

UAV-LiDAR-Based Structural Diversity of Subtropical Forests Under Different Management Practices in Southern China


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Description of the Study Area
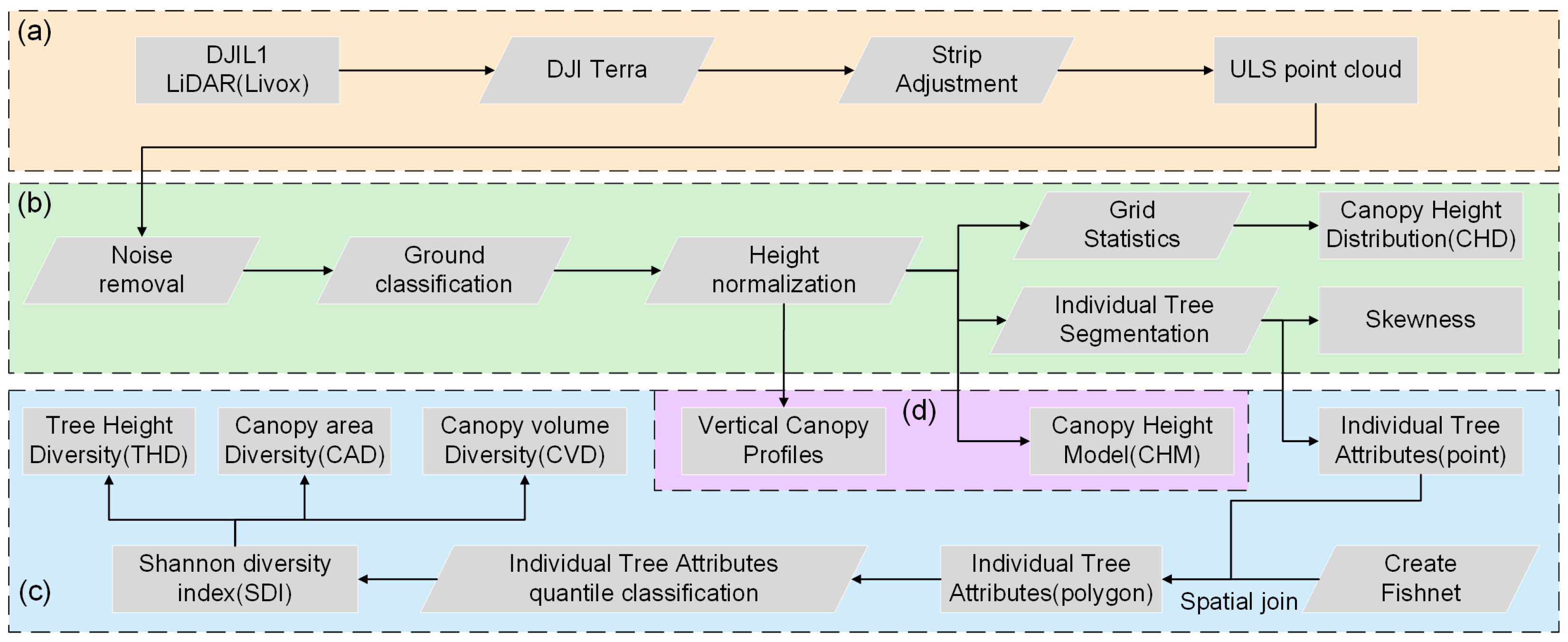
2.2. UAV Laser Scanning
2.3. Field Data
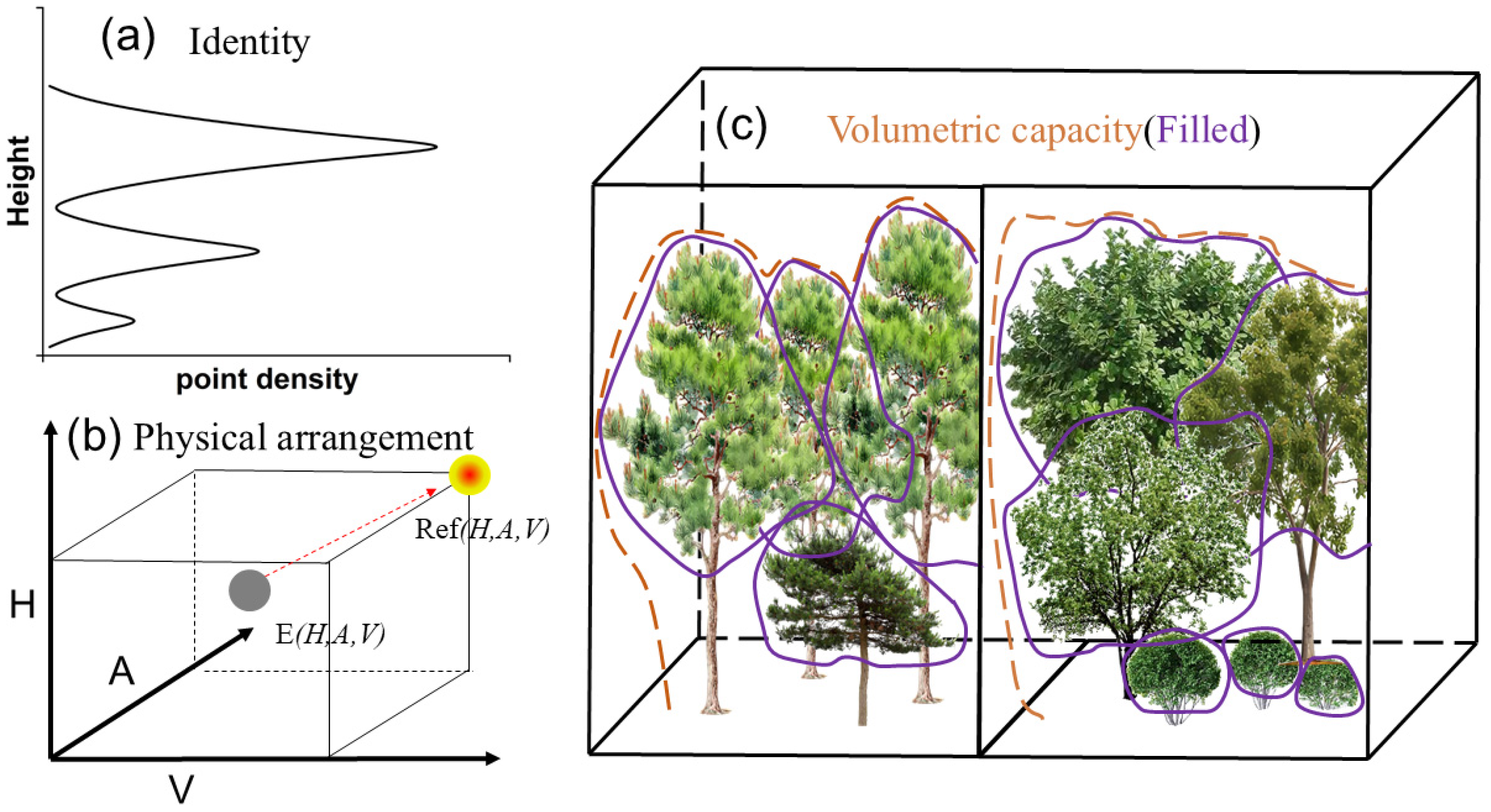
2.4. Measuring Forest Structural Diversity
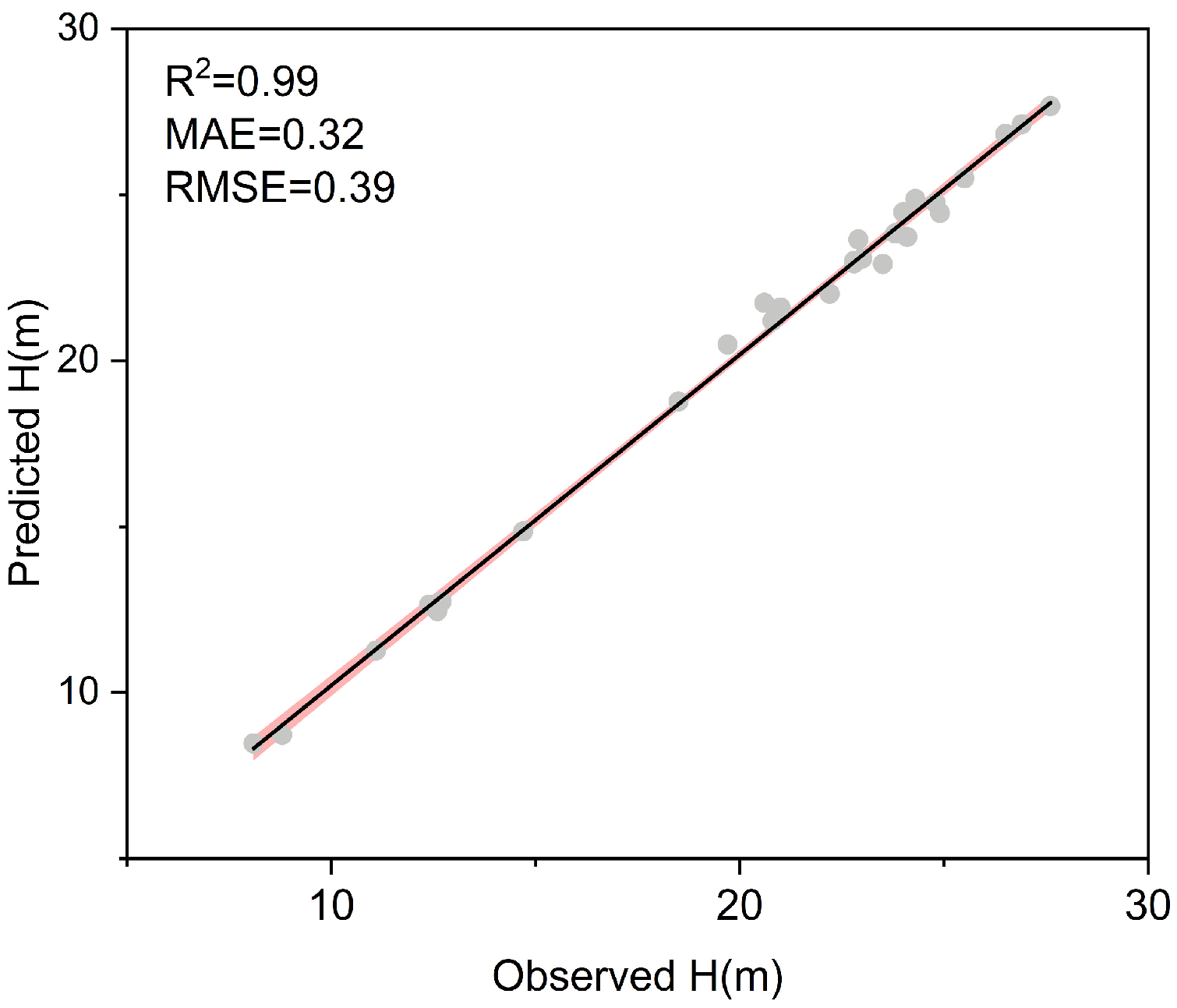
2.5. Data Processing
3. Results
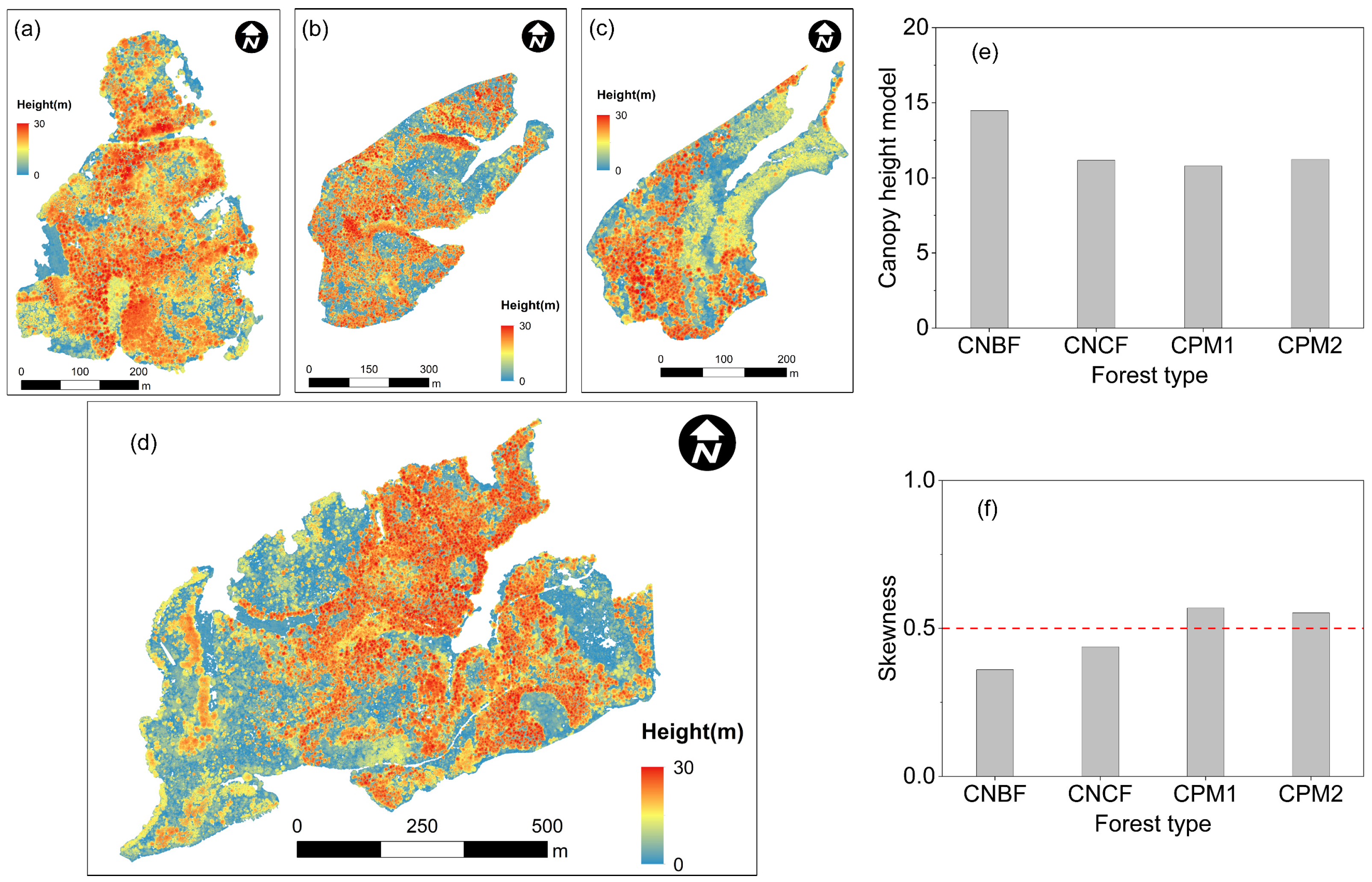
3.1. Forest Volumetric Occupation
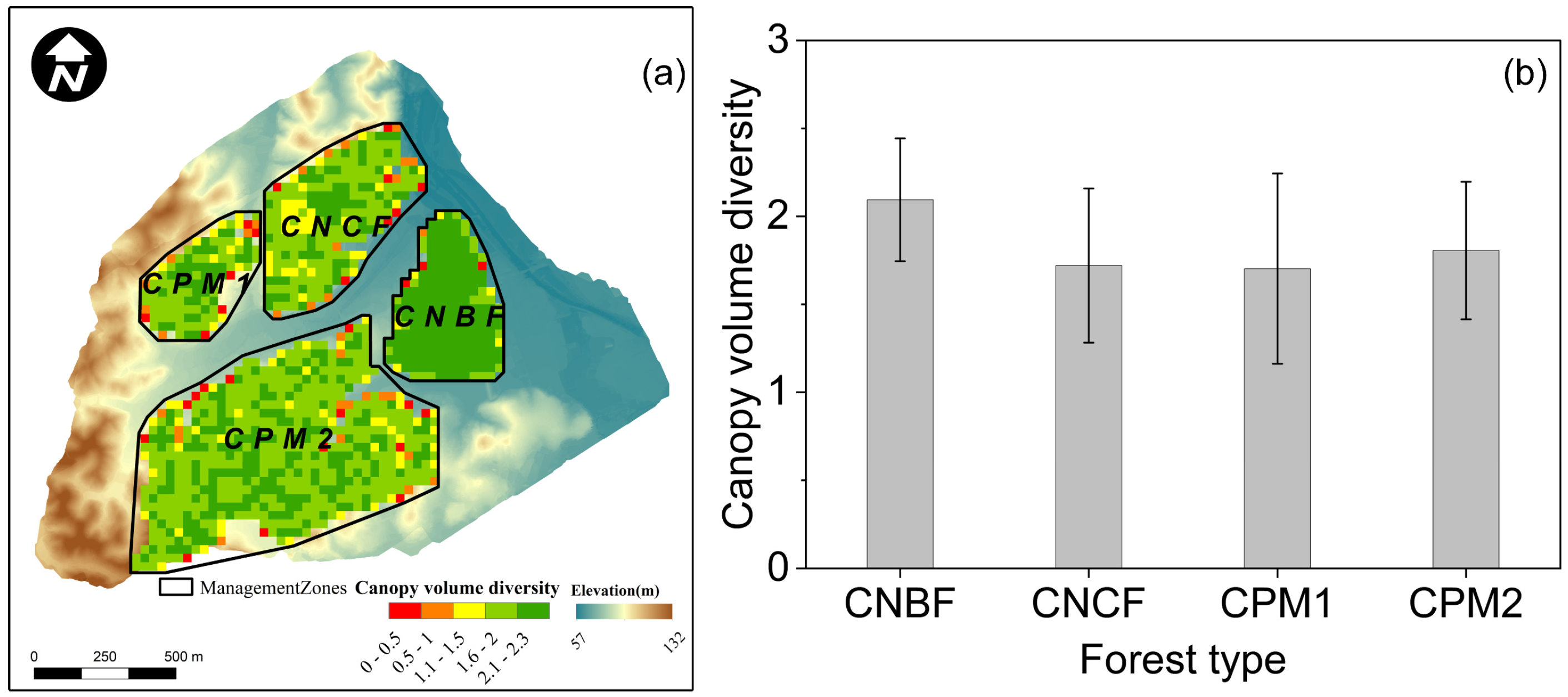
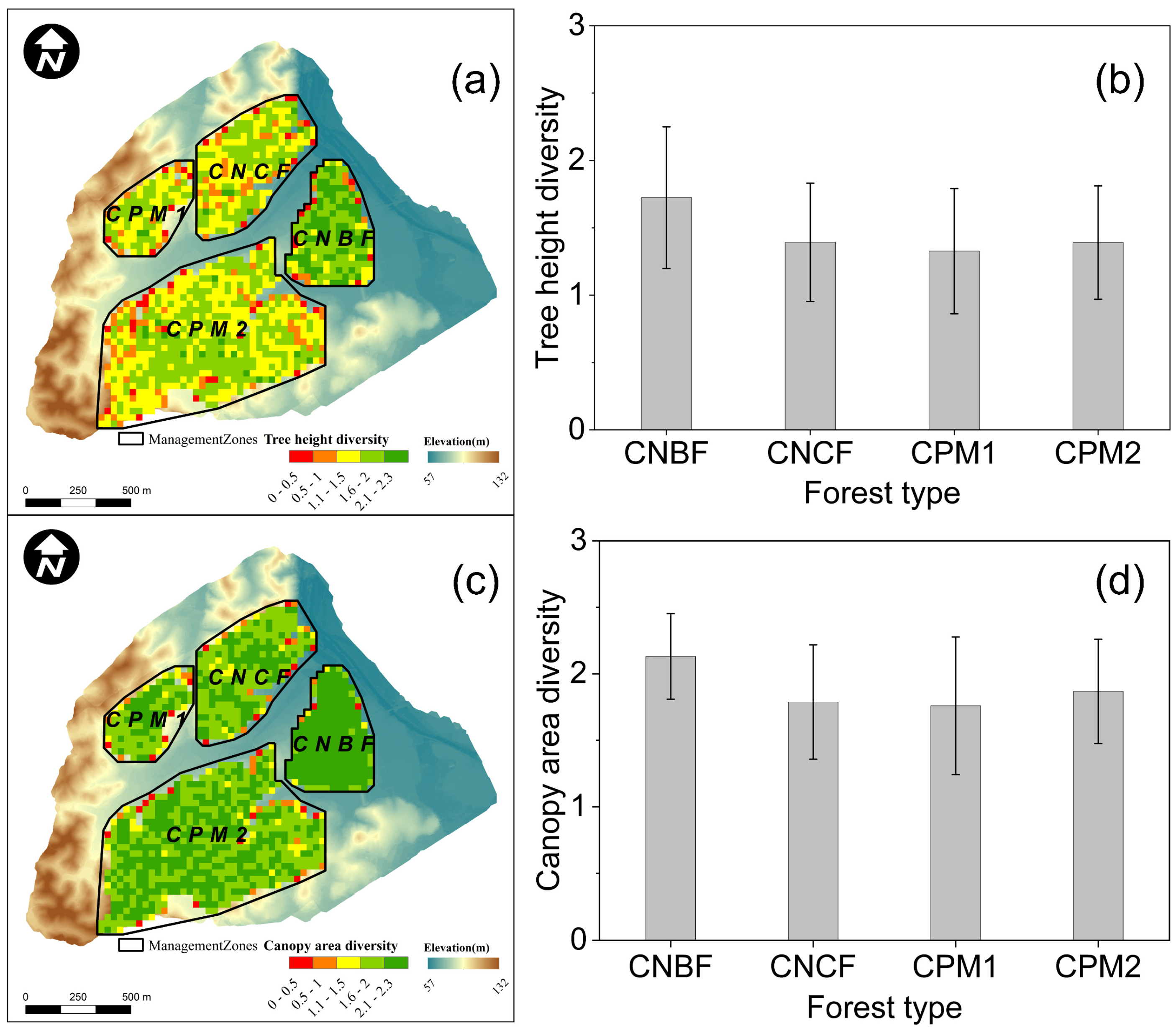
3.2. Forest Structural Diversity
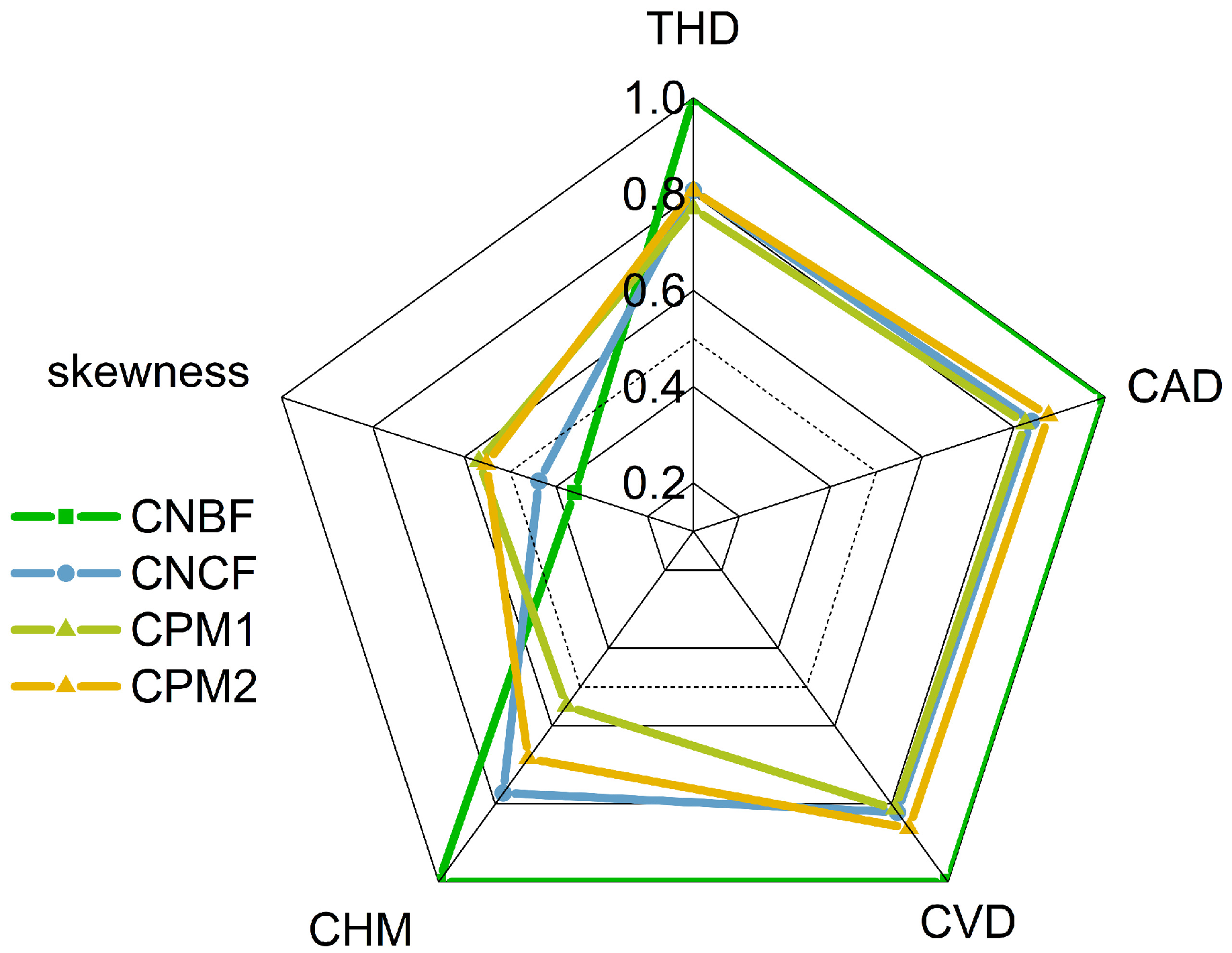
3.3. Synthesis Differences Among Forests
4. Discussion
4.1. Effectiveness of Structural Diversity
4.2. Implications for Future Forest Management
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Storch, F.; Dormann, C.F.; Bauhus, J. Quantifying forest structural diversity based on large-scale inventory data: A new approach to support biodiversity monitoring. For. Ecosyst. 2018, 5, 34. [Google Scholar] [CrossRef]
- Gao, T.; Hedblom, M.; Emilsson, T.; Nielsen, A.B. The role of forest stand structure as biodiversity indicator. For. Ecol Manag 2014, 330, 82–93. [Google Scholar] [CrossRef]
- Storch, F.; Boch, S.; Gossner, M.M.; Feldhaar, H.; Ammer, C.; Schall, P.; Polle, A.; Kroiher, F.; Müller, J.; Bauhus, J. Linking structure and species richness to support forest biodiversity monitoring at large scales. Ann. Sci 2023, 80, 3. [Google Scholar] [CrossRef]
- de Conto, T.; Armston, J.; Dubayah, R. Characterizing the structural complexity of the Earth’s forests with spaceborne lidar. Nat. Commun. 2024, 15, 8116. [Google Scholar] [CrossRef]
