
Combining Cellular Immunization and Phage Display Screening Results in Novel, FcγRI-Specific Antibodies

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture of Eukaryotic Cells
2.2. Generation of F(ab’)2 from IgG
2.3. Flow Cytometric Analyses with Antibodies and F(ab’)2
2.4. Generation of Murine scFv Antibody Library from Spleen
2.5. Phage Display Using Cellular Panning
2.6. Flow Cytometric Analyses with Polyclonal Phage Antibodies
2.7. Whole Cell ELISA with Monoclonal Phage Antibodies
2.8. Sequencing Analyses
2.9. Generation of Chimeric FcγRI-Specific Antibodies
2.10. Generation, Stimulation, and Labeling of Polymorphonuclear Cells
3. Results
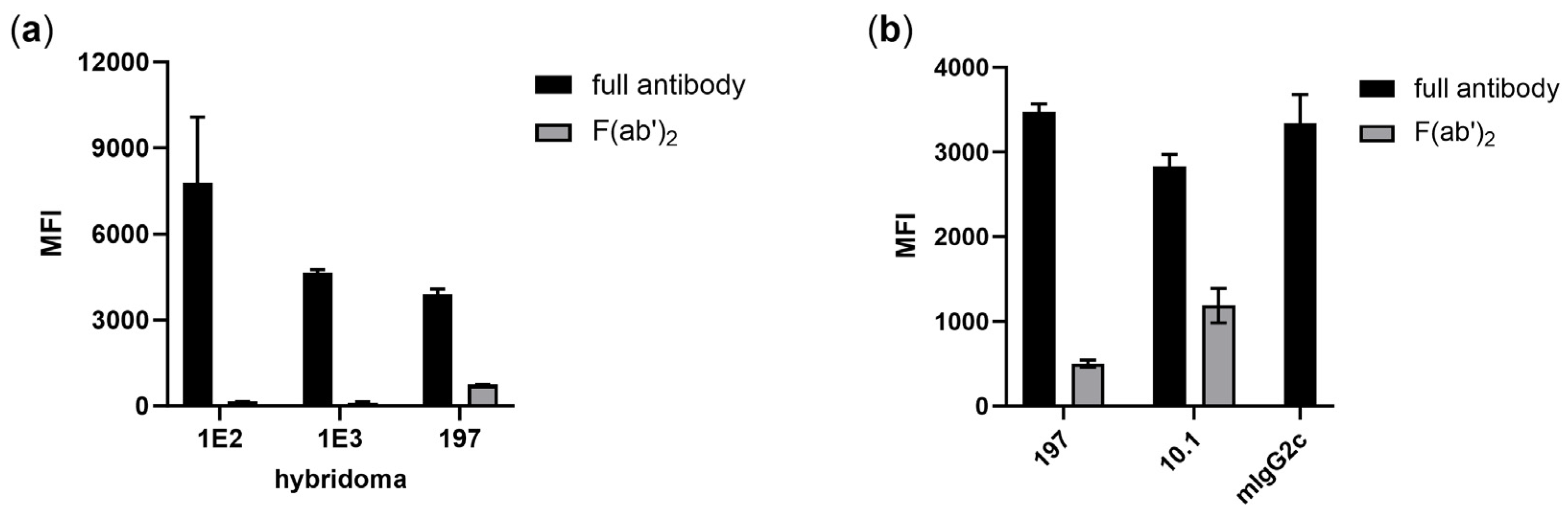
3.1. Isolation of FcγRI-Specific Antibodies by Hybridoma Technology Is Biased by Fc-Mediated Binding
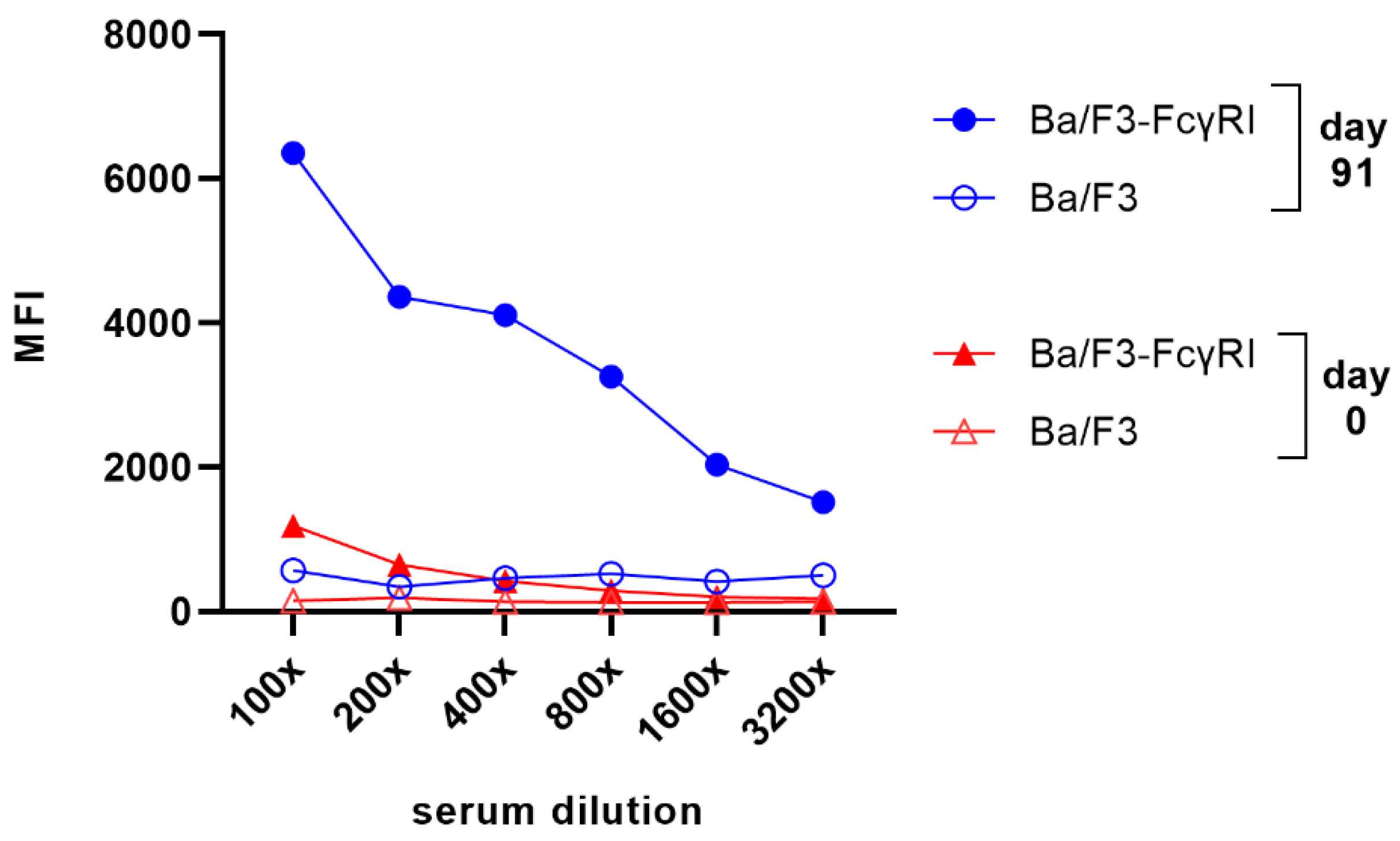
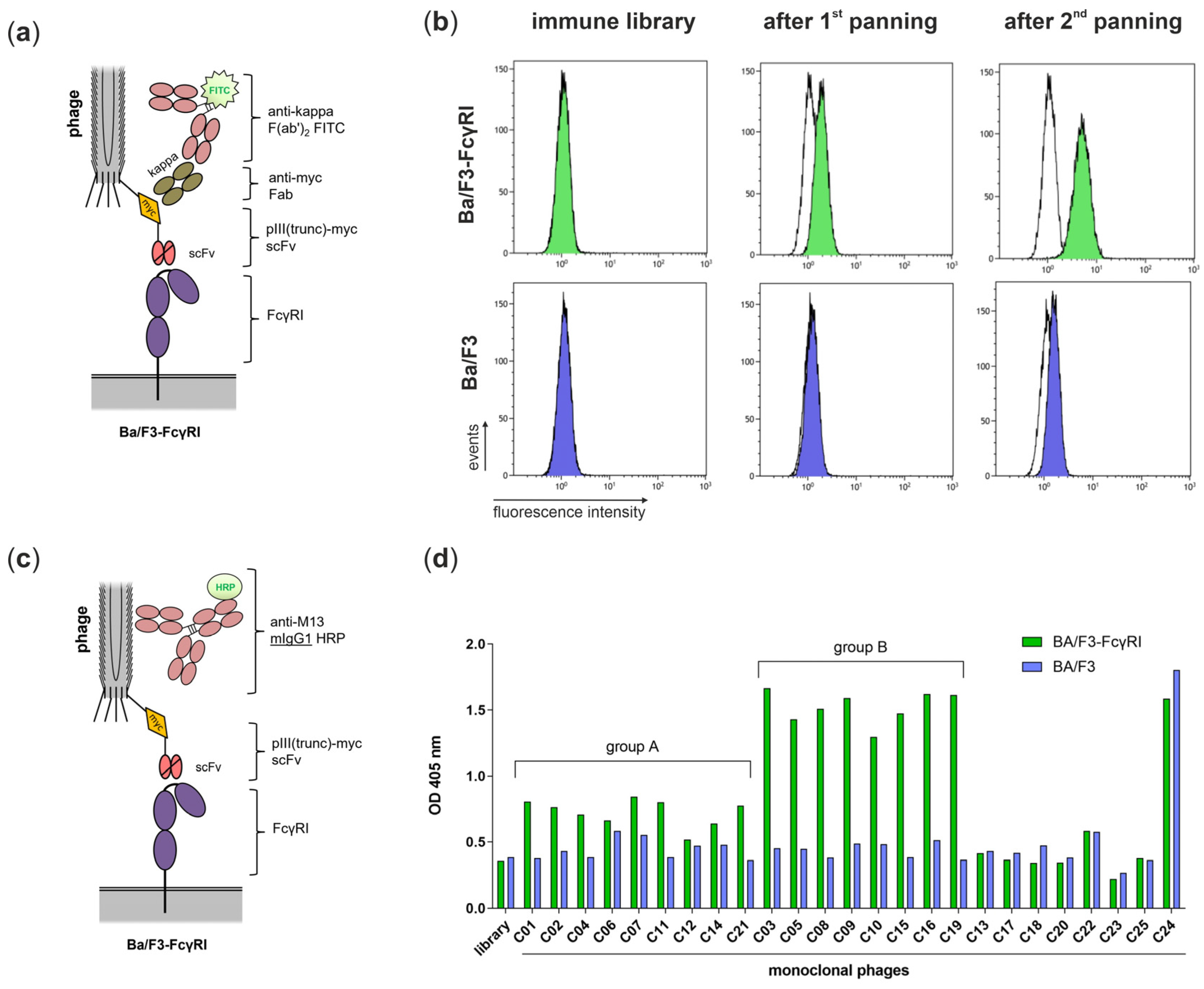
3.2. Isolation of FcγRI-Specific Antibodies by Phage Display Using Cellular Panning Obviating Fc-FcγRI Interactions
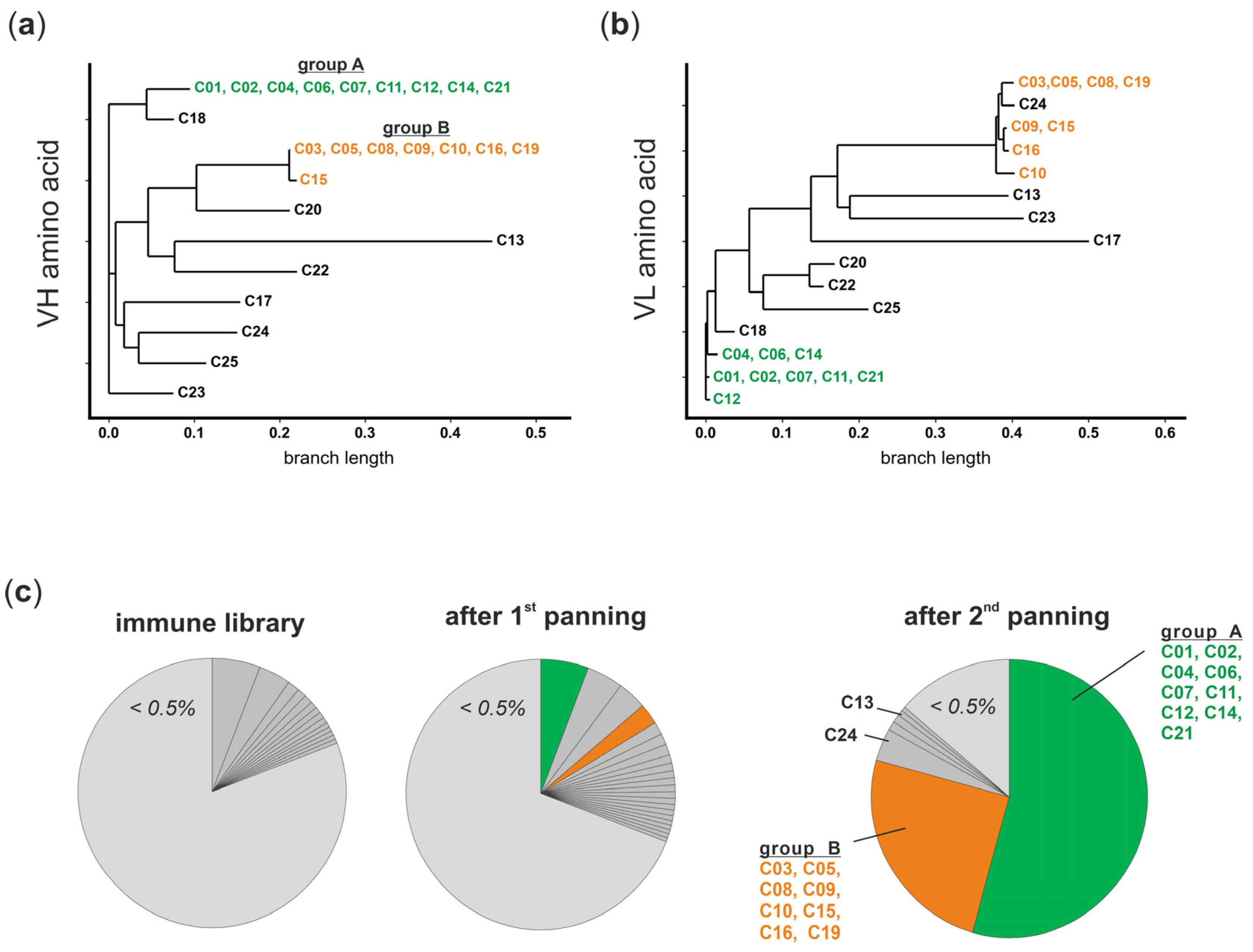
3.3. The Panel of FcγRI-Specific scFvs Can Be Grouped in Two VH Clonotypes
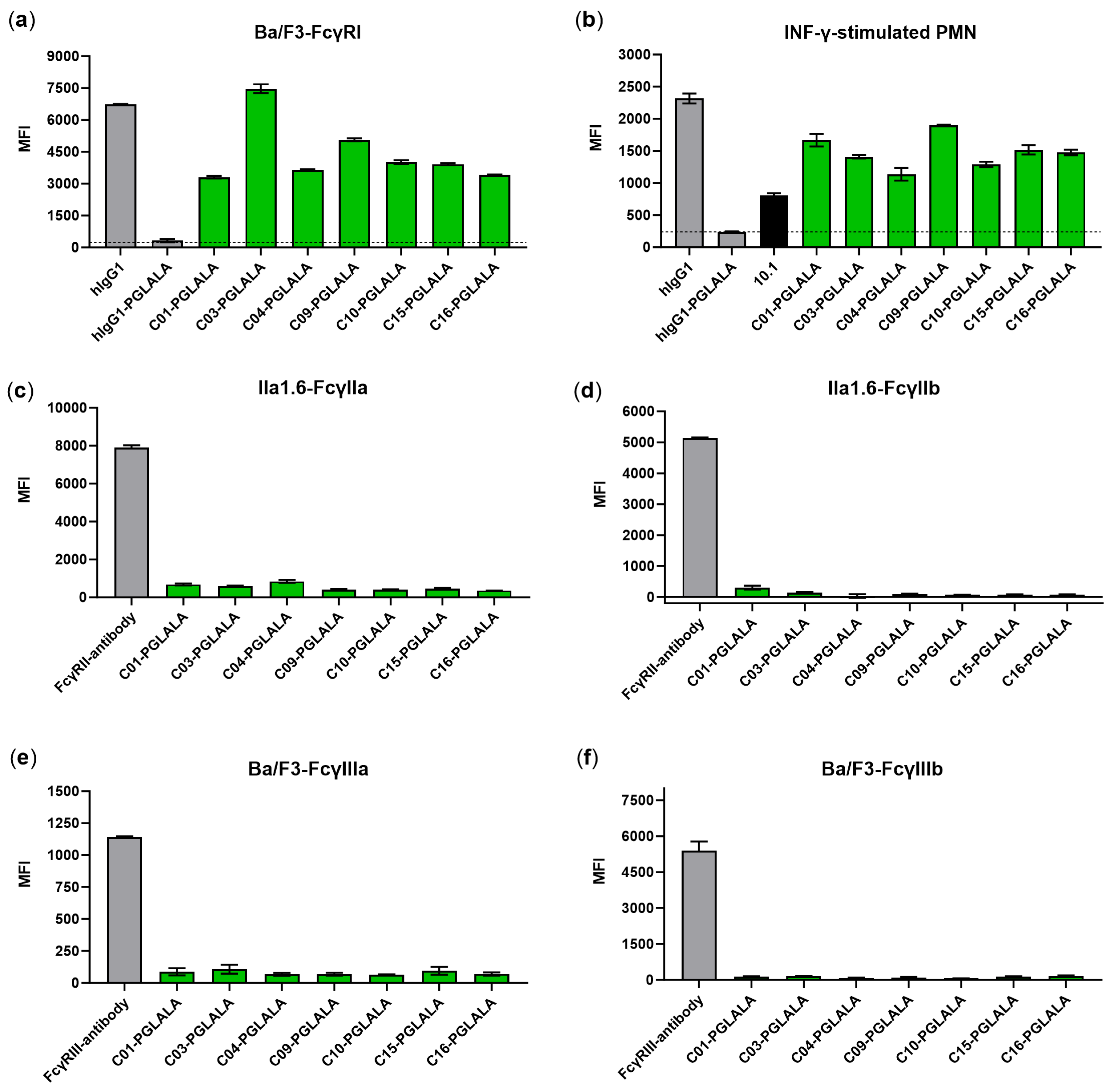
3.4. Chimeric Fc-Silent Antibodies Show FcγRI-Specific Binding
