
Multiomics Identification of Potential Targets for Alzheimer Disease and Antrocin as a Therapeutic Candidate

 ,
,  ,
,
Abstract
:1. Introduction
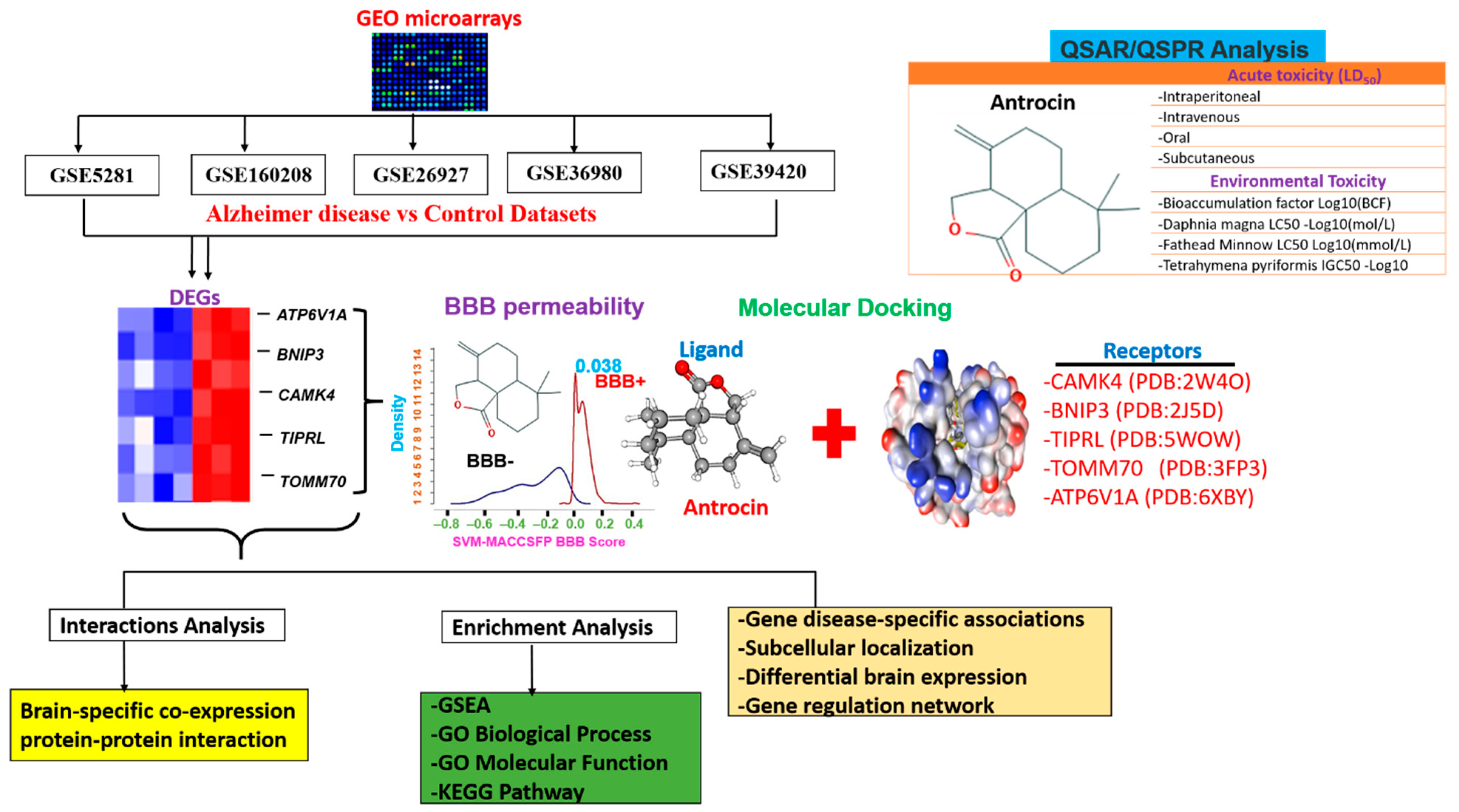
2. Methods
2.1. Collection of Microarray Data of Early Onset Alzheimer’s Disease (AD)
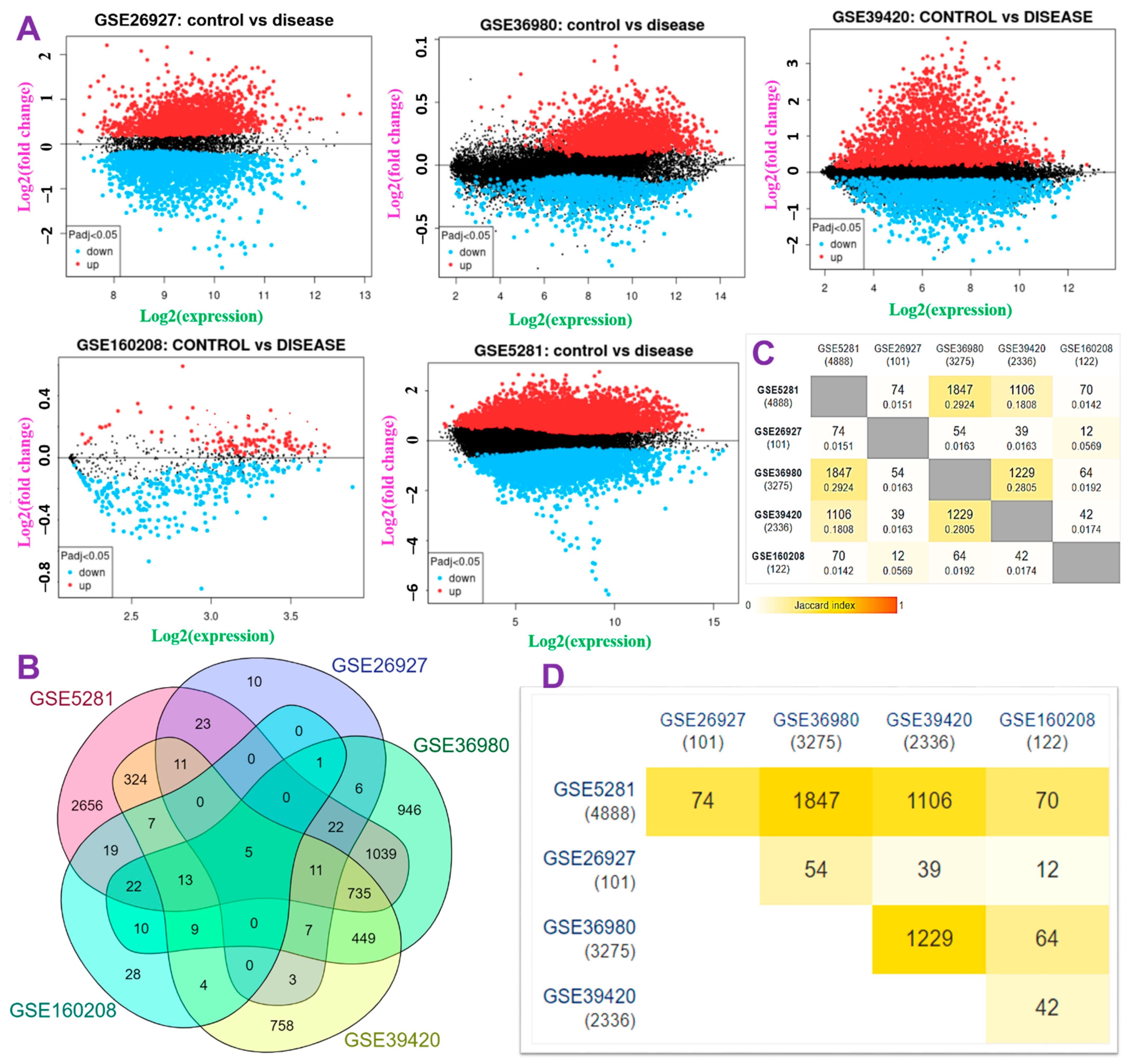
2.2. Identification of Differentially Expressed Genes (DEGs)
2.3. Brain-Specific Gene Co-Expression, Protein–Protein Interaction (PPI) Networks, and Gene-Set-Enrichment Analysis (GSEA) of DEGs
2.4. Analysis of Gene Disease-Specific Associations of the DEGs
2.5. MicroRNA (miRNA) Regulatory Network Analysis of the DEGs
2.6. In Silico Evaluation of the Drug-Likeness, Pharmacokinetics (PKs), Blood–Brain Barrier (BBB) Permeability and Acute Rat Toxicity Study of Antrocin
2.7. Molecular Docking Studies
3. Results
3.1. Deregulated Expressions of ATP6V1A, BNIP3, CAMK4, TIPRL, and TOMM70 Associated with the Pathology of Neurodegenerative Dementia
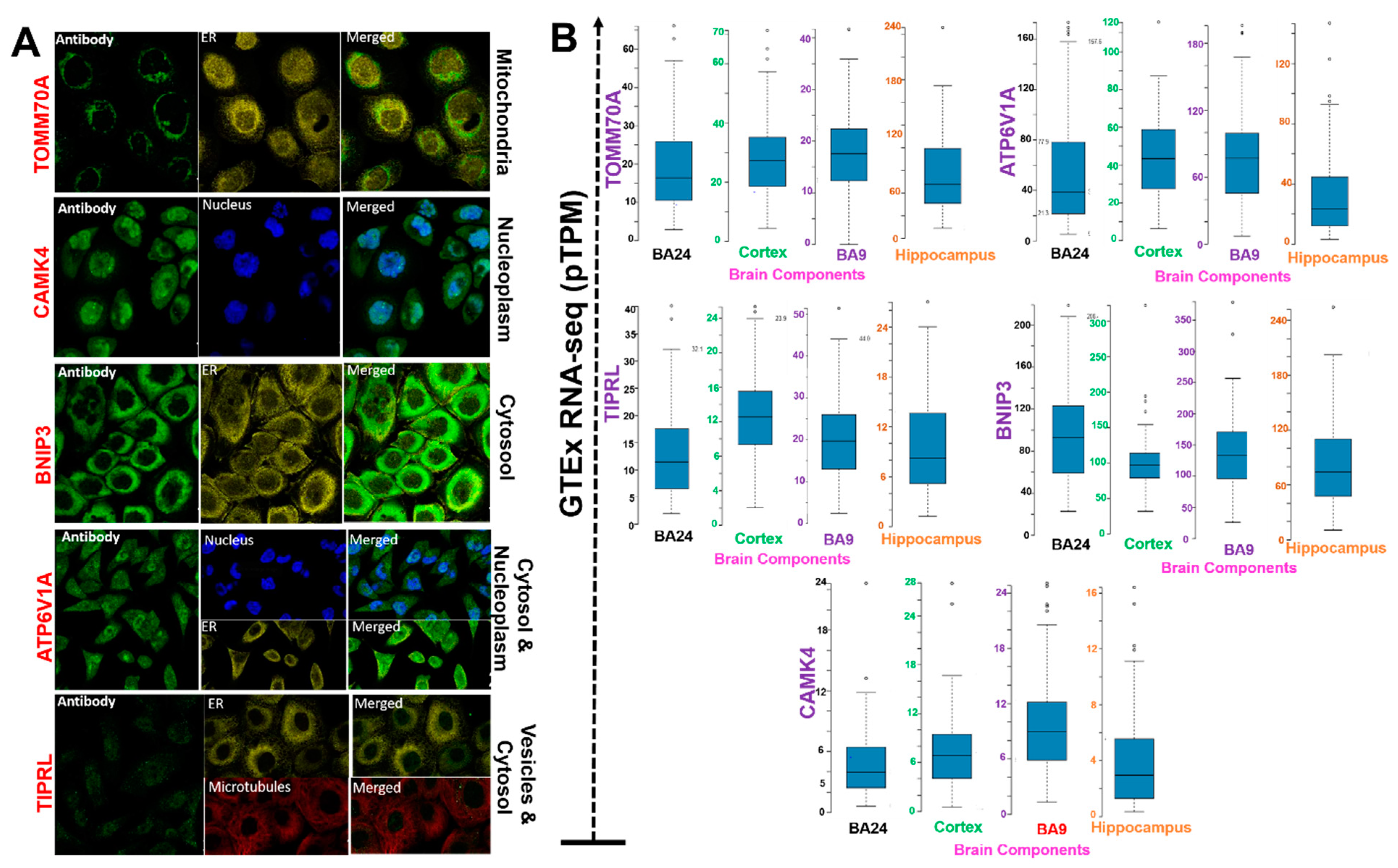
3.2. ATP6V1A, BNIP3, CAMK4, TIPRL, and TOMM70 Localization and Differential Expressions in Brain Regions
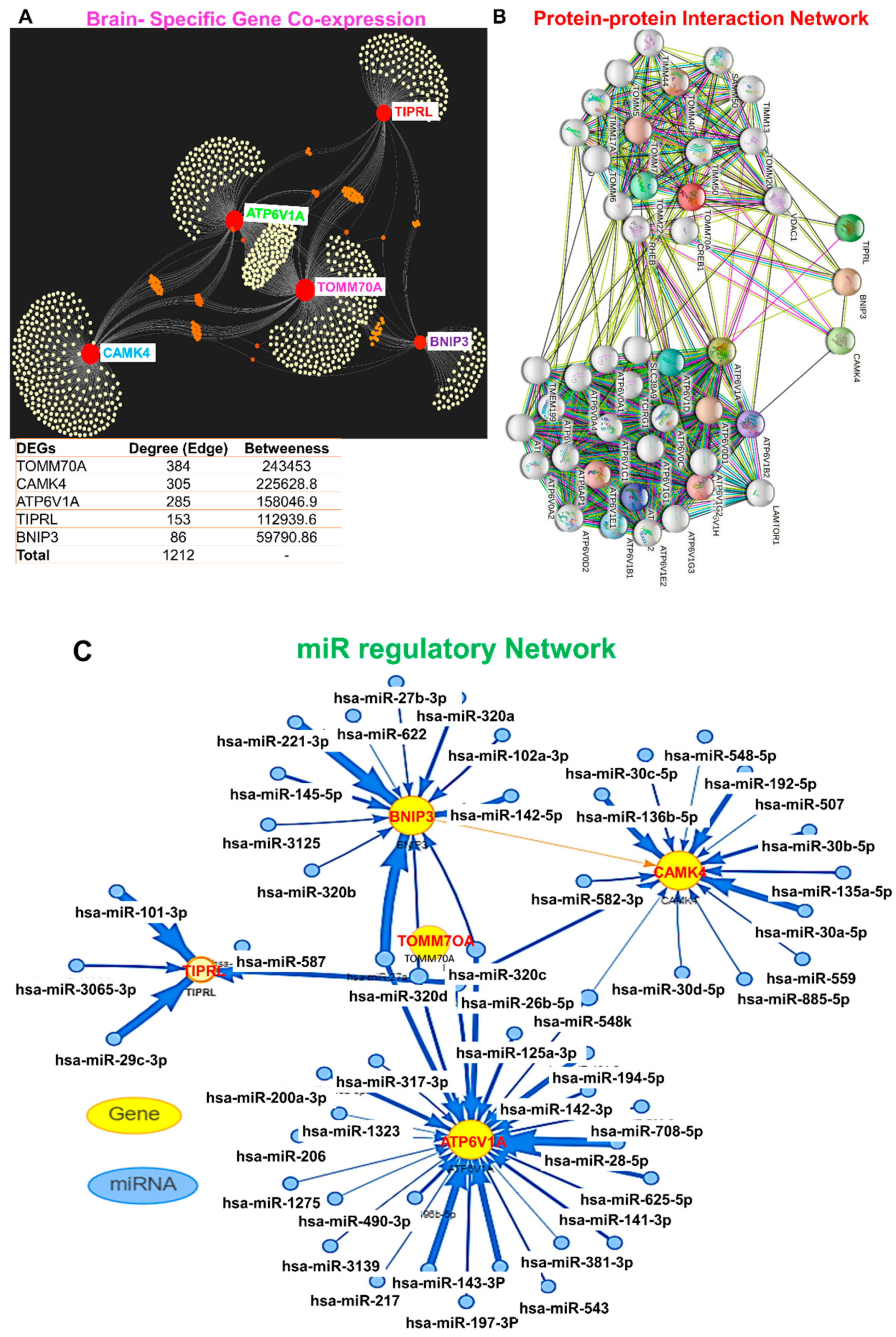
3.3. MicroRNA (miR) Regulatory Network and Brain-Specific Gene Interactions of ATP6V1A, BNIP3, CAMK4, TIPRL, and TOMM70
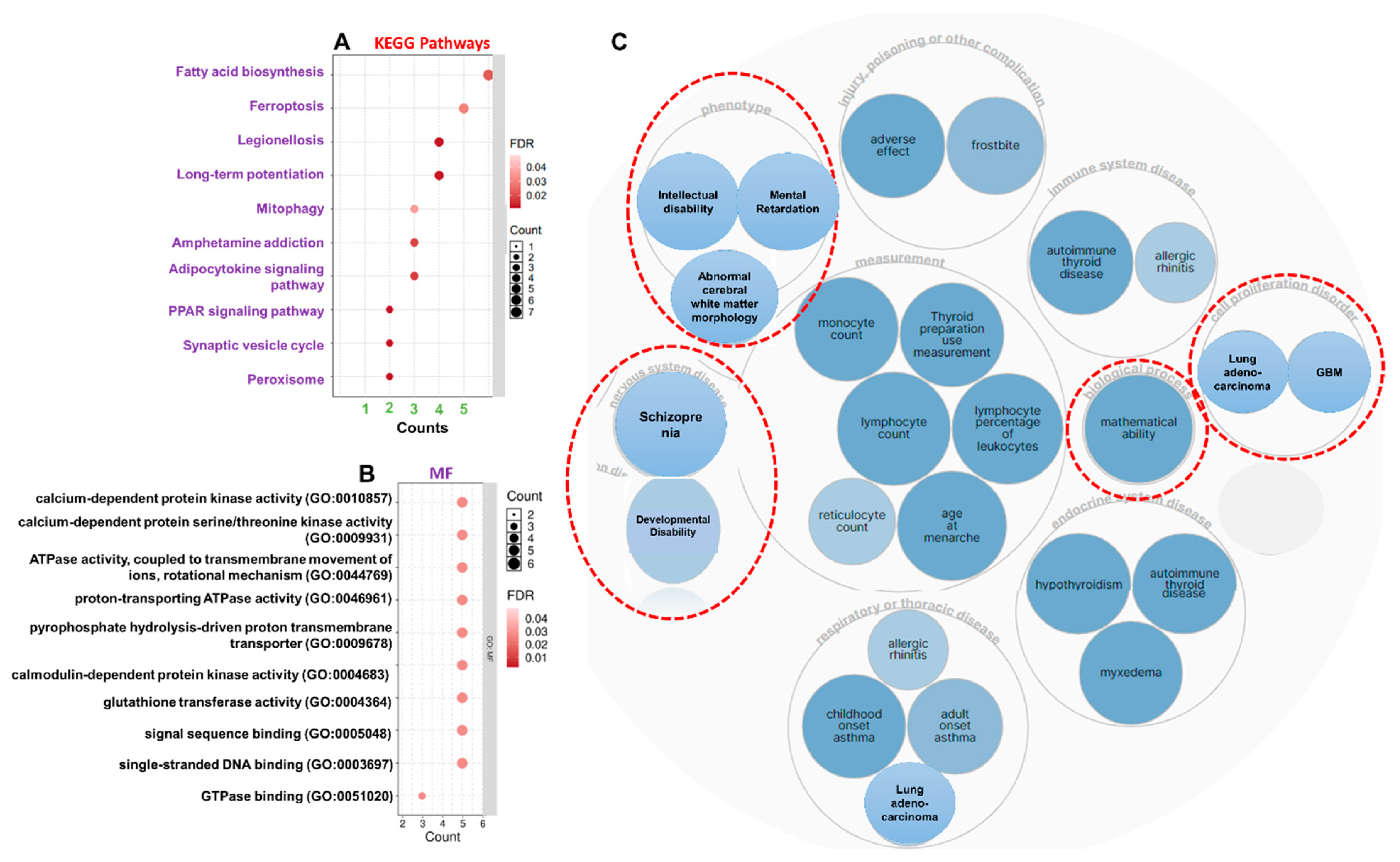
3.4. ATP6V1A/BNIP3 and CAMK4/TIPRL/TOMM70 Are Associated with Mitochondrial Dysfunction, Inflammatory Processes, and Various Pathways Involved in AD Pathogenesis
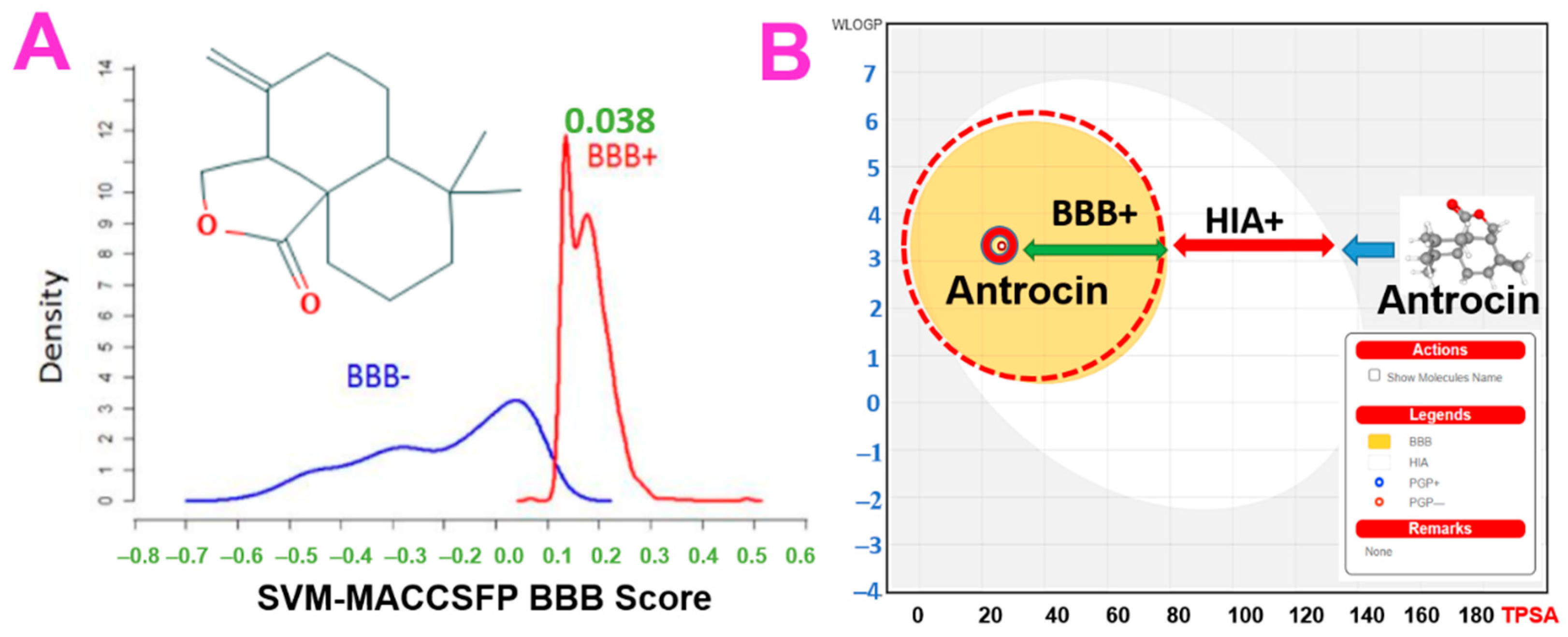
3.5. In Silico Pharmacokinetics, BBB Permeability and Acute Toxicity of Antrocin
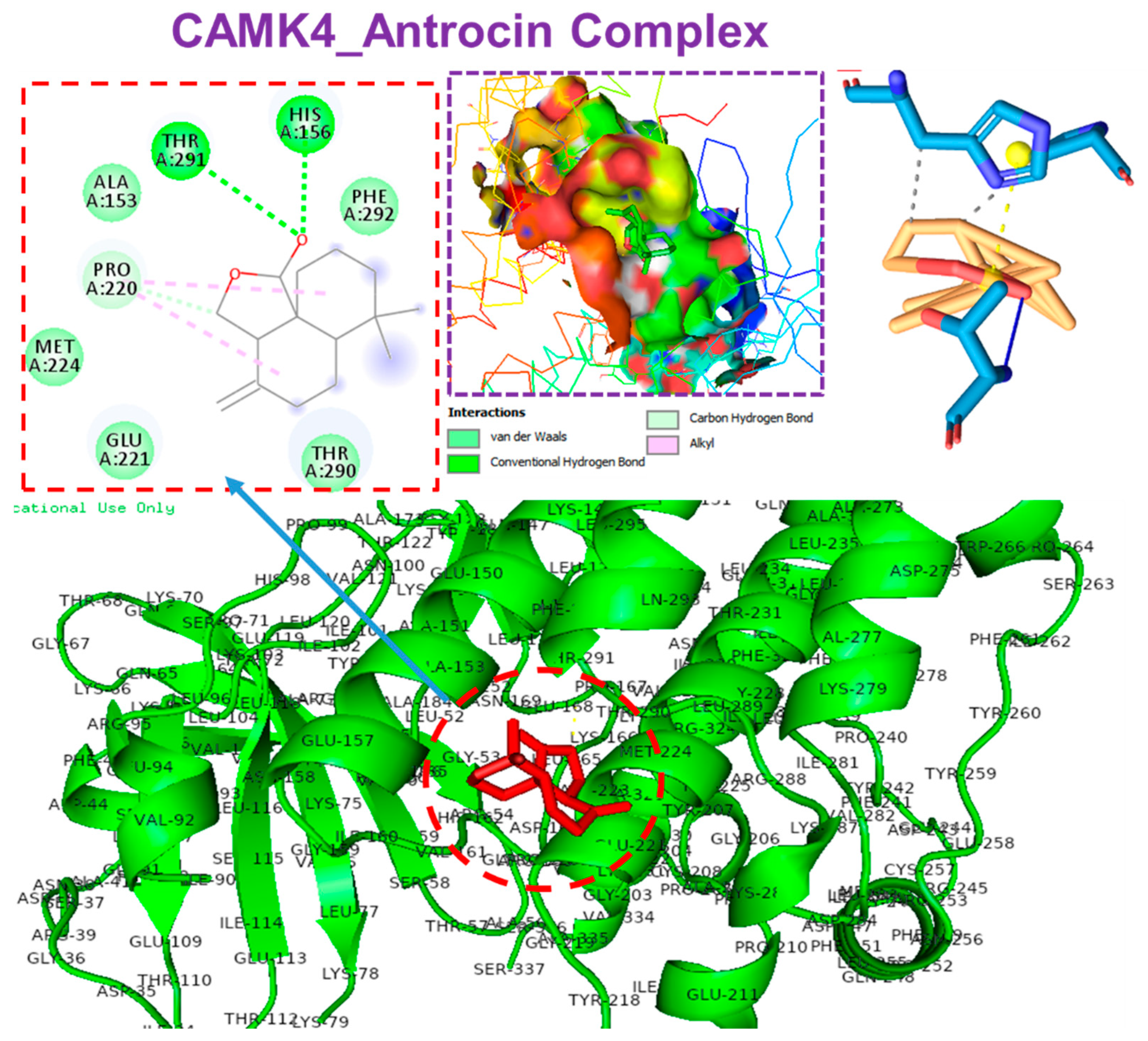
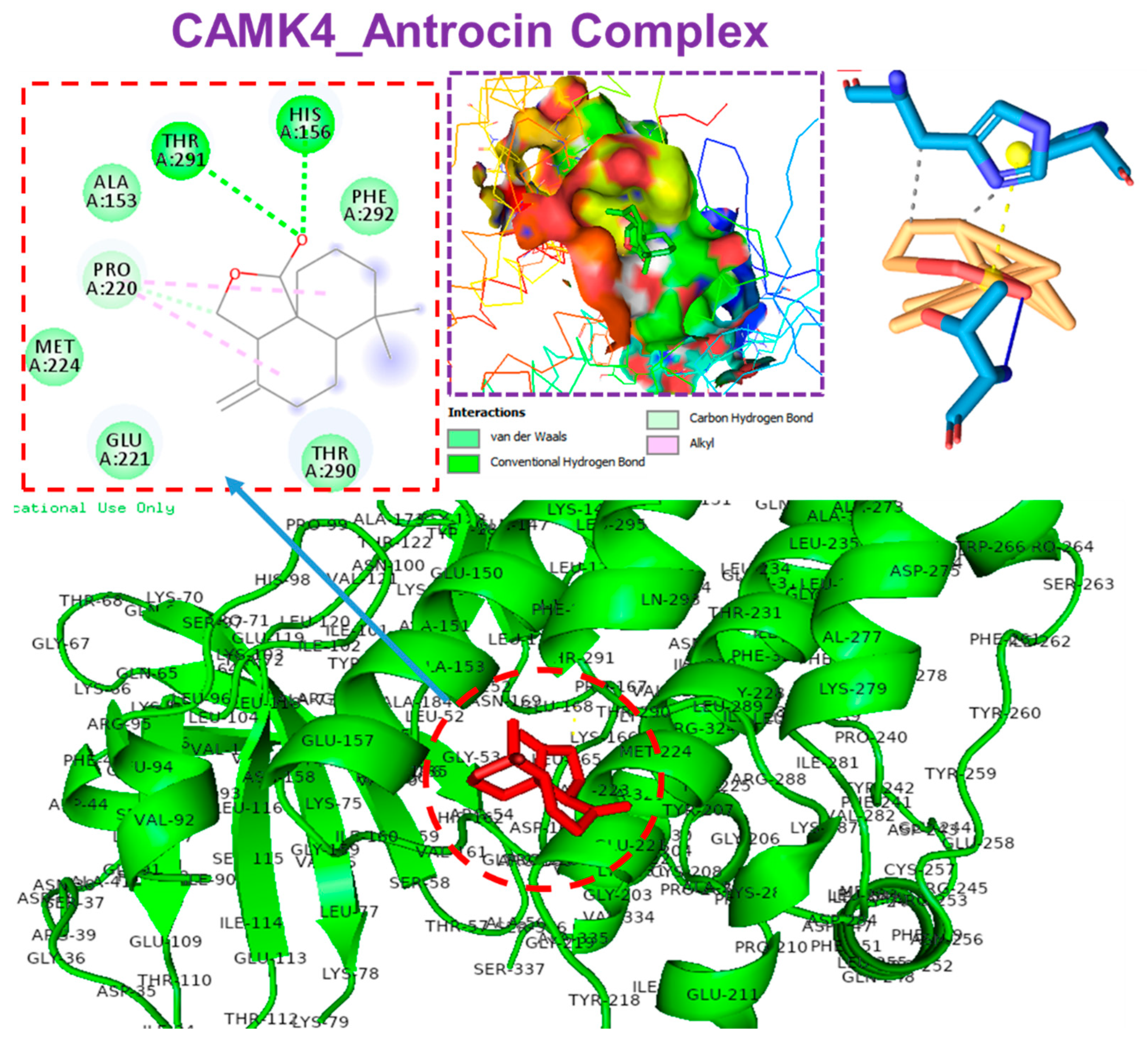
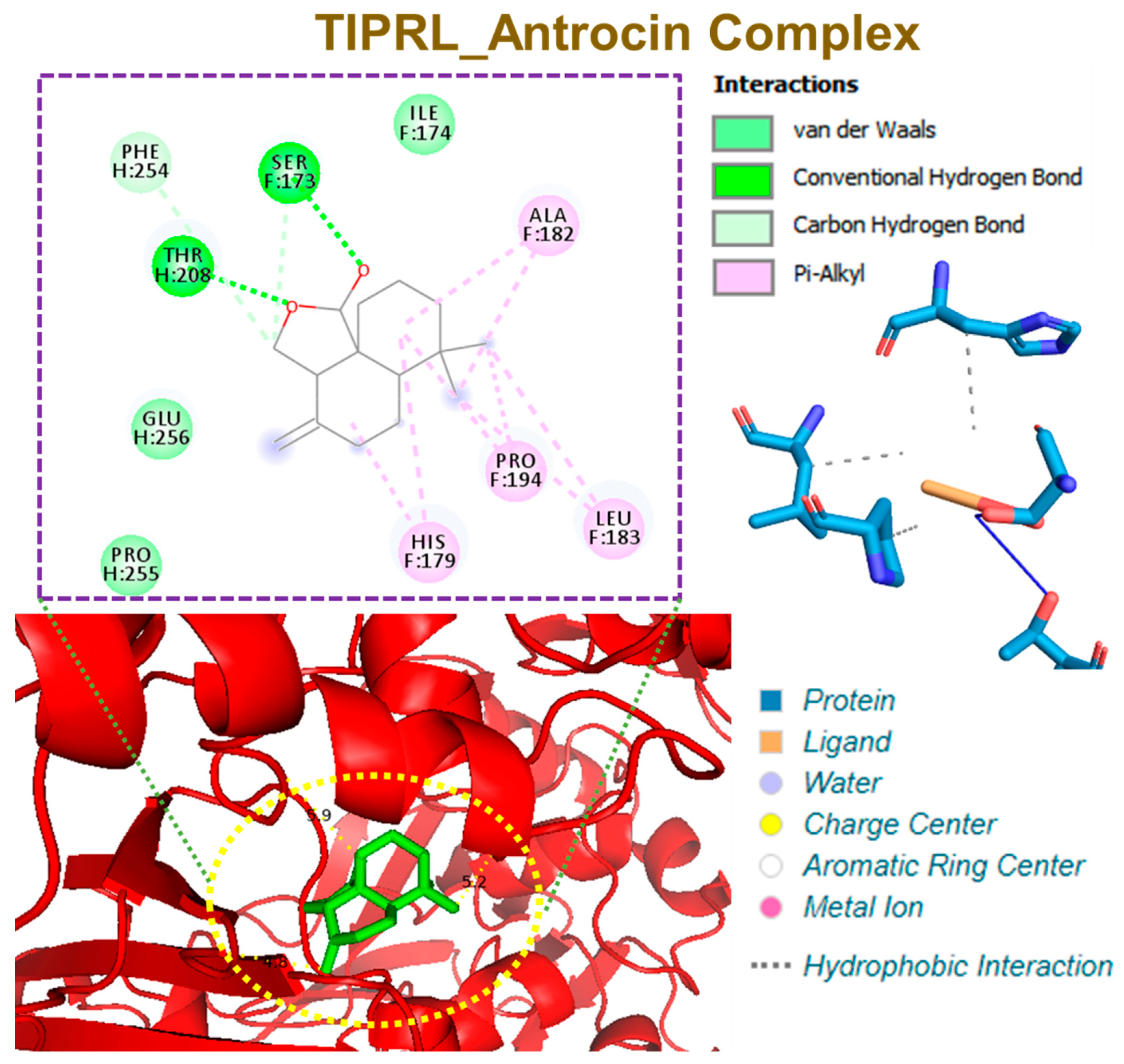
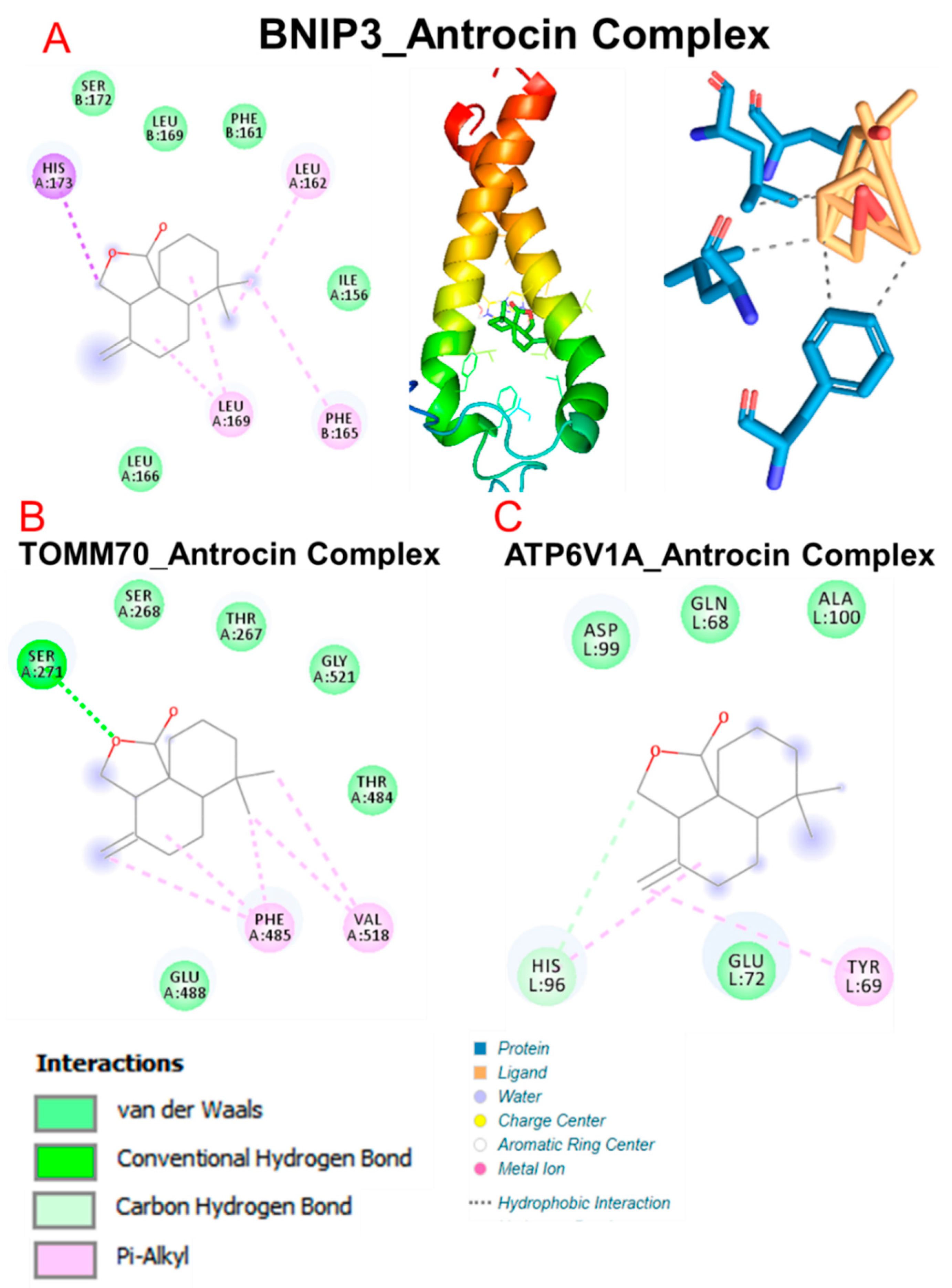
3.6. Molecular Docking Profiles of Antrocin with ATP6V1A, BNIP3, CAMK4, TIPRL, and TOMM70
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fleming, R.; Zeisel, J.; Bennett, K. World Alzheimer Report: Design Dignity Dementia: Dementia Related Design and the Built Environment; Alzheimer’s Disease International: London, UK, 2020; Volume 1. [Google Scholar]
- Evans, D.A.; Funkenstein, H.H.; Albert, M.S.; Scherr, P.A.; Cook, N.R.; Chown, M.J.; Hebert, L.E.; Hennekens, C.H.; Taylor, J.O. Prevalence of Alzheimer’s disease in a community population of older persons: Higher than previously reported. JAMA 1989, 262, 2551–2556. [Google Scholar] [CrossRef]
- Geldmacher, D.S.; Whitehouse, P.J. Evaluation of Dementia. N. Engl. J. Med. 1996, 335, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Girard, H.; Potvin, O.; Nugent, S.; Dallaire-Théroux, C.; Cunnane, S.; Duchesne, S. Faster progression from MCI to probable AD for carriers of a single-nucleotide polymorphism associated with type 2 diabetes. Neurobiol. Aging 2018, 64, 157.e111–157.e117. [Google Scholar] [CrossRef] [Green Version]
- Sabry, R.; El Sharkawy, R.E.; Gad, N.M. MiRNA -483-5p as a Potential Noninvasive Biomarker for Early Detection of Alzheimer’s Disease. Egypt. J. Immunol. 2020, 27, 59–72. [Google Scholar]
- Nestor, P.J.; Scheltens, P.; Hodges, J.R. Advances in the early detection of Alzheimer’s disease. Nat. Med. 2004, 10, S34–S41. [Google Scholar] [CrossRef]
