


Impacts of PEGylation and Glycosylation on the Biological Properties of Host Defense Peptide IDR1018


Abstract
:1. Introduction
2. Materials and Methods
2.1. Peptide Synthesis, Pegylation, and Glycosylation
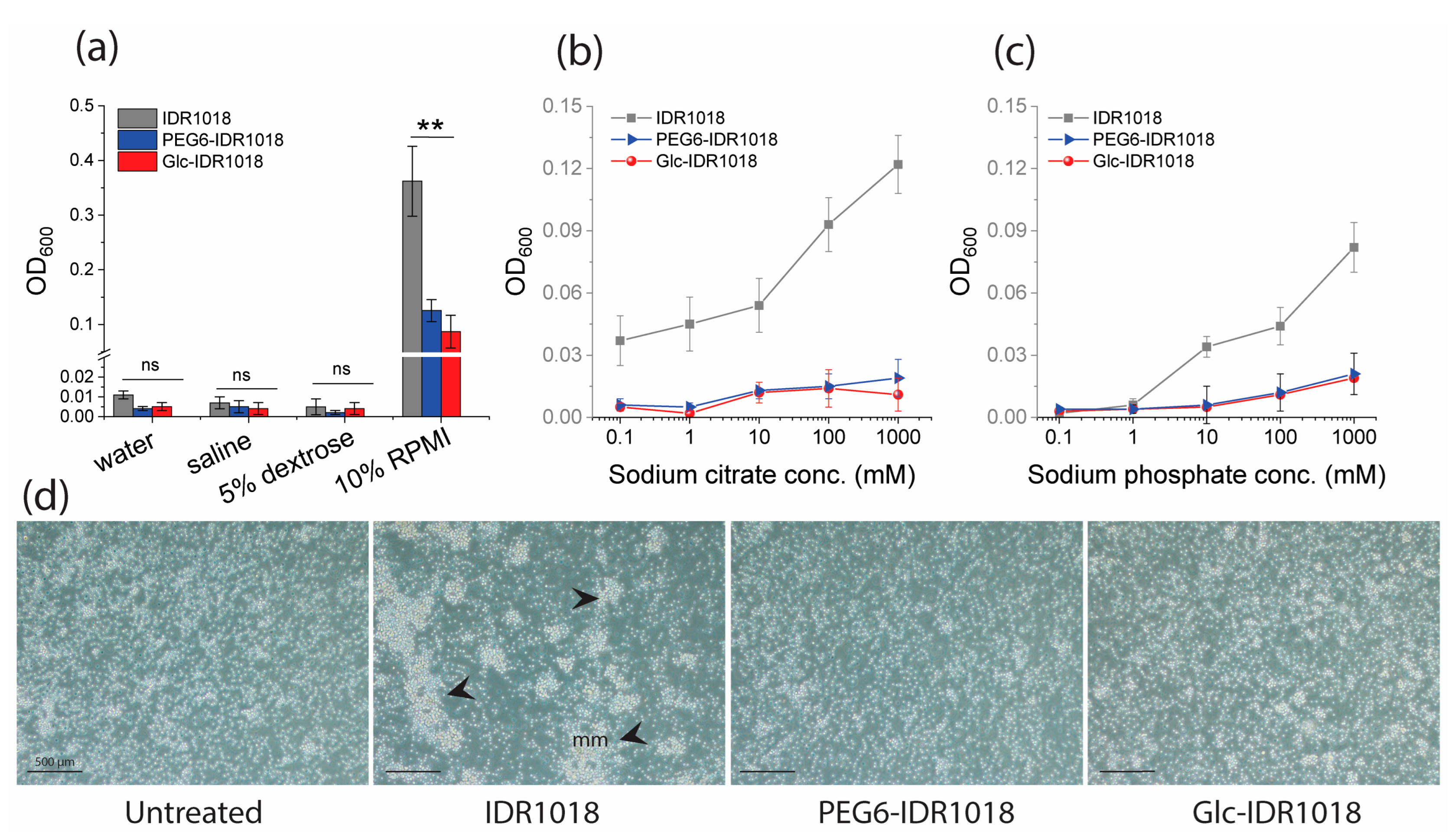
2.2. Aggregation Assay
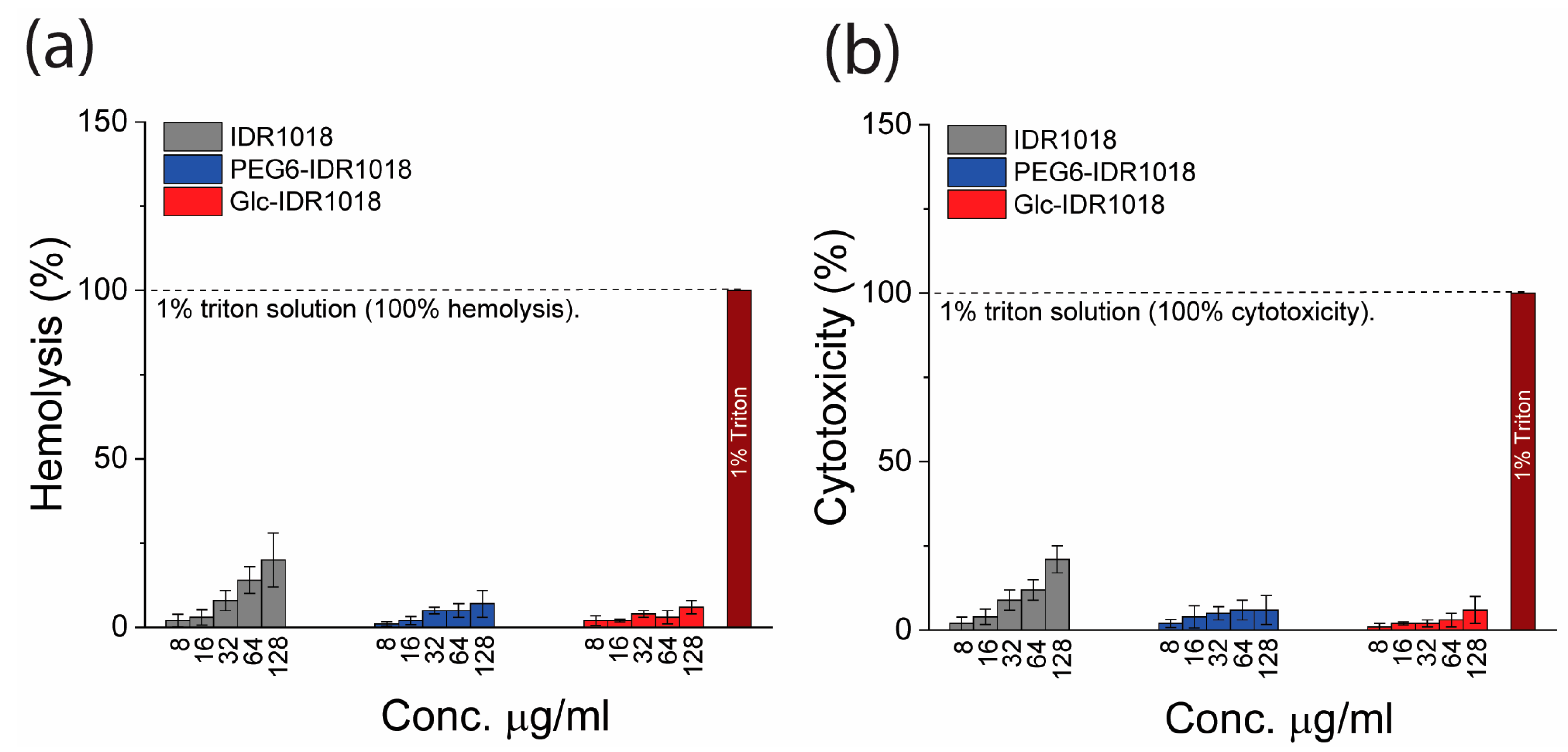
2.3. Hemolysis Test
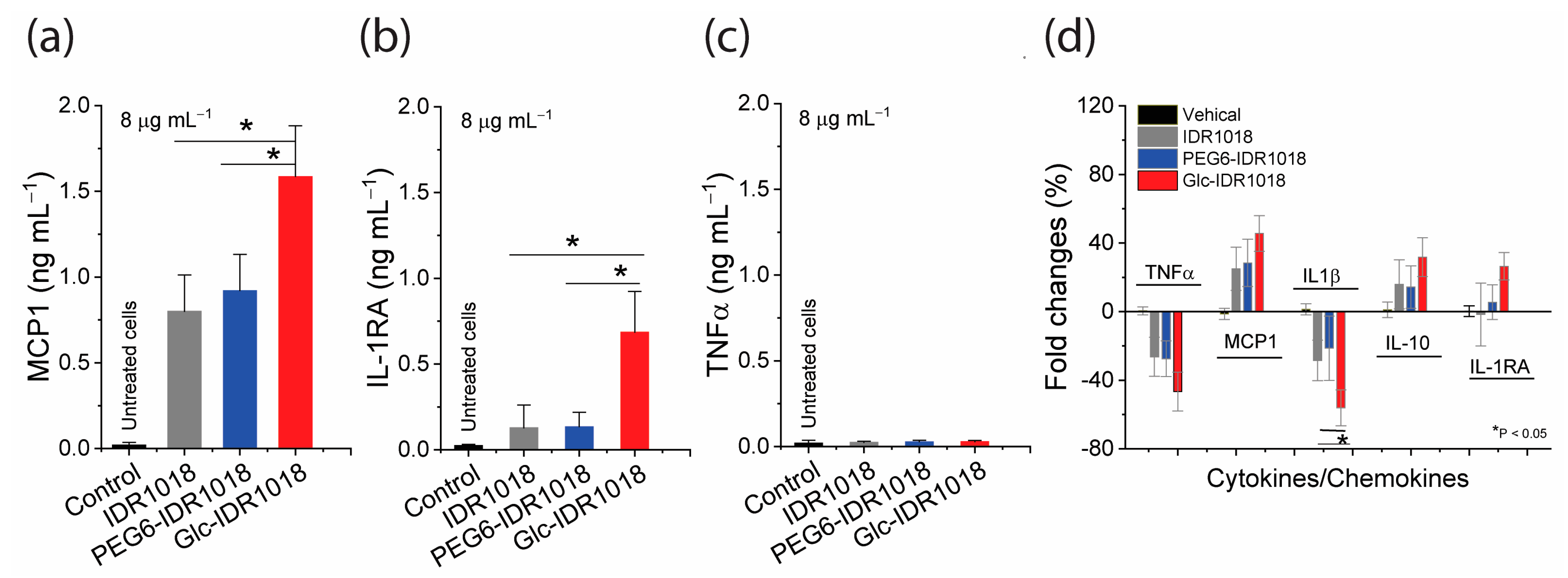
2.4. Cytotoxicity and Immunomodulatory Assays
2.5. Minimal Inhibitory Concentration (MIC)
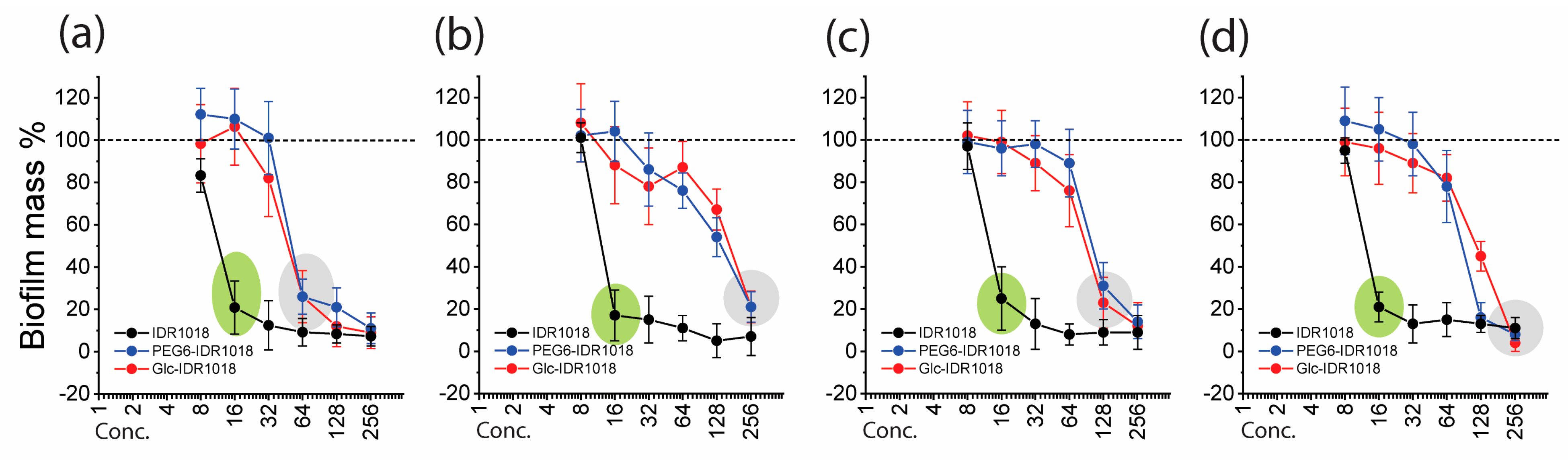
2.6. Biofilm Biomass and Biofilm Eradication Assays
2.7. Statistical Analysis
3. Results and Discussion
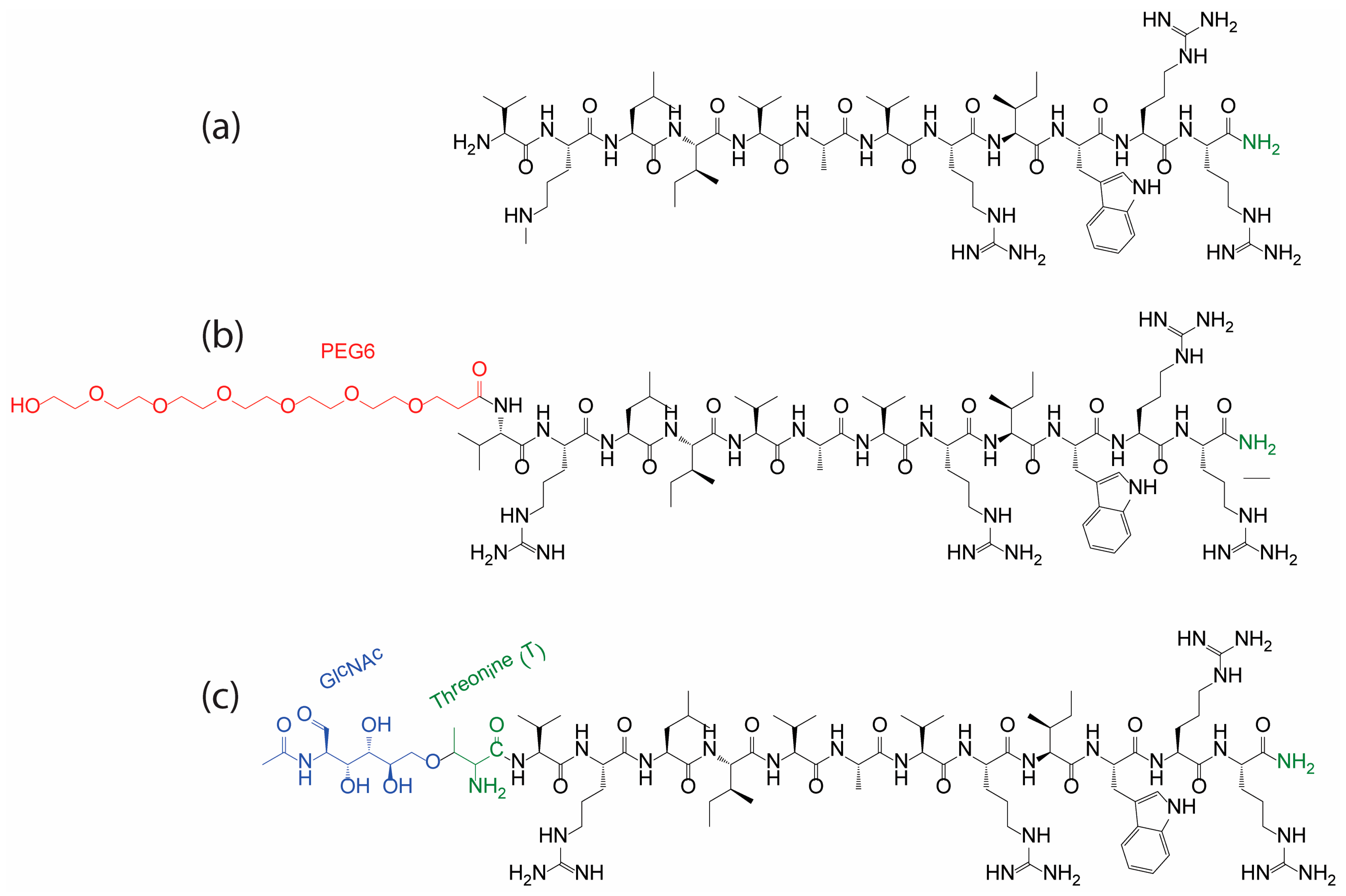
3.1. Peptide Conjugation Design
3.2. In Vitro Peptide(s) Aggregation
3.3. Hemolysis, Cytotoxicity and Immunomodulatory Activity
3.4. Antimicrobial Activity
3.5. Antibiofilm Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ikuta, K.S.; Swetschinski, L.R.; Aguilar, G.R.; Sharara, F.; Mestrovic, T.; Gray, A.P.; Weaver, N.D.; Wool, E.E.; Han, C.; Hayoon, A.G. Global mortality associated with 33 bacterial pathogens in 2019: A systematic analysis for the global burden of disease study 2019. Lancet 2022, 400, 2221–2248. [Google Scholar] [CrossRef] [PubMed]
- El-Kafrawy, S.A.; Abbas, A.T.; Oelkrug, C.; Tahoon, M.; Ezzat, S.; Zumla, A.; Azhar, E.I. IgY antibodies: The promising potential to overcome antibiotic resistance. Front. Immunol. 2023, 14, 1065353. [Google Scholar] [CrossRef] [PubMed]