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nimmerjahn, F.; Ravetch, J.V. Fcgamma Receptors as Regulators of Immune Responses. Nat. Rev. Immunol. 2008, 8, 34–47. [Google Scholar] [CrossRef] [PubMed]
- van der Poel, C.E.; Karssemeijer, R.A.; Boross, P.; van der Linden, J.A.; Blokland, M.; van de Winkel, J.G.; Leusen, J.H. Cytokine-Induced Immune Complex Binding to the High-Affinity Igg Receptor, Fcgammari, in the Presence of Monomeric Igg. Blood 2010, 116, 5327–5333. [Google Scholar] [CrossRef] [PubMed]
- Di Ceglie, I.; Kruisbergen, N.N.L.; van den Bosch, M.H.J.; van Lent, P. Fc-Gamma Receptors and S100A8/A9 Cause Bone Erosion during Rheumatoid Arthritis. Do They Act as Partners in Crime? Rheumatology 2019, 58, 1331–1343. [Google Scholar] [CrossRef] [PubMed]
- Holtrop, T.; Budding, K.; Brandsma, A.M.; Leusen, J.H.W. Targeting the High Affinity Receptor, FcgammaRI, in Autoimmune Disease, Neuropathy, and Cancer. Immunother. Adv. 2022, 2, ltac011. [Google Scholar] [CrossRef] [PubMed]
- Fleit, H.B.; Wright, S.D.; Unkeless, J.C. Human Neutrophil Fc Gamma Receptor Distribution and Structure. Proc. Natl. Acad. Sci. USA 1982, 79, 3275–3279. [Google Scholar] [CrossRef] [PubMed]
- Looney, R.J.; Abraham, G.N.; Anderson, C.L. Human Monocytes and U937 Cells Bear Two Distinct Fc receptors for IgG. J. Immunol. 1986, 136, 1641–1647. [Google Scholar] [CrossRef] [PubMed]