- Mistur, R.; Mosconi, L.; De Santi, S.; Guzman, M.; Li, Y.; Tsui, W.; de Leon, M.J. Current challenges for the early detection of Alzheimer’s disease: Brain imaging and CSF studies. J. Clin. Neurol. 2009, 5, 153–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtzman, D.M.; Morris, J.C.; Goate, A.M. Alzheimer’s disease: The challenge of the second century. Sci. Transl. Med. 2011, 3, sr71–sr77. [Google Scholar] [CrossRef] [Green Version]
- Cedazo-Minguez, A.; Winblad, B. Biomarkers for Alzheimer’s disease and other forms of dementia: Clinical needs, limitations and future aspects. Exp. Gerontol. 2010, 45, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Beckman, D.; Morrison, J.H. Towards developing a rhesus monkey model of early Alzheimer’s disease focusing on women’s health. Am. J. Primatol. 2021, e23289. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhang, Y.; Xu, L.; Jin, S. Relationship Between Alzheimer’s Disease and the Immune System: A Meta-Analysis of Differentially Expressed Genes. Front. Neurosci. 2019, 12, 1026. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Sun, Y.Z.; Gao, X.H.; Qi, R.Q. Integrated bioinformatic analysis of differentially expressed genes and signaling pathways in plaque psoriasis. Mol. Med. Rep. 2019, 20, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Durrenberger, P.F.; Fernando, F.S.; Kashefi, S.N.; Bonnert, T.P.; Seilhean, D.; Nait-Oumesmar, B.; Schmitt, A.; Gebicke-Haerter, P.J.; Falkai, P.; Grünblatt, E.; et al. Common mechanisms in neurodegeneration and neuroinflammation: A BrainNet Europe gene expression microarray study. J. Neural. Transm (Vienna) 2015, 122, 1055–1068. [Google Scholar] [CrossRef] [PubMed]
- Areškevičiūtė, A.; Litman, T.; Broholm, H.; Melchior, L.C.; Nielsen, P.R.; Green, A.; Eriksen, J.O.; Smith, C.; Lund, E.L. Regional Differences in Neuroinflammation-Associated Gene Expression in the Brain of Sporadic Creutzfeldt-Jakob Disease Patients. Int. J. Mol. Sci. 2020, 22, 140. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.S.; Dunckley, T.; Beach, T.G.; Grover, A.; Mastroeni, D.; Ramsey, K.; Caselli, R.J.; Kukull, W.A.; McKeel, D.; Morris, J.C.; et al. Altered neuronal gene expression in brain regions differentially affected by Alzheimer’s disease: A reference data set. Physiol. Genom. 2008, 33, 240–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hokama, M.; Oka, S.; Leon, J.; Ninomiya, T.; Honda, H.; Sasaki, K.; Iwaki, T.; Ohara, T.; Sasaki, T.; LaFerla, F.M.; et al. Altered expression of diabetes-related genes in Alzheimer’s disease brains: The Hisayama study. Cereb. Cortex 2014, 24, 2476–2488. [Google Scholar] [CrossRef] [PubMed]
- Antonell, A.; Lladó, A.; Altirriba, J.; Botta-Orfila, T.; Balasa, M.; Fernández, M.; Ferrer, I.; Sánchez-Valle, R.; Molinuevo, J.L. A preliminary study of the whole-genome expression profile of sporadic and monogenic early-onset Alzheimer’s disease. Neurobiol. Aging 2013, 34, 1772–1778. [Google Scholar] [CrossRef] [PubMed]
- Bertram, L.; McQueen, M.B.; Mullin, K.; Blacker, D.; Tanzi, R.E. Systematic meta-analyses of Alzheimer disease genetic association studies: The AlzGene database. Nat. Genet. 2007, 39, 17–23. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Lawal, B.; Shittu, O.K.; Oibiokpa, F.I.; Berinyuy, E.B.; Mohammed, H. African natural products with potential antioxidants and hepatoprotectives properties: A review. Clin. Phytosci. 2017, 2, 1–66. [Google Scholar] [CrossRef] [Green Version]
