
Characterization of Mycoviruses in Armillaria ostoyae and A. cepistipes in the Czech Republic
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Isolation of Double-Stranded (ds) RNA
2.3. Isolation of Total RNA
2.4. Stranded Total RNA Sequencing
2.5. Bioinformatics
2.5.1. Data Preprocessing
2.5.2. Host Reads’ Removal
2.5.3. Discovery of Known Virus
2.5.4. Discovery of Novel Viruses
2.5.5. Ribozymes’ Detection
2.6. Retro Transcription Polymerase Chain Reaction (RT-PCR) and Sanger Sequencing
2.7. Conserved Domains
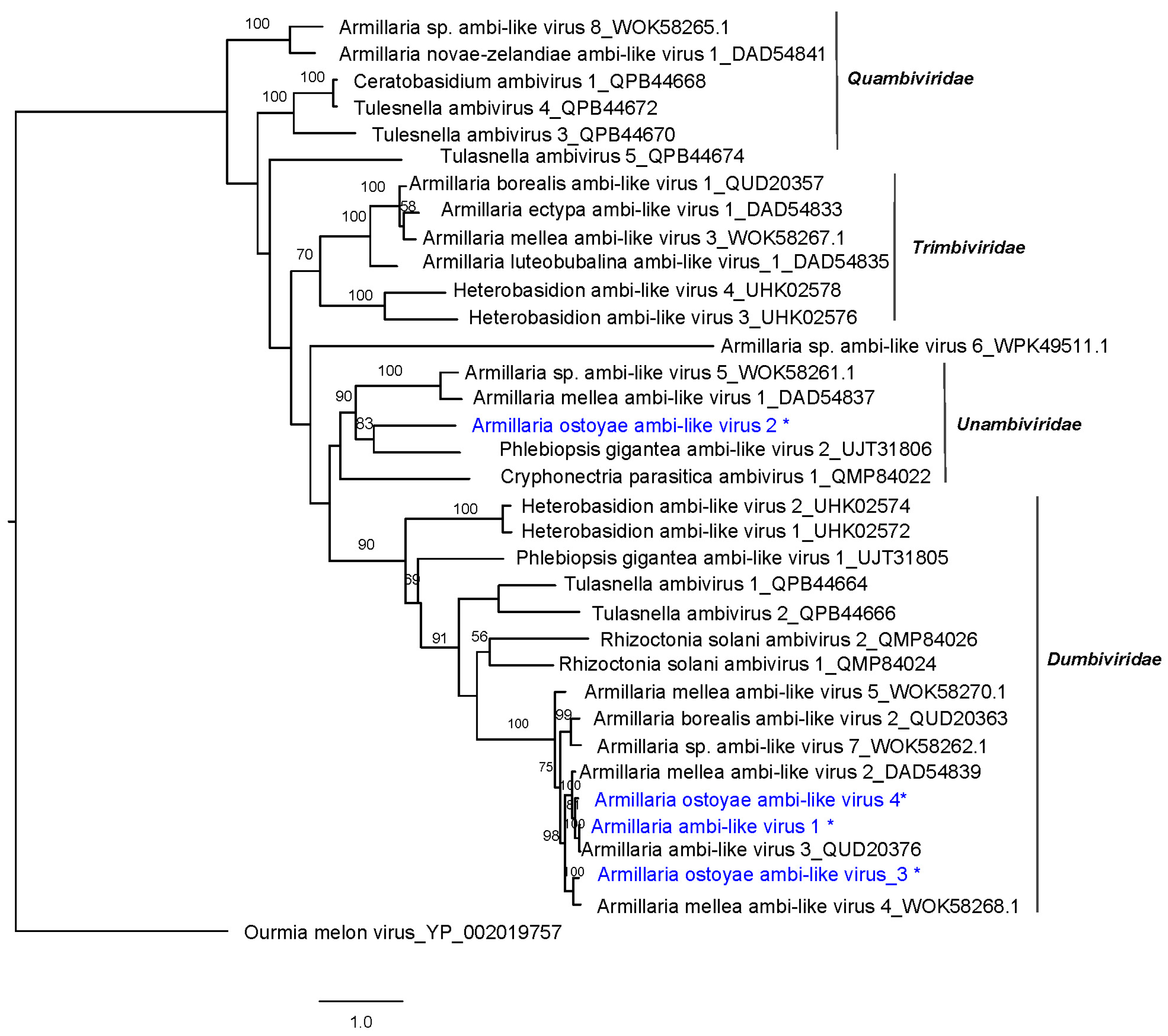
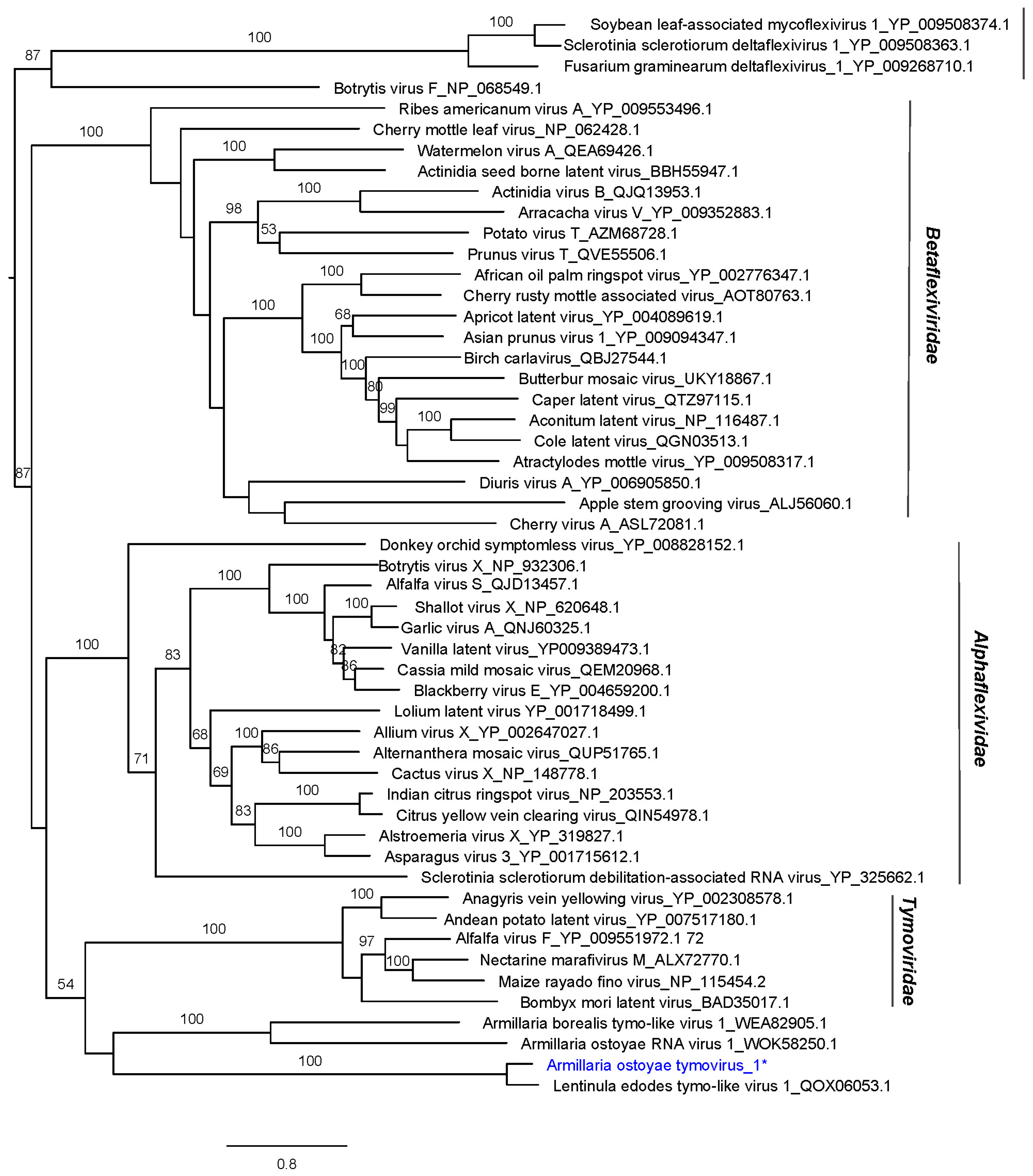
2.8. Phylogenetic Analyses
3. Results and Discussion
3.1. dsRNA Screening
3.2. Identification of ssRNA Viruses
3.3. RT-PCR Screening
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Hansen, E.M.; Goheen, E.M. Phellinus weirii and other native root pathogens as determinants of forest structure and process in Western North America. Annu. Rev. Phytopathol. 2000, 38, 515–539. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, G.S. Evolutionary ecology of plant diseases in natural ecosystems. Annu. Rev. Phytopathol. 2002, 40, 13–43. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, M.; Wingfield, B.D.; Wingfield, M.J. Armillaria root-rot pathogens: Species boundaries and global distribution. Pathogens 2018, 7, 83. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.E.; Shaw, C.G.; Wargo, P.M.; Sites, W.H. Armillaria Root Disease. In Forest Insect and Disease Leaflet 78; U.S. Department of Agriculture Forest Service: Radnor, PA, USA, 1986. [Google Scholar]
- Baumgartner, K.; Coetzee, M.; Hoffmeister, D. Secrets of the subterranean pathosystem of Armillaria. Mol. Plant Pathol. 2011, 12, 515–534. [Google Scholar] [CrossRef] [PubMed]
- Holuša, J.; Lubojacký, J.; Čurn, V.; Tonka, T.; Resnerová, K.; Horák, J. Combined effects of drought stress and Armillaria infection on tree mortality in Norway spruce plantations. For. Ecol. Manag. 2018, 427, 434–445. [Google Scholar] [CrossRef]
- Cienciala, E.; Tumajer, J.; Zatloukal, V.; Beranová, J.; Holá, Š.; Hůnová, I.; Russ, R. Recent spruce decline with biotic pathogen infestation as a result of interacting climate, deposition and soil variables. Eur. J. Forest. Res. 2017, 136, 307–317. [Google Scholar] [CrossRef]
- Turčáni, M. Podiel podkorneho hmyzu na hynutí smrekových porastov postihnutých žltnutím. In Aktuálne Problémy v Ochrane Lesa; Zúbrik, M., Ed.; Lesnický Výskumný Ústav: Banská Štiavnica, Slovakia, 2001; pp. 56–60. [Google Scholar]
- Jankovský, L. Role houbových patogenů v chřadnutí smrku. In Collection of Lectures of the Professional Seminar Budišov Nad Budišovkou; Research Institute of Forestry and Hunting: Budišov and Budišovkou, Czech Republic, 2014; pp. 20–30. [Google Scholar]
