Abstract
In 2020, 264 samples were collected from potato fields in the Turkish provinces of Bolu, Afyon, Kayseri and Niğde. RT-PCR tests, with primers which amplified its coat protein (CP), detected potato virus S (PVS) in 35 samples. Complete CP sequences were obtained from 14 samples. Phylogenetic analysis using non-recombinant sequences of (i) the 14 CP’s, another 8 from Tokat province and 73 others from GenBank; and (ii) 130 complete ORF, RdRp and TGB sequences from GenBank, found that they fitted within phylogroups, PVSI, PVSII or PVSIII. All Turkish CP sequences were in PVSI, clustering within five subclades. Subclades 1 and 4 were in three to four provinces, whereas 2, 3 and 5 were in one province each. All four genome regions were under strong negative selection constraints (ω = 0.0603–0.1825). Considerable genetic variation existed amongst PVSI and PVSII isolates. Three neutrality test methods showed PVSIII remained balanced whilst PVSI and PVSII underwent population expansion. The high fixation index values assigned to all PVSI, PVSII and PVSIII comparisons supported subdivision into three phylogroups. As it spreads more readily by aphid and contact transmission, and may elicit more severe symptoms in potato, PVSII spread constitutes a biosecurity threat for countries still free from it.
1. Introduction
Potato virus S (PVS, genus Carlavirus) is one of the most prevalent viruses infecting potato (Solanum tubersosum) crops globally. It was reported first in 1952 in Europe [1] and subsequently in all major potato production areas across the world [2,3,4,5,6,7]. Following potato virus Y (PVY, genus Potyvirus), potato leafroll virus (PLRV, genus Polerovirus) and potato virus X (PVX; genus Potexvirus), PVS is considered the fourth most important viral pathogen affecting potato production worldwide [3,4,7]. It generally causes mild symptoms or asymptomatic infection in potato foliage and only minor yield losses. However, when severe PVS strains are present, tuber yield losses can reach 20% and tuber size is diminished. Moreover, PVS commonly occurs in mixed infections with other common potato viruses, such as PVY, PLRV, PVX and potato virus A (PVA, genus Potyvirus). This co-infection can increase PVS virion content within infected plants thereby intensifying symptom expression and causing greater tuber yield losses than that occurring with individual infections [3,4,7,8,9,10,11]. As PVS is transmitted easily by foliage contact, and often causes subtle symptoms or asymptomatic infection when present on its own, managing it in healthy seed potato multiplication schemes is problematic. This is especially so in potato seed schemes that rely on visual crop inspections for symptoms without complementing such inspections with regular virus tests on randomly collected leaf samples. This means that PVS is often the first virus to invade healthy seed potato stock schemes. Thorough sample testing in the beginning phases of clonal selection or healthy min-ituber multiplication combined with phytosanitary control measures that minimize contact transmission are required to detect and remove this virus contamination [3,4,7,8,10].
As with other members of genus Carlavirus, PVS has filamentous particles containing a single-stranded, positive-sense RNA genome approximately 8.5 kb in length. This genome is composed of 34 kDa coat protein (CP) and contains a 5′ cap structure, an open reading frame (ORF) encoding an RNA-dependent RNA-polymerase (RdRp), the triple gene-block proteins (TGBp1-3) involved in virus cell-to-cell movement, and a NABP (cysteine-rich nucleic-acid-binding protein) [12]. Two PVS strains were recognized in early biological studies, PVSO (ordinary) and PVSA (Andean). These strains were distinguished depending on whether they remained in inoculated leaves of Chenopdium quinoa (PVSO; O = Ordinary) or invaded this host systemically (PVSA; A = Andean) [13,14,15,16].
Complications arose later, however, when PVS isolates were sequenced, and this same nomenclature (PVSO and PVSA) was retained to distinguish the two major phylogroups found following phylogenetic analysis. This problem arose because biologically defined PVSA isolates also occurred within major phylogroup PVSO [17] and biologically defined PVSO strain isolates occurred within major phylogroup PVSA [6,18]. The term PVSCS (CS = Chenopodium systemic) was suggested to distinguish PVSA isolates from other isolates within major phylogroup PVSO [17]. Subsequently, Cox and Jones [18] suggested the terms PVSO-CS (CS = Chenopodium systemic) for major phylogroup PVSO isolates which invaded this host systemically, and PVSA-CL (CL = Chenopodium localized) for major phylogroup PVSA isolates that remained localized within inoculated leaves. Moreover, when several Andean PVS isolates from S. phureja in Colombia were sequenced, an additional major phylogroup was found [19,20,21]. This was named PVSRVC [5]. In the same year, Santillan et al. [6] reported findings on two further Colombian isolates from S. phureja and an Ecuadorian isolate from S. tubersosum ssp. Andigena that were within this same phylogroup, referring to it as a ‘second South American lineage’ rather than as PVSRVC. Ten additional PVS isolates from five countries in the potato’s Andean domestication center (Bolivia, Chile, Colombia, Ecuador, and Peru) all fitted within the predominantly Andean phylogroup, previously called PVSA, which they referred to as the ‘first South American lineage’. However, they retained the names PVSO and PVSA for the two strains defined biologically based on their respective inabilities or abilities to invade C. quinoa systemically. Santillan et al. [6] adopted their geographically based approach towards phylogroup nomenclature to avoid the confusion arising from using the same names (PVSO and PVSA) for isolate groupings defined in two entirely different ways despite the lack of any relationship between them. To overcome the confusion caused by using names derived from biological properties or geography for plant virus phylogenetic groupings, Jones and Kehoe [22] had suggested replacing all the previous names of strains defined by biological or geographic differences within species phylogenetic group nomenclatures with Latinized numerals. Thus, according to this system, PVSO and PVSA would be retained for biologically defined PVS strains, but the names PVSI, PVSII and PVSIII would replace the major phylogroup names PVSO, PVSA and PVSRVC, respectively.
From their PVS phylogeny which included new PVS sequences from potato’s Andean region domestication center, Santillan et al. [6] concluded that their most important deduction was “amongst all three lineages (= PVSI, PVSII and PVSIII) from the root lead to existing populations infecting potato, only one (= PVSI) dominated the recent global adoption of potato as a major food crop”. After the Spanish invasion of Peru in 1532, PVS was introduced to Europe with potatoes during the “Colombian Exchange” period, which started in c.1570. However, the recent rapid diversification of PVSI only commenced after the major introduction of new potato germplasm from the Andean region which occurred following the European potato famine of the 1840s caused by potato blight (Phythopthora infestans). Moreover, Duan et al. [5] concluded from their PVS phylogeny that Europe played a major role in dispersing this virus to other continents via multiple migration pathways. They also concluded that future studies on bigger data sets with broader geographic representation were necessary to obtain a more complete picture of PVS’s evolutionary history.
RNA viruses, such as PVS, have a strong potential to grow and adapt quickly to natural selective forces because of their enormous population size, proneness to quasispecies development, lack of genome proofreading systems, and rapid generation rates allowing the development of considerable genetic variation [23]. When genetic variation results in functional gain, the high frequency of mutation, recombination, and reassortment that occurs in viral genomes encourages generation of novel forms that quickly spread across the viral population [24]. Therefore, to control viral pathogens effectively, it is important to understand virus population structure and assess its diversity [25,26].