- Geethangili, M.; Tzeng, Y.-M. Review of pharmacological effects of Antrodia camphorata and its bioactive compounds. Evid.-Based Complementary Altern. Med. 2011, 2011, 212641. [Google Scholar] [CrossRef] [Green Version]
- Geethangili, M.; Rao, Y.K.; Tzeng, Y.-M. Development and validation of a HPLC-DAD separation method for determination of bioactive antrocin in medicinal mushroom antrodia camphorata. Int. J. Appl. Sci. Eng. 2013, 11, 195–201. [Google Scholar]
- Chen, J.-H.; Wu, A.T.H.; Tzeng, D.T.W.; Huang, C.-C.; Tzeng, Y.-M.; Chao, T.-Y. Antrocin, a bioactive component from Antrodia cinnamomea, suppresses breast carcinogenesis and stemness via downregulation of β-catenin/Notch1/Akt signaling. Phytomedicine 2019, 52, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-T.; Huang, W.-C.; Rao, Y.K.; Ye, M.; Lee, W.-H.; Wang, L.-S.; Tzeng, D.T.; Wu, C.-H.; Shieh, Y.-S.; Huang, C.-Y.F. A sesquiterpene lactone antrocin from Antrodia camphorata negatively modulates JAK2/STAT3 signaling via microRNA let-7c and induces apoptosis in lung cancer cells. Carcinogenesis 2013, 34, 2918–2928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, Y.K.; Wu, A.T.; Geethangili, M.; Huang, M.-T.; Chao, W.-J.; Wu, C.-H.; Deng, W.-P.; Yeh, C.-T.; Tzeng, Y.-M. Identification of antrocin from Antrodia camphorata as a selective and novel class of small molecule inhibitor of Akt/mTOR signaling in metastatic breast cancer MDA-MB-231 cells. Chem. Res. Toxicol. 2011, 24, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-A.; Tzeng, D.T.W.; Huang, Y.-P.; Lin, C.-J.; Lo, U.-G.; Wu, C.-L.; Lin, H.; Hsieh, J.-T.; Tang, C.-H.; Lai, C.-H. Antrocin Sensitizes Prostate Cancer Cells to Radiotherapy through Inhibiting PI3K/AKT and MAPK Signaling Pathways. Cancers 2019, 11, 34. [Google Scholar] [CrossRef]
- Chiu, K.-Y.; Wu, C.-C.; Chia, C.-H.; Hsu, S.-L.; Tzeng, Y.-M. Inhibition of growth, migration and invasion of human bladder cancer cells by antrocin, a sesquiterpene lactone isolated from Antrodia cinnamomea, and its molecular mechanisms. Cancer Lett. 2016, 373, 174–184. [Google Scholar] [CrossRef]
- Hainfellner, J.A.; Wanschitz, J.; Jellinger, K.; Liberski, P.P.; Gullotta, F.; Budka, H. Coexistence of Alzheimer-type neuropathology in Creutzfeldt-Jakob disease. Acta Neuropathol. 1998, 96, 116–122. [Google Scholar] [CrossRef]
- Calero, O.; Bullido, M.J.; Clarimón, J.; Frank-García, A.; Martínez-Martín, P.; Lleó, A.; Rey, M.J.; Rábano, A.; Blesa, R.; Gómez-Isla, T.; et al. Genetic cross-interaction between APOE and PRNP in sporadic Alzheimer’s and Creutzfeldt-Jakob diseases. PLoS ONE 2011, 6, e22090. [Google Scholar] [CrossRef] [Green Version]
- Dorey, A.; Tholance, Y.; Vighetto, A.; Perret-Liaudet, A.; Lachman, I.; Krolak-Salmon, P.; Wagner, U.; Struyfs, H.; De Deyn, P.P.; El-Moualij, B.; et al. Association of Cerebrospinal Fluid Prion Protein Levels and the Distinction Between Alzheimer Disease and Creutzfeldt-Jakob Disease. JAMA Neurol. 2015, 72, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.W.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvaraj, G.; Kaliamurthi, S.; Kaushik, A.C.; Khan, A.; Wei, Y.-K.; Cho, W.C.; Gu, K.; Wei, D.-Q. Identification of target gene and prognostic evaluation for lung adenocarcinoma using gene expression meta-analysis, network analysis and neural network algorithms. J. Biomed. Inform. 