, A.; Szafranska, K.; Posmyk, M.M. Constitutive elevated accumulation of phenylpropanoids in soybean roots at low temperature. Plant Sci. 2002, 163, 369–373. [Google Scholar] [CrossRef]
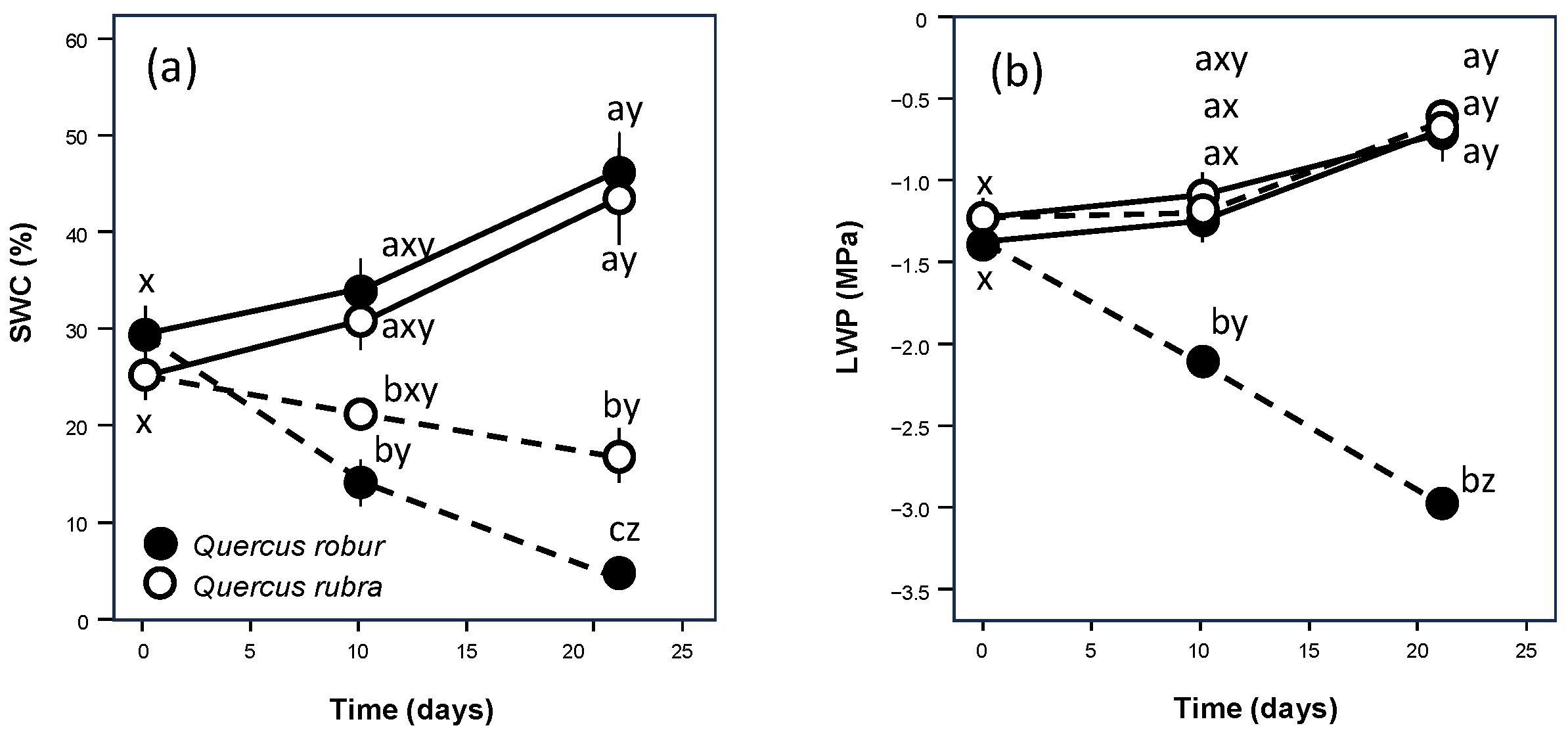
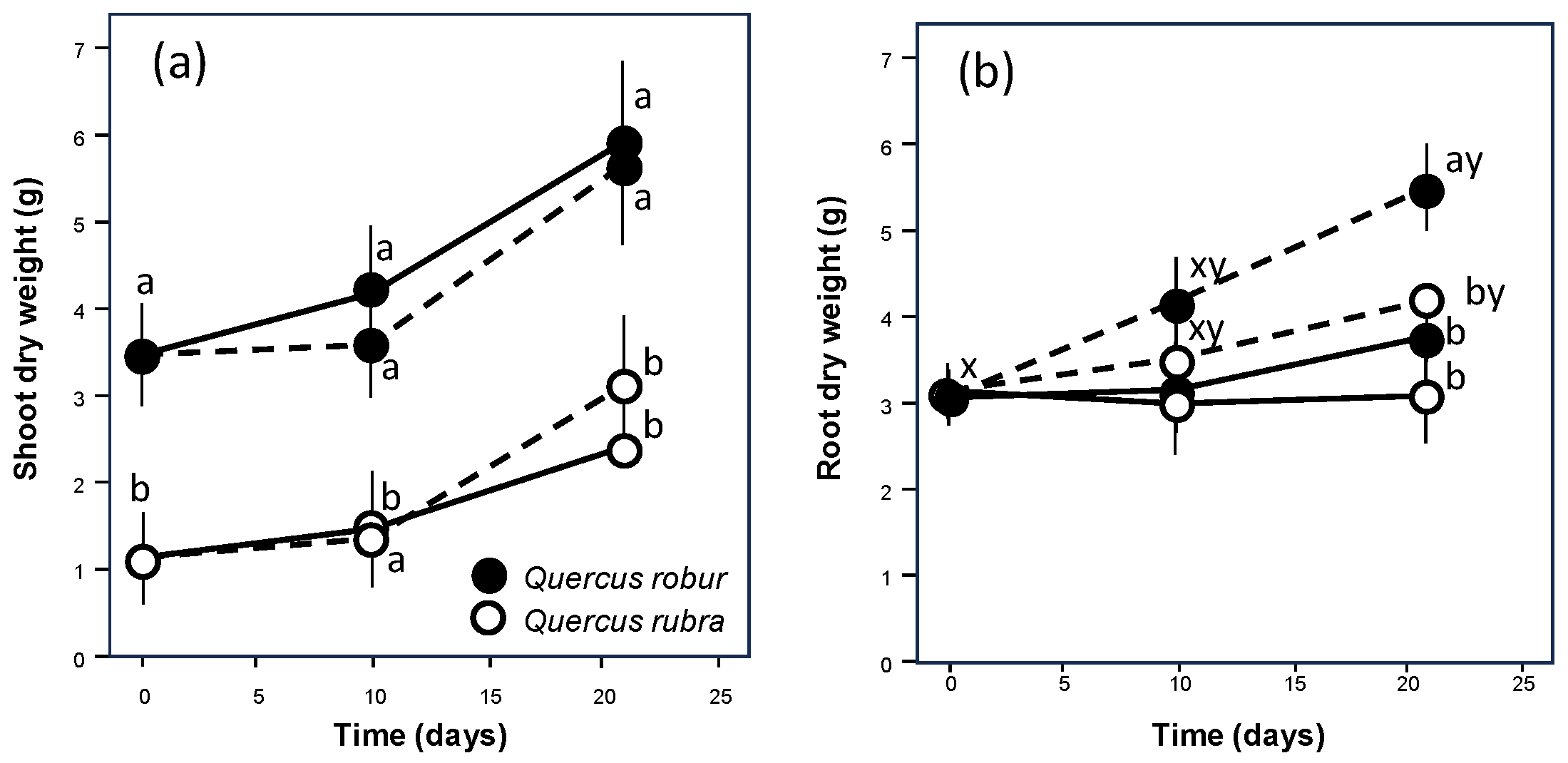
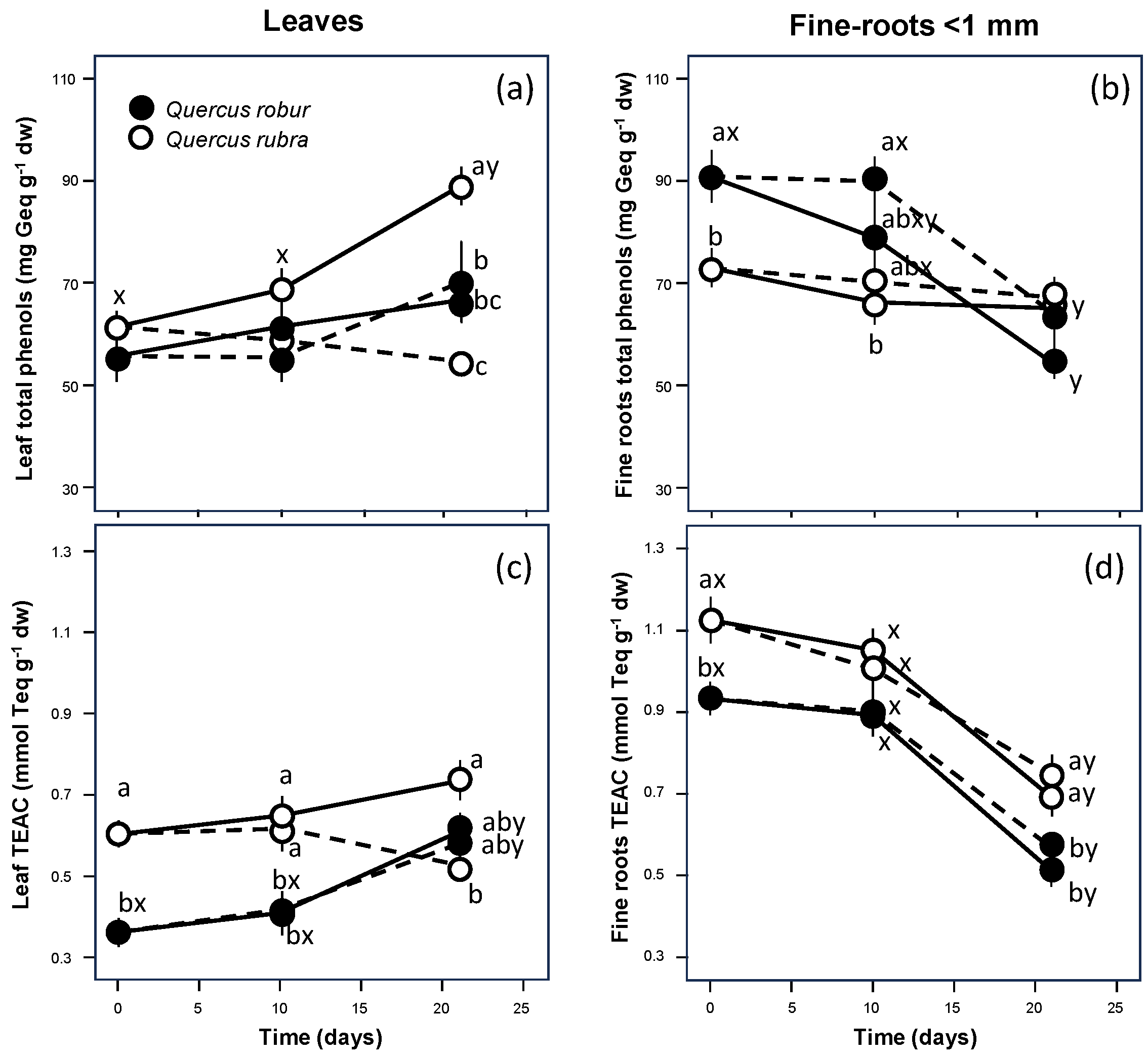
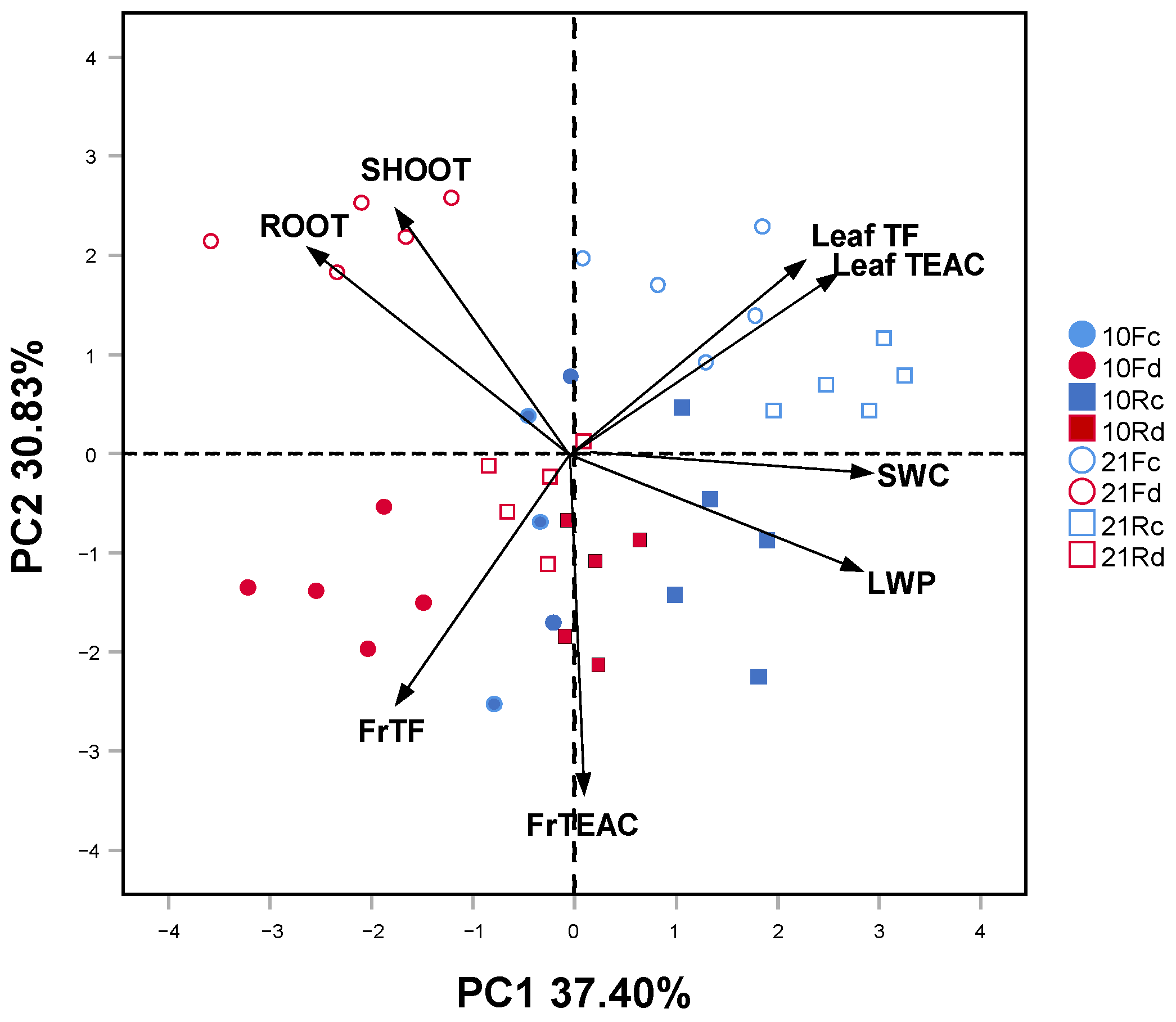

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (T) | Species (S) | Drought (D) | T × S | T × D | S × D | T × S × D | |
|---|---|---|---|---|---|---|---|
| SWC | 0.168 | 0.094 | <0.001 | - | 0.001 | - | - |