- Köhler, G.; Milstein, C. Continuous Cultures of Fused Cells Secreting Antibody of Predefined Specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Mancardi, D.A.; Albanesi, M.; Jonsson, F.; Iannascoli, B.; Van Rooijen, N.; Kang, X.; England, P.; Daeron, M.; Bruhns, P. The High-Affinity Human IgG Receptor FcgammaRI (CD64) Promotes IgG-Mediated Inflammation, Anaphylaxis, and Antitumor Immunotherapy. Blood 2013, 121, 1563–1573. [Google Scholar] [CrossRef] [PubMed]
- Temming, A.R.; Bentlage, A.E.H.; de Taeye, S.W.; Bosman, G.P.; Lissenberg-Thunnissen, S.N.; Derksen, N.I.L.; Brasser, G.; Mok, J.Y.; van Esch, W.J.E.; Howie, H.L.; et al. Cross-Reactivity of Mouse IgG Subclasses to Human Fc Gamma Receptors: Antibody Deglycosylation only Eliminates IgG2b Binding. Mol. Immunol. 2020, 127, 79–86. [Google Scholar] [CrossRef]
- Guyre, P.M.; Graziano, R.F.; Vance, B.A.; Morganelli, P.M.; Fanger, M.W. Monoclonal Antibodies That Bind to Distinct Epitopes on Fc Gamma RI Are Able to Trigger Receptor Function. J. Immunol. 1989, 143, 1650–1655. [Google Scholar] [CrossRef]