2018, 86, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Wei, Y.-K.; Kaliamurthi, S.; Cao, Y.; Nangraj, A.S.; Sui, X.; Chu, D.; Wang, H.; Wei, D.-Q.; Peslherbe, G.H.; et al. Circulating miR-1246 Targeting UBE2C, TNNI3, TRAIP, UCHL1 Genes and Key Pathways as a Potential Biomarker for Lung Adenocarcinoma: Integrated Biological Network Analysis. J. Pers. Med. 2020, 10, 162. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Carvalho-Silva, D.; Pierleoni, A.; Pignatelli, M.; Ong, C.; Fumis, L.; Karamanis, N.; Carmona, M.; Faulconbridge, A.; Hercules, A.; McAuley, E.; et al. Open Targets Platform: New developments and updates two years on. Nucleic Acids Res. 2019, 47, D1056–D1065. [Google Scholar] [CrossRef]
- Kern, F.; Fehlmann, T.; Solomon, J.; Schwed, L.; Grammes, N.; Backes, C.; Van Keuren-Jensen, K.; Craig, D.W.; Meese, E.; Keller, A. miEAA 2.0: Integrating multi-species microRNA enrichment analysis and workflow management systems. Nucleic Acids Res. 2020, 48, W521–W528. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Poroikov, V.V.; Filimonov, D.A.; Gloriozova, T.A.; Lagunin, A.A.; Druzhilovskiy, D.S.; Rudik, A.V.; Stolbov, L.A.; Dmitriev, A.V.; Tarasova, O.A.; Ivanov, S.M.; et al. Computer-aided prediction of biological activity spectra for organic compounds: The possibilities and limitations. Russ. Chem. Bull. 2019, 68, 2143–2154. [Google Scholar] [CrossRef]
- Liu, H.; Wang, L.; Lv, M.; Pei, R.; Li, P.; Pei, Z.; Wang, Y.; Su, W.; Xie, X.-Q. AlzPlatform: An Alzheimer’s Disease Domain-Specific Chemogenomics Knowledgebase for Polypharmacology and Target Identification Research. J. Chem. Inf. Modeling 2014, 54, 1050–1060. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Zoete, V. A BOILED-Egg To Predict Gastrointestinal Absorption and Brain Penetration of Small Molecules. ChemMedChem 2016, 11, 1117–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baell, J.B.; Holloway, G.A. New substructure filters for removal of pan assay interference compounds (PAINS) from screening libraries and for their exclusion in bioassays. J. Med. Chem. 2010, 53, 2719–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagunin, A.; Zakharov, A.; Filimonov, D.; Poroikov, V. QSAR Modelling of Rat Acute Toxicity on the Basis of PASS Prediction. Mol. Inform. 2011, 30, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Marcus, D.; Hanwell, D.E.C.; David, C.L.; Tim, V.; Eva, Z.; Geoffrey, R.H. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Lawal, B.; Liu, Y.-L.; Mokgautsi, N.; Khedkar, H.; Sumitra, M.R.; Wu, A.T.H.; Huang, H.-S. Pharmacoinformatics and Preclinical Studies of NSC765690 and NSC765599, Potential STAT3/CDK2/4/6 Inhibitors with Antitumor Activities against NCI60 Human Tumor Cell Lines. Biomedicines 2021, 9, 92. [Google Scholar] [CrossRef]
- Lawal, B.; Lee, C.-Y.; Mokgautsi, N.; Sumitra, M.R.; Khedkar, H.; Wu, A.T.H.; Huang, H.-S. mTOR/EGFR/iNOS/MAP2K1/FGFR/TGFB1 Are Druggable Candidates for N-(2,4-Difluorophenyl)-2′,4′-Difluoro-4-Hydroxybiphenyl-3-Carboxamide (NSC765598), With Consequent Anticancer Implications. Front. Oncol. 2021, 11, 656738. [Google Scholar] [CrossRef]
- Visualizer, D.S. BIOVIA, Dassault Systèmes, BIOVIA Workbook; Release 2020; BIOVIA Pipeline Pilot, Release 2020; Dassault Systèmes: San Diego, CA, USA, 2020. [Google Scholar]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein-ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef]