Potato is one of Turkey’s most important food crops, being grown in 14,799 ha of land and with a total annual production of 5,200,000 tons [27]. PVS presence in the country was established when potato samples were tested by ELISA using PVS antibodies [28,29,30,31] or reverse transcription-polymerase chain reaction (RT-PCR) with PVS specific primers [32,33,34,35]. Güner and Yorganci [29] found PVS in the potato production areas of Nevşehir and Niğde provinces, and Engür and Topkaya [34] reported it in the Tokat province. However, despite these studies suggesting widespread PVS occurrence, there is insufficient molecular information about Turkish PVS isolates. In this study, after initial surveys in which we collected PVS-infected leaf samples from Turkish potato fields and obtained the CP sequences of new isolates, we conducted a molecular analysis that redefines the phylogeny of the global PVS population. Our aims were (i) to provide a comprehensive overview of the phylogenetics of PVS using complete ORF, RdRp, and CP genes of different isolates to characterize its genetic diversity and evolutionary history, and (ii) to analyze Turkish isolate coat protein (CP) gene sequences to provide information about PVS biosecurity threats in Anatolia and neighboring Middle Eastern countries.
2. Materials and Methods
2.1. Surveys
During the 2020 summer growing season, 264 leaf samples were collected from potato fields in the Turkish provinces of Afyon, Bolu, Kayseri, and Nevşehir. They were stored at 4 °C for 1–2 days before use. Most of these samples were from plants showing virus-like symptoms, but a few were from symptomless plants.
2.2. RT-PCR and Sequencing
Total RNA was extracted from leaf tissue from each sample as described by Astruc et al. [36]. RT-PCR was performed on each sample individually. To create cDNA in the reverse transcription, a mixture containing 2.5 µL total RNA, 4 µL 5× RT-buffer, 0.5 µL dNTPs (25 mM), 0.25 µL hexamer primer (10 μM) and dH2O was incubated at 42 °C for 1 h. For PCR, a 25 μL mixture containing 2.5 μL of cDNA, 0.2 μL of 25 mM dNTPs, 2 μL of 25 mM MgCl2 and 5 μL of 5× green reaction buffer was used. Next, 0.5 μL of 10 μM of each of forward and reverse primers (5′-TGGGGAATCAGTCCGGCTAGTC-3′ and 5′-ACTGGACCTGCGCTTAGGCT-3′) were added to this mixture. To amplify the complete CP region of the PVS genome [37], 1.25-units GoTaq DNA polymerase (Promega, Madison, WI, USA), and sterile ultra-pure water were prepared. Amplification was performed as follows: initial denaturation for 5 min at 94 °C, 35 cycles of denaturation at 92 °C for 45 s, annealing at primer optimized temperatures 62 °C for 45 s, 72 °C for 1 min and final extension at 72 °C for 7 min. PCR products were then electrophoresed in 1.5% agarose gel stained with ethidium bromide. A single band of around 1100 bp was expected from positive samples. The PCR amplicons for 14 selected PVS positive samples were submitted to a commercial company (Macrogen, Singapore) to be sequenced bi-directionally using Sanger technology. Unfortunately, complete genomic sequencing was impossible, because of the limited research funding and facilities in the laboratory where the sequencing was performed.
2.3. Recombination and Phylogenetic Analysis
On 2 November 2022, complete genome sequences of 139 PVS isolates were retrieved from GenBank and aligned using the ClustalW algorithm included in the MEGA X software [38]. Then, their 5′ and 3′ ends were trimmed to create a ‘complete ORF’ alignment. Nine isolate sequences with recombination events detected by at least five of the RDP, MaxChi, GENECONV, BootScan, ChiMaera, 3Seq, and SiScan, and algorithms with Bonferroni-corrected p value of < 0.05 implemented in Recombination Detection Program (RDP) v.5.30 [39] were removed from the alignment. Then, the recombinant-free complete ORF alignment was subdivided sequentially to form alignments of RdRp, TGB and CP genes according to the reading frames of PVS RefSeq NC_007289. The complete CP sequences (885 nts) of the 14 novel isolates from this study, the 8 other Turkish isolates from Engür and Topkaya [34] and 73 other isolates with complete CP sequences from GenBank were added to the CP alignment of 130 sequences to create a 225-isolate dataset. Present in the ORF phylogenetic tree of Santillan et al. [6] but excluded from our analyses were the incomplete PVSIII sequences MF4I8029 (Ecuadorian S. tuberosum spp. andigena isolate with most of its CP gene missing), and JX683388 (Colombian S. phureja isolate with most of its RdRP gene missing). A list of isolates analyzed in our study is available in Supplementary Table S1.
Maximum-likelihood (ML) trees each based on complete ORF, RdRp, TGB or CP genes, were built using the best-fit Tamura–Nei parameter model [40] within MEGA X, with 1000 bootstrap replicates to determine the statistical significance of isolate clusters. The distance matrixes amongst the CP sequences of isolates tested at nucleotide (nt) and amino acid (aa) levels were analyzed using SDTv1.2 [41].
2.4. Population Structure
Estimation of population genetics-related parameters was performed for both complete ORFs and individual coding sequences (RdRp, TGB and CP) using DnaSP v.6.12.03 software [42]. The parameters calculated included number of haplotypes (h), haplotype diversity (Hd), number of variable sites (S), total number of mutations (η), average number of nt differences between sequences (k), nt diversity (per site) (π), and the acting selection pressure (ω = dN/dS). The calculations also included three neutrality tests with a window length of 100 sites and step size of 25 sites, Fu and Li’s D* and F* [43], and Tajima’s D [44]. To give insight into genetic differentiation and gene flow between the major PVS phylogroups, the parameters KS*, KST*, Z*, Snn, and FST (fixation index) metrics [45,46] were also calculated for complete ORFs and each of the coding sequences again using DnaSP v.6.12.03 software. The coefficient FST ranged between 0 (panmixia) and 1 (fully distinct populations) [45]. Therefore, a FST value > 0.33 indicates rare gene flow and expanding genetic separation amongst tested populations [47,48].
3. Results
3.1. PVS Occurrence and Sequences
The RT-PCR tests with PVS-specific primers detected this virus in 13% of the leaf samples collected (Table 1). The samples found infected mostly came from potato plants that exhibited foliage symptoms of mosaic, mottle, leaf curling and/or plant stunting. However, a few infected plants were asymptomatic, e.g., the infected potato plant in Afyon from which isolate SA12-5 was obtained (Figure 1A–E). The more severe virus symptoms most likely came from mixed infections caused by PVS and other common potato viruses. However, knowledge of other potato viruses that might have been present is lacking as the sequenced samples were not tested for the presence of other viruses. The PVS incidences amongst the samples from each province were as follows: Afyon (19%), Bolu (11%), Kayseri (9%), and Nevşehir (11%) (Table 1). Thus, the highest PVS incidence was in Afyon and the lowest in Kayseri. The best preserved PVS positive samples were selected for sequencing. Complete sequences of the CP region of 14 new PVS isolates were obtained (Afyon = 6, Bolu = 1, Kayseri = 3, Nevşehir = 4) and registered under accession nos. OP819684-OP819697 in the NCBI GenBank.

