
Safety and Efficacy of the Bordetella bronchiseptica Vaccine Combined with a Vegetable Oil Adjuvant and Multi-Omics Analysis of Its Potential Role in the Protective Response of Rabbits


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Vaccine Preparation
2.3. Experimental Design
2.4. Analysis of Bb-Specific Antibodies and Cytokine Levels
2.5. Bacterial Loads and Histopathological Examination
2.6. Transcriptomic Analysis
2.7. Proteomic Analysis
2.8. Integrated Analysis of Transcriptome and Proteome Data
2.9. Validation of DEGs/DEPs by Real Time-qPCR/PRM
2.10. Statistical Analysis
3. Results
3.1. Safety Estimation of E515 and the E515-Adjuvanted Bb Vaccine
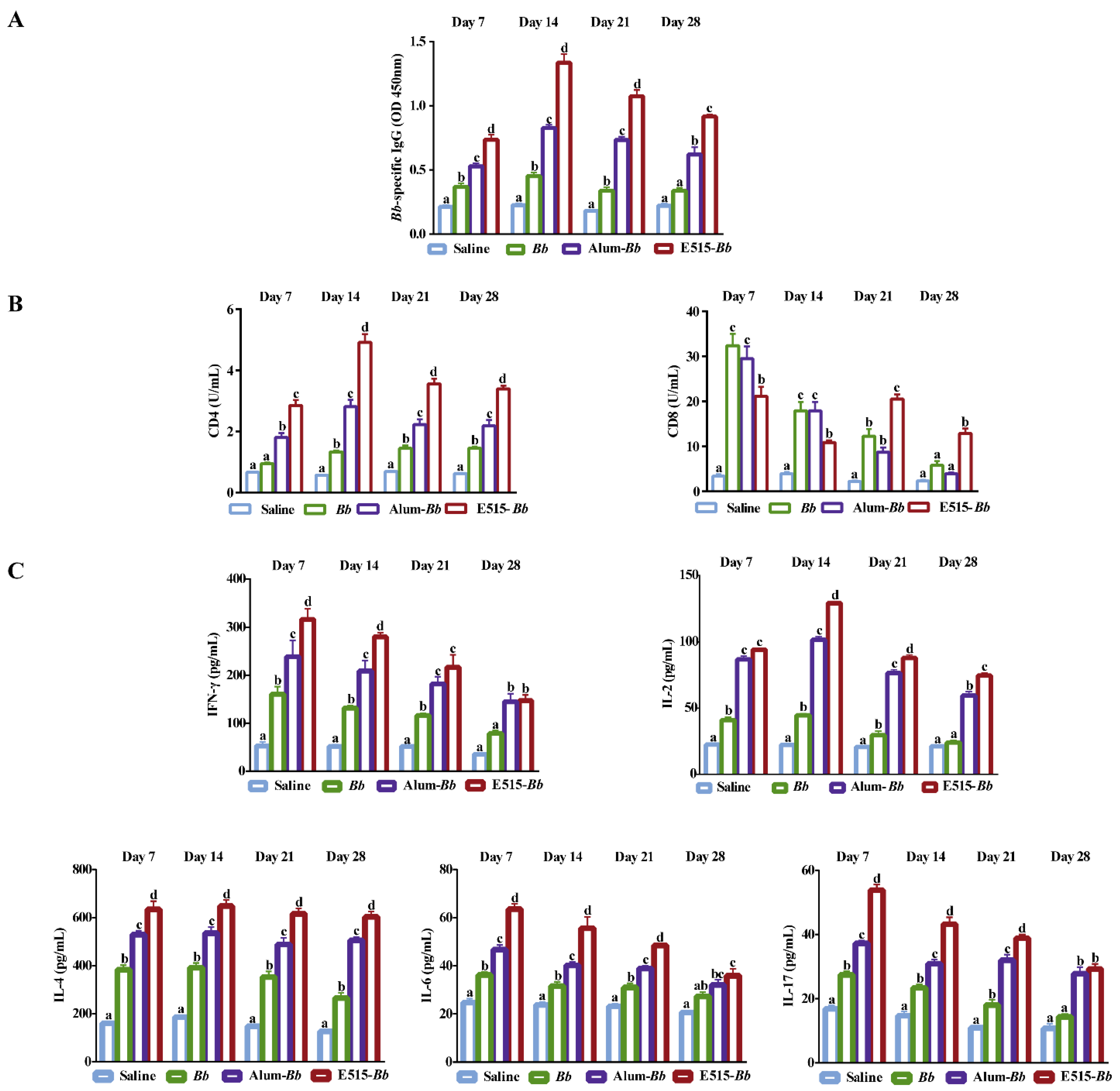
3.2. Evaluation of the Immune Effect of the E515-Bb Vaccine
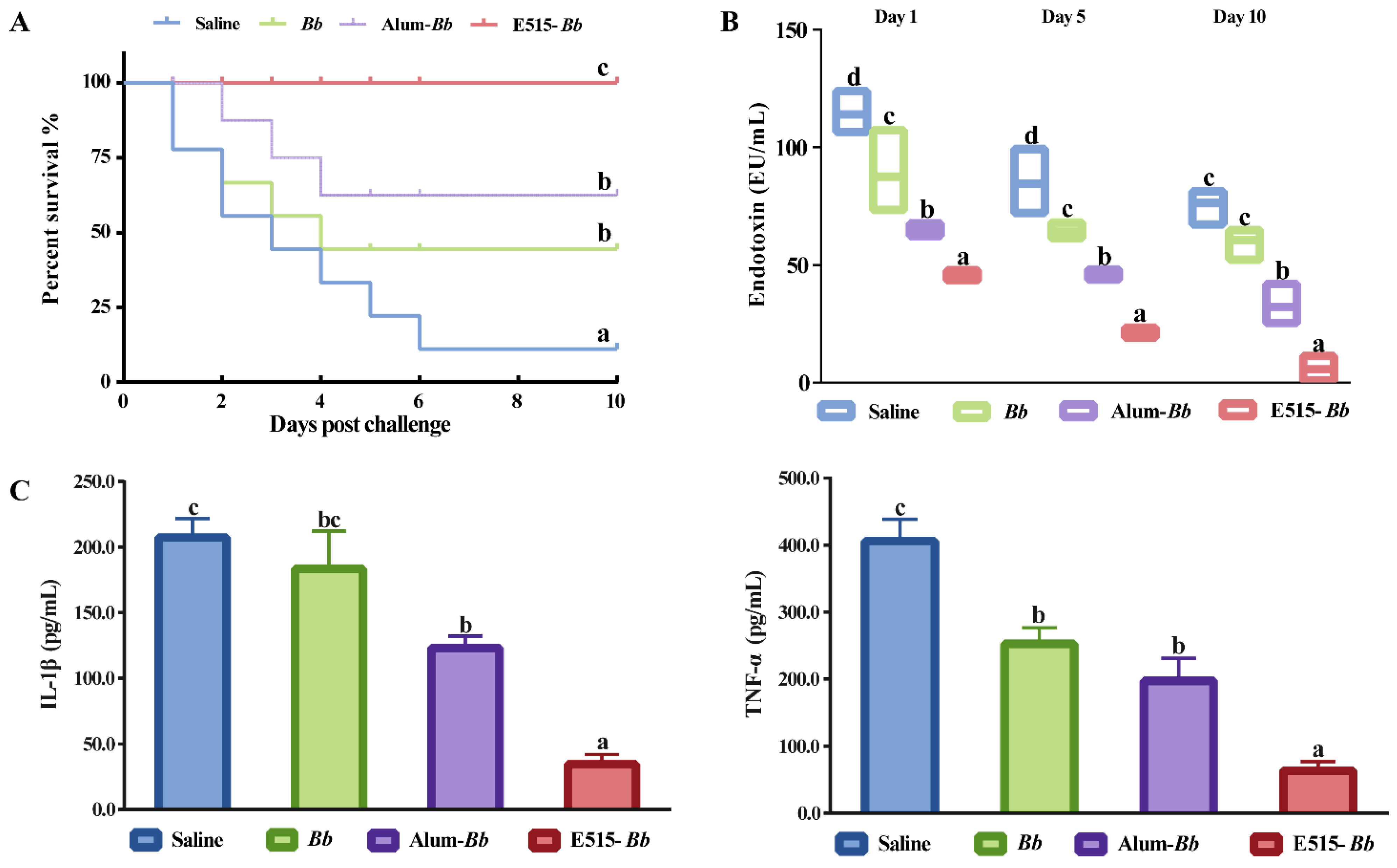
3.3. Assessment of the Role of the E515-Bb Vaccine in Protecting Rabbits from the Bb Challenge
3.4. Evaluation of the Long-Term Protection Provided by the E515-Bb Vaccine
3.5. Differential Gene Expression Analysis
3.6. Differential Protein Expression Analysis
3.7. Correlation Analysis of the Transcriptome and Proteome
3.8. Validation of DEGs/DEPs by RT–qPCR/PRM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parkhill, J.; Sebaihia, M.; Preston, A.; Murphy, L.D.; Thomson, N.; Harris, D.E.; Holden, M.T.G.; Churcher, C.M.; Bentley, S.D.; Mungall, K.L.; et al. Comparative analysis of the genome sequences of Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica. Nat. Genet. 2003, 35, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zeng, W.; Li, R.; Hoffman, L.; He, Z.; Sun, Q.; Li, H. Rabbit meat production and processing in China. Meat Sci. 2018, 145, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sun, S.; Chen, Y.; Chen, D.; Sang, L.; Xie, X. Characterisation of Bordetella bronchiseptica isolated from rabbits in Fujian, China. Epidemiol. Infect 2020, 148, e237. [Google Scholar] [CrossRef] [PubMed]
- Kameyama, H.; Fujimoto, Y.; Tomioka, Y.; Yamamoto, S.; Suyama, H.; Inoue, H.; Takahashi, E.; Ono, E. Pathogenicity of Bordetella bronchiseptica isolated from apparently healthy rabbits in guinea pig, rat, and mouse. J. Vet. Med. Sci. 2022, 84, 574–581. [Google Scholar] [CrossRef]