- Lawal, B.; Kuo, Y.-C.; Sumitra, M.R.; Wu, A.T.; Huang, H.-S. In vivo Pharmacokinetic and Anticancer Studies of HH-N25, a Selective Inhibitor of Topoisomerase I, and Hormonal Signaling for Treating Breast Cancer. J. Inflamm. Res. 2021, 14, 1–13. [Google Scholar] [CrossRef]
- Brookmeyer, R.; Johnson, E.; Ziegler-Graham, K.; Arrighi, H.M. Forecasting the global burden of Alzheimer’s disease. Alzheimers Dement. 2007, 3, 186–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takizawa, C.; Thompson, P.L.; van Walsem, A.; Faure, C.; Maier, W.C. Epidemiological and economic burden of Alzheimer’s disease: A systematic literature review of data across Europe and the United States of America. J. Alzheimers Dis. 2015, 43, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Choy, K.H.C.; Marriott, P.J.; Chai, S.Y.; Scanlon, M.J.; Porter, C.J.H.; Short, J.L.; Nicolazzo, J.A. Reduced blood-brain barrier expression of fatty acid-binding protein 5 is associated with increased vulnerability of APP/PS1 mice to cognitive deficits from low omega-3 fatty acid diets. J. Neurochem. 2018, 144, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Zhang, Y.; Shen, Y.; Wang, Y.; Zhao, M.; Sun, L. The Potential Role of Ferroptosis in Alzheimer’s Disease. J. Alzheimers Dis. 2021, 80, 907–925. [Google Scholar] [CrossRef] [PubMed]
- Prieto, G.A.; Trieu, B.H.; Dang, C.T.; Bilousova, T.; Gylys, K.H.; Berchtold, N.C.; Lynch, G.; Cotman, C.W. Pharmacological Rescue of Long-Term Potentiation in Alzheimer Diseased Synapses. J. Neurosci. 2017, 37, 1197–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo-García, L.E.; Tendilla-Beltrán, H.; Vázquez-Roque, R.A.; Jurado-Tapia, E.E.; Díaz, A.; Aguilar-Alonso, P.; Brambila, E.; Monjaraz, E.; De La Cruz, F.; Rodríguez-Moreno, A.; et al. Amphetamine sensitization alters hippocampal neuronal morphology and memory and learning behaviors. Mol. Psychiatry 2020. [Google Scholar] [CrossRef]
- Hu, Y.-S.; Xin, J.; Hu, Y.; Zhang, L.; Wang, J. Analyzing the genes related to Alzheimer’s disease via a network and pathway-based approach. Alzheimers Res. Ther. 2017, 9, 29. [Google Scholar] [CrossRef]
- Akiyama, H.; Barger, S.; Barnum, S.; Bradt, B.; Bauer, J.; Cole, G.M.; Cooper, N.R.; Eikelenboom, P.; Emmerling, M.; Fiebich, B.L. Inflammation and Alzheimer’s disease. Neurobiol. Aging 2000, 21, 383–421. [Google Scholar] [CrossRef]
- Tuppo, E.E.; Arias, H.R. The role of inflammation in Alzheimer’s disease. Int. J. Biochem. Cell Biol. 2005, 37, 289–305. [Google Scholar] [CrossRef]
- Griffin, W.; Sheng, J.; Royston, M.; Gentleman, S.; McKenzie, J.; Graham, D.; Roberts, G.; Mrak, R. Glial-neuronal interactions in Alzheimer’s disease: The potential role of a ‘cytokine cycle’in disease progression. Brain Pathol. 1998, 8, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Carelli, V.; Musumeci, O.; Caporali, L.; Zanna, C.; La Morgia, C.; Del Dotto, V.; Porcelli, A.M.; Rugolo, M.; Valentino, M.L.; Iommarini, L.; et al. Syndromic parkinsonism and dementia associated with OPA1 missense mutations. Ann. Neurol. 2015, 78, 21–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan Zhang, S.Z.; Siman, Y.; Yuting, W.; Jingjing, W.; Jinlan, H.; Dengpan, W.; Zhenguo, Z. Identification of Potential Therapeutic Targets of Alzheimer’s Disease By Weighted Gene Co-Expression Network Analysis. Chin. Med. Sci. J. 2020, 35, 330–341. [Google Scholar] [CrossRef]
