Abstract
The red turpentine beetle Dendroctonus valens is an invasive forest pest that has been rapidly spreading northward since its introduction in China. Some ophiostomatalean associates of D. valens are essential contributing factors for this beetle to become an aggressive pine killer in China. In this study, a survey was conducted in a new invasive area of this beetle’s habitat in Liaoning Province. A total of 126 ophiostomatalean fungal associates of D. valens were isolated. Based on the combination of morphological characteristics and phylogeny, these isolates were identified as Ceratocystiopsis jianpingensis sp. nov., along with two new records, Leptographium terebrantis and Ophiostoma gilletteae. Ophiostoma gilletteae was the dominant species, with an isolation rate of 85.7%. The results of this study show that more shared ophiostomatalean fungi are associated with D. valens in China and North America, and accumulate resources for the development of fungal associate-mediated bark beetle management strategies.
1. Introduction
The red turpentine beetle (Coleoptera: Scolytinae), Dendroctonus valens, is a secondary pine-infesting pest, native to North America [1]. Since its introduction to China, it has spread rapidly to seven provinces, killing more than 10 million healthy Pinus tabuliformis trees and posing a serious threat to the health of native P. bungeana and P. sylvestris var. mongolica [2]. The associate of D. valens, Leptographium procerum, is an essential contributing factor to this beetle becoming an aggressive pine killer in China [3].
Leptographium procerum is an ophiostomatalean fungus (Ascomycota, Ophiostomatales), which is commonly associated with beetles [4]. This fungus has been isolated from more than 20 conifer species, and is associated with a wide variety of bark beetles and weevils [5]. In addition, L. procerum is an agent of pine root disease [6]. The success of symbionts of D. valens and L. procerum in China comes from the facilitation of the invasions by of the both the species [7,8]. Leptographium procerum can deplete the host resistance of P. tabuliformis and induce the production of 3-carene, which attracts D. valens, thereby assisting beetle invasion. Dendroctonus valens benefits L. procerum by producing volatiles that inhibit native fungi reproduction.
To date, 31 ophiostomatalean fungal associates of D. valens have been reported [9]. Of these, 15 and 23 species were isolated from China and North America, respectively. In China, Lu et al. identified seven ophiostomatalean associates of D. valens infesting P. tabuliformis and P. bungeana from Shanxi, Shaanxi, and Henan provinces and found that L. procerum and L. sinoprocerum were the dominant species [10,11]. However, different fungal communities of D. valens were subsequently reported. Ten ophiostomatalean species associated with D. valens have been isolated from P. tabuliformis in Shanxi and Shaanxi Provinces [12]. Although only these two species are common in China, the dominant species is L. procerum. Ophiostomatalean associates of D. valens in North America show higher species diversity and fewer shared species compared to those in China, and are mostly endemic [9,13]. Overall, the interactions between most of the fungal associates and D. valens remain unknown.
Despite various management measures, D. valens has spread northward to Liaoning Province, posing a serious threat to native pine forests in northeast China [14]. In this study, an investigation was conducted in July 2024 in a new invasive area of D. valens, which was Jianping County, Liaoning Province. Fungal associates were isolated from adult beetles infesting P. tabuliformis and identified by combining their morphological characteristics and phylogeny. This study revealed additional shared fungal associates of D. valens in North America and China, contributing resources for the development and implementation of fungus-mediated bark beetle management.
2. Materials and Methods
2.1. Isolation of Fungi
Adults of D. valens were collected from dying P. tabuliformis in Jianping County, Liaoning Province, China in July 2024. The beetles were identified based on morphological characteristics [15]. The vigorous beetles were placed in a sterile centrifuge tube at 4 °C for immediate fungal isolation. Approximately 30 tissue pieces were dismembered from each beetle and placed on the surface of 2% malt extract agar medium (MEA; 10 g malt extract, 10 g agar, and 500 mL distilled water), and incubated at 25 °C. As soon as the mycelia grew from the tissue pieces, a single hyphal tip was cut off and placed in fresh medium. This step was repeated two to three times to obtain pure cultures.
2.2. Morphological and Growth Studies
An Olympus BX43 microscope (Olympus Corporation, Tokyo, Japan) was used to observe the microscopic features of fungal isolates. Typical features were recorded using a BioHD-A20c color digital camera (FluoCa Scientific, Shanghai, China). Optimum and extreme growth temperatures for undescribed species were tested under 2% MEA medium at 5–40 °C in darkness.
2.3. PCR Amplification and Phylogenetic Analysis
Thriving mycelia at the edge of the colony were scraped off the medium for DNA extraction. The Fungal Genomic DNA Extraction Kit (Solarbio Co., Ltd., Beijing, China) was used for DNA extraction, following the protocol outlined by the manufacturer. Four gene regions including the internal transcribed spacer regions 1 and 2 of the nuclear ribosomal DNA operon, including the 5.8S region (ITS); the nuclear ribosomal DNA operon, including the internal transcribed spacer 2 region and part of the large subunit (ITS2-LSU); partial β-tubulin gene region (tub2); and partial transcription elongation factor 1-α gene region (tef1-α) were amplified by the primer pairs listed in Table 1. PCR amplification was conducted using 2 × Taq Plus PCR Master Mix (Rui Biotech Co., Ltd., Beijing, China). Sequencing was performed by Rui Biotech Co., Ltd.

Table 1.
Primer pairs and PCR conditions used in this study.