- Vötsch, D.; Willenborg, M.; Baumgärtner, W.; Rohde, M.; Valentin-Weigand, P. Bordetella bronchiseptica promotes adherence, colonization, and cytotoxicity of Streptococcus suis in a porcine precision-cut lung slice model. Virulence 2021, 12, 84–95. [Google Scholar] [CrossRef]
- Xiao, C.; Bao, G.; Liu, Y.; Wei, Q.; Ji, Q.; Liu, Y.; Pan, L. Greater efficacy of the ECMS-oil adjuvant over other formulations on immune responses against Bordetella bronchiseptica in rabbits and the underlying mechanism. Int. Immunopharmacol. 2016, 38, 194–203. [Google Scholar] [CrossRef]
- Papantoniou, S.; Tsakiris, A.; Ladopoulos, T.; Kranidiotis, G.; Tamvakos, C. A Case of Bordetella bronchiseptica Bacteremia in a Patient With COVID-19: Brief Report. Cureus 2021, 13, e15976. [Google Scholar] [CrossRef]
- Kadlec, K.; Kehrenberg, C.; Schwarz, S. Molecular basis of resistance to trimethoprim, chloramphenicol and sulphonamides in Bordetella bronchiseptica. J. Antimicrob. Chemother. 2005, 56, 485–490. [Google Scholar] [CrossRef] [Green Version]
- Kadlec, K.; Kehrenberg, C.; Schwarz, S. tet(A)-mediated tetracycline resistance in porcine Bordetella bronchiseptica isolates is based on plasmid-borne Tn1721 relics. J. Antimicrob. Chemother. 2006, 58, 225–227. [Google Scholar] [CrossRef] [Green Version]
- Mayer, R.L.; Impens, F. Immunopeptidomics for next-generation bacterial vaccine development. Trends Microbiol. 2021, 29, 1034–1045. [Google Scholar] [CrossRef]
- Muhammad, A.; Kassmannhuber, J.; Rauscher, M.; Falcon, A.A.; Wheeler, D.W.; Zhang, A.A.; Lubitz, P.; Lubitz, W. Subcutaneous Immunization of Dogs with Bordetella bronchiseptica Bacterial Ghost Vaccine. Front. Immunol. 2019, 10, 1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, W.; Peng, Z.; Wang, F.; Zhang, Y.; Xie, S.; Liang, W.; Hua, L.; Chen, H.; Wu, B. A Marker-Free Bordetella bronchiseptica aroA/bscN Double Deleted Mutant Confers Protection against Lethal Challenge. Vaccines 2019, 7, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, H.; Xiong, B.; Fan, G.; Cao, Z. Immunogenicity of recombinant outer membrane porin protein and protective efficacy against lethal challenge with Bordetella bronchiseptica in rabbits. J. Appl. Microbiol. 2019, 127, 1646–1655. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, S.; Li, Z.; Ma, G.; Su, Z. Construction of a stable w/o nano-emulsion as a potential adjuvant for foot and mouth disease virus vaccine. Artif. Cells Nanomed. Biotechnol. 2017, 45, 897–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, A.S.; Marrack, P. Old and new adjuvants. Curr. Opin. Immunol. 2017, 47, 44–51. [Google Scholar] [CrossRef]