- Lang, A.K.; LaRue, E.A.; Kivlin, S.N.; Edwards, J.D.; Phillips, R.P.; Gallion, J.; Kong, N.; Parker, J.D.; McCormick, M.K.; Domke, G.; et al. Forest structural diversity is linked to soil microbial diversity. Ecosphere 2023, 14, e4702. [Google Scholar] [CrossRef]
- Godlee, J.L.; Ryan, C.M.; Bauman, D.; Bowers, S.J.; Carreiras, J.M.B.; Chisingui, A.V.; Cromsigt, J.P.G.M.; Druce, D.J.; Finckh, M.; Gonçalves, F.M.; et al. Structural diversity and tree density drives variation in the biodiversity–ecosystem function relationship of woodlands and savannas. New Phytol. 2021, 232, 579–594. [Google Scholar] [CrossRef]
- LaRue, E.A.; Hardiman, B.S.; Elliott, J.M.; Fei, S. Structural diversity as a predictor of ecosystem function. Environ. Res. Lett. 2019, 14, 114011. [Google Scholar] [CrossRef]
- Atkins, J.W.; Bhatt, P.; Carrasco, L.; Francis, E.; Garabedian, J.E.; Hakkenberg, C.R.; Hardiman, B.S.; Jung, J.; Koirala, A.; LaRue, E.A.; et al. Integrating forest structural diversity measurement into ecological research. Ecosphere 2023, 14, e4633. [Google Scholar] [CrossRef]
- Zhai, L.; Will, R.E.; Zhang, B. Structural diversity is better associated with forest productivity than species or functional diversity. Ecology 2024, 105, e4269. [Google Scholar] [CrossRef]
- LaRue, E.A.; Fahey, R.T.; Alveshere, B.C.; Atkins, J.W.; Bhatt, P.; Buma, B.; Chen, A.; Cousins, S.; Elliott, J.M.; Elmore, A.J.; et al. A theoretical framework for the ecological role of three-dimensional structural diversity. Front. Ecol. Environ. 2023, 21, 4–13. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S.; Lindgren, P.M.F. Stand structure and small mammals in young lodgepole pine forest: 10-year results after thinning. Ecol. Appl. 2001, 11, 1151–1173. [Google Scholar] [CrossRef]