- Dougherty, G.J.; Selvendran, Y.; Murdoch, S.; Palmer, D.G.; Hogg, N. The Human Mononuclear Phagocyte High-Affinity Fc Receptor, FcRI, Defined by a Monoclonal Antibody, 10.1. Eur. J. Immunol. 1987, 17, 1453–1459. [Google Scholar] [CrossRef] [PubMed]
- Beekman, J.M.; van der Poel, C.E.; van der Linden, J.A.; van den Berg, D.L.; van den Berghe, P.V.; van de Winkel, J.G.; Leusen, J.H. Filamin A Stabilizes Fc Gamma RI Surface Expression and Prevents Its Lysosomal Routing. J. Immunol. 2008, 180, 3938–3945. [Google Scholar] [CrossRef] [PubMed]
- Krohn, S.; Boje, A.S.; Gehlert, C.L.; Lutz, S.; Darzentas, N.; Knecht, H.; Herrmann, D.; Bruggemann, M.; Scheidig, A.J.; Weisel, K.; et al. Identification of New Antibodies Targeting Malignant Plasma Cells for Immunotherapy by Next-Generation Sequencing-Assisted Phage Display. Front. Immunol. 2022, 13, 908093. [Google Scholar] [CrossRef]
- Krebber, A.; Bornhauser, S.; Burmester, J.; Honegger, A.; Willuda, J.; Bosshard, H.R.; Pluckthun, A. Reliable Cloning of Functional Antibody Variable Domains from Hybridomas and Spleen Cell Repertoires Employing a Reengineered Phage Display System. J. Immunol. Methods 1997, 201, 35–55. [Google Scholar] [CrossRef] [PubMed]
- Peipp, M.; Kupers, H.; Saul, D.; Schlierf, B.; Greil, J.; Zunino, S.J.; Gramatzki, M.; Fey, G.H. A Recombinant CD7-Specific Single-Chain Immunotoxin is a Potent Inducer of Apoptosis in Acute Leukemic T Cells. Cancer Res. 2002, 62, 2848–2855. [Google Scholar] [PubMed]