- Charerntantanakul, W. Adjuvants for swine vaccines: Mechanisms of actions and adjuvant effects. Vaccine 2020, 38, 6659–6681. [Google Scholar] [CrossRef]
- Portuondo, D.L.; Batista-Duharte, A.; Ferreira, L.S.; de Andrade, C.R.; Quinello, C.; Téllez-Martínez, D.; de Aguiar Loesch, M.L.; Carlos, I.Z. Comparative Efficacy and Toxicity of Two Vaccine Candidates against Sporothrix schenckii Using Either MontanideTM Pet Gel A or Aluminum Hydroxide Adjuvants in Mice. Vaccine 2017, 35, 4430–4436. [Google Scholar] [CrossRef] [Green Version]
- Riaz, M.; Rahman, N.U.; Zia-Ul-Haq, M.; Jaffar, H.Z.E.; Manea, R. Ginseng: A Dietary Supplement as Immune-Modulator in Various Diseases. Trends Food Sci. Technol. 2019, 83, 12–30. [Google Scholar] [CrossRef]
- Patel, S.; Faraj, Y.; Duso, D.K.; Reiley, W.W.; Karlsson, E.A.; Schultz-Cherry, S.; Vajdy, M. Comparative Safety and Efficacy Profile of a Novel Oil in Water Vaccine Adjuvant Comprising Vitamins A and E and a Catechin in Protective Anti-Influenza Immunity. Nutrients 2017, 9, 516. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Wang, Y.; Maqbool, B.; Yuan, L.; He, S.; Zhang, C.; Xu, W.; Hu, S. Early IgG Response to Foot and Mouth Disease Vaccine Formulated with a Vegetable Oil Adjuvant. Vaccines 2019, 7, 143. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Wang, Y.; Guan, R.; Lu, M.; Yuan, L.; Xu, W.; Hu, S. Enhanced Immune Responses with Serum Proteomic Analysis of Hu Sheep to Foot-and-Mouth Disease Vaccine Emulsified in a Vegetable Oil Adjuvant. Vaccines 2020, 8, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chenwen, X.; Quanan, J.; Yee, H.; Yan, L.; Jiaoyu, W.; Qiang, W.; Litao, Q.; Li, N.; Guolian, B. Efficacy of Rg1-Oil Adjuvant on Inducing Immune Responses against Bordetella bronchiseptica in Rabbits. J. Immunol. Res. 2021, 2021, 8835919. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Huang, Y.; Wei, Q.; Liu, Y.; Ji, Q.; Li, K.; Bao, G. Comparative Proteomic Analysis Reveals Complex Responses to Bordetella bronchiseptica Infections in the Spleen of Rabbits. Proteomics 2020, 20, 2000117. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Petrovsky, N. Comparative Safety of Vaccine Adjuvants: A Summary of Current Evidence and Future Needs. Drug Saf. 2015, 38, 1059–1074. [Google Scholar] [CrossRef] [PubMed]