- Tyrrell, L.E.; Crow, T.R. Structural characteristics of old-growth hemlock-hardwood forests in relation to age. Ecology 1994, 75, 370–386. [Google Scholar] [CrossRef]
- Fei, S.; LaRue, E.A.; Hardiman, B.S.; Dahlin, K.M. Structural diversity: A digital revolution. Front. Ecol. Environ. 2023, 21, 3. [Google Scholar] [CrossRef]
- Hovi, A.; Korhonen, L.; Vauhkonen, J.; Korpela, I. LiDAR waveform features for tree species classification and their sensitivity to tree- and acquisition related parameters. Remote Sens. Environ. 2016, 173, 224–237. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Q.; Wan, B.; Qin, H.; Wang, D.; Xu, K.; Song, S.; Sun, Q.; Zhao, X.; Yang, M.; et al. Current status and prospect of three-dimensional dynamic monitoring of natural resources based on LiDAR. Natl. Remote Sens. Bull. 2021, 25, 381–402. [Google Scholar] [CrossRef]
- Du, L.; Pang, Y.; Wang, Q.; Huang, C.; Bai, Y.; Chen, D.; Lu, W.; Kong, D. A LiDAR biomass index-based approach for tree- and plot-level biomass mapping over forest farms using 3D point clouds. Remote Sens. Environ. 2023, 290, 113543. [Google Scholar] [CrossRef]
- Fahey, R.T.; Atkins, J.W.; Gough, C.M.; Hardiman, B.S.; Nave, L.E.; Tallant, J.M.; Nadehoffer, K.J.; Vogel, C.; Scheuermann, C.M.; Stuart-Haëntjens, E.; et al. Defining a spectrum of integrative trait-based vegetation canopy structural types. Ecol. Lett. 2019, 22, 2049–2059. [Google Scholar] [CrossRef]