- Garcia, O. A simple and effective forest stand mortality model. Math. Comput. For. Nat.-Resour. Sci. 2009, 1, 1–9. [Google Scholar]
- Van Mantgem, P.J.; Stephenson, N.L.; Byrne, J.C.; Daniels, L.D.; Franklin, J.F.; Fulé, P.Z.; Harmon, M.E.; Larson, A.J.; Smith, J.M.; Taylor, A.H.; et al. Widespread increase of tree mortality rates in the western United States. Science 2009, 323, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Cleary, M.R.; Arhipova, N.; Morrison, D.J.; Thomsen, I.M.; Sturrock, R.N.; Vasaitis, R.; Gaitnieks, T.; Stenlid, J. Stump removal to control root disease in Canada and Scandinavia: A synthesis of results from long-term trial. For. Ecol. Manag. 2013, 290, 5–14. [Google Scholar] [CrossRef]
- Linnakoski, R.; Sutela, S.; Coetzee, M.P.A.; Duong, T.A.; Pavlov, I.N.; Litovka, Y.A. Armillaria root rot fungi host single-stranded RNA viruses. Sci. Rep. 2021, 11, 7336. [Google Scholar] [CrossRef]
- Prospero, S.; Botella, L.; Santini, A.; Robin, C. Biological control of emerging forest diseases: How can we move from dreams to reality? For. Ecol. Manag. 2021, 496, 119377. [Google Scholar] [CrossRef]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef] [PubMed]
- García-Pedrajas, M.D.; Cañizares, M.C.; Sarmiento-Villamil, J.L.; Jacquat, A.G.; Dambolena, J.S. Mycoviruses in Biological Control: From Basic Research to Field Implementation. Phytopathology 2019, 109, 1828–1839. [Google Scholar] [CrossRef]
- Tonka, T.; Walterova, L.; Curn, V. Biological control of pathogenic fungi: Can mycoviruses play an important role? J. Cent. Eur. Agric. 2022, 23, 540–551. [Google Scholar] [CrossRef]
- Muñoz-Adalia, E.J.; Fernández, M.M.; Diez, J.J. The use of mycoviruses in the control of forest diseases. Biocontrol Sci. Technol. 2016, 26, 577–604. [Google Scholar] [CrossRef]
- Vainio, E.J.; Jurvansuu, J.; Hyder, R.; Kashif, M.; Piri, T.; Tuomivirta, T.; Poimala, A.; Xu, P.; Mäkelä, S.; Nitisa, D.; et al. Heterobasidion partitivirus 13 mediates severe growth debilitation and major alterations in the gene expression of a fungal forest pathogen. J. Virol. 2018, 92, 1744. [Google Scholar] [CrossRef]
- Sutela, S.; Piri, T.; Vainio, E.J. Discovery and community dynamics of novel ssRNA mycoviruses in the conifer pathogen Heterobasidion parviporum. Front. Microbiol. 2021, 12, 770787. [Google Scholar] [CrossRef] [PubMed]
- Blattny, C.; Kralik, O.; Veselsky, J.; Kasala, B.; Herzova, H. Particles resembling virions accompanying the proliferation of Agaric mushrooms. Česká Mykol. 1973, 27, 1–5. [Google Scholar]