- Powell, B.S.; Andrianov, A.K.; Fusco, P.C. Polyionic Vaccine Adjuvants: Another Look at Aluminum Salts and Polyelectrolytes. Clin. Exp. Vaccine Res. 2015, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Laguna, J.; Salguero, F.J.; Pallarés, F.J.; Rodríguez-Gómez, I.M.; Barranco, I.; Carrasco, L. Acute Phase Proteins as Biomarkers in Animal Health and Welfare. In Acute Phase Proteins as Early Non-Specific Biomarkers of Human and Veterinary Diseases; InTechOpen: Rijeka, Croatia, 2011; p. 13. [Google Scholar]
- Khalil, R.H.; Al-Humadi, N. Types of Acute Phase Reactants and Their Importance in Vaccination (Review). Biomed. Rep. 2020, 12, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Caravaca, I.; Gourgues, S.F.; Rodríguez, V.; Estrada, E.D.; Cerón, J.J.; Escribano, D. Serum Acute Phase Response Induced by Different Vaccination Protocols against Circovirus Type 2 and Mycoplasma hyopneumoniae in Piglets. Res. Vet. Sci. 2017, 114, 69–73. [Google Scholar] [CrossRef]
- Pomorska-Mól, M.; Markowska-Daniel, I.; Kwit, K.; Stępniewska, K.; Pejsak, Z. C-Reactive Protein, Haptoglobin, Serum Amyloid A and Pig Major Acute Phase Protein Response in Pigs Simultaneously Infected with H1N1 Swine Influenza Virus and Pasteurella multocida. BMC Vet. Res. 2013, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Green, M.D. Acute Phase Responses to Novel, Investigational Vaccines in Toxicology Studies. Int. J. Toxicol. 2015, 34, 379–383. [Google Scholar] [CrossRef]
- Destexhe, E.; Prinsen, M.K.; van Schöll, I.; Kuper, C.F.; Garçon, N.; Veenstra, S.; Segal, L. Evaluation of C-Reactive Protein as an Inflammatory Biomarker in Rabbits for Vaccine Nonclinical Safety Studies. J. Pharmacol. Toxicol. Methods 2013, 68, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Garçon, N.; Vaughn, D.W.; Didierlaurent, A.M. Development and Evaluation of AS03, an Adjuvant System Containing α-Tocopherol and Squalene in an Oil-in-Water Emulsion. Expert Rev. Vaccines 2012, 11, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Wang, Y.; Li, Z.; Ma, X.; Cui, X.; Chi, X.; Xu, W.; Hu, S. Sunflower Seed Oil Containing Ginseng Stem–Leaf Saponins (E515-D) Is a Safe Adjuvant for Newcastle Disease Vaccine. Poult. Sci. 2020, 99, 4795–4803. [Google Scholar] [CrossRef] [PubMed]