- Liu, S.; Yang, Y.; Wang, H. Development strategy and management countermeasures of planted forests in China: Transforming from timber-centered single objective management towards multi-purpose management for enhancing quality and benefits of ecosystem services. Acta Ecol. Sin. 2018, 38, 1–10. [Google Scholar]
- Lu, Y.; Luan, S.; Zhang, S.; der, H.B.v.; Lei, X.; Bao, Y. From normal forest to close-to-nature forest: Multi-functional forestry and its practice at national, regional and forest management unit levels in Germany. World For. Res. 2010, 23, 1–11. [Google Scholar]
- Li, T.; Lu, Y.; Jiang, J. Assessment of forest management model of pinus massoniana plantation. J. Northwest For. Univ. 2015, 30, 164–171. [Google Scholar]
- Franklin, J.F. Ecological Science: A Conceptual Basis for FEMAT. J. For. 1994, 92, 21–23. [Google Scholar] [CrossRef]
- Larsen, J.B.; Angelstam, P.; Bauhus, J.; Carvalho, J.F.; Diaci, J.; Dobrowolska, D.; Gazda, A.; Gustafsson, L.; Krumm, F.; Knoke, T.; et al. Closer-to-Nature Forest Management: From Science to Policy 12; European Forest Institute: Joensuu, Finland, 2022. [Google Scholar] [CrossRef]
- Schütz, J.-P.; Saniga, M.; Diaci, J.; Vrška, T. Comparing close-to-naturesilviculture with processes in pristine forests: Lessons from Central Europe. Ann. Sci. 2016, 73, 911–921. [Google Scholar] [CrossRef]
- Li, S.; Yu, G.; Yu, X.; He, H.; Guo, X. A Brief Introduction to Chinese Ecosystem Research Network (CERN). J. Resour. Ecol. 2015, 6, 192–196. [Google Scholar]
- Hao, Z.; Gao, Y.; Zhang, J.Z.; Xu, Y.J.; Yu, G.R. Characteristics of atmospheric nitrogen wet deposition and associated impact on n transport in the watershed of red soil area in southern China. Environ. Sci. 2015, 36, 1630–1638. [Google Scholar] [CrossRef]