- Li, W. Prospective Application of aptamer-based assays and therapeutics in bloodstream infections. Mini Rev. Med. Chem. 2020, 20, 831–840. [Google Scholar] [CrossRef]
- Sun, Y.; Meng, L.; Zhang, Y.; Zhao, D.; Lin, Y. The application of nucleic acids and nucleic acid materials in antimicrobial research. Curr. Stem Cell Res. Ther. 2021, 16, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Pino, P.; Bosco, F.; Mollea, C.; Onida, B. Antimicrobial nano-zinc oxide biocomposites for wound healing applications: A Review. Pharmaceutics 2023, 15, 970. [Google Scholar] [CrossRef]
- Akshaya, S.; Rowlo, P.K.; Dukle, A.; Nathanael, A.J. Antibacterial coatings for titanium implants: Recent trends and future perspectives. Antibiotics 2022, 11, 1719. [Google Scholar] [CrossRef]
- Sun, C.; Wang, X.; Dai, J.; Ju, Y. Metal and metal oxide nanomaterials for fighting planktonic bacteria and biofilms: A review emphasizing on mechanistic aspects. Int. J. Mol. Sci. 2022, 23, 11348. [Google Scholar] [CrossRef]
- Zha, G.F.; Preetham, H.D.; Rangappa, S.; Sharath Kumar, K.S.; Girish, Y.R.; Rakesh, K.P.; Ashrafizadeh, M.; Zarrabi, A.; Rangappa, K.S. Benzimidazole analogues as efficient arsenals in war against methicillin-resistance Staphylococcus aureus (MRSA) and its SAR studies. Bioorg. Chem. 2021, 115, 105175. [Google Scholar] [CrossRef]
- Mousavifar, L.; Roy, R. Recent development in the design of small ‘drug-like’ and nanoscale glycomimetics against Escherichia coli infections. Drug Discov. Today 2021, 26, 2124–2137. [Google Scholar] [CrossRef]
