
Tetraethylenepentamine-Coated β Cyclodextrin Nanoparticles for Dual DNA and siRNA Delivery

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Functionalization and Characterization of TEPA-BCD Carrier
2.2. Effects of Complex Ratio on Physicochemical Properties of DNA- and siRNA-Loaded TEPA-BCD
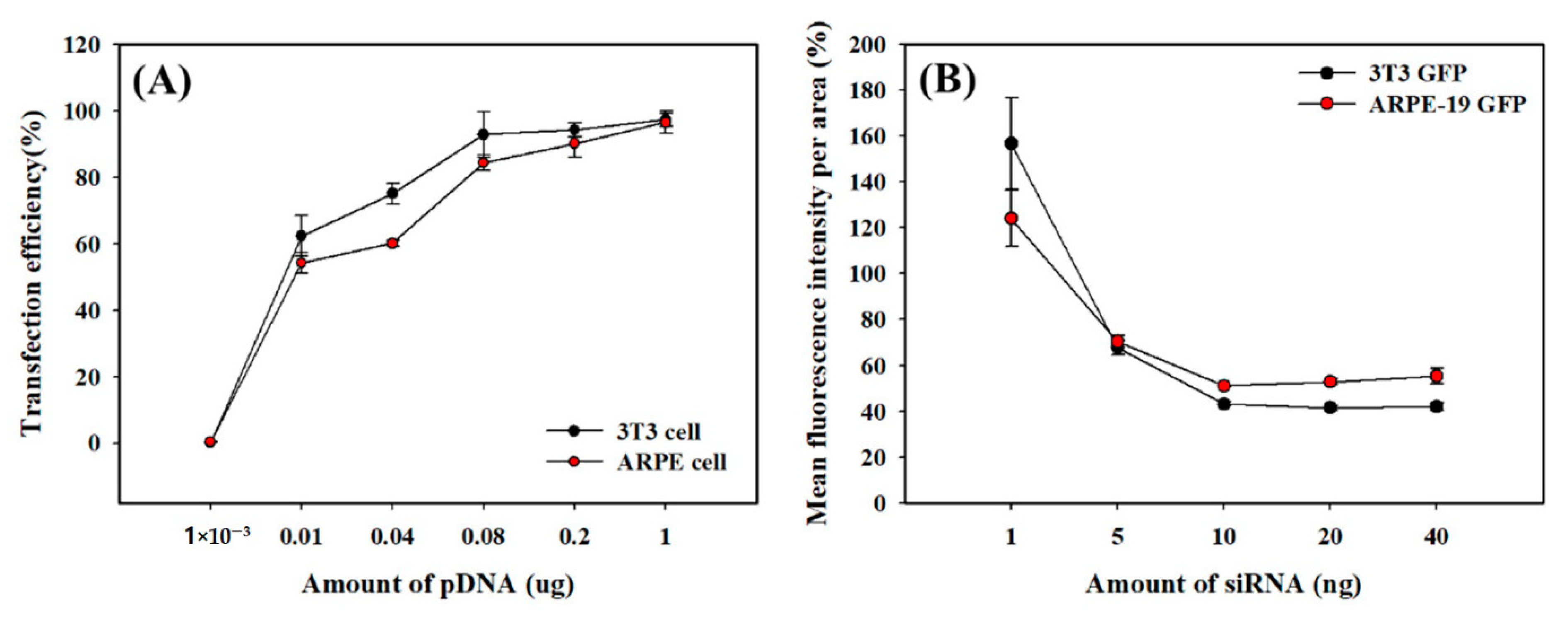
2.3. Transfection Efficiency and Viability of the TEPA-BCD Polyplexes
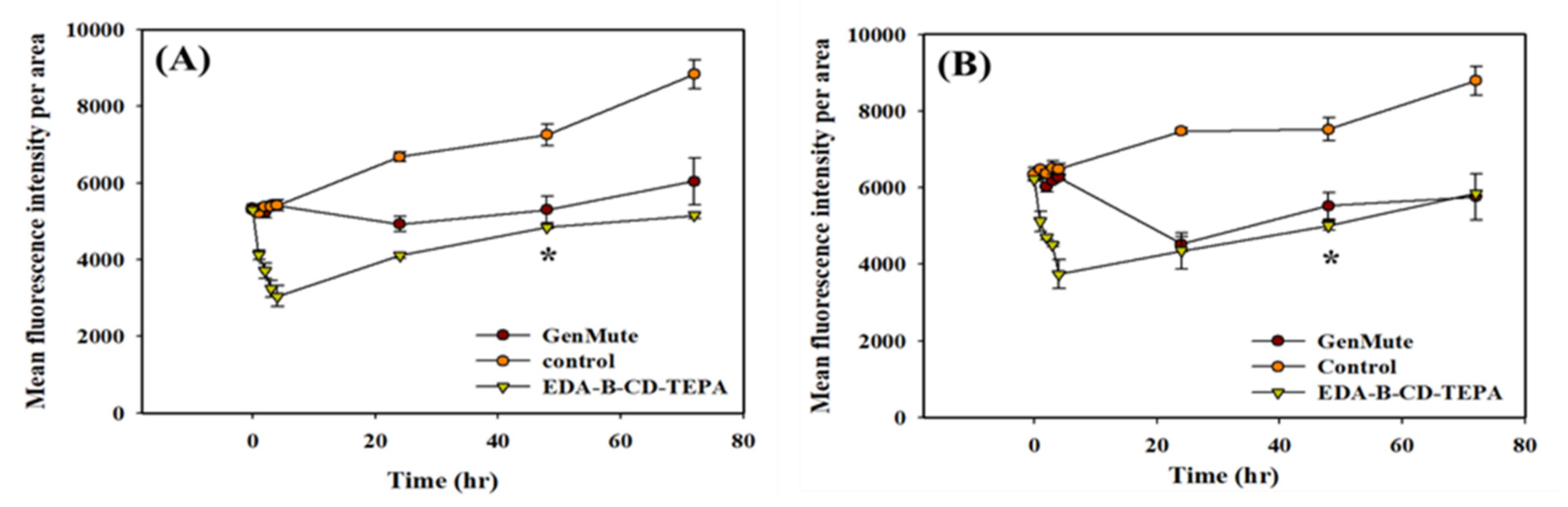
2.4. Kinetics and Internalization Mechanisms of GFP Expression and Gene Silencing after DNA and siRNA Delivery
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. TEPA-BCD Synthesis
4.3. Preparation of TEPA-BCD/Plasmid DNA and TEPA-BCD/siRNA
4.4. Characterization of TEPA-BCD
4.5. GFP Silencing and Transfection In Vitro
4.6. Effects of Endocytic Inhibitors on Transfection and Gene-Silencing Efficiency
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Setten, R.L.; Rossi, J.J.; Han, S.-P. The current state and future directions of RNAi-based therapeutics. Nat. Rev. Drug Discov. 2019, 18, 421–446. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Siegwart, D.J.; Anderson, D.G. Strategies, design, and chemistry in siRNA delivery systems. Adv. Drug Deliv. Rev. 2019, 144, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yan, Y. A review of ph-responsive organic–inorganic hybrid nanoparticles for rnai-based therapeutics. Macromol. Biosci. 2021, 21, 2100183. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, J.; Li, B.; Meng, L.; Tian, Z. Recent advances in mechanism-based chemotherapy drug-sirna pairs in co-delivery systems for cancer: A review. Colloids Surf. B Biointerfaces 2017, 157, 297–308. [Google Scholar] [CrossRef]
- Sarett, S.M.; Werfel, T.A.; Chandra, I.; Jackson, M.A.; Kavanaugh, T.E.; Hattaway, M.E.; Giorgio, T.D.; Duvall, C.L. Hydrophobic interactions between polymeric carrier and palmitic acid-conjugated siRNA improve PEGylated polyplex stability and enhance in vivo pharmacokinetics and tumor gene silencing. Biomaterials 2016, 97, 122–132. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Ellert-Miklaszewska, A.; Garofalo, S.; Dey, A.K.; Tang, J.; Jiang, Y.; Clément, F.; Marche, P.N.; Liu, X.; Kaminska, B.; et al. Synthesis and use of an amphiphilic dendrimer for siRNA delivery into primary immune cells. Nat. Protoc. 2020, 16, 327–351. [Google Scholar] [CrossRef]