- Jiang, P.; Wang, H.; Fu, X.; Dai, X.; Kou, L.; Wang, J. Elaborate differences between trees and understory plants in the deployment of fine roots. Plant Soil 2018, 431, 433–447. [Google Scholar] [CrossRef]
- Hu, Q.; Hu, L.; Li, X. Plant diversity in qianyanzhou after 20 years of small watershed treatment. Acta Phytoecol. Sin. 2005, 29, 766–774. [Google Scholar]
- Listopad, C.M.C.S.; Masters, R.E.; Drake, J.; Weishampel, J.; Branquinho, C. Structural diversity indices based on airborne LiDAR as ecological indicators for managing highly dynamic landscapes. Ecol. Indic. 2015, 57, 268–279. [Google Scholar] [CrossRef]
- Walz, U. Indicators to monitor the structural diversity of landscapes. Ecol. Model. 2015, 295, 88–106. [Google Scholar] [CrossRef]
- Juchheim, J.; Ammer, C.; Schall, P.; Seidel, D. Canopy space filling rather than conventional measures of structural diversity explains productivity of beech stands. For. Ecol. Manag. 2017, 395, 19–26. [Google Scholar] [CrossRef]
- Zhao, K.; Popescu, S.; Nelson, R. Lidar remote sensing of forest biomass: A scale-invariant estimation approach using airborne lasers. Remote Sens. Environ. 2009, 113, 182–196. [Google Scholar] [CrossRef]
- Weishampel, J.F.; Drake, J.B.; Cooper, A.; Blair, J.B.; Hofton, M. Forest canopy recovery from the 1938 hurricane and subsequent salvage damage measured with airborne LiDAR. Remote Sens. Environ. 2007, 109, 142–153. [Google Scholar] [CrossRef]