- Hemmati, F.; Rezaee, M.A.; Ebrahimzadeh, S.; Yousefi, L.; Nouri, R.; Kafil, H.S.; Gholizadeh, P. Novel strategies to combat bacterial biofilms. Mol. Biotechnol. 2021, 63, 569–586. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Alford, M.A.; Haney, E.F. Antibiofilm activity of host defence peptides: Complexity provides opportunities. Nat. Rev. Microbiol. 2021, 19, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The value of antimicrobial peptides in the age of resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef]
- Etayash, H.; Hancock, R.E.W. Host defense peptide-mimicking polymers and polymeric-brush-tethered host defense peptides: Recent developments, limitations, and potential success. Pharmaceutics 2021, 13, 1820. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Cho, B.H.; Jang, Y.S. Understanding the roles of host defense peptides in immune modulation: From antimicrobial action to potential as adjuvants. J. Microbiol. Biotechnol. 2023, 33, 288–298. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Haney, E.F.; Gill, E.E. The immunology of host defence peptides: Beyond antimicrobial activity. Nat. Rev. Immunol. 2016, 16, 321–334. [Google Scholar] [CrossRef]
- Ting, D.S.J.; Beuerman, R.W.; Dua, H.S.; Lakshminarayanan, R.; Mohammed, I. Strategies in translating the therapeutic potentials of host defense peptides. Front. Immunol. 2020, 11, 983. [Google Scholar] [CrossRef]