- Reaves, J.L.; Allen, T.C.; Shaw, C.G.; Dashek, W.V.; Mayfield, J.E. Occurrence of virus like particles in isolates of Armillaria. J. Ultrastruct. Mol. Struct. Res. 1988, 98, 217–221. [Google Scholar] [CrossRef]
- Tonka, T.; Walterová, L.; Hejna, O.; Čurn, V. Molecular characterization of a ssRNA mycovirus isolated from the forest pathogenic fungus Armillaria ostoyae. Acta Virol. 2022, 66, 290–294. [Google Scholar] [CrossRef]
- Shamsi, W.; Heinzelmann, R.; Ulrich, S.; Kondo, H.; Cornejo, C. Decoding the RNA virome of the tree parasite Armillaria provides new insights into the viral community of soil-borne fungi. Environ. Microbiol. 2024, 26, e16583. [Google Scholar] [CrossRef] [PubMed]
- Morris, T.J.; Dodds, J.A. Isolation and analysis of double-stranded RNA from virus-infected plant and fungal tissue. Phytopathology 1979, 69, 854. [Google Scholar] [CrossRef]
- Tonka, T.; Walterová, L.; Hejna, O.; Čurn, V. Metodika Identifikace, Determinace a Přenosu Mykovirů u Hub Rodu Armillaria; University of South Bohemia in České Budějovice: České Budějovice, Czech Republic, 2021; ISBN 978-80-7394-891-7. [Google Scholar]
- Botella, L.; Dvořák, M.; Capretti, P.; Luchi, N. Effect of temperature on GaRV6 accumulation and its fungal host, the conifer pathogen Gremmeniella abietina. For. Pathol. 2017, 47, 12291. [Google Scholar] [CrossRef]
- Botella, L.; Jung, T. Multiple viral infections detected in Phytophthora condilina by total and small RNA sequencing. Viruses 2021, 13, 620. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. Babraham Bioinformatics—FastQC. A Quality Control Tool for High Throughput Sequence Data. Soil. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 26 May 2023).
- Illumina Adapter Sequences. Available online: https://emea.support.illumina.com/downloads/illumina-adapter-sequences-document-1000000002694.html (accessed on 26 May 2023).
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. 2012. Available online: https://cutadapt.readthedocs.io/en/stable (accessed on 26 May 2023).
- GitHub. Available online: https://github.com/marcelm/cutadapt (accessed on 27 May 2023).
- Dobin, A.; Davies, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Sonali, J.; Philippe, B.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Viral NCBI. Available online: https://www.ncbi.nlm.nih.gov/labs/virus/vssi/#/ (accessed on 29 May 2023).
- Burrows-Wheeler Alingner. Available online: https://bio-bwa.sourceforge.net/ (accessed on 27 May 2023).
- IGV. Available online: https://software.broadinstitute.org/software/igv/ (accessed on 27 May 2023).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Viral UniProtKB. Available online: https://www.uniprot.org/uniprotkb?query=10239 (accessed on 29 May 2023).