The newly generated sequences underwent the BLASTn tool in GenBank database to identify the genus and complex to which they belonged. The reference sequences were obtained from GenBank. All sequences were aligned using the MAFFT v.7 online web server [21]. Alignments were edited using MEGA 7.0.21 [22]. Phylogenies were calculated using Maximum likelihood (ML) and Bayesian Inference (BI) methods. ML phylogenetic inferences were conducted using RAxML-HPC v.8.2.3 [23], the GTR-GAMMA model was selected, and 1000 bootstrap replicates were run. BI phylogenetic analyses were conducted using MrBayes v. 3.1.2 [24]. The best-fit substitution model was selected using jModelTest v.2.1.7 [25]. One to five million generations were run in four Markov chain Monte Carlo chains to guarantee that the average standard deviation of the split frequencies across different runs was less than 0.01.
3. Results
3.1. Isolation of Fungi
In total, 126 ophiostomatalean strains were isolated from 15 adult D. valens infesting P. tabuliformis. Based on microscopic and cultural characteristics, 13 strains were selected for phylogenetic analysis and detailed morphological and growth studies (Table 2).
Table 2.
Strains isolated from Dendroctonus valens infesting Pinus tabuliformis.
3.2. Phylogenetic Analysis
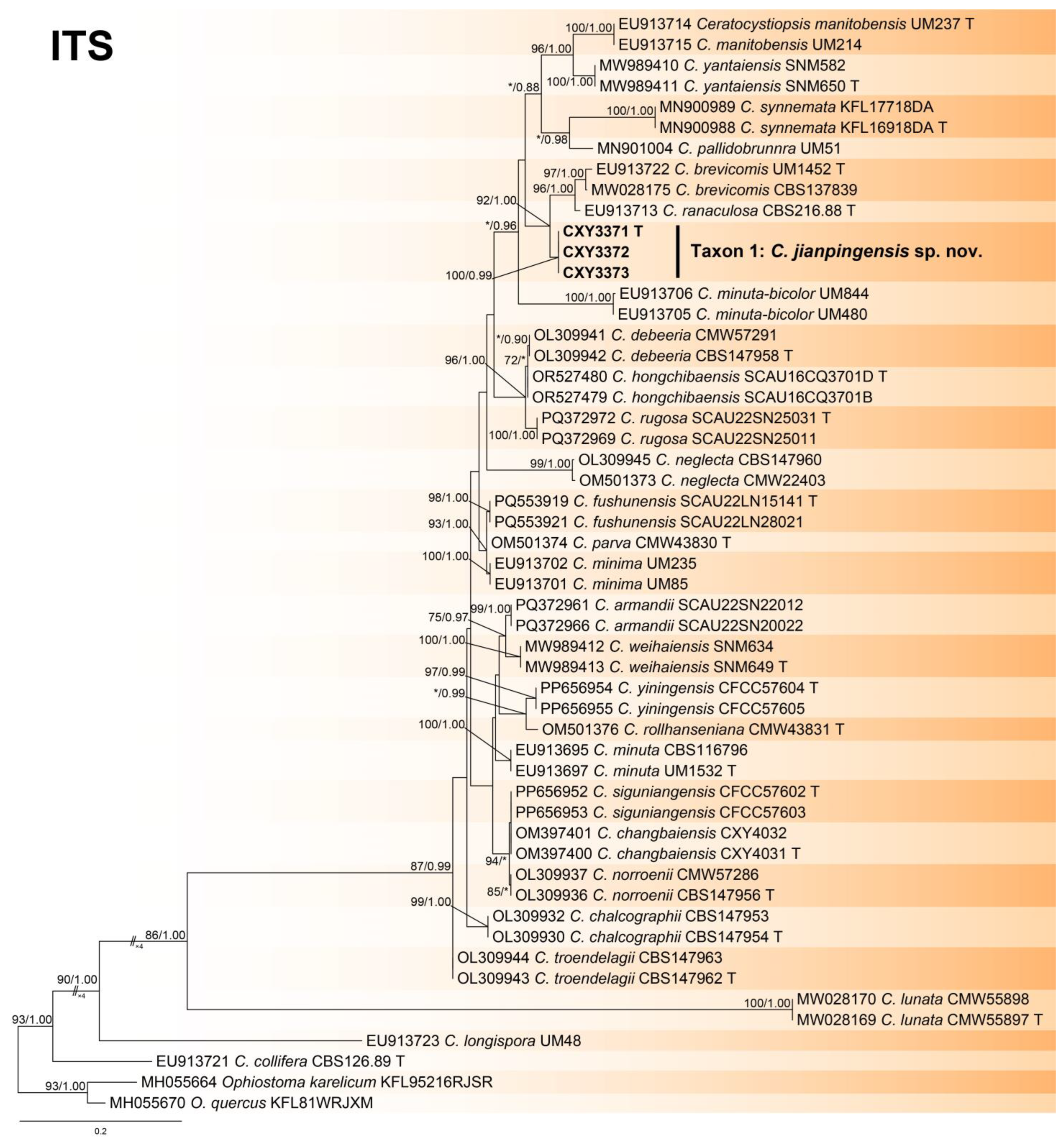
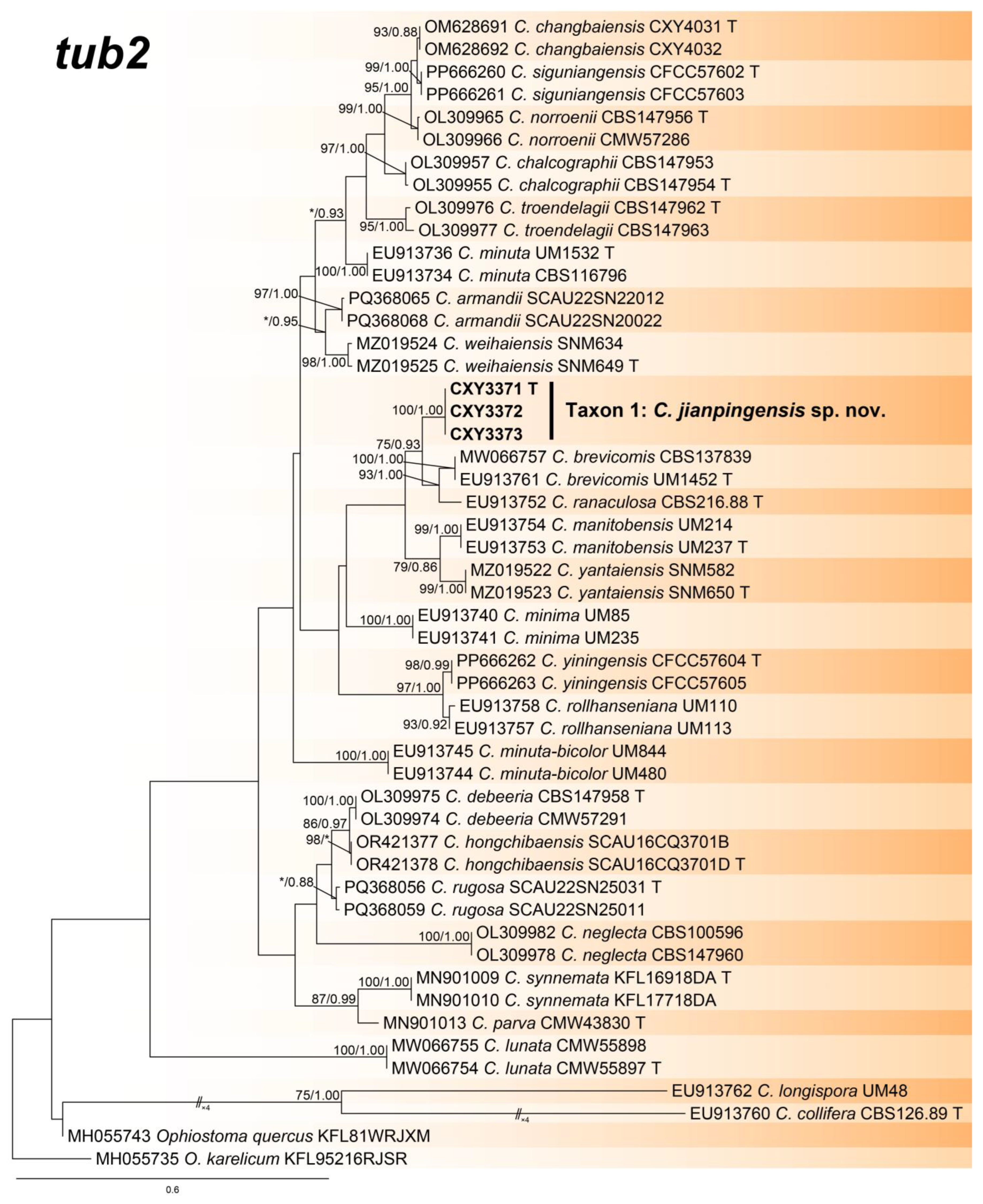
The ITS, tub2, and the combined sequence datasets (containing 659, 435, and 1094 characters, respectively) were used for the phylogenetic analyses of Ceratocystiopsis spp., using the GTR+I+G (ITS sequence dataset) and GTR+G (tub2 and the combined sequence datasets) models, respectively. In three trees, our three isolates formed a robust terminal clade representing an undescribed species, Taxon 1, which was a phylogenetic sister to C. brevicomis and C. ranaculosa (Figure 1, Figure 2 and Figure 3).

Figure 1.
Phylogeny from the maximum likelihood (ML) analysis of the combined sequence dataset (ITS and tub2) of Ceratocystiopsis spp. ML bootstrap values (≥70%)/Bayesian posterior probability values (≥0.85) are presented at the nodes. If not, “*” is presented at the nodes. The newly generated sequences in this study are shown in bold. “T” is ex-type.
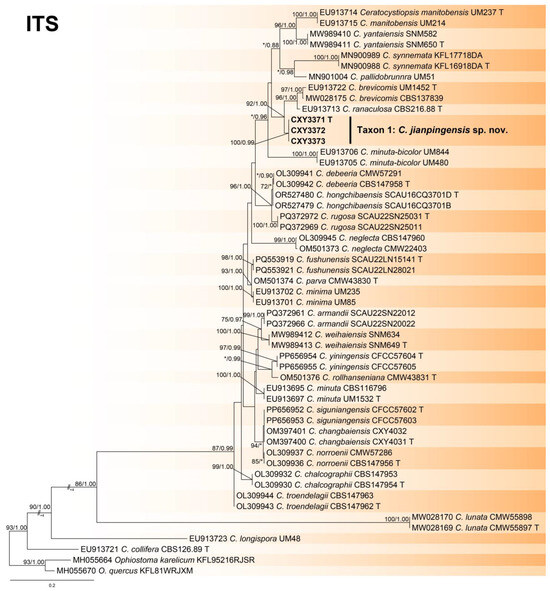
Figure 2.