- Panagiotidis, D.; Abdollahnejad, A.; Slavík, M. 3D point cloud fusion from UAV and TLS to assess temperate managed forest structures. Int. J. Appl. Earth Obs. Geoinf. 2022, 112, 102917. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Chapter 4—Multidimensional quantitative data. In Developments in Environmental Modelling; Legendre, P., Legendre, L., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 24, pp. 143–194. [Google Scholar]
- Staudhammer, C.L.; LeMay, V.M. Introduction and evaluation of possible indices of stand structural diversity. Can. J. For. Res. 2001, 31, 1105–1115. [Google Scholar] [CrossRef]
- LaRue, E.A.; Knott, J.A.; Domke, G.M.; Chen, H.Y.H.; Guo, Q.; Hisano, M.; Oswalt, C.; Oswalt, S.; Kong, N.; Potter, K.M.; et al. Structural diversity as a reliable and novel predictor for ecosystem productivity. Front. Ecol. Environ. 2023, 21, 33–39. [Google Scholar] [CrossRef]
- Balta, H.; Velagic, J.; Bosschaerts, W.; De Cubber, G.; Siciliano, B. Fast statistical outlier removal based method for large 3D point clouds of outdoor environments. IFAC-PapersOnLine 2018, 51, 348–353. [Google Scholar] [CrossRef]
- Zhao, X.; Guo, Q.; Su, Y.; Xue, B. Improved progressive TIN densification filtering algorithm for airborne LiDAR data in forested areas. ISPRS J. Photogramm. Remote Sens. 2016, 117, 79–91. [Google Scholar] [CrossRef]
- Li, W.; Guo, Q.; Jakubowski, M.K.; Kelly, M. A new method for segmenting individual trees from the lidar point cloud. Photogramm. Eng. Remote Sens. 2012, 78, 75–84. [Google Scholar] [CrossRef]
- Atkins, J.W.; Bohrer, G.; Fahey, R.T.; Hardiman, B.S.; Morin, T.H.; Stovall, A.E.L.; Zimmerman, N.; Gough, C.M. Quantifying vegetation and canopy structural complexity from terrestrial LiDAR data using the FORESTR R package. Methods Ecol. Evol. 2018, 9, 2057–2066. [Google Scholar] [CrossRef]
