
In Vitro Pharmacokinetic/Pharmacodynamic Modelling and Simulation of Amphotericin B against Candida auris

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Time-Kill Experiments
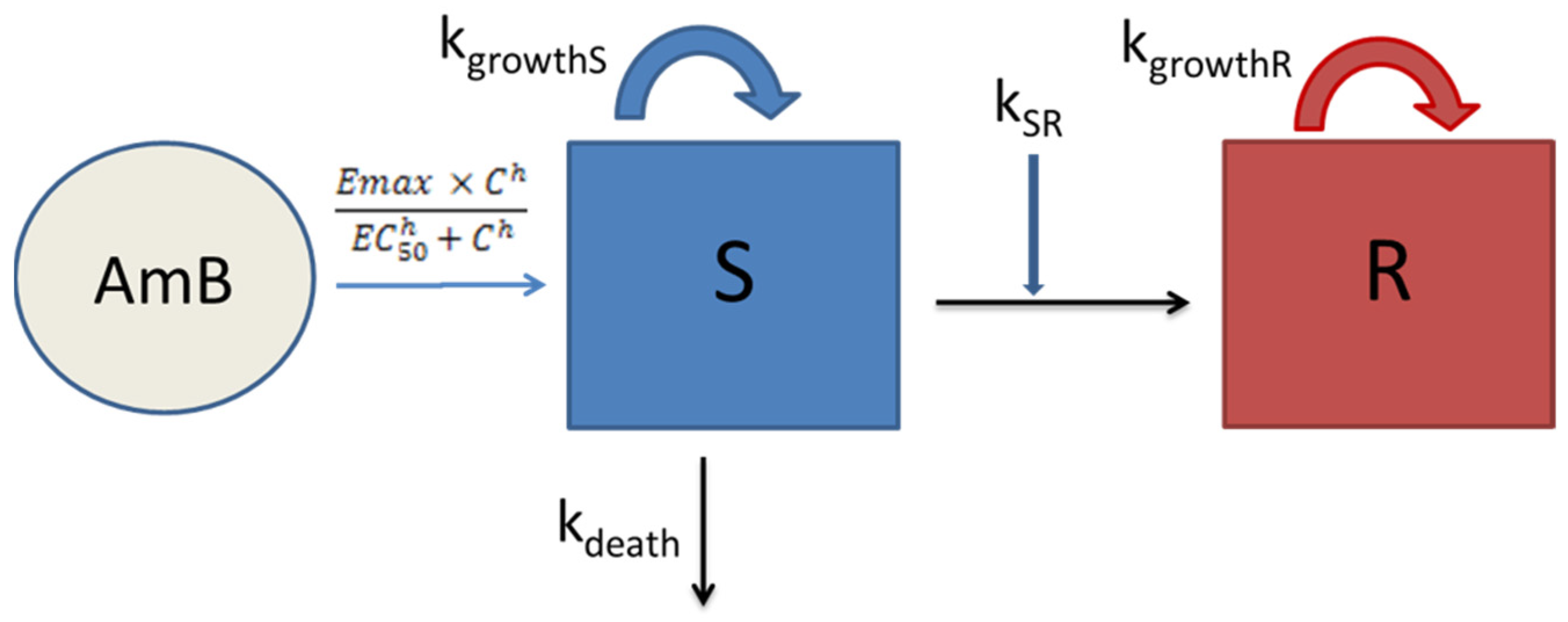
3.2. Semi-Mechanistic PK/PD Modelling
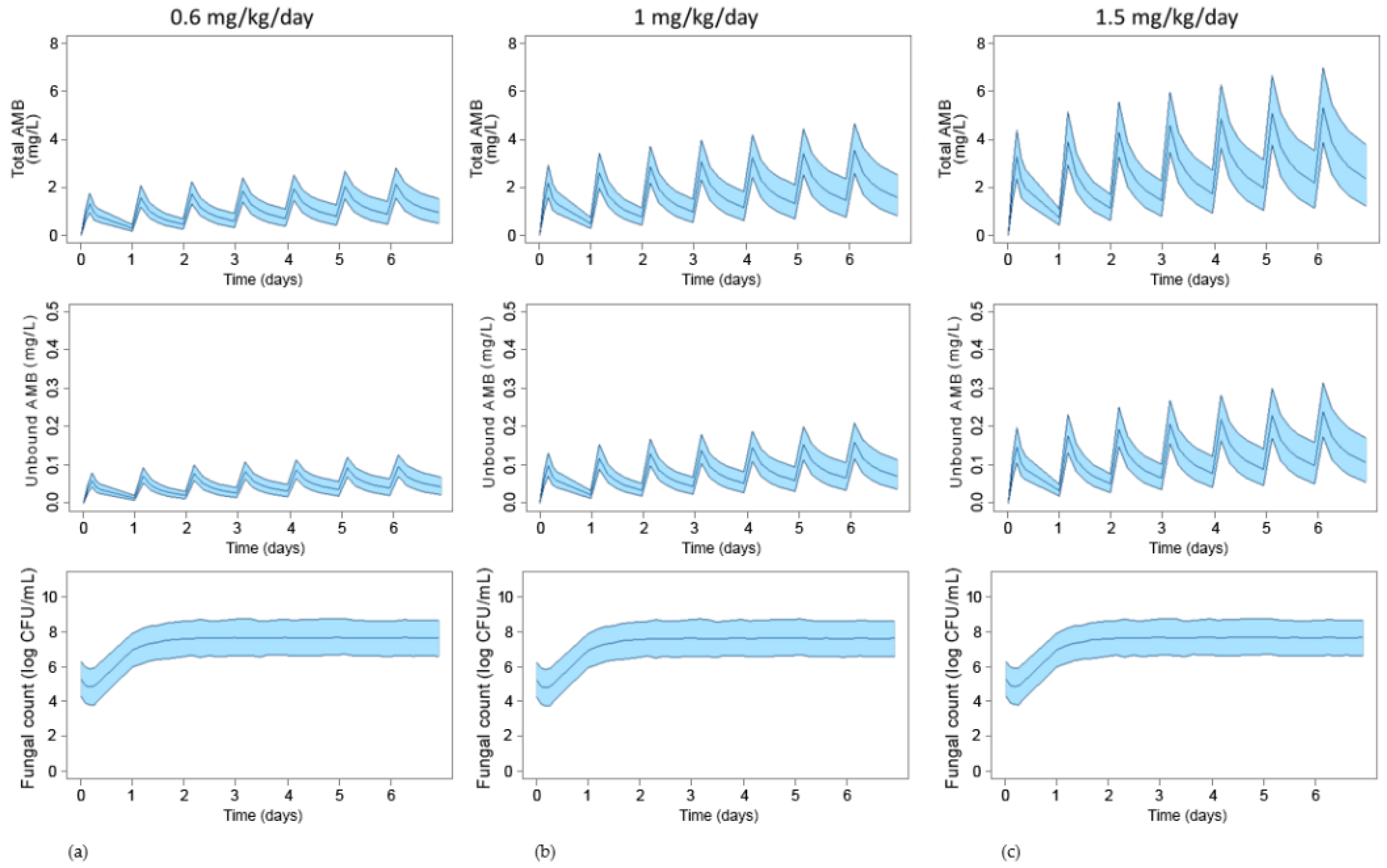
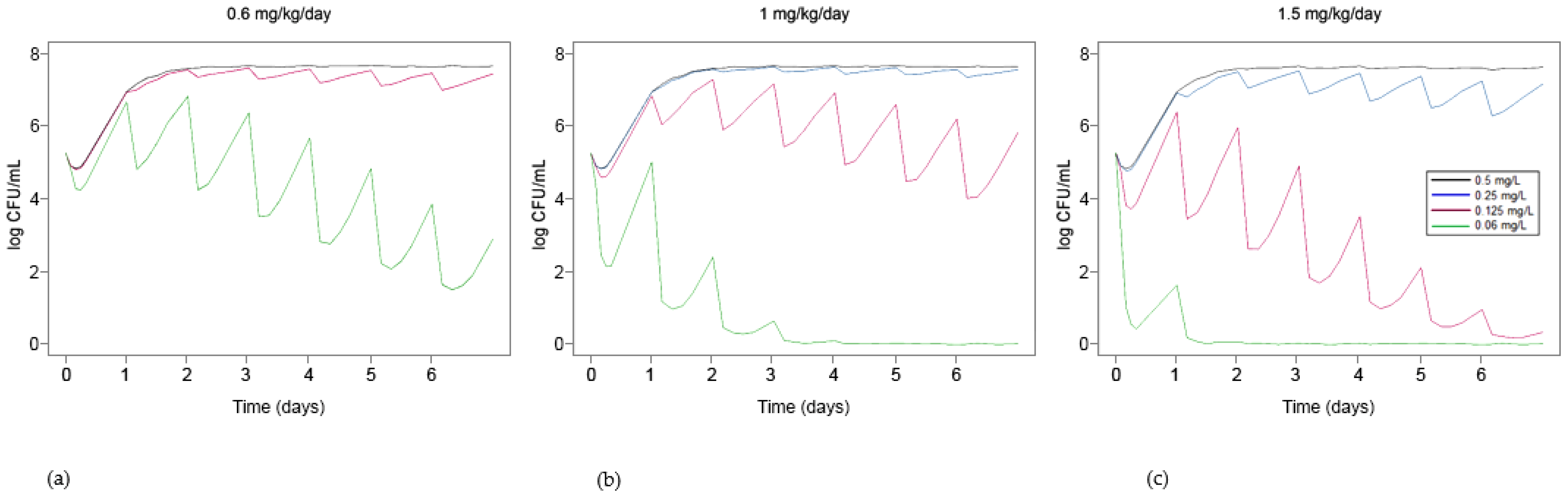
3.3. Simulation of Standard Treatments Using Human PK Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Chowdhary, A.; Sharma, C.; Meis, J.F. Candida auris: A rapidly emerging cause of hospital-acquired multidrug-resistant fungal infections globally. PLoS Pathog. 2017, 13, e1006290. [Google Scholar] [CrossRef] [PubMed]
- Quindós, G.; Marcos-Arias, C.; San-Millán, R.; Mateo, E.; Eraso, E. The continuous changes in the aetiology and epidemiology of invasive candidiasis: From familiar Candida albicans to multiresistant Candida auris. Int. Microbiol. 2018, 21, 107–119. [Google Scholar] [CrossRef]
- Sekyere, J.O. Candida auris: A systematic review and meta-analysis of current updates on an emerging multidrug-resistant pathogen. Microbiology 2018, 7, e00578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pemán, J.; Ruiz-Gaitán, A.; García-Vidal, C.; Salavert, M.; Ramírez, P.; Puchades, F.; García-Hita, M.; Alastruey-Izquierdo, A.; Quindós, G. Fungal co-infection in COVID-19 patients: Should we be concerned? Rev. Iberoam. Micol. 2020, 37, 41–46. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Prakash, A.; Meletiadis, J.; Sharma, C.; Chowdhary, A. Comparison of EUCAST and CLSI reference microdi-lution MICs of eight antifungal compounds for Candida auris and associated tentative epidemiological cutoff values. Antimicrob. Agents Chemother. 2017, 61, e00485-17. [Google Scholar] [CrossRef] [Green Version]
- Chaabane, F.; Graf, A.; Jequier, L.; Coste, A.T. Review on Antifungal Resistance Mechanisms in the Emerging Pathogen Candida auris. Front. Microbiol. 2019, 10, 2788. [Google Scholar] [CrossRef]
- Lockhart, S.R. Candida auris and multidrug resistance: Defining the new normal. Fungal Genet. Biol. 2019, 131, 103243. [Google Scholar] [CrossRef]