- Fassio, A.; Esposito, A.; Kato, M.; Saitsu, H.; Mei, D.; Marini, C.; Conti, V.; Nakashima, M.; Okamoto, N.; Olmez Turker, A.; et al. De novo mutations of the ATP6V1A gene cause developmental encephalopathy with epilepsy. Brain 2018, 141, 1703–1718. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.S.; Anderson, A.J.; Stojanovski, D. Mitochondrial protein import dysfunction: Mitochondrial disease, neurodegenerative disease and cancer. FEBS Lett. 2021, 595, 1107–1131. [Google Scholar] [CrossRef]
- Shafqat, N.; Kavanagh, K.L.; Sass, J.O.; Christensen, E.; Fukao, T.; Lee, W.H.; Oppermann, U.; Yue, W.W. A structural mapping of mutations causing succinyl-CoA:3-ketoacid CoA transferase (SCOT) deficiency. J. Inherit. Metab. Dis. 2013, 36, 983–987. [Google Scholar] [CrossRef] [Green Version]
- Kitchen, D.B.; Decornez, H.; Furr, J.R.; Bajorath, J. Docking and scoring in virtual screening for drug discovery: Methods and applications. Nat. Rev. Drug Discov. 2004, 3, 935–949. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-H.; Wu, A.T.H.; Lawal, B.; Tzeng, D.T.W.; Lee, J.-C.; Ho, C.-L.; Chao, T.-Y. Identification of Cancer Hub Gene Signatures Associated with Immune-Suppressive Tumor Microenvironment and Ovatodiolide as a Potential Cancer Immunotherapeutic Agent. Cancers 2021, 13, 3847. [Google Scholar] [CrossRef]
- Meng, X.-Y.; Zhang, H.-X.; Mezei, M.; Cui, M. Molecular docking: A powerful approach for structure-based drug discovery. Curr. Comput.-Aided Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef]
- Jorgensen, W.L. The many roles of computation in drug discovery. Science 2004, 303, 1813–1818. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Huang, D. Hydrogen bonding penalty upon ligand binding. PLoS ONE 2011, 6, e19923. [Google Scholar] [CrossRef]
- Lawal, B.; Lo, W.-C.; Mokgautsi, N.; Sumitra, M.R.; Khedkar, H.; Wu, A.T.; Huang, H.-S. A preclinical report of a cobimetinib-inspired novel anticancer small-molecule scaffold of isoflavones, NSC777213, for targeting PI3K/AKT/mTOR/MEK in multiple cancers. Am. J. Cancer Res. 2021, 11, 2590–2617. [Google Scholar] [PubMed]
- Arthur, D.E.; Uzairu, A. Molecular docking studies on the interaction of NCI anticancer analogues with human Phosphatidylinositol 4,5-bisphosphate 3-kinase catalytic subunit. J. King Saud Univ. Sci. 2019, 31, 1151–1166. [Google Scholar] [CrossRef]
- Yang, H.; Wang, W.; Jia, L.; Qin, W.; Hou, T.; Wu, Q.; Li, H.; Tian, Y.; Jia, J. The Effect of Chronic Cerebral Hypoperfusion on Blood-Brain Barrier Permeability in a Transgenic Alzheimer’s Disease Mouse Model (PS1V97L). J. Alzheimers Dis. 2020, 74, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Brain Drug Targeting: The Future of Brain Drug Development; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Cohen, B.; Bangham, A. Diffusion of small non-electrolytes across liposome membranes. Nature 1972, 236, 173–174. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.; Gottschlich, R.; Seelig, A. Blood-brain barrier permeation: Molecular parameters governing passive diffusion. J. Membr. Biol. 1998, 165, 201–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldmann, C.; Bajorath, J. Machine learning reveals that structural features distinguishing promiscuous and non-promiscuous compounds depend on target combinations. Sci. Rep. 2021, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Ji, Y.; Youn, K.; Lim, G.; Lee, J.; Kim, D.H.; Jun, M. Baicalein as a Potential Inhibitor against BACE1 and AChE: Mechanistic Comprehension through In Vitro and Computational Approaches. Nutrients 2019, 11, 2694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chedik, L.; Mias-Lucquin, D.; Bruyere, A.; Fardel, O. In Silico Prediction for Intestinal Absorption and Brain Penetration of Chemical Pesticides in Humans. Int. J. Environ. Res. Public Health 2017, 14, 708. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Datasets | Platform | No. of Cases | No. of Controls | Total No. | Mean Age (Years) |
|---|---|---|---|---|---|
| GSE5281 | GPL570 | 87 | 74 | 161 | 79.8 0 ± 9.10 |
| GSE160208 | GPL29311 | 27 | 20 | 47 | NA |
| GSE26927 | GPL6255 | 63 | 56 | 119 | 63.65 ± 10.83 |
| GSE36980 | GPL6244 | 33 | 47 | 80 | NA |
| GSE39420 | GPL11532 | 14 | 7 | 21 | 55.66 ± 1.93 |
| GTEx RNA-seq (pTPM) | |||||
|---|---|---|---|---|---|
| Brain Region | TIPRL | TOMM70A | CAMK4 | ATPV1A | BNIPS |
| Anterior cingulate cortex (BA24) | 12.90 ± 7.90 b | 19.10 ± 11.70 b | 4.70 ± 3.30 a | 53.40 ± 42.60 b | 98.90 ± 45.70 b |
| Cortex (central) | 12.30 ± 4.50 b | 19.20 ± 7.20 b | 7.50 ± 4.00 b | 43.81 ± 20.00 b | 98.63 ± 33.20 b |
| Frontal cortex (BA9) | 19.90 ± 9.80 c | 27.80 ± 13.10 c | 9.30 ± 5.20 b | 77.60 ± 42.50 b | 136.72 ± 55.03 b |
| Hippocampus | 9.70 ± 5.70 a | 14.00 ± 8.20 a | 4.00 ± 3.30 a | 31.70 ± 27.50 a | 80.43 ± 45.05 a |
| Index | Name | p-Value | Adjusted p-Value | Odds Ratio | Combined Score |
|---|---|---|---|---|---|
| 1 | catabolism of mitochondrial proteins (GO:0035694) | 0.003296 | 0.04581 | 399.68 | 2284.21 |
| 2 | response to increased oxygen levels (GO:0036296) | 0.003844 | 0.04581 | 333.05 | 1852.16 |
| 3 | regulation of myeloid leukocyte differentiation (GO:0002761) | 0.004392 | 0.04581 | 285.46 | 1549.44 |
| 4 | negative regulation of mitochondrial fusion (GO:0010637) | 0.004392 | 0.04581 | 285.46 | 1549.44 |
| 5 | positive regulation of dendritic cell cytokine production (GO:0002732) | 0.004392 | 0.04581 | 285.46 | 1549.44 |
| 6 | mitochondrial fragmentation involved in apoptotic process (GO:0043653) | 0.004940 | 0.04581 | 249.76 | 1326.33 |
| 7 | negative regulation of membrane potential (GO:0045837) | 0.004940 | 0.04581 | 249.76 | 1326.33 |
| 8 | positive regulation of mitochondrial membrane permeability involved in apoptotic process (GO:1902110) | 0.004940 | 0.04581 | 249.76 | 1326.33 |
| 9 | cellular response to oxygen levels (GO:0071453) | 0.005488 | 0.04581 | 222.00 | 1155.57 |
| 10 | myeloid dendritic cell differentiation (GO:0043011) | 0.005488 | 0.04581 | 222.00 | 1155.57 |
| LD50 | |||
| Administration Route | Log10(mmol/kg) | mg/kg | OECD classification |
| Intraperitoneal (i.p) | 0.421 | 618 | Class 5 |
| Intravenous (i.v) | −0.955 | 26 | Class 3 |
| Oral gavage (o.p) | 1.536 | 804.3 | Non-toxic |
| Subcutaneous (s.c) | 0.344 | 517.3 | Class 4 |
| Environmental Toxicity | |||
| Bioaccumulation factor Log10(BCF) | 1.521 | ||
| Daphnia magna LC50-Log10(mol/L) | 4.594 | ||
| Fathead Minnow LC50 Log10(mmol/L) | −1.648 | ||
| Tetrahymena pyriformis IGC50-Log10(mol/L) | 0.856 | ||
| Properties | Antrocin | Reference Value |
|---|---|---|