- Ali, A. Forest stand structure and functioning: Current knowledge and future challenges. Ecol. Indic. 2019, 98, 665–677. [Google Scholar] [CrossRef]
- Dănescu, A.; Albrecht, A.T.; Bauhus, J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. Oecologia 2016, 182, 319–333. [Google Scholar] [CrossRef]
- Migliavacca, M.; Musavi, T.; Mahecha, M.D.; Nelson, J.A.; Knauer, J.; Baldocchi, D.D.; Perez-Priego, O.; Christiansen, R.; Peters, J.; Anderson, K.; et al. The three major axes of terrestrial ecosystem function. Nature 2021, 598, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.J.; Paquette, A.; Cavender-Bares, J.; Messier, C.; Reich, P.B. Spatial complementarity in tree crowns explains overyielding in species mixtures. Nat. Ecol. Evol. 2017, 1, 63. [Google Scholar] [CrossRef] [PubMed]
- Forrester, D.I.; Bauhus, J. A review of processes behind diversity—productivity relationships in forests. Curr. For. Rep. 2016, 2, 45–61. [Google Scholar] [CrossRef]
- Bazzaz, F.A. Plant species diversity in old-field successional ecosystems in Southern Illinois. Ecology 1975, 56, 485–488. [Google Scholar] [CrossRef]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin Iii, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Camarretta, N.; Harrison, P.A.; Bailey, T.; Potts, B.; Lucieer, A.; Davidson, N.; Hunt, M. Monitoring forest structure to guide adaptive management of forest restoration: A review of remote sensing approaches. New For. 2020, 51, 573–596. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, S.G.; Lei, X.D.; Ning, J.; Wang, Y. Theoretical basis and implementation techniques on close-to-nature transformation of plantations. World For. Res. 2009, 22, 20–27. [Google Scholar]