- Chowdhary, A.; Prakash, A.; Sharma, C.; Kordalewska, M.; Kumar, A.; Sarma, S.; Tarai, B.; Singh, A.; Upadhyaya, G.; Upadhyay, S.; et al. A multicentre study of antifungal susceptibility patterns among 350 Candida auris isolates (2009–2017) in India: Role of the ERG11 and FKS1 genes in azole and echinocandin resistance. J. Antimicrob. Chemother. 2018, 73, 891–899. [Google Scholar] [CrossRef]
- Kenters, N.; Kiernan, M.; Chowdhary, A.; Denning, D.W.; Pemán, J.; Saris, K.; Schelenz, S.; Tartari, E.; Widmer, A.; Meis, J.F.; et al. Control of Candida auris in healthcare institutions: Outcome of an International Society for Antimicrobial Chemotherapy expert meeting. Int. J. Antimicrob. Agents 2019, 54, 400–406. [Google Scholar] [CrossRef]
- Biagi, M.J.; Wiederhold, N.P.; Gibas, C.; Wickes, B.; Lozano, V.; Bleasdale, S.C.; Danziger, L. Development of High-Level Echinocandin Resistance in a Patient with Recurrent Candida auris Candidemia Secondary to Chronic Candiduria. Open Forum Infect. Dis. 2019, 6, ofz262. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Kim, M.-N.; Jang, S.J.; Ju, M.Y.; Kim, S.H.; Shin, M.G.; Suh, S.P.; Ryang, D.W. Detection of Amphotericin B Resistance in Candida haemulonii and Closely Related Species by Use of the Etest, Vitek-2 Yeast Susceptibility System, and CLSI and EUCAST Broth Microdilution Methods. J. Clin. Microbiol. 2012, 50, 1852–1855. [Google Scholar] [CrossRef] [Green Version]
- Morales, S.; Giraldo, C.M.P.; Garzón, A.C.; Martínez, H.P.; Rodríguez, G.J.; Moreno, C.A.A.; Rodriguez, J.Y. Invasive Infections with Multidrug-Resistant Yeast Candida auris, Colombia. Emerg. Infect. Dis. 2017, 23, 162–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhary, A.; Sharma, C.; Duggal, S.; Agarwal, K.; Prakash, A.; Singh, P.K.; Jain, S.; Kathuria, S.; Randhawa, H.S.; Hagen, F.; et al. New Clonal Strain of Candida auris, Delhi, India. Emerg. Infect. Dis. 2013, 19, 1670–1673. [Google Scholar] [CrossRef] [Green Version]
- Calvo, B.; Melo, A.S.A.; Perozo-Mena, A.; Hernandez, M.; Francisco, E.C.; Hagen, F.; Meis, J.F.; Colombo, A.L. First report of Candida auris in America: Clinical and microbiological aspects of 18 episodes of candidemia. J. Infect. 2016, 73, 369–374. [Google Scholar] [CrossRef]