Phylogeny from the maximum likelihood (ML) analysis of the ITS sequence dataset of Ceratocystiopsis spp. ML bootstrap values (≥70%)/Bayesian posterior probability values (≥0.85) are presented at the nodes. If not, “*” is presented at the nodes. The newly generated sequences in this study are shown in bold. “T” is ex-type.
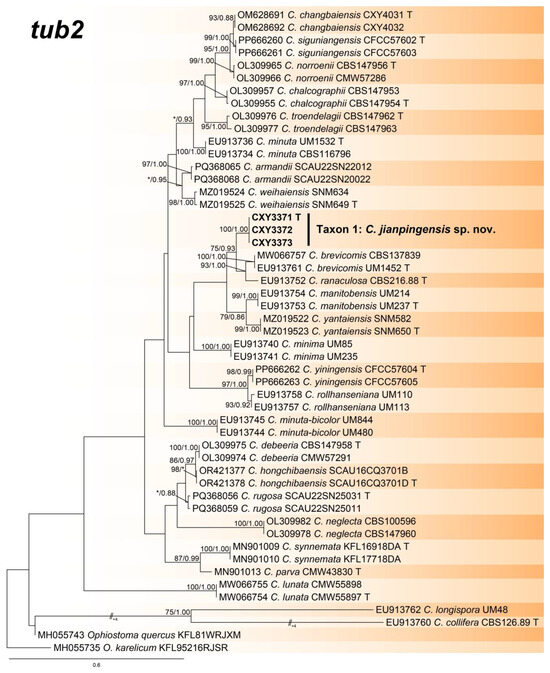
Figure 3.
Phylogeny from the maximum likelihood (ML) analysis of tub2 sequence dataset of Ceratocystiopsis spp. ML bootstrap values (≥70%)/Bayesian posterior probability values (≥0.85) are presented at the nodes. If not, “*” is presented at the nodes. The newly generated sequences in this study are shown in bold. “T” is ex-type.
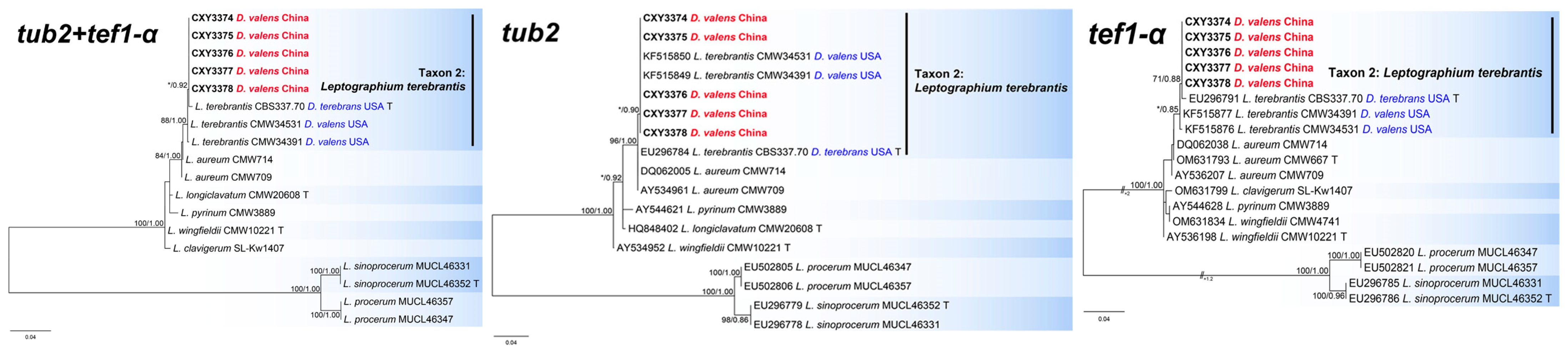
The tub2, tef1-α, and the combined sequence datasets (containing 353, 656, and 1009 characters, respectively) were used for phylogenetic analyses of L. clavigerum complex under K80 (tub2 sequence dataset) and GTR+G (tef1-α and the combined sequence datasets) models, respectively. In tub2 tree, five isolates from this study formed a terminal clade with two isolates associated with D. valens from the USA, representing L. terebrantis (Figure 4). Although the type of L. terebrantis was not distinguishable from the two strains of L. aureum in tub2 tree, all eight strains of L. terebrantis grouped a clade distinct from L. aureum in the tef1-α and the combined trees (Figure 4).

Figure 4.
Phylogenies from the maximum likelihood (ML) analysis of tub2, tef1-α, and the combined sequence datasets of Leptographium clavigerum complex. ML bootstrap values (≥70%)/Bayesian posterior probability values (≥0.85) are presented at the nodes. If not, “*” is presented at the nodes. The newly generated sequences in this study are shown in bold. “T” is ex-type.