- Etayash, H.; Alford, M.; Akhoundsadegh, N.; Drayton, M.; Straus, S.K.; Hancock, R.E.W. Multifunctional antibiotic-host defense peptide conjugate kills bacteria, eradicates biofilms, and modulates the innate immune response. J. Med. Chem. 2021, 64, 16854–16863. [Google Scholar] [CrossRef] [PubMed]
- Etayash, H.; Qian, Y.; Pletzer, D.; Zhang, Q.; Xie, J.; Cui, R.; Dai, C.; Ma, P.; Qi, F.; Liu, R.; et al. Host defense peptide-mimicking amphiphilic β-peptide polymer (Bu:DM) exhibiting anti-biofilm, immunomodulatory, and in vivo anti-infective activity. J. Med. Chem. 2020, 63, 12921–12928. [Google Scholar] [CrossRef]
- Etayash, H.; Pletzer, D.; Kumar, P.; Straus, S.K.; Hancock, R.E.W. Cyclic derivative of host-defense peptide IDR-1018 improves proteolytic stability, suppresses inflammation, and enhances in vivo activity. J. Med. Chem. 2020, 63, 9228–9236. [Google Scholar] [CrossRef]
- Gonçalves da Costa Sousa, M.; Conceição de Almeida, G.; Martins Mota, D.C.; Andrade da Costa, R.; Dias, S.C.; Limberger, S.N.; Ko, F.; Lin, L.T.; Haney, E.F.; Etayash, H.; et al. Antibiofilm and immunomodulatory resorbable nanofibrous filing for dental pulp regenerative procedures. Bioact. Mater. 2022, 16, 173–186. [Google Scholar] [CrossRef]
- Thapa, R.K.; Diep, D.B.; Tønnesen, H.H. Topical antimicrobial peptide formulations for wound healing: Current developments and future prospects. Acta Biomater. 2020, 103, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Gaglio, S.C.; Jabalera, Y.; Montalbán-López, M.; Millán-Placer, A.C.; Lázaro-Callejón, M.; Maqueda, M.; Carrasco-Jimenez, M.P.; Laso, A.; Aínsa, J.A.; Iglesias, G.R.; et al. Embedding biomimetic magnetic nanoparticles coupled with peptide AS-48 into PLGA to treat intracellular pathogens. Pharmaceutics 2022, 14, 2744. [Google Scholar] [CrossRef] [PubMed]