- RVDB. Available online: https://rvdb.dbi.udel.edu/ (accessed on 29 May 2023).
- Virus-Host DB. Available online: https://www.genome.jp/virushostdb/ (accessed on 29 May 2023).
- Lee, B.D.; Neri, U.; Roux, S.; Wolf, Y.I.; Camargo, A.P.; Krupovic, M.; Simmonds, P.; Kyrpides, N.; Gophna, U.; Dolja, V.V.; et al. Mining metatranscriptomes reveals a vast world of viroid-like circular RNAs. Cell 2023, 186, 646–661. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Xu, T.; Lin, W.; Jia, Q.; He, Q.; Liu, K.; Du, J.; Chen, L.; Yang, X.; Du, F.; et al. Reference-free and de novo Identification of Circular RNAs. bioRxiv 2020. [Google Scholar] [CrossRef]
- Nawrocki, E.P.; Eddy, S.R. Infernal 1.1: 100-fold faster RNA homology searches. Bioinformatics 2013, 29, 2933–2935. [Google Scholar] [CrossRef]
- Macke, T.J.; Ecker, D.J.; Gutell, R.R.; Gautheret, D.; Case, D.A.; Sampath, R. RNAMotif, an RNA secondary structure definition and search algorithm. Nucleic Acids Res. 2001, 29, 4724–4735. [Google Scholar] [CrossRef]
- Tonka, T.; Walterová, L.; Čurn, V. Development of RT-PCR for rapid detection of ssRNA ambi-like mycovirus in a root rot fungi (Armillaria spp.). Acta Virol. 2022, 66, 287–289. [Google Scholar] [CrossRef]
- Čurn, V.; Tonka, T.; Křížová, L.; Jozová, E. Metodika Izolace DNA Analýzy Molekulárních Markerů u Hub; University of South Bohemia in České Budějovice: České Budějovice, Czech Republic, 2019; ISBN 978-80-7394-781-1. [Google Scholar]
- NCBI CD-Search Tool. Available online: https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi (accessed on 27 May 2023).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J.; Diego, S.; Jolla, L. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop, New Orleans, LA, USA, 14 November 2010. [Google Scholar] [CrossRef]
- Sutela, S.; Forgia, M.; Vainio, E.J.; Chiapello, M.; Daghino, S.; Vallino, M.; Martino, E.; Girlanda, M.; Perotto, S.; Turina, M. The virome from a collection of endomycorrhizal fungi reveals new viral taxa with unprecedented genome organization. Virus Evol. 2020, 6, 76. [Google Scholar] [CrossRef]