- Yount, K.S.; Jennings-Gee, J.; Caution, K.; Fullen, A.R.; Corps, K.N.; Quataert, S.; Deora, R.; Dubey, P. Bordetella Colonization Factor A (BcfA) Elicits Protective Immunity against Bordetella bronchiseptica in the Absence of an Additional Adjuvant. Infect. Immun. 2019, 87, e00506-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.R.; Hino, A.; Yoshimoto, T.; Nagase, H.; Kato, T.; Hirokawa, K.; Matsuzawa, A.; Nariuchi, H. Impaired Delayed-Type Hypersensitivity Response in Mutant Mice Secreting Soluble CD4 without Expression of Membrane-Bound CD4. Immunology 2000, 100, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Huang, X.; Han, K.; Liu, Y.; Yang, J.; Liu, Q.; An, F.; Li, Y. Protective Immune Response against Newly Emerging Goose Tembusu Virus Infection Induced by Immunization with a Recombinant Envelope Protein. Lett. Appl. Microbiol. 2015, 61, 318–324. [Google Scholar] [CrossRef]
- Lu, Y.-J.; Barreira-Silva, P.; Boyce, S.; Powers, J.; Cavallo, K.; Behar, S.M. CD4 T Cell Help Prevents CD8 T Cell Exhaustion and Promotes Control of Mycobacterium Tuberculosis Infection. Cell Rep. 2021, 36, 109696. [Google Scholar] [CrossRef]
- Bunte, K.; Beikler, T. Th17 Cells and the IL-23/IL-17 Axis in the Pathogenesis of Periodontitis and Immune-Mediated Inflammatory Diseases. Int. J. Mol. Sci. 2019, 20, 3394. [Google Scholar] [CrossRef] [Green Version]
- Abusleme, L.; Moutsopoulos, N. IL-17: Overview and Role in Oral Immunity and Microbiome. Oral Dis. 2017, 23, 854–865. [Google Scholar] [CrossRef]
- Su, F.; Wu, Y.; Li, J.; Huang, Y.; Yu, B.; Xu, L.; Xue, Y.; Xiao, C.; Yuan, X. Escherichia Coli Heat-Labile Enterotoxin B Subunit Combined with Ginsenoside Rg1 as an Intranasal Adjuvant Triggers Type I Interferon Signaling Pathway and Enhances Adaptive Immune Responses to an Inactivated PRRSV Vaccine in ICR Mice. Vaccines 2021, 9, 266. [Google Scholar] [CrossRef]