- Schelenz, S.; Hagen, F.; Rhodes, J.L.; Abdolrasouli, A.; Chowdhary, A.; Hall, A.; Ryan, L.; Shackleton, J.; Trimlett, R.; Meis, J.F.; et al. First hospital outbreak of the globally emerging Candida auris in a European hospital. Antimicrob. Resist. Infect. Control. 2016, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Dudiuk, C.; Berrio, I.; Leonardelli, F.; Morales-Lopez, S.; Theill, L.; Macedo, D.; Rodriguez, J.Y.; Salcedo, S.; Marin, A.; Gamarra, S.; et al. Antifungal activity and killing kinetics of anidulafungin, caspofungin and amphotericin B against Candida auris. J. Antimicrob. Chemother. 2019, 74, 2295–2302. [Google Scholar] [CrossRef] [PubMed]
- Spivak, E.S.; Hanson, K.E. Candida auris: An Emerging Fungal Pathogen. J. Clin. Microbiol. 2018, 56, e01588-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alastruey-Izquierdo, A.; Asensio, A.; Besoli, A.; Calabuig, E.; Fernández-Ruiz, M.; Garcia-Vidal, C.; Gasch, O.; Guinea, J.; Martín-Gomez, M.T.; Paño, J.R. GEMICOMED/GEIRAS-SEIMC recommendations for the management of Candida auris infection and colonization. Rev. Iberoam. Micol. 2019, 36, 109–114. [Google Scholar] [CrossRef]
- Venisse, N.; Grégoire, N.; Marliat, M.; Couet, W. Mechanism-Based Pharmacokinetic-Pharmacodynamic Models of In Vitro Fungistatic and Fungicidal Effects against Candida albicans. Antimicrob. Agents Chemother. 2008, 52, 937–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Nguyen, M.H.; Cheng, S.; Schmidt, S.; Zhong, L.; Derendorf, H.; Clancy, C.J. A pharmacokinetic/pharmacodynamic mathematical model accurately describes the activity of voriconazole against Candida spp. in vitro. Int. J. Antimicrob. Agents 2008, 31, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Gil-Alonso, S.; Jauregizar, N.; Ortega, I.; Eraso, E.; Suarez, E.; Quindós, G. In vitro pharmacodynamic modelling of anidulafungin against Candida spp. Int. J. Antimicrob. Agents 2016, 47, 178–183. [Google Scholar] [CrossRef]
- Ruiz-Gaitán, A.; Moret, A.M.; Tasias-Pitarch, M.; Aleixandre-López, A.I.; Morel, H.M.; Calabuig, E.; Salavert-Lletí, M.; Ramírez, P.; López-Hontangas, J.L.; Hagen, F.; et al. An outbreak due to Candida auris with prolonged colonisation and candidaemia in a tertiary care European hospital. Mycoses 2018, 61, 498–505. [Google Scholar] [CrossRef] [Green Version]