- Godinho, B.M.D.C.; Ogier, J.R.; Darcy, R.; O’Driscoll, C.M.; Cryan, J.F. Self-assembling Modified β-Cyclodextrin Nanoparticles as Neuronal siRNA Delivery Vectors: Focus on Huntington’s Disease. Mol. Pharm. 2013, 10, 640–649. [Google Scholar] [CrossRef]
- Ramsay, E.; Raviña, M.; Sarkhel, S.; Hehir, S.; Cameron, N.R.; Ilmarinen, T.; Skottman, H.; Kjems, J.; Urtti, A.; Ruponen, M.; et al. Avoiding the Pitfalls of siRNA Delivery to the Retinal Pigment Epithelium with Physiologically Relevant Cell Models. Pharmaceutics 2020, 12, 667. [Google Scholar] [CrossRef]
- Mitrach, F.; Schmid, M.; Toussaint, M.; Dukic-Stefanovic, S.; Deuther-Conrad, W.; Franke, H.; Ewe, A.; Aigner, A.; Wölk, C.; Brust, P.; et al. Amphiphilic Anionic Oligomer-Stabilized Calcium Phosphate Nanoparticles with Prospects in siRNA Delivery via Convection-Enhanced Delivery. Pharmaceutics 2022, 14, 326. [Google Scholar] [CrossRef]
- Lamberti, G.; Barba, A.A. Drug Delivery of siRNA Therapeutics. Pharmaceutics 2020, 12, 178. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ye, L.; Xi, J.; Wang, J.; Feng, Z.-G. Cyclodextrin Polymers: Structure, Synthesis, and Use as Drug Carriers. Prog. Polym. Sci. 2021, 118, 101408. [Google Scholar] [CrossRef]
- Qie, S.; Hao, Y.; Liu, Z.; Wang, J.; Xi, J. Advances in Cyclodextrin Polymers and Their Applications in Biomedicine. Acta Chim. Sin. 2020, 78, 232. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Hidalgo, T.; Cazade, P.-A.; Darcy, R.; Cronin, M.F.; Dorin, I.; O’Driscoll, C.M.; Thompson, D. Self-Assembled Cationic β-Cyclodextrin Nanostructures for siRNA Delivery. Mol. Pharm. 2019, 16, 1358–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, B.; Liu, Y.; Liu, J. Smart stimuli-responsive drug delivery systems based on cyclodextrin: A review. Carbohydr. Polym. 2020, 251, 116871. [Google Scholar] [CrossRef] [PubMed]
- Ceborska, M. Folate appended cyclodextrins for drug, DNA, and sirna delivery. Eur. J. Pharm. Biopharm. 2017, 120, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Lv, P.; Zhou, C.; Zhao, Y.; Liao, X.; Yang, B. Modified-epsilon-polylysine-grafted-pei-beta-cyclodextrin supramolecular carrier for gene delivery. Carbohydr. Polym. 2017, 168, 103–111. [Google Scholar] [CrossRef]
- Evans, J.C.; Malhotra, M.; Sweeney, K.; Darcy, R.; Nelson, C.C.; Hollier, B.G.; O’Driscoll, C.M. Folate-targeted amphiphilic cyclodextrin nanoparticles incorporating a fusogenic peptide deliver therapeutic siRNA and inhibit the invasive capacity of 3D prostate cancer tumours. Int. J. Pharm. 2017, 532, 511–518. [Google Scholar] [CrossRef] [Green Version]