Table 1.
Details of potato virus S (PVS) detections in potato samples collected from four Turkish provinces, isolates sequenced and accession numbers allocated.

Figure 1.
Sampled potato plants infected with potato virus S alone or in combination with other viruses. (A) Mild mosaic in young leaves of the infected plant from which isolate Ka1-7 was obtained; (B) Asymptomatic infection in the plant from which isolate SA12-5 was secured; (C) Leaf curling, mosaic, rugosity and chlorosis plus plant stunting in the plant from which isolate B9 was obtained; (D) Leaf curling, rugosity and mosaic in the plant from which isolate B14 was secured; (E) Leaf curling in young leaves of the infected the plant from which isolate Bo7 was obtained.
3.2. Recombination and Phylogenetic Analyses
Significant recombination events were detected using RDP5 analysis in different regions of the genomes of 9 out of 139 PVS isolates available in GenBank (Table 2). Five of these had been reported previously (AJ863510, LN851189, LN851192, LN851193 and LN851194) [6], but four others were new (KC430335, LC375227, MN689463 and MK096268). These recombinants came from Eastern Europe, East Asia and East Africa, and following were their countries of origin: Ukraine (3), Czech Republic (1), Poland (1), China (2), Japan (1) and Kenya (1). Their parents mainly belonged to phylogroup PVSI. However, MN689463 from Kenya had a minor PVSII parent also from Kenya, and AJ863510 from the Czech Republic had a minor PVSII parent from the Andean country of Bolivia. It had two recombination events, one at each end of its genome and both involving the same Bolivian minor parent. Additionally, LC375227 from Japan had Chinese recombinant sequence MK096268 as its major parent, and LN851194 from Poland had recombinant sequence LN851192 from Ukraine as its minor parent. With the exception of two Ukrainian recombinant sequences (LN851192 and LN851193) which had identical major and minor parents, and breakpoints at nucleotide sequence position 64–2751, all the minor parental nucleotide sequence positions were different. Thus, no recombination hotspots were revealed. None of the 10 recombinants were from Turkey or the potato’s Andean domestication center in South America.
Table 2.
Putative recombination events detected in 9 out of 139 complete potato virus S genomes tested using RDP5 analysis.
The subsequent phylogenetic and population genetic analyses for complete ORF, RdRp, and TGB included only the 130 non-recombinant sequences.
Sixty-three GenBank isolates (three from Tokat and sixty others) were selected to represent PVS phylogroups I, II and III in nt and aa comparative analyses. According to SDT matrix analysis (Figure 2), the 14 new Turkish PVS isolates shared 94–99% nt and 97–100% aa identities with each other, 94–99% aa and 97–99% nt identities with the three Tokat isolates, and 79–99% nt and 93–100% aa identities with 60 other isolates from GenBank. The most genetically divergent new Turkish isolates were B13, B9, NP49, and SA125, which was confirmed by their phylogenetic placement (see next paragraph).
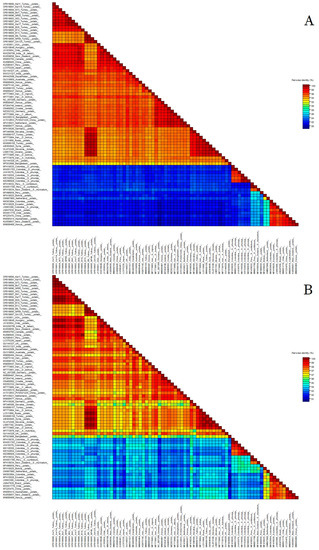
Figure 2.
Percentage identities of the coat protein regions between 14 new Turkish Potato virus S isolates and 63 isolates from the NCBI GenBank. (A) nucleotide; (B) amino acid. Individual isolate names can be seen by increasing the magnification. Additionally, the phylogroup each isolate belonged to is shown in both Figure 3 and Supplementary Table S1.
3.3. Phylogenetic Analysis
In our study, all of the complete ORF, RdRP, TGB and CP trees showed identical topographies in which PVS isolates were clustered into the same three major phylogroups found previously, PVSI, PVSII and PVSIII (Figure 3 and Figure 4). However, during the intervening 4–5 years since the two most recently published PVS phylogenies [5,6], the addition of further new PVS sequences to GenBank has substantially altered the spectrum of geographical origins of the PVS sequences within PVSI and PVSII. Our ORF, RdRp and TGB phylogenetic trees revealed that PVSII now contains the following sequences: 15 Andean (Bolivia, Chile, Colombia, Ecuador, and Peru), one non-Andean South America (Brazil), 10 African (Kenya), two each from Australasia (New Zealand), East Asia (China) and Europe (the Netherlands), and one from Central Asia (Kazakhstan) (Figure 3A–C). Similarly, our PVSII CP phylogenetic tree now contains the following sequences: 19 Andean (Bolivia, Chile, Colombia, Ecuador, Peru), two non-Andean South American (Brazil), 12 East Asian (China), 10 East African (Kenya), two each from Australasia (New Zealand) and Europe (the Netherlands), and one South Asian (India) (Figure 4). In our four different types of phylogenetic trees, PVSII shares a basal node with PVSIII and all six sequences within PVSIII are Andean (Colombia) from S. phureja. All of the Turkish isolate CP sequences fitted within major phylogroup PVSI within our CP tree (Figure 4). In addition, in all four of our phylogenetic trees (especially our CP tree), PVSI now contains sequences from more continents (Figure 3 and Figure 4), primarily the ones from Kenya, Africa (45 sequences), and also ones from other major regions worldwide (South America, North America, Europe, Middle East, Central Asia, South Asia, East Asia, South East Asia, and Australasia). Notably, however, only four of these sequences are Andean (from Chile, Peru, and Ecuador), so PVSI consists of an increasingly ‘rest of the world’ phylogroup.
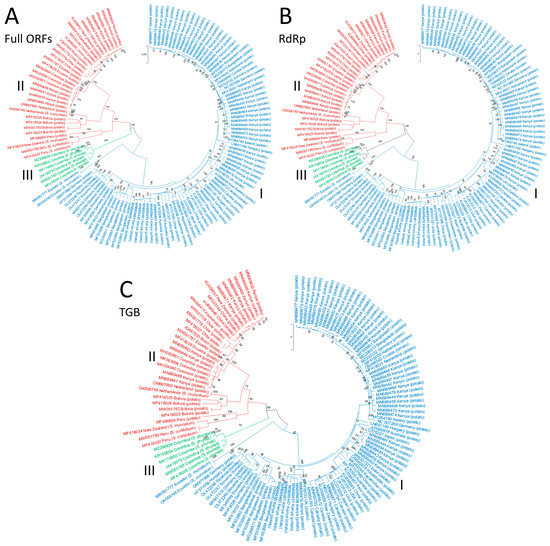
Figure 3.