- van Gent, M.E.; Ali, M.; Nibbering, P.H.; Kłodzińska, S.N. Current advances in lipid and polymeric antimicrobial peptide delivery systems and coatings for the prevention and treatment of bacterial infections. Pharmaceutics 2021, 13, 1840. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, M.; Jenssen, H.; Kindrachuk, J.; Scott, W.R.; Elliott, M.; Hilpert, K.; Cheng, J.T.; Hancock, R.E.W.; Straus, S.K. Structural studies of a peptide with immune modulating and direct antimicrobial activity. Chem. Biol. 2010, 17, 970–980. [Google Scholar] [CrossRef]
- Hilchie, A.L.; Wuerth, K.; Hancock, R.E.W. Immune modulation by multifaceted cationic host defense (antimicrobial) peptides. Nat. Chem. Biol. 2013, 9, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Rezende, S.B.; Oshiro, K.G.N.; Júnior, N.G.O.; Franco, O.L.; Cardoso, M.H. Advances on chemically modified antimicrobial peptides for generating peptide antibiotics. Chem. Commun. 2021, 57, 11578–11590. [Google Scholar] [CrossRef] [PubMed]
- Grimsey, E.; Collis, D.W.P.; Mikut, R.; Hilpert, K. The effect of lipidation and glycosylation on short cationic antimicrobial peptides. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183195. [Google Scholar] [CrossRef]
- Manteghi, R.; Pallagi, E.; Olajos, G.; Csóka, I. Pegylation and formulation strategy of anti-microbial peptide (AMP) according to the quality by design approach. Eur. J. Pharm. Sci. 2020, 144, 105197. [Google Scholar] [CrossRef]