- Sherje, A.P.; Dravyakar, B.R.; Kadam, D.; Jadhav, M. Cyclodextrin-based nanosponges: A critical review. Carbohydr. Polym. 2017, 173, 37–49. [Google Scholar] [CrossRef]
- Andrzejewska, W.; Wilkowska, M.; Skrzypczak, A.; Kozak, M. Ammonium Gemini Surfactants Form Complexes with Model Oligomers of siRNA and dsDNA. Int. J. Mol. Sci. 2019, 20, 5546. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, D.W.; Su, H.; Hildebrandt, I.J.; Weber, W.A.; Davis, M.E. Impact of tumor-specific targeting on the biodistribution and efficacy of siRNA nanoparticles measured by multimodality in vivo imaging. Proc. Natl. Acad. Sci. USA 2007, 104, 15549–15554. [Google Scholar] [CrossRef] [Green Version]
- Hu-Lieskovan, S.; Heidel, J.D.; Bartlett, D.W.; Davis, M.E.; Triche, T.J. Sequence-specific knockdown of ews-fli1 by targeted, nonviral delivery of small interfering rna inhibits tumor growth in a murine model of metastatic ewing’s sarcoma. Cancer Res. 2005, 65, 8984–8992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salcher, E.E.; Kos, P.; Fröhlich, T.; Badgujar, N.; Scheible, M.; Wagner, E. Sequence-defined four-arm oligo(ethanamino)amides for pDNA and siRNA delivery: Impact of building blocks on efficacy. J. Control. Release 2012, 164, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Higashi, T.; Motoyama, K.; Jono, H.; Ando, Y.; Arima, H. In vitro and in vivo siRNA delivery to hepatocyte utilizing ternary complexation of lactosylated dendrimer/cyclodextrin conjugates, siRNA and low-molecular-weight sacran. Int. J. Biol. Macromol. 2017, 107, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, A.; Higashi, T.; Motoyama, K.; Arima, H. Ternary complexes of folate-PEG-appended dendrimer (G4)/α-cyclodextrin conjugate, siRNA and low-molecular-weight polysaccharide sacran as a novel tumor-selective siRNA delivery system. Int. J. Biol. Macromol. 2017, 99, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-H.; Lee, G.-W.; Wu, W.-C.; Wang, C.-C. Encapsulating curcumin in ethylene diamine-β-cyclodextrin nanoparticle improves topical cornea delivery. Colloids Surfaces B Biointerfaces 2020, 186, 110726. [Google Scholar] [CrossRef]
- Crini, G.; Torri, G.; Guerrini, M.; Martel, B.; Lekchiri, Y.; Morcellet, M. Linear Cyclodextrin-Poly(Vinylamine): Synthesis and Nmr Characterization. Eur. Polym. J. 1997, 33, 1143–1151. [Google Scholar] [CrossRef]