Phylogenetic trees based on the nucleotide sequences of three regions of potato virus S genome: (A) Complete ORFs; (B) RdRp; and (C) TGB. The trees were generated using the Tamura–Nei parameter model (TN93) in MEGA X software, with uniform rates among sites and 1000 bootstrap replicates (only values > 50% were shown). Different PVS phylogroups are highlighted with colors: PVSI = blue, PVSIII = green, PVSII = red.
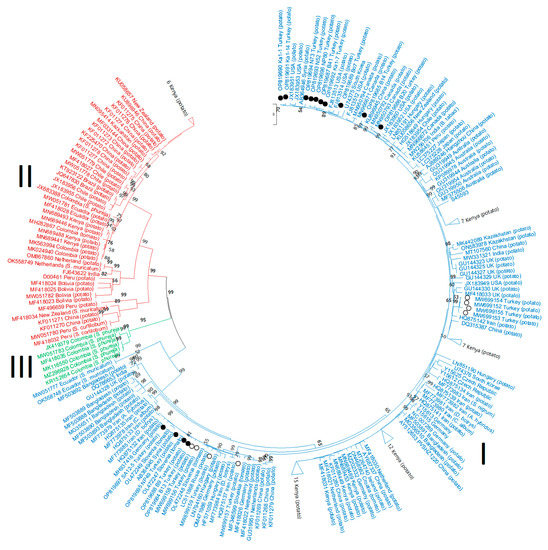
Figure 4.
Phylogenetic trees based on the nucleotide sequences of the coat protein (CP) region of the potato virus S genome. The trees were generated using the Tamura–Nei parameter model (TN93) in MEGA X software, with uniform rates among sites and 1000 bootstrap replicates (only values > 50% were shown). Different PVS phylogroups are highlighted with colors: PVSI = blue, PVSIII = green, PVSII = red. The 14 new Turkish isolates are indicated with black circle symbols and the 8 Turkish isolates from Engür and Topkaya [34] with white circles.
The CP nt sequences from all the 22 Turkish isolates fitted within 5 different PVSI subclades (Figure 4): (1) 8 Turkish isolates (PA3-3, PN14-3, B9, B13, NP49, PN5-2, PN3-6, and SA12-5) grouped closest to 5 isolates from the neighboring countries of Syria and Iran (AB364945, HQ875132, HQ875137, and MF773984-5) and 10 from Netherlands, Slovakia, Russia, Hungary, Germany and Slovenia (GU319951, MF418026, MF418022, MF346599, OL472247-9, LC511868, HF571059, LN794160, MH937416, and OM471986); (2) 4 Turkish isolates (PA3-2, PB5-4, PB6-2, and PB7-1) grouped closest to 1 isolate from the neighboring country of Iran (HQ875142) and 7 from Kenya (MN689446-7, MN689456-7, MN689481, MN689495, and MN689460); (3) B14 was closest to 1 from China (KU896945) and 1 from South Korea (U74375); (4) 8 isolates (B41, N52, NP80, Bo7, N73, Ka1-1, Ka1-7, and Ka1-14) grouped with 1 isolate from the neighboring country of Syria (AB364946) and 4 from the USA (FJ813513-14, JX183951, and JX183953); and (5) B20 was closest to 2 from Canada (MN950792–93). These results are consistent with 5 different PVS introductions to Turkey.
When the 22 Turkish isolates sequences were each mapped to the province they originally came from, Afyon contained the most diverse population with representatives of subclades 1, 3, 4 and 5 all present (Table 1 and Figure 5). Tokat and Nevşehir contained isolates from two subclades each, 1 and 4 for Nevşehir and 1 and 2 for Tokat. Kayseri and Bolu each only contained subclade 4. Thus, subclades 2, 3 and 5 each only occurred in one province whereas subclades 1 and 4 were the most widespread as they were found in three (subclade 1) or four (subclade 4) provinces.
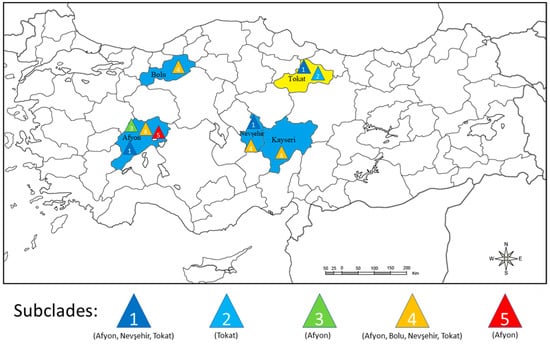
Figure 5.
Distribution according to coat protein sequence of five different potato virus phylogroup PVSI subclades within the five Turkish provinces sampled. Each colored triangle indicates a different PVSI subclade. On the map, blue color identifies the four Turkish provinces where potato leaf samples were collected in this study, and yellow color the Turkish province sampled by Engür and Topkaya [34].
3.4. Genetic Diversity and Selection Pressure Analyses
According to analyses based on their complete ORFs and RdRp, TGB and CP coding regions, major phylogroup PVSII isolate sequences showed high divergence despite being relatively few in number (Table 3). The assigned k (nt differences between sequences) and π (nt diversity) values for the PVSII population were the most obvious parameters as they were consistently higher than those of PVSI and PVSIII across all regions studied. The highest S (variable sites) and η (mutation) values were exhibited by PVSI, closely followed by PVSII. The CP was the most conserved region probably because it is shorter than both of the RdRp and TGB regions. Different regions of the PVS genome were all under strong transcriptional selective pressure as shown by the low dN/dS ratio (ω < 0.2) obtained. Except in the RdRp analysis, PVSII always received smaller ω values than PVSI. Overall, the CP had a lower ω value than other coding regions (Table 3).
Table 3.
Summary of genetic diversity and polymorphism analyses on four genomic regions from different potato virus S populations.
3.5. Neutrality Tests
When three methods were applied in the neutrality tests of complete ORFs and RdRp, only for PVSIII were the values estimated for the TGB and CP coding regions consistently positive (Table 4). Additionally, in the analysis of complete ORFs and RdRp, the Fu and Li’s D* test also gave positive values with PVSII isolates. However, there was no statistically significant support for these differences, indicating there were insufficient data to draw strong conclusions for all populations.
Table 4.
Summary of demography test statistics for four genomic regions from different potato virus S populations.
3.6. Gene Flow and Genetic Separation amongst Populations
In comparisons between PVS isolate sequences belonging to all three major PVS phylogroups, DnaSP analysis estimated high and statistically significant KS, KST, and Z values (genetic differentiation statistics). According to all complete ORFs, RdRp, TGB and CP coding region comparisons, the Snn parameter (genetic differentiation) also always reached its maximum value (1.0000). Furthermore, all FST (coefficient of gene differentiation) values for the three phylogroup comparison were > 0.6 (Table 5). These results provided strong evidence that the division of PVS into three major phylogroups is justified.
Table 5.
Genetic differentiation estimates for the major potato virus S phylogroups based on comparisons between sequences from four different genomic regions.