- EUCAST. The European Committee for Antimicrobial Susceptibility Testing. Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Yeasts. EUCAST Definitive Document E.def 7.3.2. 2020. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Files/EUCAST_E_Def_7.3.2_Yeast_testing_definitive_revised_2020.pdf (accessed on 8 March 2021).
- Gil-Alonso, S.; Jauregizar, N.; Canton, E.; Eraso, E.; Quindos, G. In vitro fungicidal activities of anidulafungin, caspofungin, and micafungin against Candida glabrata, Candida bracarensis, and Candida nivariensis evaluated by time-kill studies. Antimicrob. Agents Chemother. 2015, 59, 3615–3618. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, E.I.; Friberg, L.E. Pharmacokinetic-Pharmacodynamic Modeling of Antibacterial Drugs. Pharmacol. Rev. 2013, 65, 1053–1090. [Google Scholar] [CrossRef] [Green Version]
- Bekersky, I.; Fielding, R.; Dressler, D.E.; Lee, J.W.; Buell, D.N.; Walsh, T.J. Pharmacokinetics, Excretion, and Mass Balance of Liposomal Amphotericin B (AmBisome) and Amphotericin B Deoxycholate in Humans. Antimicrob. Agents Chemother. 2002, 46, 828–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekersky, I.; Fielding, R.; Dressler, D.E.; Lee, J.W.; Buell, D.N.; Walsh, T.J. Plasma Protein Binding of Amphotericin B and Pharmacokinetics of Bound versus Unbound Amphotericin B after Administration of Intravenous Liposomal Amphotericin B (AmBisome) and Amphotericin B Deoxycholate. Antimicrob. Agents Chemother. 2002, 46, 834–840. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.; Schuck, E.; Kumar, V.; Burkhardt, O.; Derendorf, H. Integration of pharmacokinetic/pharmacodynamic modeling and simulation in the development of new anti-infective agents—Minimum inhibitory concentration versus time-kill curves. Expert Opin. Drug Discov. 2007, 2, 849–860. [Google Scholar] [CrossRef]
- Mueller, M.; de la Peña, A.; Derendorf, H. Issues in Pharmacokinetics and Pharmacodynamics of Anti-Infective Agents: Kill Curves versus MIC. Antimicrob. Agents Chemother. 2004, 48, 369–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klepser, M.E.; Wolfe, E.J.; Jones, R.N.; Nightingale, C.H.; Pfaller, M.A. Antifungal pharmacodynamic characteristics of fluconazole and amphotericin B tested against Candida albicans. Antimicrob. Agents Chemother. 