- Bystry, V.; Reigl, T.; Krejci, A.; Demko, M.; Hanakova, B.; Grioni, A.; Knecht, H.; Schlitt, M.; Dreger, P.; Sellner, L.; et al. ARResT/Interrogate: An Interactive Immunoprofiler for IG/TR NGS Data. Bioinformatics 2017, 33, 435–437. [Google Scholar] [CrossRef] [PubMed]
- Schlothauer, T.; Herter, S.; Koller, C.F.; Grau-Richards, S.; Steinhart, V.; Spick, C.; Kubbies, M.; Klein, C.; Umana, P.; Mossner, E. Novel Human IgG1 and IgG4 Fc-Engineered Antibodies with Completely Abolished Immune Effector Functions. Protein Eng. Des. Sel. 2016, 29, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Ericson, S.G.; Coleman, K.D.; Wardwell, K.; Baker, S.; Fanger, M.W.; Guyre, P.M.; Ely, P. Monoclonal Antibody 197 (Anti-Fc Gamma RI) Infusion in a Patient with Immune Thrombocytopenia Purpura (ITP) Results in Down-Modulation of Fc Gamma RI on Circulating Monocytes. Br. J. Haematol. 1996, 92, 718–724. [Google Scholar] [CrossRef]
- Hayes, J.M.; Wormald, M.R.; Rudd, P.M.; Davey, G.P. Fc Gamma Receptors: Glycobiology and Therapeutic Prospects. J. Inflamm. Res. 2016, 9, 209–219. [Google Scholar] [CrossRef]