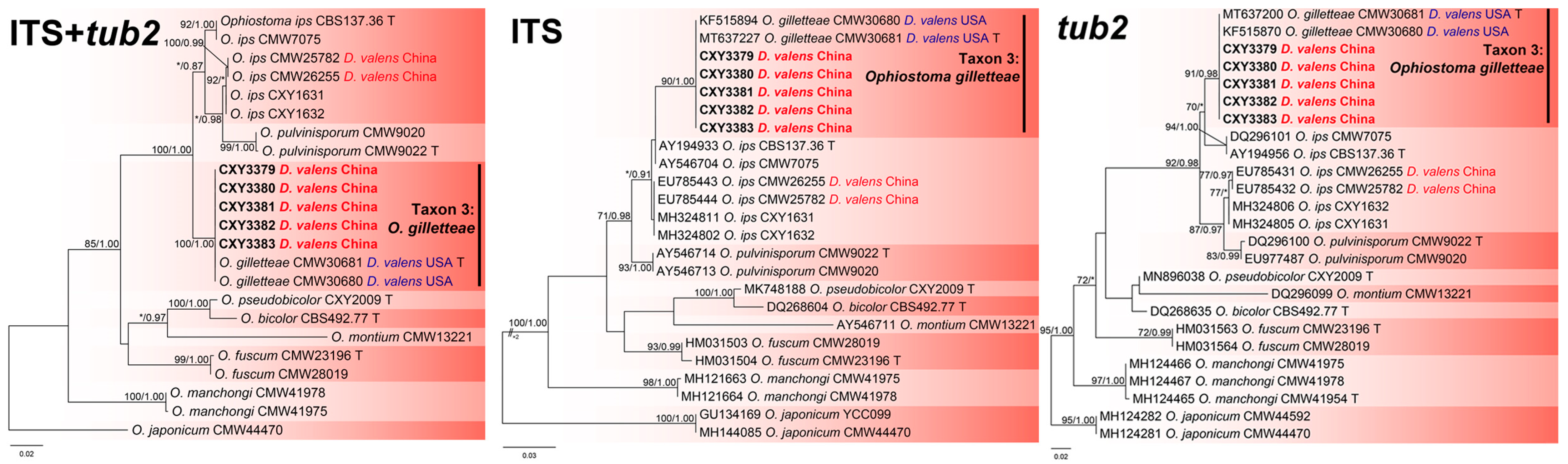
The ITS, tub2, and the combined sequence datasets (containing 685, 271, and 956 characters, respectively) were used for phylogenetic analyses of O. ips complex using the GTR+G (ITS and the combined sequence datasets) and HKY+I (tub2 dataset) models, respectively. The five isolates isolated from this study formed a terminal clade, with two isolates associated with D. valens from the USA, representing O. gilletteae, based on phylogenetic analyses of the three datasets (Figure 5).
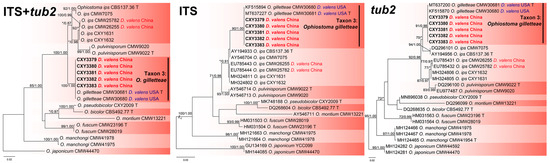
Figure 5.
Phylogenies from the maximum likelihood (ML) analysis of ITS, tub2, and the combined sequence datasets of Ophiostoma ips complex. ML bootstrap values (≥70%)/Bayesian posterior probability values (≥0.85) are presented at the nodes. If not, “*” is presented at the nodes. The newly generated sequences in this study are shown in bold. “T” is ex-type.
3.3. Taxonomy
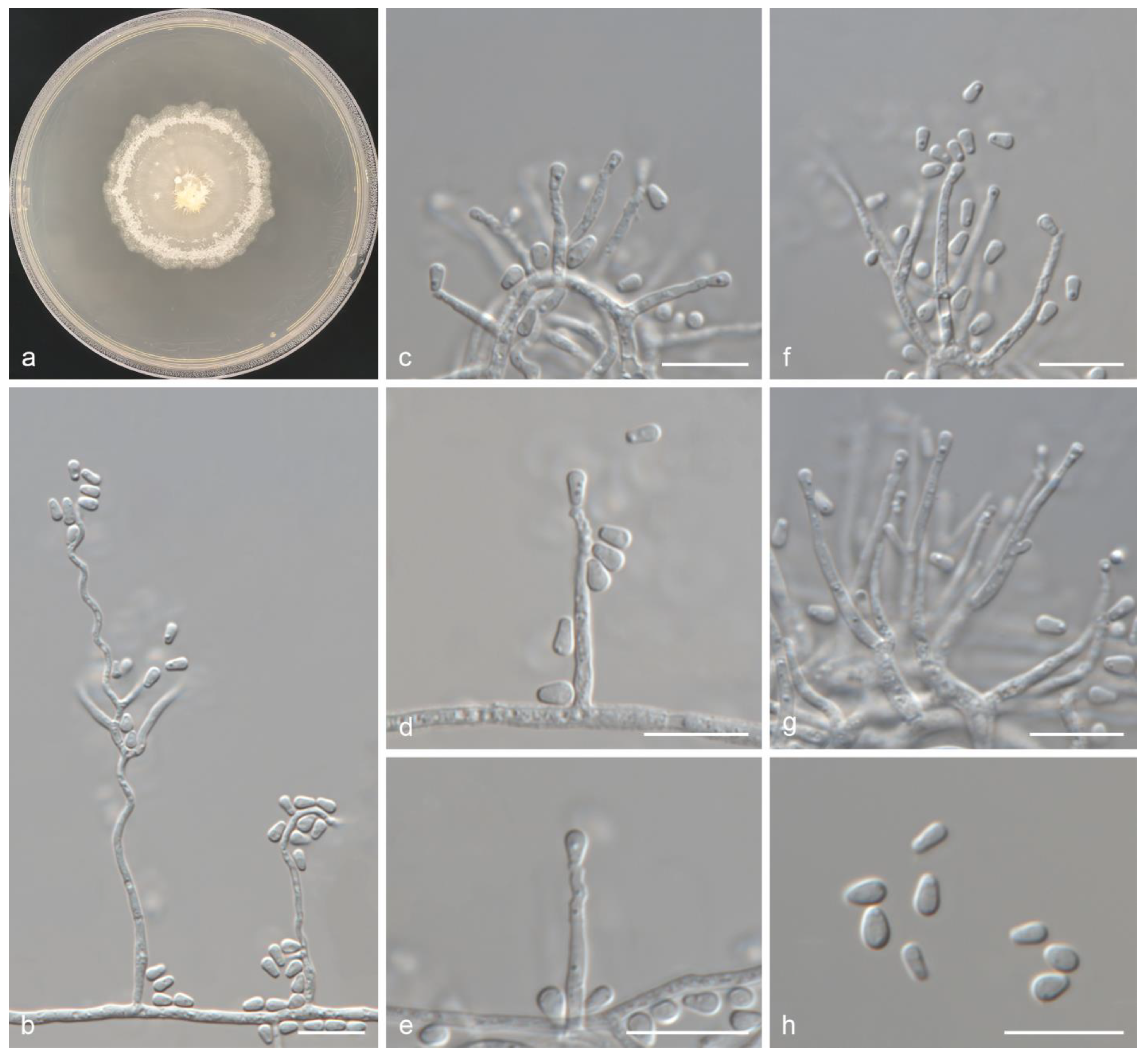
- Ceratocystiopsis jianpingensis Z. Wang and Q. Lu, sp. nov. (Taxon 1, Figure 6).