- Bednarska, N.G.; Wren, B.W.; Willcocks, S.J. The importance of the glycosylation of antimicrobial peptides: Natural and synthetic approaches. Drug Discov. Today 2017, 22, 919–926. [Google Scholar] [CrossRef]
- Harris, J.M.; Chess, R.B. Effect of pegylation on pharmaceuticals. Nat. Rev. Drug Discov. 2003, 2, 214–221. [Google Scholar] [CrossRef]
- Bellavita, R.; Braccia, S.; Galdiero, S.; Falanga, A. Glycosylation and lipidation strategies: Approaches for improving antimicrobial peptide efficacy. Pharmaceuticals 2023, 16, 439. [Google Scholar] [CrossRef] [PubMed]
- Moradi, S.V.; Hussein, W.M.; Varamini, P.; Simerska, P.; Toth, I. Glycosylation, an effective synthetic strategy to improve the bioavailability of therapeutic peptides. Chem. Sci. 2016, 7, 2492–2500. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Wu, B.C.; Lee, K.; Hilchie, A.L.; Hancock, R.E.W. Aggregation and its influence on the immunomodulatory activity of synthetic innate defense regulator peptides. Cell Chem. Biol. 2017, 24, 969–980.e964. [Google Scholar] [CrossRef]
- Rodríguez, A.; Villegas, E.; Montoya-Rosales, A.; Rivas-Santiago, B.; Corzo, G. Characterization of antibacterial and hemolytic activity of synthetic pandinin 2 variants and their inhibition against Mycobacterium tuberculosis. PLoS ONE 2014, 9, e101742. [Google Scholar] [CrossRef] [PubMed]