4. Discussion
4.1. Research Highlights
Here, we provide new insights into the phylogeny of the global PVS population, and the genetic variation present within its local population in the Middle Eastern country of Turkey. We provide further evidence justifying the renaming of its three major phylogroups as PVSI, PVSII and PVSIII rather than continuing with biologically or geographically based nomenclature, and confirm that the smallest phylogroup, PVSIII, shares a basal node with PVSII. PVSIII remains entirely Andean in distribution, PVSII has spread further outside the Andes, especially to East Africa and East Asia, and PVSI now occurs worldwide. Indeed, because of its greater virulence, the likelihood of further PVSII spread is a cause of concern for the biosecurity authorities of counties it has yet to reach. We find substantial genetic variation between the isolates within PVSI and PVSII, but not within PVSIII, and according to our neutrality tests, PVSIII isolates remain balanced whilst PVSI and PVSII exhibit expansion of their respective isolate populations. The high FST values assigned to PVSI, PVSII and PVSIII comparisons is consistent with their separation into three major phylogroups. In addition, the ORF, RdRp, TGB and CP regions of PVS are under strong negative selection constraints. Eight of the major parents and six of the minor parents of the nine recombinant sequences belonged to PVSI, but one recombinant sequence had a recombinant major parent and three of them had minor parents that were either recombinants or belonged to PVSII. Moreover, our study expanded the geographical distribution of recombinant PVS sequences to include East Africa and East Asia in addition to Europe despite not finding any recombinant sequences amongst potato’s Andean domestication center where PVS originated. The twenty-two Turkish CP sequences belonged to five PVSI subclades which is consistent with five separate incursions into the country, three of which likely represent recent arrivals. In our surveys of potato fields within Turkey, the incidence of PVS infection averaged 13% overall, and varied between incidences of 9% and 19% between individual provinces. This shows that PVS occurs commonly, so it is likely to be of economic importance for Turkish potato production.
4.2. Incursion History
When the origins of the 14 new CP sequences within PVSI subclades 1–5 from our study, along with the 8 CP sequences of Engür and Topkaya [34], were mapped to different Turkish provinces, this information provided an insight into the likely history of the 5 PVS incursions into the country. Sequences from subclades 1 and 4 were each found in 3–4 of the 5 provinces, but subclades 2, 3 and 5 were only present in 1 province each (Figure 5). Therefore, the sequences within subclades 1 and 4 might represent 2 earlier PVS introductions that spread to other provinces subsequently, whereas, the sequences within subclades 2, 3 and 5 might represent more recent introductions yet to spread to other provinces. Moreover, since subclade 1, 2 and 4 isolate sequences grouped closely with Syrian and/or Iranian isolates, their introduction to Turkey might have occurred from these neighboring countries, rather than from more distant countries or from other continents. However, there was no evidence of this possible scenario with subclades 3 and 5, as their isolate sequences were closest to ones from countries much further away (Canada, China, and South Korea).
4.3. Recombination Findings
When Lin et al. 2014 [49] used the recombination program RDP4 to examine 44 nt sequences consisting of PVS’s CP and 11K genes (11K encodes an nt-binding protein), they reported detection of 19 potential recombination events by at least three of the seven recombination methods available. Santillan et al. [6] also used RDP4 when they tested 40 complete genomic PVS sequences. However, the level of stringency they applied was much higher as they ignored all anomalies not found by at least five of these same seven recombination methods. They reported presence of recombination events in five PVS genomic sequences, AJ863510, LN851189 and LN851192–4. Our study, which used the more sensitive RDP5 recombination program to examine many more (139) complete genomic PVS sequences, not only confirmed these five sequences as recombinants, but also found four others: one Kenyan (MN689463), one Japanese (LC375227) and two Chinese (KC430335, MK096268) (Table 2). The minor parent of European sequence AJ863510 was from the Bolivian Andes, but otherwise none of the Middle Eastern or Andean region sequences were recombinants themselves or parents of recombinants. Moreover, phylogenetic analysis of different coding regions of 130 recombinant-free PVS sequences generated trees with homogeneous topography (Figure 3), which substantiated their recombinant-free status.
4.4. Phylogenetics
When Santillan et al. [6] prepared phylogenetic trees of the non-recombinant PVS ORFs then available, phylogroup PVSII was predominantly Andean, consisting of 10 new sequences they added from Andean region countries (Bolivia, Chile, Colombia, Ecuador, and Peru), but only 2 from elsewhere, 1 from Australasia (New Zealand) and 1 from East Asia (China). Similarly, after Duan et al. [5] added 10 new sequences from East Asia (China) to a phylogenetic tree of PVS CP sequences they published in the same year (2018), the only others their PVSII contained were 3 Andean (Chile, Colombia) and 2 from non-Andean South America (Brazil). Over the 4–5 years period separating our PVS phylogenetic studies undertaken in 2022 from these two studies published in 2018 [5,6], our phylogenetic trees (ORF, RdRp, TGB, and CP; Figure 3 and Figure 4) revealed considerable differences in the geographical origins of the sequences within PVSI and PVSII. PVSI had become an increasingly ‘rest of world’ phylogroup. Although four Andean region sequences had been added to it by 2022, by that time many others from elsewhere were present from all continents except Antarctica (Africa, Australasia, Europe, North America, Southeast Asia, Middle East, and Central, South and East Asia). Notably, these included 45 sequences from just one African country (Kenya). Within PVSII, the biggest difference between 2018 and 2022 was the expansion of geographical origins of its non-South American sequences. This sequence expansion within PVSII ranged from 11 sequences from East Asia (China) and 1 from New Zealand in 2018, to 12 from East Asia (China), 10 African (Kenya), 2 each from Australasia (New Zealand) and Europe (the Netherlands), and 1 from Central Asia (Kazakhstan) in 2022. Thus, more non-Andean sequences from different world regions were now present, especially from Africa and East Asia, making PVSII global instead of being predominantly Andean. By contrast, PVSIII remained entirely Andean, despite a modest increase in its sequence numbers to six. Although the isolate RVC Andean (JX419379) sequence was first proposed as a distinct PVS lineage having a closer genetic relationship with PVSI than PVSII [21], our analysis of complete ORFs and three coding regions placed this sequence and five other isolate sequences from S. phureja within a separate phylogroup (PVSIII). PVSIII had a closer genetic relationship with PVSII than PVSI and shared a basal node with PVSII (Figure 3 and Figure 4).