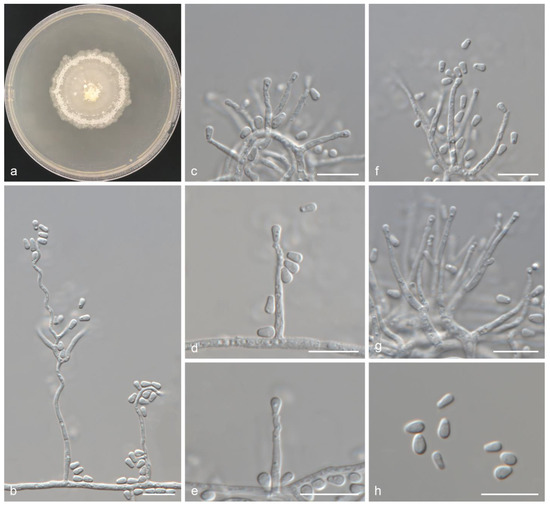 Figure 6. Ceratocystiopsis jianpingensis sp. nov. (a) Twenty-day-old culture on 2% MEA; (b) Serpentine conidiophores; (c–e) Sporothrix-like anamorph: conidiogenous cells and conidia; (f,g) Hyalorhinocladiella-like anamorph: conidiogenous cells and conidia; (h) Conidia. Scale bars: (b–h) = 10 μm.
Figure 6. Ceratocystiopsis jianpingensis sp. nov. (a) Twenty-day-old culture on 2% MEA; (b) Serpentine conidiophores; (c–e) Sporothrix-like anamorph: conidiogenous cells and conidia; (f,g) Hyalorhinocladiella-like anamorph: conidiogenous cells and conidia; (h) Conidia. Scale bars: (b–h) = 10 μm. - Mycobank: 856888
- Etymology: The name refers to Jianping, the county of Liaoning Province, where the fungus was collected.
- Holotype: CXY3371
- Description: Teleomorph unknown. Anamorphs: sporothrix-like and hyalorhinocladiella-like. Conidiophores macronematous or micronematous, simple or branched, erect, occasionally serpentine, arising from the mycelium; conidiogenous cells rough or smooth, hyaline, (5.7–)8.1–13.2(–14.6) × 1.3–1.6(–1.7) μm. Conidia hyaline, smooth, aseptate, obovoid with upper part swollen, (3.1–)3.3–3.9(–4.5) × (1.7–)1.9–2.4(–2.9) μm.
- Culture features: Colonies grew slowly and reached 42.9 mm in 20 days, pure white, occasionally faint yellow. Abundant aerial mycelia, which were either clumpy or spiny, developed on the inoculated plugs. Colonies were dense with irregular margins. Colonies grew fastest at 25 °C and slowest at 5 °C. No growth was observed at 35 °C.
- Associated insects: Dendroctonus valens.
- Hosts: Pinus tabuliformis.
- Material examined: CHINA, Liaoning Province, Chaoyang City, Jianping County, from Dendroctonus valens infesting Pinus tabuliformis, July 2024, Z. Wang and Q. Lu, holotype: CXY3371, ex-type culture CFCC72301, ibid. CFCC72302, and CFCC72303.
- Notes: Ceratocystiopsis jianpingensis was a phylogenetic sister to C. brevicomis and C. ranaculosa (Figure 1, Figure 2 and Figure 3) [26]. Although all three species produce a sporothrix-like asexual morph, C. jianpingensis also produces a hyalorhinocladiella-like asexual morph. Furthermore, C. jianpingensis grew slower than C. brevicomis at 25 °C on 2% MEA (42.9 mm in 20 days vs. 40–50 mm in 13 days). Ceratocystiopsis jianpingensis, C. brevicomis, and C. ranaculosa are associated with pine-infesting Dendroctonus. However, C. jianpingensis is an associate of D. valens in China, and C. brevicomis and C. ranaculosa are associated with D. brevicomis and D. frontalis in North America, respectively [26].