- Forgia, M.; Isgandarli, E.; Aghayeva, D.N.; Huseynova, I.; Turina, M. Virome characterization of Cryphonectria parasitica isolates from Azerbaijan unveiled a new mymonavirus and a putative new RNA virus unrelated to described viral sequences. Virology 2021, 553, 51–61. [Google Scholar] [CrossRef]
- Turina, M.; Lee, B.D.; Sabanadzovic, S.; Vainio, E.J.; Navarro, B.; Simmonds, P. Create One New Phylum, Ambiviricota, Including One New Class, One New Order, Four New Families, Four New Genera, and 20 New Species, in Kingdom Orthornavirae (Realm Riboviria). ICTV TaxoProp 2023.007F. 2023. Available online: https://ictv.global/files/proposals/pending?fid=11741#block-teamplus-page-title (accessed on 27 May 2023).
- Forgia, M.; Navarro, B.; Daghino, S.; Cervera, A.; Gisel, A.; Perotto, S.; Aghayeva, D.N.; Akinyuwa, M.F.; Gobbi, E.; Zheludev, I.N.; et al. Hybrids of RNA viruses and viroid-like elements replicate in fungi. Nat. Commun. 2023, 14, 2591. [Google Scholar] [CrossRef]
- Dvořák, J. Výskyt RNA Elementů u Rodu Armillaria. Master’s Thesis, Charles University, Prague, Czech Republic, 2008. [Google Scholar]
- Drenkhan, T.; Sutela, S.; Veeväli, V.; Vainio, E.J. Phlebiopsis gigantea strains from Estonia show potential as native biocontrol agents against Heterobasidion root rot and contain diverse dsRNA and ssRNA viruses. Biol. Control 2022, 167, 104837. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Kreuze, J.F.; Martelli, G.P. Tymovirales. In Virus Taxonomy: Classification and Nomenclature of Viruses; Elsevier: Waltham, MA, USA, 2011; pp. 901–903. [Google Scholar]
- King, A.M.Q.; Michael, J.; Carstens, E.B.; Lefkowitz, E.J. Virus Taxonomy: Classification and Nomenclature of Viruses; American Press/Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Howitt, R.L.; Beever, R.E.; Pearson, M.N.; Forster, R.L. Genome characterization of botrytis virus F, aflexuous rod-shaped mycovirus resembling plant ‘potex-like’ viruses. J. Gen. Virol. 2001, 82, 67–78. [Google Scholar] [CrossRef]