- Wang, J.; Xie, X.; Jiang, S.; Lu, L. Immunoengineered Adjuvants for Universal Vaccines against Respiratory Viruses. Fundam. Res. 2021, 1, 189–192. [Google Scholar] [CrossRef]
- Kalita, P.; Dasgupta, A.; Sritharan, V.; Gupta, S. Nanoparticle–Drug Bioconjugate as Dual Functional Affinity Ligand for Rapid Point-of-Care Detection of Endotoxin in Water and Serum. Anal. Chem. 2015, 87, 11007–11012. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Pan, C.; Liu, Z.; Sun, P.; Hua, X.; Feng, E.; Yu, Y.; Wu, J.; Zhu, L.; Wang, H. Safety and Immunogenicity of a New Glycoengineered Vaccine against Acinetobacter Baumannii in Mice. Microb. Biotechnol. 2022, 15, 703–716. [Google Scholar] [CrossRef]
- Cibulski, S.; Rivera-Patron, M.; Suárez, N.; Pirez, M.; Rossi, S.; Yendo, A.C.; de Costa, F.; Gosmann, G.; Fett-Neto, A.; Roehe, P.M.; et al. Leaf Saponins of Quillaja Brasiliensis Enhance Long-Term Specific Immune Responses and Promote Dose-Sparing Effect in BVDV Experimental Vaccines. Vaccine 2018, 36, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Savoji, M.A.; Sereshgi, M.M.A.; Ghahari, S.M.M.; Asgarhalvaei, F.; Mahdavi, M. Formulation of HBsAg in Montanide ISA 51VG Adjuvant: Immunogenicity Study and Monitoring Long-Lived Humoral Immune Responses. Int. Immunopharmacol. 2021, 96, 107599. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yu, L.; Gu, P.; Bo, R.; Wusiman, A.; Liu, J.; Hu, Y.; Wang, D. Preparation of Lentinan-Calcium Carbonate Microspheres and Their Application as Vaccine Adjuvants. Carbohydr. Polym. 2020, 245, 116520. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.E.-S.; Gamal, W.M.; Hassan, A.I.; Mahdy, S.E.-D.; Hegazy, A.Z.; Abdel-Atty, M.M. Comparative Study on the Immunopotentiator Effect of ISA 201, ISA 61, ISA 50, ISA 206 Used in Trivalent Foot and Mouth Disease Vaccine. Vet. World 2015, 8, 1189–1198. [Google Scholar] [CrossRef]
- Lin, L.; Ibrahim, A.S.; Xu, X.; Farber, J.M.; Avanesian, V.; Baquir, B.; Fu, Y.; French, S.W.; Edwards, J.E., Jr.; Spellberg, B. Th1-Th17 Cells Mediate Protective Adaptive Immunity against Staphylococcus Aureus and Candida Albicans Infection in Mice. PLoS Pathog. 2009, 5, e1000703. [Google Scholar] [CrossRef]