As mentioned in the introduction, it makes no sense to retain the names PVSO and PVSA for groups of isolates defined both biologically and phylogenetically that do not coincide with their properties. Thus, there is no logic in using the name PVSA (A = Andean) for PVSII as many isolates outside the Andean region are now present within this phylogroup. Likewise, there is no logic in retaining the name PVSRVC (this name is from isolate RVC) for PVSII, or the name PVSO (O = ordinary) for PVSI which now consists of isolates from all continents, except Antarctica. The likely reason for the expansion in both geographical distribution and numbers of PVSII isolate sequences from outside the Andean region might be the importation of Andean potato germplasm inadvertently infected with PVS by potato breeding programs in other continents. Since the non-Andean countries of Kenya and China have the most new PVSII sequences, this reflects the presence of highly active potato breeding programs in these two countries [50]. Such germplasm importation probably occurred mainly in the past when quarantine testing for viruses in potato germplasm dispatched as tubers were less stringent than present, and when virus sequencing technologies were less advanced and less virus sequencing was performed than present [51]. Although there are far fewer sequences within phylogroup PVSIII and all of these are currently Andean, more sequencing will likely reveal that it too has reached other continents. This might occur particularly via the distribution of germplasm consisting of S. phureja tubers, since currently, with one exception (MF4I8029 from S. tuberosum ssp. andigena referred above in the sections Introduction, and Materials and Methods) [6], this is the only potato species found so far infected with it (Figure 3 and Figure 4). Within PVSI, expansion of the international trade in seed potatoes between non-Andean countries seems the probable reason for the major diversification of global geographical isolate origins within this phylogroup. This is because this export trade consists mostly of seed tuber exports from Europe and North America to the rest of the world, with only a minor Andean region seed potato export contribution (from Chile which exports within South America) [52].
4.5. Population Genetics
When Lin et al. [49] undertook a population genetics study of 69 CP and 44 11K genes of PVS, they reported greater nt diversity (higher π value) with CP than 11K, suggesting greater variation in the CP gene. Additionally, they reported that PVSI sequences had lower π values than PVSII sequences, which was consistent with PVSII having greater diversity. In addition, when they used three neutrality tests (Tajima’s D, Fu and Li’s D*, and Fu and Li’s F*), the values they obtained with both CP and 11K genes were positive for PVSII but negative for PVSI. This suggested a balancing selection pressure on CP and 11K proteins for PVSII but a negative selection pressure on them for PVSI. In our study, which was with many more sequences and which included complete ORF, RdRp and TGB regions in addition to CP sequences, the PVSII population values for nt differences between sequences and nt diversity were consistently higher than those obtained with the PVSI and PVSIII populations across all regions. In addition, there was considerable genetic variation amongst PVSI and PVSII isolate sequences, but low genetic variation within the PVSIII population. When the same three neutrality tests were used, PVSI and PVSII sequences always received negative values, but the PVSIII sequence values were always positive. This differs from the findings of Lin et al. [49] in which positive values were obtained with PVSII sequences. This difference was presumably because they analyzed a much smaller number of PVSII sequences (only 6) than we did (48), and a greater number of ours were from outside the Andean region. Our results indicate that both PVSI and PVSII are experiencing population expansion across all four genomic regions, but the smaller PVSIII population is undergoing balancing selection. Moreover, our FST values from comparisons between PVSIII versus either PVSI or PVSII at different genomic regions were always > 0.6 (Table 4), suggesting lack of gene flow, and expanding genetic separation between PVSIII and the other two phylogroups. This provides additional evidence that PVSIII is distinct. Furthermore, in our study, analysis of the sequences of 225 PVS isolates showed that the CP was the most conserved region, which was probably due to the strong purifying pressure on its short nt sequence. Thus, when sequences from other genomic regions are absent, the CP gene can still be used to represent PVS phylogeny. Other members of genus Carlavirus (potato virus M (PVM) [53] and garlic common latent virus (GCLV) [54]) also experience a strong negative evolutionary constraint on the CP. Thus, data obtained in this study could also contribute to other evolutionary studies of the genus Carlavirus.
4.6. Biosecurity Implications
Studies in Europe and North America reported that not only did more symptoms develop in S. tuberosum spp. tubersosum potato foliage with strain group PVSA, to which most phylogroup PVSII isolates belong rather than to phylogroup PVSI, but also aphid transmission is more efficient with strain group PVSA than with PVSO [16,55,56,57]. Moreover, although symptomatology studies did not confirm this for phylogroup PVSII when Andean S. tuberosum ssp. andigena or S. tuberosum ssp. tuberosum x andigena potato cultivars were used in South America, they did confirm it for PVSII aphid transmission [6]. In addition, the South American studies [6] reported evidence that isolates within phylogroup PVSII are not only more stable in infective sap than those in phylogroup PVSI, but also attain higher virion concentrations within infected plants, both properties likely to favor more efficient aphid and contact transmission in the field. Therefore, PVSII is more likely to constitute a threat to seed and ware potato production for countries to which it is yet to spread, especially when co-infection then occurs with other viruses. Our study demonstrates that plant biosecurity and quarantine organizations in such countries should consider conducting surveillance programs to establish whether PVSII has arrived, and taking additional precautions to help avoid its establishment. This is suggested as potato is the fourth most important staple food crop in the world and plays a major role in addressing food insecurity in developing countries [58,59].
5. Conclusions
We compared (i) all the non-recombinant sequences of complete ORF, RdRp and TGB available from GenBank, and (ii) the new CP’s we obtained from four Turkish provinces, and others obtained previously from a fifth province, with all other CP sequences in GenBank. Our comparisons involved recombination, phylogenetic and population genetic analyses. The more comprehensive sequence data set available now than in 2018, when the last genomic and CP analyses were conducted with this virus, enabled us to make important new deductions that helped build a better understanding of its evolution, the spread of its distinct phylogroups to other continents and the practical significance of this ongoing process. The suggested renaming of PVS’s three major phylogroups as PVSI, PVSII and PVSIII was supported by recent spread of PVSI and PVSII around the world, especially the recent spread of PVSII away from the Andean region of South America, where potato was first domesticated, to East Africa and East Asia. The genetic variation within phylogroups PVSI and PVSII was considerable, and both of their populations were expanding. By contrast, the PVSIII population had low variation and was undergoing balancing selection. The recombination involved found mostly parental sequences belonging to PVSI. As PVSI infection was widespread in potato crops in Turkey’s potato producing regions, it is likely to have economic significance for the country, especially when it occurs in mixed infection with other viruses. The incursion history of the Turkish CP sequences within phylogroup PVSI subclades 1–5 indicated presence of two earlier PVS introductions that spread to other provinces subsequently, and three more recent introductions yet to spread from the province in which they were found first. Further international spread of phylogroup PVSII poses a biosecurity threat to world regions in which it is still absent. Such spread seems most likely to occur through distribution of germplasm for potato breeding purposes or further expansion of the international seed potato tuber trade.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/v15051104/s1, Supplementary Table S1. Potato virus S isolates examined in this study: accession numbers, names, origins, original hosts and phylogroups.
Author Contributions
Conceptualization, Ş.T. and R.A.C.J.; Methodology, Ş.T., A.Ç. and A.I.S.; Formal analysis, A.Ç. and A.I.S.; Investigation, Ş.T. and R.A.C.J.; Resources, Ş.T.; Writing—Original Draft, A.Ç., A.I.S., Ş.T. and R.A.C.J.; Writing—Review and Editing, A.Ç. and R.A.C.J.; Funding acquisition, Ş.T. All authors have read and agreed to the published version of the manuscript.