1997, 41, 1392–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantón, E.; Pemán, J.; Gobernado, M.; Viudes, A.; Espinel-Ingroff, A. Patterns of Amphotericin B Killing Kinetics against Seven Candida Species. Antimicrob. Agents Chemother. 2004, 48, 2477–2482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canton, E.; Peman, J.; Sastre, M.; Romero, M.; Espinel-Ingroff, A. Killing kinetics of caspofungin, micafungin, and ampho-tericin B against Candida guilliermondii. Antimicrob. Agents Chemother. 2006, 50, 2829–2832. [Google Scholar] [CrossRef] [Green Version]
- Brill, M.; Kristoffersson, A.; Zhao, C.; Nielsen, E.; Friberg, L. Semi-mechanistic pharmacokinetic–pharmacodynamic modelling of antibiotic drug combinations. Clin. Microbiol. Infect. 2018, 24, 697–706. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, E.I.; Viberg, A.; Lowdin, E.; Cars, O.; Karlsson, M.O.; Sandstrom, M. Semimechanistic pharmacokinet-ic/pharmacodynamic model for assessment of activity of antibacterial agents from time-kill curve experiments. Antimicrob. Agents Chemother. 2007, 51, 128–136. [Google Scholar] [CrossRef] [Green Version]
- Sasse, C.; Dunkel, N.; Schäfer, T.; Schneider, S.; Dierolf, F.; Ohlsen, K.; Morschhäuser, J. The stepwise acquisition of fluconazole resistance mutations causes a gradual loss of fitness inCandida albicans. Mol. Microbiol. 2012, 86, 539–556. [Google Scholar] [CrossRef]
- Asner, S.; Giulieri, S.; Diezi, M.; Marchetti, O.; Sanglard, D. Acquired Multidrug Antifungal Resistance in Candida lusitaniae During Therapy. Open Forum Infect. Dis. 2015, 59, 7715–7722. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. Amphotericin B: Rationale for the Clinical Breakpoints, Version 2.0. 2020. Available online: http://www.eucast.org (accessed on 25 February 2021).
- Lepak, A.J.; Zhao, M.; Berkow, E.L.; Lockhart, S.R.; Andes, D.R. Pharmacodynamic Optimization for Treatment of Invasive Candida auris Infection. Antimicrob. Agents Chemother. 2017, 61, 00791-17. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization Model List of Essential Medicines, 21st List, 2019. World Health Organization: Geneva, Switzerland, 2019. Available online: https://www.who.int/medicines/publications/essentialmedicines/en/ (accessed on 8 March 2021).