| LWP | 0.065 | 0.014 | 0.052 | - | - | 0.002 | - |
| - | - | - | - | ||||
| SHOOT | 0.008 | <0.001 | 0.828 | - | - | - | - |
| ROOT | 0.006 | 0.082 | 0.006 | - | - | - | - |
| - | - | - | - | ||||
| Leaf TP | 0.004 | 0.083 | 0.007 | - | - | 0.015 | 0.04 |
| Leaf TEAC | 0.002 | <0.001 | 0.118 | 0.008 | - | - | - |
| Fr TP | <0.001 | 0.005 | 0.136 | 0.002 | - | - | - |
| Fr TEAC | <0.001 | <0.001 | 0.622 | - | - | - | - |
| LWP | SHOOT | ROOT | Leaf TP | Fr TP | Leaf TEAC | Fr TEAC | |
|---|---|---|---|---|---|---|---|
| SWC | 0.686 ** | −0.137 | −0.625 ** | 0.395 | −0.271 | 0.274 | −0.072 |
| <0.001 | 0.523 | 0.001 | 0.056 | 0.201 | 0.195 | 0.74 | |
| LWP | −0.413 * | −0.582 ** | 0.07 | −0.215 | 0.244 | 0.122 | |
| 0.045 | 0.003 | 0.744 | 0.314 | 0.25 | 0.569 | ||
| SHOOT | 0.567 ** | −0.092 | −0.128 | −0.198 | −0.634 ** | ||
| 0.004 | 0.67 | 0.552 | 0.353 | 0.001 | |||
| ROOT | −0.144 | −0.023 | −0.197 | −0.437 * | |||
| 0.501 | 0.915 | 0.357 | 0.033 | ||||
| Leaf TP | −0.415 * | 0.749 ** | −0.336 | ||||
| 0.044 | <0.001 | 0.108 | |||||
| Fr TP | −0.597 ** | 0.516 ** | |||||
| 0.002 | 0.01 | ||||||
| Leaf TEAC | −0.247 | ||||||