- Popr, M.; Hybelbauerová, S.; Jindřich, J. A complete series of 6-deoxy-monosubstituted tetraalkylammonium derivatives of α-, β-, and γ-cyclodextrin with 1, 2, and 3 permanent positive charges. Beilstein J. Org. Chem. 2014, 10, 1390–1396. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Y.; Fan, X.D.; Gao, L. Synthesis and characterization of β-cyclodextrin based functional monomers and its copolymers with n-isopropylacrylamide. Macromol. Biosci. 2003, 3, 715–719. [Google Scholar] [CrossRef]
- Migneault, I.; Dartiguenave, C.; Bertrand, M.J.; Waldron, K.C. Glutaraldehyde: Behavior in aqueous solution, reaction with proteins, and application to enzyme crosslinking. Biotechniques 2004, 37, 790–802. [Google Scholar] [CrossRef]
- Kumari, M.; Liu, C.-H.; Wu, W.-C.; Wang, C.-C. Gene delivery using layer-by-layer functionalized multi-walled carbon nanotubes: Design, characterization, cell line evaluation. J. Mater. Sci. 2021, 56, 7022–7033. [Google Scholar] [CrossRef]
- Kumari, M.; Liu, C.-H.; Wu, W.-C. Oligochitosan modified albumin as plasmid DNA delivery vector: Endocytic trafficking, polyplex fate, in vivo compatibility. Int. J. Biol. Macromol. 2019, 142, 492–502. [Google Scholar] [CrossRef] [PubMed]
- Kanasty, R.L.; Dorkin, J.R.; Vegas, A.; Anderson, D.G. Delivery materials for siRNA therapeutics. Nat. Mater. 2013, 12, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Hammoud, Z.; Khreich, N.; Auezova, L.; Fourmentin, S.; Elaissari, A.; Greige-Gerges, H. Cyclodextrin-membrane interaction in drug delivery and membrane structure maintenance. Int. J. Pharm. 2019, 564, 59–76. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.Y.; Alnakhli, M.; Bhardwaj, R.; Apostolou, S.; Sinha, S.; Fraser, C.; Kuchel, T.; Kuss, B.; Voelcker, N.H. Delivery of siRNA in vitro and in vivo using PEI-capped porous silicon nanoparticles to silence MRP1 and inhibit proliferation in glioblastoma. J. Nanobiotechnol. 2018, 16, 38. [Google Scholar] [CrossRef]
- Xia, Y.; Tang, G.; Wang, C.; Zhong, J.; Chen, Y.; Hua, L.; Li, Y.; Liu, H.; Zhu, B. Functionalized selenium nanoparticles for targeted siRNA delivery silence Derlin1 and promote antitumor efficacy against cervical cancer. Drug Deliv. 2020, 27, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Ross, N.L.; Munsell, E.V.; Sabanayagam, C.; Sullivan, M.O. Histone-targeted Polyplexes Avoid Endosomal Escape and Enter the Nucleus during Postmitotic Redistribution of ER Membranes. Mol. Ther. Nucleic Acids 2015, 4, e226. [Google Scholar] [CrossRef]