4. Discussion
This study revealed three ophiostomatalean associates of D. valens with P. tabuliformis, including C. jianpingensis sp. nov., and two new records (L. terebrantis and O. gilletteae). Ophiostoma gilletteae was the dominant species, with 108 isolates, and an isolation rate of 85.7%. This species is most frequently isolated from D. valens in western and eastern North America [9,13]. However, O. gilletteae has never been reported in China; therefore, its origin needs to be clarified through population genetics, as was the case with L. procerum [27]. Although we isolated only 14 strains of L. terebrantis (11.1%), this does not prevent it from becoming a species of concern. Similarly to L. procerum, L. terebrantis is associated with a wide variety of weevils and bark beetles in North America, and has been isolated from many dying pine trees [6]. Notably, L. terebrantis has an extremely variable pathogenicity to different hosts [6]. Therefore, the pathogenicity of this species in pine trees native to China needs to be tested urgently. In addition, surveys with larger sample sizes should be conducted to clarify the diversity of ophiostomatalean fungi associated with D. valens.
Leptographium procerum is considered the dominant associate of D. valens in China. However, aside from this species, the three ophiostomatalean fungi we identified in this study are different from the fifteen previously reported in China [9,10,11,12]. The causes of this variation in the fungal assemblages associated with D. valens in different regions may require further investigation using larger sample sizes. Additionally, some associates were neutral or antagonistic to D. valens. For example, O. minus and L. sinoprocerum retard weight gain in larvae of D. valens by depleting the immune defenses of the beetle [28]. Hence, while we are concerned about the pathogenicity of ophiostomatalean fungi to trees, we should also analyze the interactions between these associates and beetles.
Highly heterogeneous forest ecosystems limit the effectiveness of traditional pest control strategies. Innovative biological control programs provide new hope in the fight against forest pests. Dendroctonus valens employs mutualistic microbes to facilitate its invasion success [29]. For instance, gut bacteria are essential for the development of D. valens by regulating glucose transport [30]. Consequently, these mutualistic microbes are expected to be potential targets for beetle control. Furthermore, the early detection of a D. valens infestation using unmanned aerial vehicle (UAV)-based thermal and hyperspectral imaging is conducive to the formulation of effective biological control strategies for this beetle [31]. Some emerging technologies, such as RNA interference (RNAi), have already been applied in pest management [32]. Beetles are generally highly sensitive to RNAi, and feeding and injection of double-stranded RNA (dsRNA) can kill bark beetles [33]. For example, feeding Dendroctonus frontalis dsRNA targeting heat shock protein genes resulted in up to 100% mortality [34]. However, highly heterogeneous forest ecosystems limit the feasibility of delivering dsRNAs to pests. Microbial-mediated dsRNA delivery is increasingly used for pest management [35]. This means that there are prospects for using genetic engineering techniques to modify fungal associates to produce and deliver dsRNA to their vector, D. valens. The discovery of a large number of ophiostomatalean fungi associated with D. valens has led to the accumulation of resources helpful in realizing this vision.
5. Conclusions
We isolated and identified three ophiostomatalean fungal associates of D. valens from P. tabuliformis, including the new species, C. jianpingensis. Leptographium terebrantis and O. gilletteae, which are associates of D. valens in North America, were found in China for the first time. This study provides new understanding of the ophiostomatalean assemblage of D. valens, and enriches the resources for the development of fungal associate-mediated bark beetle management strategies.
Author Contributions
Conceptualization, Z.W., H.W. and Q.L.; methodology, Z.W.; software, Z.W.; validation, Z.W., H.W. and Q.L.; formal analysis, Z.W. and S.Y.; investigation, Z.W. and L.L.; resources, Z.W. and Q.L.; data curation, Z.W.; writing—original draft preparation, Z.W.; writing—review and editing, Z.W.; visualization, Z.W. and Q.L.; supervision, Z.W., H.W. and Q.L.; project administration, Z.W. and Q.L.; funding acquisition, Z.W. and Q.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key R&D Program of China (Project No.: 2023YFC2604800).
Data Availability Statement
The data presented in this study are openly available in [GenBank].