- Etayash, H.; Haney, E.F.; Hancock, R.E.W. Assessing biofilm inhibition and immunomodulatory activity of small amounts of synthetic host defense peptides synthesized using SPOT-array technology. Nat. Protoc. 2021, 16, 1850–1870. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Haney, E.F.; Trimble, M.J.; Hancock, R.E.W. Microtiter plate assays to assess antibiofilm activity against bacteria. Nat. Protoc. 2021, 16, 2615–2632. [Google Scholar] [CrossRef]
- Mansour, S.C.; de la Fuente-Núñez, C.; Hancock, R.E.W. Peptide IDR-1018: Modulating the immune system and targeting bacterial biofilms to treat antibiotic-resistant bacterial infections. J. Pept. Sci. 2015, 21, 323–329. [Google Scholar] [CrossRef]
- Zou, R.; Zhu, X.; Tu, Y.; Wu, J.; Landry, M.P. Activity of antimicrobial peptide aggregates decreases with increased cell membrane embedding free energy cost. Biochemistry 2018, 57, 2606–2610. [Google Scholar] [CrossRef]
- Zai, Y.; Xi, X.; Ye, Z.; Ma, C.; Zhou, M.; Chen, X.; Siu, S.W.I.; Chen, T.; Wang, L.; Kwok, H.F. Aggregation and its influence on the bioactivities of a novel antimicrobial peptide, Temporin-PF, and its analogues. Int. J. Mol. Sci. 2021, 22, 4509. [Google Scholar] [CrossRef]
- Kumar, P.; Pletzer, D.; Haney, E.F.; Rahanjam, N.; Cheng, J.T.J.; Yue, M.; Aljehani, W.; Hancock, R.E.W.; Kizhakkedathu, J.N.; Straus, S.K. Aurein-derived antimicrobial peptides formulated with pegylated phospholipid micelles to target methicillin-resistant Staphylococcus aureus skin infections. ACS Infect. Dis. 2019, 5, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Moreira