- Lai, W.-F. Cyclodextrins in non-viral gene delivery. Biomaterials 2014, 35, 401–411. [Google Scholar] [CrossRef]
- Akinc, A.; Thomas, M.; Klibanov, A.M.; Langer, R. Exploring polyethylenimine-mediated DNA transfection and the proton sponge hypothesis. J. Gene Med. 2005, 7, 657–663. [Google Scholar] [CrossRef]
- Manzanares, D.; Pérez-Carrión, M.D.; Blanco, J.L.J.; Mellet, C.O.; Fernández, J.M.G.; Ceña, V. Cyclodextrin-Based Nanostructure Efficiently Delivers siRNA to Glioblastoma Cells Preferentially via Macropinocytosis. Int. J. Mol. Sci. 2020, 21, 9306. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Guo, J.; Raftery, R.M.; Castaño, I.M.; Curtin, C.M.; Gooding, M.; Darcy, R.; O’ Brien, F.J.; O’ Driscoll, C.M. Nanoparticle-mediated sirna delivery assessed in a 3d co-culture model simulating prostate cancer bone metastasis. Int. J. Pharm. 2016, 511, 1058–1069. [Google Scholar] [CrossRef]
- Malhotra, M.; Gooding, M.; Evans, J.C.; O’Driscoll, D.; Darcy, R.; O’Driscoll, C.M. Cyclodextrin-sirna conjugates as versatile gene silencing agents. Eur. J. Pharm. Sci. 2018, 114, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-Q.; Yu, Q.-L.; Liu, Y.-H.; Yu, H.-J.; Chen, Y.; Liu, Y. Highly efficient photocontrolled targeted delivery of siRNA by a cyclodextrin-based supramolecular nanoassembly. Chem. Commun. 2020, 56, 3907–3910. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, A.; Higashi, T.; Motoyama, K.; Arima, H. In vitro and in vivo tumor-targeting sirna delivery using folate-peg-appended dendrimer (g4)/α-cyclodextrin conjugates. Bioconjugate Chem. 2016, 27, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Dalal, C.; Saha, A.; Jana, N.R. Nanoparticle Multivalency Directed Shifting of Cellular Uptake Mechanism. J. Phys. Chem. C 2016, 120, 6778–6786. [Google Scholar] [CrossRef] [Green Version]
- Carro, A.C.; Piccini, L.E.; Damonte, E.B. Blockade of dengue virus entry into myeloid cells by endocytic inhibitors in the presence or absence of antibodies. PLoS Neglected Trop. Dis. 2018, 12, e0006685. [Google Scholar] [CrossRef]
- Salatin, S.; Khosroushahi, A.Y. Overviews on the cellular uptake mechanism of polysaccharide colloidal nanoparticles. J. Cell. Mol. Med. 2017, 21, 1668–1686. [Google Scholar] [CrossRef] [Green Version]