- Shi, W.; Kou, Y.; Xiao, J.; Zhang, L.; Gao, F.; Kong, W.; Su, W.; Jiang, C.; Zhang, Y. Comparison of Immunogenicity, Efficacy and Transcriptome Changes of Inactivated Rabies Virus Vaccine with Different Adjuvants. Vaccine 2018, 36, 5020–5029. [Google Scholar] [CrossRef]
- Wang, Y.; Yuan, L.; Cui, X.; Xu, W.; Fang, S.; Li, Z.; Lu, M.; Wu, Y.; Ma, X.; Chi, X.; et al. Ginseng Stem-Leaf Saponins in Combination with Selenium Promote the Immune Response in Neonatal Mice with Maternal Antibody. Vaccines 2020, 8, 755. [Google Scholar] [CrossRef]
- Yuan, L.; Wang, Y.; Ma, X.; Cui, X.; Lu, M.; Guan, R.; Chi, X.; Xu, W.; Hu, S. Sunflower Seed Oil Combined with Ginseng Stem-Leaf Saponins as an Adjuvant to Enhance the Immune Response Elicited by Newcastle Disease Vaccine in Chickens. Vaccine 2020, 38, 5343–5354. [Google Scholar] [CrossRef] [PubMed]
- Lesur, A.; Schmit, P.-O.; Bernardin, F.; Letellier, E.; Brehmer, S.; Decker, J.; Dittmar, G. Highly Multiplexed Targeted Proteomics Acquisition on a TIMS-QTOF. Anal. Chem. 2021, 93, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Loginov, D.S.; Fiala, J.; Chmelik, J.; Brechlin, P.; Kruppa, G.; Novak, P. Benefits of Ion Mobility Separation and Parallel Accumulation–Serial Fragmentation Technology on TimsTOF Pro for the Needs of Fast Photochemical Oxidation of Protein Analysis. ACS Omega 2021, 6, 10352–10361. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pan, H.; Yang, H.; Wang, C.; Liu, H.; Zhou, H.; Li, P.; Li, C.; Lu, X.; Tian, Y. Rhamnolipid Enhances the Nitrogen Fixation Activity of Azotobacter Chroococcum by Influencing Lysine Succinylation. Front. Microbiol. 2021, 12, 697963. [Google Scholar] [CrossRef]
- Zhang, H.; Jiang, H.; Fan, Y.; Chen, Z.; Li, M.; Mao, Y.; Karrow, N.A.; Loor, J.J.; Moore, S.; Yang, Z. Transcriptomics and ITRAQ-Proteomics Analyses of Bovine Mammary Tissue with Streptococcus agalactiae-Induced Mastitis. J. Agric. Food Chem. 2018, 66, 11188–11196. [Google Scholar] [CrossRef]
- Li, L.; Dong, L.; Xiao, Z.; He, W.; Zhao, J.; Pan, H.; Chu, B.; Cheng, J.; Wang, H. Integrated Analysis of the Proteome and Transcriptome in a MCAO Mouse Model Revealed the Molecular Landscape during Stroke Progression. J. Adv. Res. 2020, 24, 13–27. [Google Scholar] [CrossRef]
- Guo, C.; Xu, Y.; Han, X.; Liu, X.; Xie, R.; Cheng, Z.; Fu, X. Transcriptomic and Proteomic Study on the High-Fat Diet Combined With AOM/DSS-Induced Adenomatous Polyps in Mice. Front. Oncol. 2021, 11, 736225. [Google Scholar] [CrossRef]