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sun, J.H.; Lu, M.; Gillette, N.E.; Wingfield, M.J. Red turpentine beetle: Innocuous native becomes invasive tree killer in China. Annu. Rev. Entomol. 2013, 58, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Zhang, S.; Du, R.; Wang, H. Occurrence status of main forestry invasive species in China and their research trends. Plant Prot. 2022, 48, 21–38. (In Chinese) [Google Scholar]
- Lu, M.; Hulcr, J.; Sun, J. The role of symbiotic microbes in insect invasions. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 487–505. [Google Scholar] [CrossRef]
- De Beer, Z.W.; Procter, M.; Wingfield, M.J.; Marincowitz, S.; Duong, T.A. Generic boundaries in the Ophiostomatales reconsidered and revised. Stud. Mycol. 2022, 101, 57–120. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, K.; Wingfield, M.J. Leptographium Species: Tree Pathogens, Insect Associates and Agents of Bluestain; APS Press: St Paul, MN, USA, 2001. [Google Scholar]
- Eckhardt, L.G. Blackstain root disease and other Leptographium diseases. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CAB International: Wallingford, UK, 2013; pp. 283–297. [Google Scholar]
- Lu, M.; Wingfield, M.J.; Gillette, N.E.; Mori, S.R.; Sun, J.H. Complex interactions among host pines and fungi vectored by an invasive bark beetle. New Phytol. 2010, 187, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Wingfield, M.J.; Gillette, N.; Sun, J.H. Do novel genotypes drive the success of an invasive bark beetle–fungus complex? Implications for potential reinvasion. Ecology 2011, 92, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Marincowitz, S.; Duong, T.A.; Taerum, S.J.; De Beer, Z.W.; Wingfield, M.J. Fungal associates of an invasive pine-infesting bark beetle, Dendroctonus valens, including seven new ophiostomatalean fungi. Persoonia 2020, 45, 177–195. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Decock, C.; Zhang, X.Y.; Maraite, H. Leptographium sinoprocerum sp. nov., an undescribed species associated with Pinus tabuliformis-Dendroctonus valens in northern China. Mycologia 2008, 100, 275–290. [Google Scholar] [CrossRef]
- Lu, Q.; Decock, C.; Zhang, X.Y.; Maraite, H. Ophiostomatoid fungi (Ascomycota) associated with Pinus tabuliformis infested by Dendroctonus valens (Coleoptera) in northern China and an assessment of their pathogenicity on mature trees. Antonie Leeuwenhoek 2009, 96, 275–293. [Google Scholar] [CrossRef]
- Lu, M.; Zhou, X.; De Beer, Z.W.; Wingfield, M.J.; Sun, J.H. Ophiostomatoid fungi associated with the invasive pine-infesting bark beetle, Dendroctonus valens, in China. Fungal Divers. 2009, 38, 133–145. [Google Scholar]
- Taerum, S.J.; Duong, T.A.; De Beer, Z.W.; Gillette, N.; Sun, J.H.; Owen, D.R.; Wingfield, M.J. Large shift in symbiont assemblage in the invasive red turpentine beetle. PLoS ONE 2013, 8, e78126. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Li, D.; Luo, Z.; He, S.; Jiang, S.; Song, Y. Disaster risk analysis of Dendroctonus valens in Northeast China. For. Pest. Dis. 2022, 41, 22–28. (In Chinese) [Google Scholar]
- Huang, F.; Lu, J. The Classification Outline of Scolytidae from China; Tongji University Press: Shanghai, China, 2015; pp. 37–38. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.J.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Jacobs, K.; Bergdahl, D.R.; Wingfield, M.J.; Halik, S.; Seifert, K.A.; Bright, D.E.; Wingfield, B.D. Leptographium wingfieldii introduced into North America and found associated with exotic Tomicus piniperda and native bark beetles. Mycol. Res. 2004, 108, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Stamatakis, A. RaxML Version 8: A tool phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Hsiau, P.T.W.; Harrington, T.C. Ceratocystiopsis brevicomi sp. nov., a mycangial fungus from Dendroctonus brevicomis (Coleoptera: Scolytidae). Mycologia 1997, 89, 661–669. [Google Scholar]
- Taerum, S.J.; Hoareau, T.B.; Duong, T.A.; De Beer, Z.W.; Jankowiak, R.; Wingfield, M.J. Putative origins of the fungus Leptographium procerum. Fungal Biol. 2017, 121, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.H.; Wang, B.; Clarke, S.R.; Sun, J.H. Effect of associated fungi on the immunocompetence of red turpentine beetle larvae, Dendroctonus valens (Coleoptera: Curculionidae: Scolytinae). Insect Sci. 2012, 19, 579–584. [Google Scholar]
- Koski, T.M.; Zhang, B.; Wickham, J.D.; Bushley, K.E.; Blanchette, R.A.; Kang, L.; Sun, J. Chemical interactions under the bark: Bark-, ambrosia-, and wood-boring beetles and their microbial associates. Rev. Environ. Sci. Bio Technol. 2024, 23, 932–948. [Google Scholar]
- Liu, F.; Ye, F.; Yang, Y.; Kang, Z.; Liu, Y.; Chen, W.; Wang, S.; Kou, H.; Kang, L.; Sun, J. Gut bacteria are essential for development of an invasive bark beetle by regulating glucose transport. Proc. Natl. Acad. Sci. USA 2024, 121, e2410889121. [Google Scholar] [CrossRef]
- Bi, P.; Yu, L.; Zhou, Q.; Kuang, J.; Tang, R.; Ren, L.; Luo, Y. Early detection of Dendroctonus valens infestation with UAV-based thermal and hyperspectral images. Remote Sens. 2024, 16, 3840. [Google Scholar] [CrossRef]
- Singewar, K.; Fladung, M. Double-stranded RNA (dsRNA) technology to control forest insect pests and fungal pathogens: Challenges and opportunities. Funct. Integr. Genom. 2023, 23, 185. [Google Scholar]
- Joga, M.R.; Mogilicherla, K.; Smagghe, G.; Roy, A. RNA interference-based forest protection products (FPPs) against wood-boring coleopterans: Hope or hype? Front. Plant Sci. 2021, 12, 733608. [Google Scholar]
- Kyre, B.R.; Rodrigues, T.B.; Rieske, L.K. RNA interference and validation of reference genes for gene expression analyses using qPCR in southern pine beetle, Dendroctonus frontalis. Sci. Rep. 2019, 9, 5640. [Google Scholar]
- Xue, Q.; Avila dos Santos, É.; Smagghe, G.; Zotti, M.J.; Taning, N.T.C. Engineering strategies for insect viruses and microbes for dsRNA production and delivery to insects for targeted gene silencing. Entomol. Gen. 2023, 43, 31–53. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).