| 0.245 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masante, T.; Cafà, S.; Di Iorio, A. Total Polyphenol Content and Antioxidant Activity of Leaves and Fine Roots as Indicators of Drought Resistance in the Native Quercus robur and Alien Quercus rubra. Forests 2024, 15, 1647. https://doi.org/10.3390/f15091647
Masante T, Cafà S, Di Iorio A. Total Polyphenol Content and Antioxidant Activity of Leaves and Fine Roots as Indicators of Drought Resistance in the Native Quercus robur and Alien Quercus rubra. Forests. 2024; 15(9):1647. https://doi.org/10.3390/f15091647
Chicago/Turabian StyleMasante, Tsvetana, Sofia Cafà, and Antonino Di Iorio. 2024. "Total Polyphenol Content and Antioxidant Activity of Leaves and Fine Roots as Indicators of Drought Resistance in the Native Quercus robur and Alien Quercus rubra" Forests 15, no. 9: 1647. https://doi.org/10.3390/f15091647
APA StyleMasante, T., Cafà, S., & Di Iorio, A. (2024). Total Polyphenol Content and Antioxidant Activity of Leaves and Fine Roots as Indicators of Drought Resistance in the Native Quercus robur and Alien Quercus rubra. Forests, 15(9), 1647. https://doi.org/10.3390/f15091647