- Eisenhardt, S.U.; Peter, K. Phage Display and Subtractive Selection on Cells. In Antibody Engineering; Kontermann, R., Dübel, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 165–182. [Google Scholar]
- Prassler, J.; Thiel, S.; Pracht, C.; Polzer, A.; Peters, S.; Bauer, M.; Norenberg, S.; Stark, Y.; Kolln, J.; Popp, A.; et al. HuCAL PLATINUM, a Synthetic Fab library Optimized for Sequence Diversity and Superior Performance in Mammalian Expression Systems. J. Mol. Biol. 2011, 413, 261–278. [Google Scholar] [CrossRef]
- Kugler, J.; Wilke, S.; Meier, D.; Tomszak, F.; Frenzel, A.; Schirrmann, T.; Dubel, S.; Garritsen, H.; Hock, B.; Toleikis, L.; et al. Generation and Analysis of the Improved Human HAL9/10 Antibody Phage Display Libraries. BMC Biotechnol. 2015, 15, 10. [Google Scholar] [CrossRef] [PubMed]
- Frenzel, A.; Schirrmann, T.; Hust, M. Phage Display-Derived Human Antibodies in Clinical Development and Therapy. MAbs 2016, 8, 1177–1194. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Input Phages | Output Phages | Ratio | Enrichment | |
|---|---|---|---|---|
| after 1st panning | 1012 CFU | 19,200 CFU | 1.92 × 10−8 | - |