Funding
Tokat Gaziosmanpasa University provided funding for sample collection, testing and virus sequencing. This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Coat protein sequences of 14 novel Turkish PVS isolates have been made available in GenBank, reference numbers OP819684-OP819697.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wetter, C. AAB/CMI Descriptions of Plant Viruses, No. 60: Potato virus S; Association of Applied Biologists: Warwick, UK, 1971; p. 3. [Google Scholar]
- McKee, R.K. Virus infection in South American potatoes. Potato Res. 1964, 7, 145–151. [Google Scholar] [CrossRef]
- Stevenson, W.R.; Loria, R.; Franc, G.D.; Weingartner, D.P. Compendium of Potato Diseases, 2nd ed.; APS Press: St. Paul, MN, USA, 2001; p. 106. [Google Scholar]
- Jones, R.A.C. Virus disease problems facing potato industries worldwide: Viruses found, climate change implications, rationalizing virus strain nomenclature and addressing the Potato virus Y issue. In The Potato: Botany, Production and Uses; Navarre, R., Pavek, M.J., Eds.; CABI: Wallingford, UK, 2014; pp. 202–224. [Google Scholar]
- Duan, G.; Zhan, F.; Du, Z.; Ho, S.Y.; Gao, F. Europe was a hub for the global spread of potato virus S in the 19th century. Virology 2018, 525, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Santillan, F.W.; Fribourg, C.E.; Adams, I.P.; Gibbs, A.J.; Boonham, N.; Kehoe, M.A.; Maina, S.; Jones, R.A.C. The biology and phylogenetics of Potato virus S isolates from the Andean region of South America. Plant Dis. 2018, 102, 869–885. [Google Scholar] [CrossRef]
- Kreuze, J.F.; Souza-Dias, J.A.C.; Jeevalatha, A.; Figueira, A.R.; Valkonen, J.P.T.; Jones, R.A.C. Viral Diseases in Potato. In The Potato Crop; Campos, H., Ortiz, O., Eds.; Springer Cham: New York, NY, USA, 2020; pp. 389–430. [Google Scholar] [CrossRef]
- De Bokx, J.A.; van der Want, J.P.H. Viruses of Potatoes and Seed-Potato Production, 2nd ed.; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1987; p. 256. [Google Scholar]
- Jeffries, C. FAO/IPGRI Technical Guidelines for the Safe Movement of Germplasm; No. 19. Potato; Plant Genetic Resources Institute, Food and Agriculture Organization of the United Nations Rome/International: Rome, Italy, 1998; p. 177. [Google Scholar]
- Loebenstein, G.; Berger, P.H.; Brunt, A.A.; Lawson, R.H. (Eds.) Virus and Virus-Like Diseases of Potatoes and Production of Seed-Potatoes; Springer: Dordercht, The Netherlands, 2001; p. 460. [Google Scholar]
- Nyalugwe, E.P.; Wilson, C.R.; Coutts, B.A.; Jones, R.A.C. Biological properties of Potato virus X in potato: Effects of mixed infection with Potato virus S and resistance phenotypes in cultivars from three continents. Plant Dis. 2012, 96, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Martelli, G.P.; Adams, M.J.; Kreuze, J.F.; Dolja, V.V. Family Flexiviridae: A case study in virion and genome plasticity. Annu. Rev. Phytopathol. 2007, 45, 73–100. [Google Scholar] [CrossRef]
- Hinostroza-Orihuela, A.M. Some properties of potato virus S isolated from Peruvian potato varieties. Potato Res. 1973, 16, 244–250. [Google Scholar] [CrossRef]
- Jones, R.A.C. The ecology of viruses infecting wild and cultivated potatoes in the Andean Region of South America. In Pests, Pathogens and Vegetation; Thresh, J.M., Ed.; Pitman: London, UK, 1981; pp. 89–107. [Google Scholar]
- Foster, G.D.; Mills, P.R. The 3′-nucleotide sequence of an ordinary strain of potato virus S. Virus Genes 1992, 6, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Slack, S.A. Identification of an isolate of the Andean strain of potato virus S in North America. Plant Dis. 1983, 67, 786–789. [Google Scholar] [CrossRef]
- Matoušek, J.; Schubert, J.; Ptáček, J.; Kozlová, P.; Dědič, P. Complete nucleotide sequence and molecular probing of potato virus S genome. Acta Virol. 2005, 49, 195–205. [Google Scholar]
- Cox, B.A.; Jones, R.A.C. Genetic variability of the coat protein gene of Potato virus S and distinguishing its biologically distinct strains. Arch. Virol. 2010, 155, 1163–1169. [Google Scholar] [CrossRef]
- Gil, J.F.; Cotes, J.M.; Marín, M. Detection and molecular characterization of Potato virus S (PVS, Carlavirus) from Colombia. Rev. Biol. Trop. 2013, 61, 565–575. (In Spanish) [Google Scholar] [CrossRef]
- Gutiérrez, P.A.; Alzate, J.F.; Montoya, M.M. Pirosecuenciación del genoma de una cepa andina de Potato virus S (PVS, Carlavirus) infectando Solanum phureja (Solanaceae) en Colombia. Rev. Fac. Cienc Agrar 2012, 8, 84–93. (In Spanish) [Google Scholar] [CrossRef]
- Gutiérrez, P.A.; Alzate, J.F.; Marín-Montoya, M.A. Complete genome sequence of a novel potato virus S strain infecting Solanum phureja in Colombia. Arch. Virol. 2013, 158, 2205–2208. [Google Scholar] [CrossRef]
- Jones, R.A.C.; Kehoe, M.A. A proposal to rationalize within-species plant virus nomenclature: Benefits and implications of inaction/in silico characterization of coat protein of PVS-bitlis isolate and docking analysis with host protein. Arch. Virol. 2016, 161, 2051–2057. [Google Scholar] [CrossRef]
- Hanssen, I.M.; Thomma, B.P.H.J. Pepino mosaic virus: A successful pathogen that rapidly evolved from emerging to endemic in tomato crops. Mol. Plant Pathol. 2010, 11, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Moya, A.; Holmes, E.C.; González-Candelas, F. The population genetics and evolutionary epidemiology of RNA viruses. Nat. Rev. Microbiol. 2004, 2, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Coşkan, S.; Morca, A.F.; Akbaş, B.; Çelik, A.; Santosa, A.I. Comprehensive surveillance and population study on plum pox virus in Ankara Province of Turkey. J. Plant Dis. Prot. 2022, 129, 981–991. [Google Scholar] [CrossRef]
- Çelik, A.; Coşkan, S.; Morca, A.F.; Santosa, A.I.; Koolivand, D. Insight into population structure and evolutionary analysis of the emerging tomato brown rugose fruit virus. Plants 2022, 11, 3279. [Google Scholar] [CrossRef]
- TÜİK-Turkish Statistical Institute: Crop Production Statistic. 2021. Available online: https://www.tuik.gov.tr/ (accessed on 19 November 2022).