- Groll, A.H.; Rijnders, B.; Walsh, T.J.; Adler-Moore, J.; Lewis, R.E.; Brüggemann, R.J.M. Clinical Pharmacokinetics, Pharmacodynamics, Safety and Efficacy of Liposomal Amphotericin B. Clin. Infect. Dis. 2019, 68, S260–S274. [Google Scholar] [CrossRef]
- Arendrup, M.; Friberg, N.; Mares, M.; Kahlmeter, G.; Meletiadis, J.; Guinea, J.; Andersen, C.; Arikan-Akdagli, S.; Barchiesi, F.; Chryssanthou, E.; et al. How to interpret MICs of antifungal compounds according to the revised clinical breakpoints v. 10.0 European committee on antimicrobial susceptibility testing (EUCAST). Clin. Microbiol. Infect. 2020, 26, 1464–1472. [Google Scholar] [CrossRef]
- Kardos, T.; Kovács, R.; Kardos, G.; Varga, I.; Bozó, A.; Tóth, Z.; Nagy, F.; Majoros, L. Poor in vivo efficacy of caspofungin, micafungin and amphotericin B against wild-type Candida krusei clinical isolates does not correlate with in vitro susceptibility results. J. Chemother. 2018, 30, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Mariné, M.; Espada, R.; Torrado, J.; Pastor, F.J.; Guarro, J. Efficacy of a new formulation of amphotericin B in murine disseminated infections by Candida glabrata or Candida tropicalis. Int. J. Antimicrob. Agents 2009, 34, 566–569. [Google Scholar] [CrossRef] [Green Version]
- Elefanti, A.; Mouton, J.W.; Verweij, P.E.; Zerva, L.; Meletiadis, J. Susceptibility Breakpoints for Amphotericin B and Aspergillus Species in anIn VitroPharmacokinetic-Pharmacodynamic Model Simulating Free-Drug Concentrations in Human Serum. Antimicrob. Agents Chemother. 2014, 58, 2356–2362. [Google Scholar] [CrossRef] [Green Version]
- Mohr, J.F.; Hall, A.C.; Ericsson, C.D.; Ostrosky-Zeichner, L. Fatal Amphotericin B Overdose Due to Administration of Nonlipid Formulation Instead of Lipid Formulation. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2005, 25, 426–428. [Google Scholar] [CrossRef]
- De la Peña, A.; Gräbe, A.; Rand, K.H.; Rehak, E.; Gross, J.; Thyroff-Friesinger, U.; Müller, M.; Derendorf, H. PK–PD modelling of the effect of cefaclor on four different bacterial strains. Int. J. Antimicrob. Agents 2004, 23, 218–225. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Model Estimate and RSE (CV %) | Bootstrap Estimate (Mean and 95% CI) |