| Formula | C15H22O2 | − |
| M.W(g/mol) | 234.33 | 150−500 |
| R-bonds | 0 | 0−9 |
| H-bond ACC. | 2 | 0−10 |
| H-bond DON. | 0 | 0−5 |
| Molar Refractivity | 68.17 | 40 ~ 130 |
| TPSA (Ų) | 26.30 | 20−130 |
| Fraction Csp3 | 0.80 | 0.25 ~ < 1 |
| Log Po/w (XLOGP3) | 3.44 | −0.7 ~ 5 |
| Consensus Log Po/w | 3.31 | ≤3.5 |
| Drug-likeness (Lipinski rule) | Yes (0 violation) | MLOGP ≤ 4.15, M.W ≤ 500, H-bond ACC ≤ 10, H-bond DON ≤ 5 |
| Bioavailability Score | 0.55 | >0.1 (10%) |
| BBB-permeation (SVM_MACCSFP) | 0.038 | ≥0.02 |
| Synthetic accessibility | 4.18 | 1 (very easy) to 10 (very difficult). |
| PAINS | 0 alert | No alert |
| P-gp substrate | No | |
| CYP1A2 inhibitor | No | |
| CYP2C19 inhibitor | No | |
| CYP2D6 inhibitor | No | |
| CYP3A4 inhibitor | No | |
| Log Kp (skin permeation) | −5.29 cm/s | <−3.5 |
| Interaction | CAMK4 | BNIP3 | TIPRL | TOMM70 | ATP6V1A |
|---|---|---|---|---|---|
| ΔG = (kcal/mol) | −6.70 | 5.80 | −6.60 | −6.80 | −5.90 |
| Conventional H-bonds | THR291 (2.72 Å) HIS156 (1.92 Å) PRO220 (3.65 Å) | THR208 (2.03 Å) SER173 (2.97 Å) PHE254 (3.78 Å) | SER271 (2.40 Å) | HIS96 (2.16 Å) | |
| π-alkyl | PRO220 | LEU169 PHE165 LEU162 | ALA182 PRO194 LEU183 HIS179 | PHE485 VAL518 | TYR69 |
| π-sigma | HIS173 | ||||
| van der Waals forces | MET224 GLU221 THR290 PHE292 ALA153 | SER172 LEU169 PHE161 ILE156 LEU166 | PHE254 GLU256 PRO255 | SER268 THR267 GLY521 THR484 GLU488 | ASP99 GLN68 ALA100 GLU72 |
| Target | Amino Acid Residue | Ligand Atom | Protein Atom | Distance (Å) |
|---|---|---|---|---|
| BNIP3 | PHE165B | 868 | 628 | 3.69 |
| PHE165B | 863 | 629 | 3.66 | |
| LEU166A | 873 | 207 | 3.59 | |
| LEU169A | 869 | 231 | 3.35 | |
| LEU169A | 872 | 230 | 3.85 | |
| LEU169B | 868 | 661 | 3.40 | |
| TIPRL | HIS179F | 42301 | 19984 | 3.68 |
| LEU183F | 42299 | 20029 | 3.63 | |
| PRO194F | 42295 | 20139 | 3.72 | |
| TOMM70 | THR484A | 5020 | 3672 | 3.69 |
| PHE485A | 5014 | 3684 | 3.62 | |
| PHE485A | 5013 | 3686 | 3.51 | |
| PHE485A | 5020 | 3685 | 3.66 | |
| VAL518A | 5020 | 4029 | 3.53 | |
| ATP6V1A | TYR69L | 17251 | 3978 | 3.69 |
| GLU72L | 17251 | 4001 | 3.58 | |
| HIS96L | 17252 | 4191 | 3.64 | |
| CAMK4 | HIS156A | 2587 | 1013 | 3.90 |
| PRO220A | 2588 | 2588 | 3.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, A.T.H.; Lawal, B.; Wei, L.; Wen, Y.-T.; Tzeng, D.T.W.; Lo, W.-C. Multiomics Identification of Potential Targets for Alzheimer Disease and Antrocin as a Therapeutic Candidate. Pharmaceutics 2021, 13, 1555. https://doi.org/10.3390/pharmaceutics13101555
Wu ATH, Lawal B, Wei L, Wen Y-T, Tzeng DTW, Lo W-C. Multiomics Identification of Potential Targets for Alzheimer Disease and Antrocin as a Therapeutic Candidate. Pharmaceutics. 2021; 13(10):1555. https://doi.org/10.3390/pharmaceutics13101555
Chicago/Turabian StyleWu, Alexander T. H., Bashir Lawal, Li Wei, Ya-Ting Wen, David T. W. Tzeng, and Wen-Cheng Lo. 2021. "Multiomics Identification of Potential Targets for Alzheimer Disease and Antrocin as a Therapeutic Candidate" Pharmaceutics 13, no. 10: 1555. https://doi.org/10.3390/pharmaceutics13101555
APA StyleWu, A. T. H., Lawal, B., Wei, L., Wen, Y.-T., Tzeng, D. T. W., & Lo, W.-C. (2021). Multiomics Identification of Potential Targets for Alzheimer Disease and Antrocin as a Therapeutic Candidate. Pharmaceutics, 13(10), 1555. https://doi.org/10.3390/pharmaceutics13101555