- Bostan, H.; Haliloglu, K. Distribution of PLRV, PVS, PVX and PVY (PVYN, PVYo and PVYc) in the seed potato tubers in Turkey. Pak. J. Biol. Sci. 2004, 7, 1140–1143. [Google Scholar] [CrossRef]
- Güner, Ü.; Yorganci, Ü. Niğde ve Nevşehir İlleri patates ekiliş alanlarında saptanan viral etmenler/Viral species detected in potato cultivation areas of Niğde and Nevşehir Provinces. Bitki Koruma Bülteni 2006, 46, 35–49. (In Turkish) [Google Scholar]
- Güner, Ü. Afyon, Bolu, Nevşehir ve Niğde Illerinde Patateslerdeki virüs Hastalıklarının Tanılanması, Hastalık Oranları ve en Yaygın Üretilen Çeşitlerin bazı Virüslere Karşı Reaksiyonlarının Belirlenmesi/Identification of Virus Diseases of Potato in Afyon, Bolu, Nevşehir, and Niğde Provinces, Disease Rates and Detection of the Most Common Grown Potato Varieties at the Variety Reactions through Some Plant Viruses. Ph.D. Thesis, Department of Plant Protection, Graduate School of Natural and Applied Sciences, Ege University, Izmir, Turkey, 2007. (In Turkish). [Google Scholar]
- Yardımcı, N.; Kılıç, H.C.; Demir, Y. Detection of PVY, PVX, PVS, PVA, and PLRV on different potato varieties in Turkey using DAS-ELISA. J. Agric. Sci. Technol. 2015, 17, 757–764. [Google Scholar]
- Bostan, H.; Guclu, C.; Ozturk, E.; Ozdemir, I.; Ilbagi, H. Influence of aphids on the epidemiology of potato virus diseases (PVY, PVS and PLRV) in the high-altitude areas of Turkey. Pak. J. Biol. Sci. 2006, 9, 759–765. [Google Scholar] [CrossRef]
- Kökten, M. Ödemiş bölgesinde üretimi yapılan patates yumrularında PVY, PVX, PVS ve PLRV’lerinin RT-PCR yöntemiyle saptanması/Determination of PVY, PVX, PVS and PLRV at potato tubers which were produced in Ödemiş region with RT-PCR method. Master’s Thesis, Graduate School of Natural and Applied Sciences, Adnan Menderes University, Aydın, Turkey, 2007. (In Turkish). [Google Scholar]
- Engür, A.M.; Topkaya, S. Prevalence and molecular characterization of important potato viruses in the Tokat province of Turkey. Mol. Biol. Rep. 2023, 50, 2171–2181. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, G.; Mustafa, U.; Demirel, S. Potato virus S (PVS)-Bitlis izolatının kılıf proteinin in silico karakterizasyonu ve konak proteini ile Docking analizi/In Silico characterization of coat protein of PVS-Bitlis isolate and docking analysis with host protein. J. Agric. 2022, 5, 58–69. (In Turkish) [Google Scholar] [CrossRef]
- Astruc, N.; Marcos, J.F.; Macquaire, G.; Candresse, T.; Pallás, V. Studies on the diagnosis of hop stunt viroid in fruit trees: Identification of new hosts and application of a nucleic acid extraction procedure based on non-organic solvents. Eur. J. Plant Pathol. 1996, 102, 837–846. [Google Scholar] [CrossRef]
- Salari, K.; Massumi, H.; Heydarnejad, J.; Pour, A.H.; Varsani, A. Analysis of Iranian Potato virus S isolates. Virus Genes 2011, 43, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Martin, D.P.; Varsani, A.; Roumagnac, P.; Botha, G.; Maslamoney, S.; Schwab, T.; Kelz, Z.; Kumar, V.; Murrell, B. RDP5: A computer program for analyzing recombination in, and removing signals of recombination from, nucleotide sequence datasets. Virus Evol. 2020, 7, veaa087. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Fu, Y.X.; Li, W.H. Statistical tests of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Hudson, R.R.; Boos, D.D.; Kaplan, N.L. A statistical test for detecting population subdivision. Mol. Biol. Evol. 1992, 9, 138–151. [Google Scholar] [PubMed]
- Hudson, R.R. A new statistic for detecting genetic differentiation. Genetics 2000, 155, 2011–2014. [Google Scholar] [CrossRef] [PubMed]
- Çelik, A.; Santosa, A.I.; Gibbs, A.J.; Ertunç, F. Prunus necrotic ringspot virus in Turkey: An immigrant population. Arch. Virol. 2022, 167, 553–562. [Google Scholar] [CrossRef]
- Santosa, A.I.; Randa-Zelyüt, F.; Karanfil, A.; Korkmaz, S.; Hartono, S.; Ertunç, F. Phylogenetic and diversity analyses revealed that leek yellow stripe virus population consists of three types: S, L, and N. Virus Genes 2023, 59, 121–131. [Google Scholar] [CrossRef]
- Lin, Y.H.; Abad, J.A.; Maroon-Lango, C.J.; Perry, K.L.; Pappu, H.R. Molecular characterization of domestic and exotic potato virus S isolates and a global analysis of genomic sequences. Arch. Virol. 2014, 159, 2115–2122. [Google Scholar] [CrossRef]
- Bradshaw, J.E. Potato Breeding: Theory and Practice; Springer: Cham, Switzerland, 2021; p. 563. [Google Scholar] [CrossRef]
- Martin, R.R.; Constable, F.; Tzanetakis, I.E. Quarantine regulations and the impact of modern detection methods. Annu. Rev. Phytopathol. 2016, 54, 189–205. [Google Scholar] [CrossRef]
- Historical Data of Export and Import of Potatoes Seed, Fresh or Chilled. Available online: https://oec.world/en/profile/hs/potatoes-seed-fresh-or-chilled (accessed on 20 December 2022).
- Ge, B.; He, Z.; Zhang, Z.; Wang, H.; Li, S. Genetic variation in potato virus M isolates infecting pepino (Solanum muricatum) in China. Arch. Virol. 2014, 159, 3197–3210. [Google Scholar] [CrossRef]
- Santosa, A.I.; Ertunc, F. Phylogenetic and diversity analyses of Garlic common latent virus based on the TGB and CP gene sequence. Plant Prot. Sci. 2021, 57, 179–187. [Google Scholar] [CrossRef]
- Dolby, C.A.; Jones, R.A.C. Occurrence of the Andean strain of Potato virus S in imported potato material and its effects on potato cultivars. Plant Pathol. 1987, 36, 381–388. [Google Scholar] [CrossRef]
- Rose, D.G. Some properties of an unusual isolate of potato virus S. Potato Res. 1983, 26, 49–62. [Google Scholar] [CrossRef]
- Wardrop, E.A.; Gray, A.B.; Singh, R.P.; Peterson, J.F. Aphid transmission of potato virus S. Amer. Potato J. 1989, 66, 449–459. [Google Scholar] [CrossRef]
- Birch, P.R.; Bryan, G.; Fenton, B.; Gilroy, E.M.; Hein, I.; Jones, J.T.; Prashar, A.; Taylor, M.A.; Torrance, L.; Toth, I.K. Crops that feed the world 8: Potato: Are the trends of increased global production sustainable? Food Secur. 2012, 4, 477–508. [Google Scholar] [CrossRef]
- Scott, G.J. A review of root, tuber and banana crops in developing countries: Past, present and future. Int. J. Food Sci. Technol. 2021, 56, 1093–1114. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).