- Ma, Z.; Wang, H.; Yang, F.; Fu, X.; Fang, H.; Wang, J.; Dai, X.; Kou, X.; Kou, L.; Zhao, B. Ecological restoration and sustainable development of forest ecosystem in subtropical red soil hilly region based on long-term observation and research. Bull. Chin. Acad. Sci. 2020, 35, 1525–1536. [Google Scholar]
- Brunet-Navarro, P.; Sterck, F.J.; Vayreda, J.; Martinez-Vilalta, J.; Mohren, G.M.J. Self-thinning in four pine species: An evaluation of potential climate impacts. Ann. For. Sci. 2016, 73, 1025–1034. [Google Scholar] [CrossRef]
- Oliver, C.; Larson, B. Forest Stand Dynamics; Wiley: New York, NY, USA, 1996. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, X.; Liu, Y. UAV-LiDAR-Based Structural Diversity of Subtropical Forests Under Different Management Practices in Southern China. Forests 2025, 16, 723. https://doi.org/10.3390/f16050723
Hao X, Liu Y. UAV-LiDAR-Based Structural Diversity of Subtropical Forests Under Different Management Practices in Southern China. Forests. 2025; 16(5):723. https://doi.org/10.3390/f16050723
Chicago/Turabian StyleHao, Xiaobo, and Yu Liu. 2025. "UAV-LiDAR-Based Structural Diversity of Subtropical Forests Under Different Management Practices in Southern China" Forests 16, no. 5: 723. https://doi.org/10.3390/f16050723
APA StyleHao, X., & Liu, Y. (2025). UAV-LiDAR-Based Structural Diversity of Subtropical Forests Under Different Management Practices in Southern China. Forests, 16(5), 723. https://doi.org/10.3390/f16050723