- Takemon, Y.; Chick, J.M.; Gyuricza, I.G.; Skelly, D.A.; Devuyst, O.; Gygi, S.P.; Churchill, G.A.; Korstanje, R. Proteomic and Transcriptomic Profiling Reveal Different Aspects of Aging in the Kidney. eLife 2021, 10, 1–21. [Google Scholar] [CrossRef]
- Yang, M.; Cao, X.; Wu, R.; Liu, B.; Ye, W.; Yue, X.; Wu, J. Comparative Proteomic Exploration of Whey Proteins in Human and Bovine Colostrum and Mature Milk Using ITRAQ-Coupled LC-MS/MS. Int. J. Food Sci. Nutr. 2017, 68, 671–681. [Google Scholar] [CrossRef]
- Yanase, Y.; Takahagi, S.; Ozawa, K.; Hide, M. The Role of Coagulation and Complement Factors for Mast Cell Activation in the Pathogenesis of Chronic Spontaneous Urticaria. Cells 2021, 10, 1759. [Google Scholar] [CrossRef]
- Harris, S.L.; Levine, A.J. The P53 Pathway: Positive and Negative Feedback Loops. Oncogene 2005, 24, 2899–2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, X.; Xu, X.; Huang, P.; Bao, G.; Liu, Y. Safety and Efficacy of the Bordetella bronchiseptica Vaccine Combined with a Vegetable Oil Adjuvant and Multi-Omics Analysis of Its Potential Role in the Protective Response of Rabbits. Pharmaceutics 2022, 14, 1434. https://doi.org/10.3390/pharmaceutics14071434
Cui X, Xu X, Huang P, Bao G, Liu Y. Safety and Efficacy of the Bordetella bronchiseptica Vaccine Combined with a Vegetable Oil Adjuvant and Multi-Omics Analysis of Its Potential Role in the Protective Response of Rabbits. Pharmaceutics. 2022; 14(7):1434. https://doi.org/10.3390/pharmaceutics14071434
Chicago/Turabian StyleCui, Xuemei, Xiangfei Xu, Pan Huang, Guolian Bao, and Yan Liu. 2022. "Safety and Efficacy of the Bordetella bronchiseptica Vaccine Combined with a Vegetable Oil Adjuvant and Multi-Omics Analysis of Its Potential Role in the Protective Response of Rabbits" Pharmaceutics 14, no. 7: 1434. https://doi.org/10.3390/pharmaceutics14071434
APA StyleCui, X., Xu, X., Huang, P., Bao, G., & Liu, Y. (2022). Safety and Efficacy of the Bordetella bronchiseptica Vaccine Combined with a Vegetable Oil Adjuvant and Multi-Omics Analysis of Its Potential Role in the Protective Response of Rabbits. Pharmaceutics, 14(7), 1434. https://doi.org/10.3390/pharmaceutics14071434