- Adams, M.J.; Candresse, T.; Hammond, J.; Kreuze, J.F.; Martelli, G.P.; Mamba, S.; Pearson, M.N.; Ryu, K.H.; Vaira, A.M. Alphaflexiviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses; Elsevier: Waltham, MA, USA, 2011; pp. 904–919. [Google Scholar] [CrossRef]
- Howitt, R.L.; Beever, R.E.; Pearson, M.N.; Forster, R.L. Genome characterization of a flexuous rod-shaped mycovirus, botrytis virus X, reveals high amino acid identity to genes from plant ‘potex-like’ viruses. Arch. Virol. 2006, 151, 563–579. [Google Scholar] [CrossRef]
- Li, K.; Zheng, D.; Cheng, J.; Chen, T.; Fu, Y.; Jiang, D.; Xie, J. Characterization of a novel Sclerotinia sclerotiorum RNA virus as the prototype of a new proposed family within the order Tymovirales. Virus Res. 2016, 219, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lin, Y.; Zhang, H.; Wang, S.; Qiu, D.; Guo, L. Molecular characterization of a novel mycovirus of the family Tymoviridae isolated from the plant pathogenic fungus Fusarium graminearum. Virology 2016, 489, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Bartholomaus, A.; Wibberg, D.; Winkler, A.; Puhler, A.; Schluter, A.; Varrelmann, M. Identification of a novel mycovirus isolated from Rhizoctonia solani (AG 2-2 IV) provides further information about genome plasticity within the order Tymovirales. Arch. Virol. 2017, 162, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, Y.; Abraham, A.; Uesaka, K.; Kondo, H.; Suga, H.; Suzuki, N.; Chiba, S. Novel mitoviruses and a unique tymo-like virus in hypovirulent and virulent strains of the Fusarium head blight fungus, Fusarium boothii. Viruses 2018, 10, 584. [Google Scholar] [CrossRef] [PubMed]
- Coenen, A.; Kevei, F.; Hoekstra, R.F. Factors affecting the spread of double-stranded RNA viruses in Aspergillus nidulans. Genet. Res. 1997, 69, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Melzer, M.S.; Ikeda, S.S.; Boland, G.J. Interspecific transmission of double-stranded RNA and hypovirulence from Sclerotinia sclerotiorum to S. minor. Phytopathology 2002, 92, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Linder-Basso, D.; Hillman, B.; Kaneko, S.; Milgroom, M.G. Evidence for interspecies transmission of viruses in natural populations of filamentous fungi in the genus Cryphonectria. Mol. Ecol. 2003, 12, 1619–1628. [Google Scholar] [CrossRef]