|---|---|---|---|
| kgrowthS (h−1) | Fungal growth rate constant of the S subpopulation | 0.111 (3%) | 0.111 (0.101–0.116) |
| kgrowthR (h−1) | Fungal growth rate constant of the R subpopulation | 0.01 (fixed) | - |
| kdeath (h−1) | Fungal death rate constant | 0.01 (fixed) | - |
| Emax (h−1) | Maximum kill rate constant of amphotericin B | 0.784 (12%) | 0.795 (0.635–1.04) |
| EC50 (mg/L) | Concentration of amphotericin B at which 50% of the Emax is achieved | 1.88 (3%) | 1.89 (1.78–2.05) |
| h | Hill factor that that modifies the steepness of the slope and smoothens the curve | 4 (fixed) | - |
| α (control) | Delay in fungal growth in the absence of drug | 0.748 (3%) | 0.754 (0.664–0.882) |
| α (drug) | Delay in fungal growth in the presence of drug | 0.231 (10%) | 0.233 (0.193–0.274) |
| Nmax (log CFU/mL) | Maximum fungal density | 7.66 (1%) | 7.67 (7.47–7.87) |
| σ (log CFU/mL) | Residual error | 0.271 (14%) | 0.270 (0.190–0.327) |
| π1 (%CV) | Occasion 1 | 0 (fixed) | - |
| π2 (%CV) | Occasion 2 | 9.5 (35%) | 9.22 (2.45–15.34) |
| π3 (%CV) | Occasion 3 | 18.4 (24%) | 18.76 (10.07–28.12) |
| π4 (%CV) | Occasion 4 | 7.5 (37%) | 7.13 (2.75–13.19) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caballero, U.; Eraso, E.; Pemán, J.; Quindós, G.; Vozmediano, V.; Schmidt, S.; Jauregizar, N. In Vitro Pharmacokinetic/Pharmacodynamic Modelling and Simulation of Amphotericin B against Candida auris. Pharmaceutics 2021, 13, 1767. https://doi.org/10.3390/pharmaceutics13111767
Caballero U, Eraso E, Pemán J, Quindós G, Vozmediano V, Schmidt S, Jauregizar N. In Vitro Pharmacokinetic/Pharmacodynamic Modelling and Simulation of Amphotericin B against Candida auris. Pharmaceutics. 2021; 13(11):1767. https://doi.org/10.3390/pharmaceutics13111767
Chicago/Turabian StyleCaballero, Unai, Elena Eraso, Javier Pemán, Guillermo Quindós, Valvanera Vozmediano, Stephan Schmidt, and Nerea Jauregizar. 2021. "In Vitro Pharmacokinetic/Pharmacodynamic Modelling and Simulation of Amphotericin B against Candida auris" Pharmaceutics 13, no. 11: 1767. https://doi.org/10.3390/pharmaceutics13111767