- Scholz, C.; Wagner, E. Therapeutic plasmid DNA versus siRNA delivery: Common and different tasks for synthetic carriers. J. Control. Release 2012, 161, 554–565. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, A.; Miyata, K.; Kataoka, K. Recent progress in development of siRNA delivery vehicles for cancer therapy. Adv. Drug Deliv. Rev. 2016, 104, 61–77. [Google Scholar] [CrossRef] [Green Version]
- Mousazadeh, H.; Pilehvar-Soltanahmadi, Y.; Dadashpour, M.; Zarghami, N. Cyclodextrin based natural nanostructured carbohydrate polymers as effective non-viral siRNA delivery systems for cancer gene therapy. J. Control. Release 2020, 330, 1046–1070. [Google Scholar] [CrossRef]
- Petter, R.C.; Salek, J.S.; Sikorski, C.T.; Kumaravel, G.; Lin, F.T. Cooperative binding by aggregated mono-6-(alkylamino)-β-cyclodextrins. J. Am. Chem. Soc. 1990, 112, 3860–3868. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibitor | Target of the Inhibitor | Conc. | Plasmid Transfection | siRNA Silencing | ||
|---|---|---|---|---|---|---|
| 3T3 | ARPE | 3T3- GFP | ARPE- GFP | |||
| Chlorpromazine (CPZ) | Clathrin-mediated endocytosis | 30 mM | + | + | + | + |
| Genistein (Gen) | Caveolae-mediated endocytosis | 200 mM | - | - | + | + |
| Monensin (Mon) | Transport to plasma membrane | 3 mM | + | + | - | - |
| Sodium azide (SA) | Active transport | 1% | + | + | + | + |
| Methyl-β-cyclodextrin (MCD) | Cholesterol-dependent endocytosis | 10 mM | - | - | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.-H.; Shih, P.-Y.; Lin, C.-H.; Chen, Y.-J.; Wu, W.-C.; Wang, C.-C. Tetraethylenepentamine-Coated β Cyclodextrin Nanoparticles for Dual DNA and siRNA Delivery. Pharmaceutics 2022, 14, 921. https://doi.org/10.3390/pharmaceutics14050921
Liu C-H, Shih P-Y, Lin C-H, Chen Y-J, Wu W-C, Wang C-C. Tetraethylenepentamine-Coated β Cyclodextrin Nanoparticles for Dual DNA and siRNA Delivery. Pharmaceutics. 2022; 14(5):921. https://doi.org/10.3390/pharmaceutics14050921
Chicago/Turabian StyleLiu, Chi-Hsien, Pei-Yin Shih, Cheng-Han Lin, Yi-Jun Chen, Wei-Chi Wu, and Chun-Chao Wang. 2022. "Tetraethylenepentamine-Coated β Cyclodextrin Nanoparticles for Dual DNA and siRNA Delivery" Pharmaceutics 14, no. 5: 921. https://doi.org/10.3390/pharmaceutics14050921
APA StyleLiu, C.-H., Shih, P.-Y., Lin, C.-H., Chen, Y.-J., Wu, W.-C., & Wang, C.-C. (2022). Tetraethylenepentamine-Coated β Cyclodextrin Nanoparticles for Dual DNA and siRNA Delivery. Pharmaceutics, 14(5), 921. https://doi.org/10.3390/pharmaceutics14050921