- Ihrmark, K.; Johannesson, H.; Stenström, E.; Stenlid, J. Transmission of double-stranded RNA in Heterobasidion annosum. Fungal Genet. Biol. 2002, 36, 147–154. [Google Scholar] [CrossRef]
- Vainio, E.J.; Korhonen, K.; Tuomivirta, T.T.; Hantula, J. A novel putative partitivirus of the saprotrophic fungus Heterobasidion ecrustosum infects pathogenic species of the Heterobasidion annosum complex. Fungal Biol. 2010, 114, 955–965. [Google Scholar] [CrossRef]
- Vainio, E.J.; Hakanpää, J.; Dai, Y.C.; Hansen, E.; Korhonen, K.; Hantula, J. Species of Heterobasidion host a diverse pool of partitiviruses with global distribution and interspecies transmission. Fungal Biol. 2011, 115, 1234–1243. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Code | Fungal Species | Site Coordinates | Country | Tree Host | Fungal Material | Collection Date (Day.Month.Year) |
|---|---|---|---|---|---|---|
| 1 | A. cepistipes | 49.5147885 N, 17.5546327 E | Czech Republic | Picea abies | Rhizomorphs | 29.9.2020 |
| 2 | A. ostoyae | 48.9813906 N, 14.4205031 E | Czech Republic | Picea abies | Fruiting body | 31.10.2019 |
| 3 | A. ostoyae | 49.81582 N, 17.34951 E | Czech Republic | Picea abies | Fruiting body | 27.10.2019 |
| 4 | A. ostoyae | 48.6232622 N, 14.6442581 E | Czech Republic | Picea abies | Fruiting body | 16.10.2019 |
| 5 | A. ostoyae | 50.10238 N, 16.0683 E | Czech Republic | Picea abies | Fruiting body | 23.10.2019 |
| 6 | A. ostoyae | 49.5226228 N, 17.5717912 E | Czech Republic | Picea abies | Rhizomorphs | 5.9.2019 |
| 7 | A. ostoyae | 49.8084936 N, 17.4939458 E | Czech Republic | Picea abies | Rhizomorphs | 4.8.2020 |
| 8 | A. ostoyae | 49.31372 N, 16.77221 E | Czech Republic | Picea abies | Fruiting body | 23.10.2019 |
| 9 | A. ostoyae | 48.6078636 N, 14.6688486 E | Czech Republic | Picea abies | Fruiting body | 21.10.2019 |
| 10 | A. ostoyae | 49.8084936 N, 17.4939458 E | Czech Republic | Picea abies | Rhizomorphs | 4.8.2020 |