| after 2nd panning | 6 × 1011 CFU | 617,600 CFU | 1.03 × 10−6 | 54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krohn, S.; Holtrop, T.; Brandsma, A.M.; Moerer, P.; Nederend, M.; Darzentas, N.; Brüggemann, M.; Klausz, K.; Leusen, J.H.W.; Peipp, M. Combining Cellular Immunization and Phage Display Screening Results in Novel, FcγRI-Specific Antibodies. Viruses 2024, 16, 596. https://doi.org/10.3390/v16040596
Krohn S, Holtrop T, Brandsma AM, Moerer P, Nederend M, Darzentas N, Brüggemann M, Klausz K, Leusen JHW, Peipp M. Combining Cellular Immunization and Phage Display Screening Results in Novel, FcγRI-Specific Antibodies. Viruses. 2024; 16(4):596. https://doi.org/10.3390/v16040596
Chicago/Turabian StyleKrohn, Steffen, Tosca Holtrop, Arianne M. Brandsma, Petra Moerer, Maaike Nederend, Nikos Darzentas, Monika Brüggemann, Katja Klausz, Jeanette H. W. Leusen, and Matthias Peipp. 2024. "Combining Cellular Immunization and Phage Display Screening Results in Novel, FcγRI-Specific Antibodies" Viruses 16, no. 4: 596. https://doi.org/10.3390/v16040596
APA StyleKrohn, S., Holtrop, T., Brandsma, A. M., Moerer, P., Nederend, M., Darzentas, N., Brüggemann, M., Klausz, K., Leusen, J. H. W., & Peipp, M. (2024). Combining Cellular Immunization and Phage Display Screening Results in Novel, FcγRI-Specific Antibodies. Viruses, 16(4), 596. https://doi.org/10.3390/v16040596