| 11 | A. ostoyae | 49.32247 N, 16.78645 E | Czech Republic | Picea abies | Fruiting body | 23.10.2019 |
| 12 | A. ostoyae | 48.9815597 N, 14.4162544 E | Czech Republic | Picea abies | Fruiting body | 4.11.2020 |
| 13 | A. ostoyae | 49.5419285 N, 17.3919515 E | Czech Republic | Picea abies | Rhizomorphs | 14.8.2019 |
| Virus Name | Acronym | L | Accession Number a | Most Similar Virus b | E Value | Q (%) | I (%) | Mapped Reads | Depth of Coverage |
|---|---|---|---|---|---|---|---|---|---|
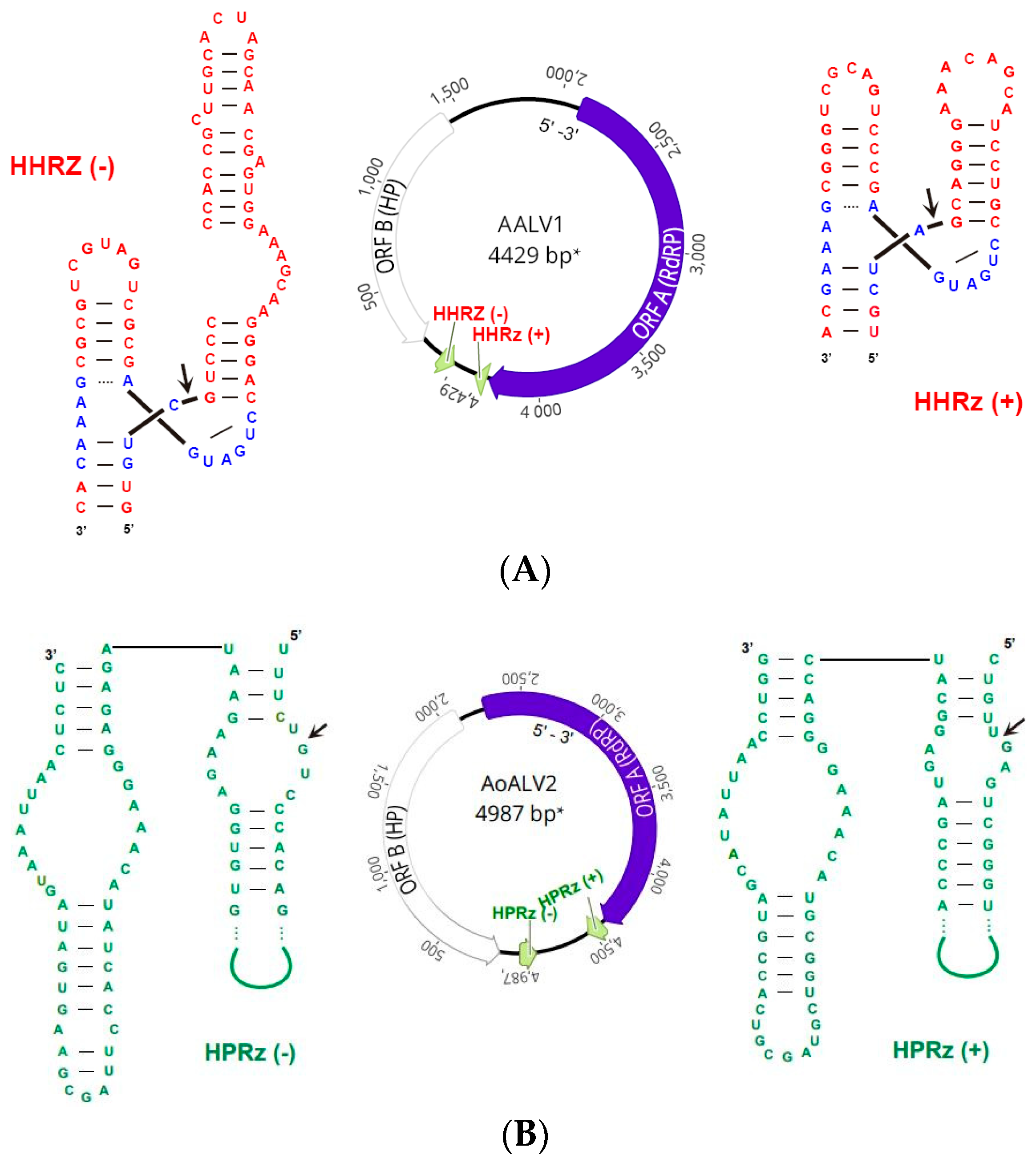
| Armillaria ambi-like virus 1 | AALV1 | 4663 | ON380550 | Armillaria spp. ambi-like virus 3 | 0.0 | 96 | 96.31 | 24,902 | 780.91 |
| Armillaria ostoyae ambi-like virus 2 | AoALV2 | 4541 | ON380551 | Phlebiopsis gigantea ambi-like virus 2 | 2 × 10−103 | 41 | 36.65 | 27,239 | 863.10 |
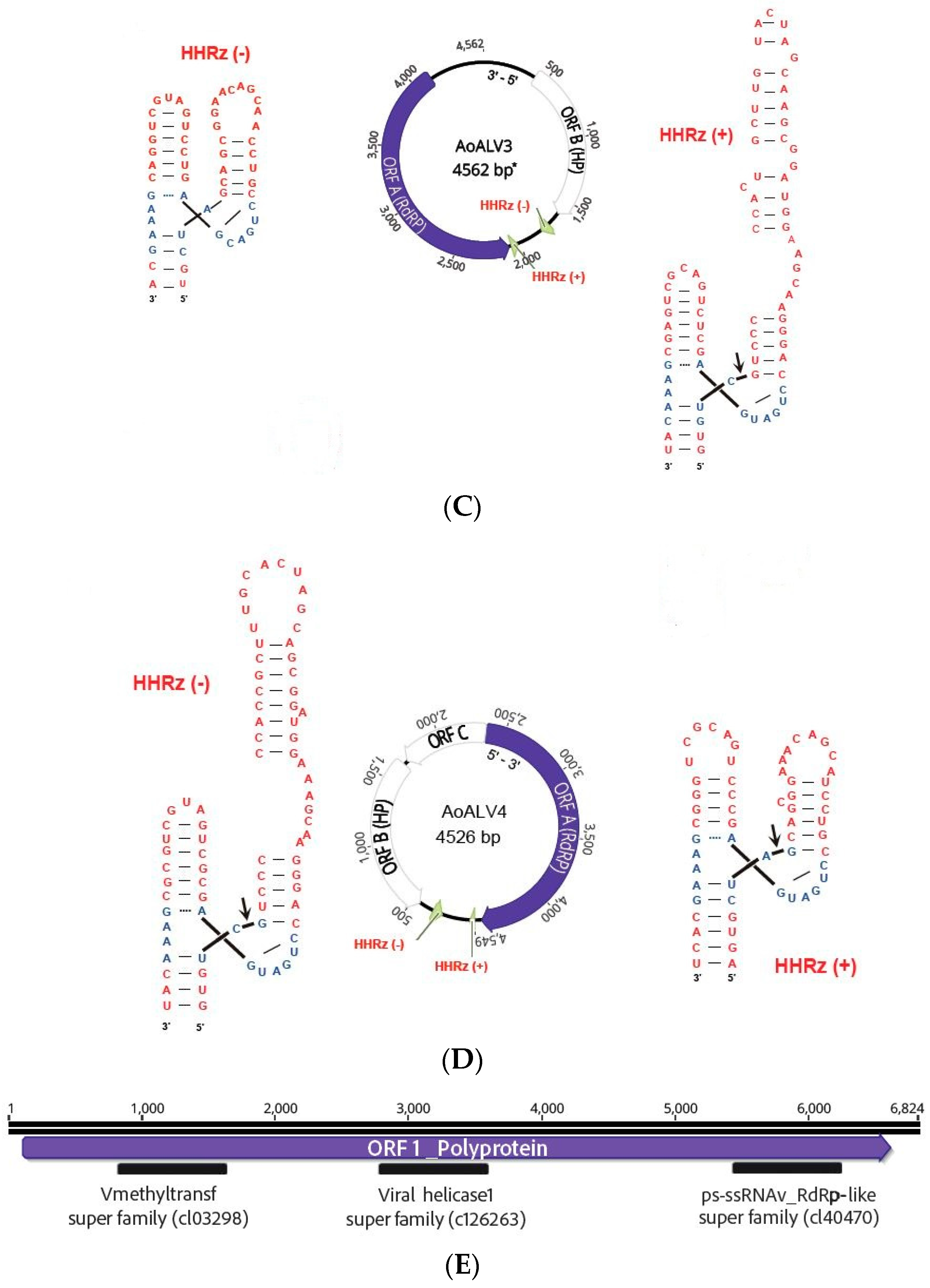
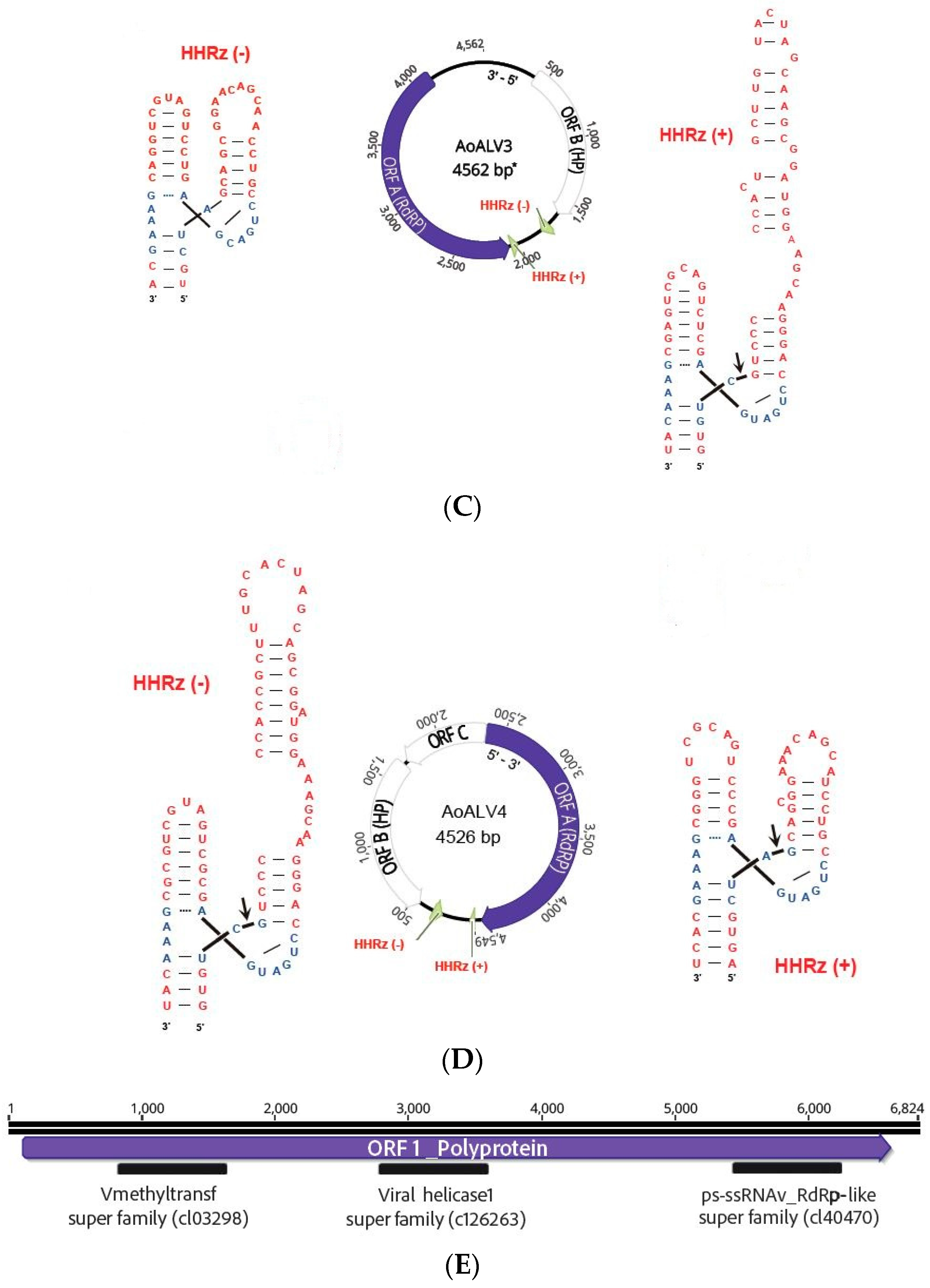
| Armillaria ostoyae ambi-like virus 3 | AoALV3 | 4562 | ON380552 | Armillaria mellea ambi like virus 4 | 0.0 | 56 | 80.94 | 21,362 | 678.90 |
| Armillaria ostoyae ambi-like virus 4 | AoALV4 | 4549 | ON380553 | Armillaria ambi-like virus 3 | 0.0 | 46 | 91.57 | 6169 | 196.26 |
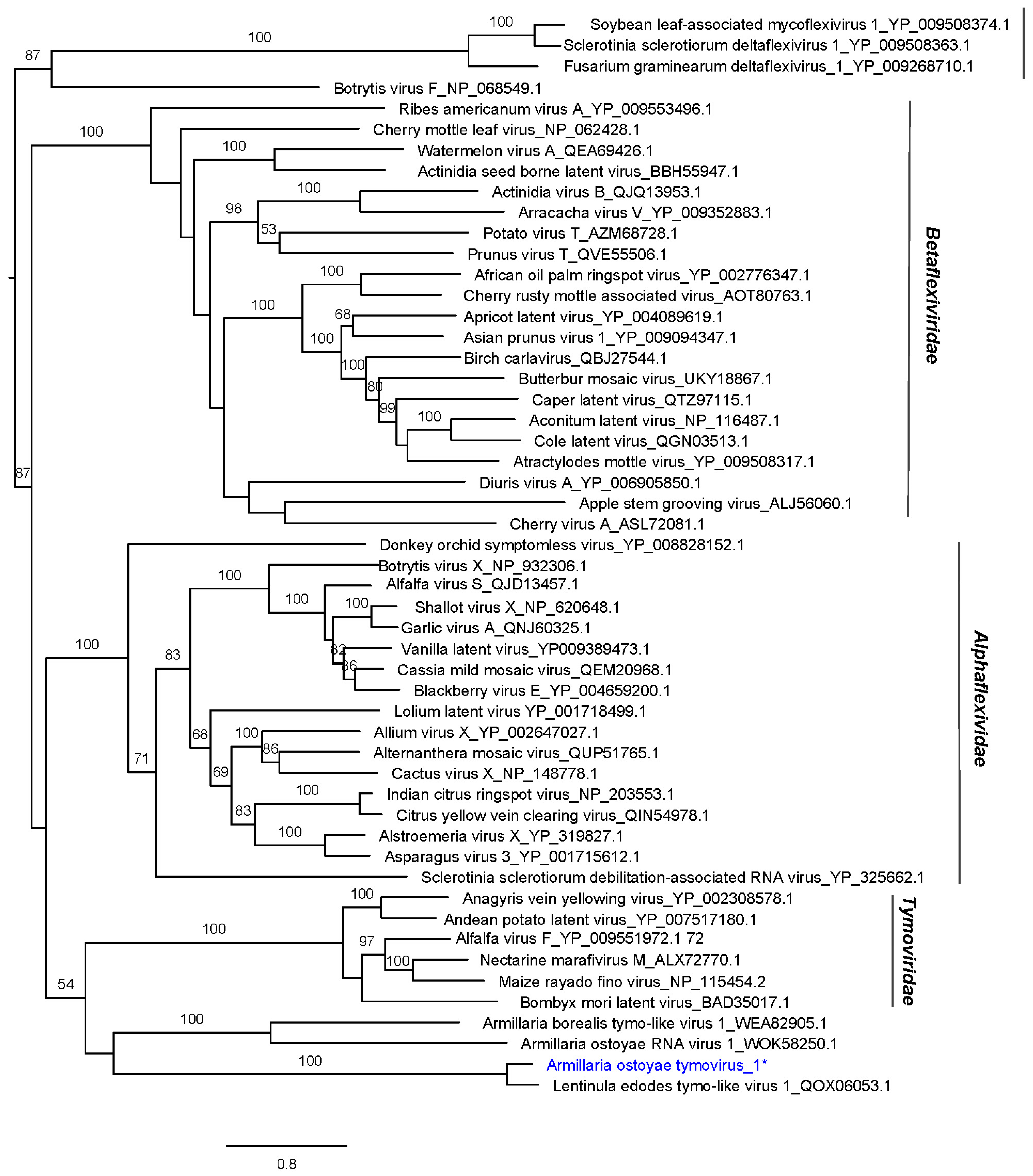
| Armillaria ostoyae tymovirus 1 | AoTV1 | 6824 | ON380554 | Lentinula edodes tymo-like virus 1 | 0.0 | 94 | 68.16 | 4959 | 106.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walterová, L.; Botella, L.; Hejna, O.; de la Peña, M.; Tonka, T.; Čurn, V. Characterization of Mycoviruses in Armillaria ostoyae and A. cepistipes in the Czech Republic. Viruses 2024, 16, 610. https://doi.org/10.3390/v16040610
Walterová L, Botella L, Hejna O, de la Peña M, Tonka T, Čurn V. Characterization of Mycoviruses in Armillaria ostoyae and A. cepistipes in the Czech Republic. Viruses. 2024; 16(4):610. https://doi.org/10.3390/v16040610
Chicago/Turabian StyleWalterová, Lucie, Leticia Botella, Ondřej Hejna, Marcos de la Peña, Tomáš Tonka, and Vladislav Čurn. 2024. "Characterization of Mycoviruses in Armillaria ostoyae and A. cepistipes in the Czech Republic" Viruses 16, no. 4: 610. https://doi.org/10.3390/v16040610