Brito, J.C.; Carvalho, L.R.; Neves de Souza, A.; Carneiro, G.; Magalhães, P.P.; Farias, L.M.; Guimarães, N.R.; Verly, R.M.; Resende, J.M.; Elena de Lima, M. PEGylation of the antimicrobial peptide LyeTx I-b maintains structure-related biological properties and improves selectivity. Front. Mol. Biosci. 2022, 9, 1001508. [Google Scholar] [CrossRef]
- Singh, S.; Papareddy, P.; Mörgelin, M.; Schmidtchen, A.; Malmsten, M. Effects of PEGylation on membrane and lipopolysaccharide interactions of host defense peptides. Biomacromolecules 2014, 15, 1337–1345. [Google Scholar] [CrossRef] [PubMed]
- Steinstraesser, L.; Hirsch, T.; Schulte, M.; Kueckelhaus, M.; Jacobsen, F.; Mersch, E.A.; Stricker, I.; Afacan, N.; Jenssen, H.; Hancock, R.E.W.; et al. Innate defense regulator peptide 1018 in wound healing and wound infection. PLoS ONE 2012, 7, e39373. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Santiago, B.; Castañeda-Delgado, J.E.; Rivas Santiago, C.E.; Waldbrook, M.; González-Curiel, I.; León-Contreras, J.C.; Enciso-Moreno, J.A.; del Villar, V.; Mendez-Ramos, J.; Hancock, R.E.W.; et al. Ability of innate defence regulator peptides IDR-1002, IDR-HH2 and IDR-1018 to protect against Mycobacterium tuberculosis infections in animal models. PLoS ONE 2013, 8, e59119. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interf. Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Yazdi, A.S.; Ghoreschi, K. The interleukin-1 family. Adv. Exp. Med. Biol. 2016, 941, 21–29. [Google Scholar]
- Haney, E.F.; Wuerth, K.C.; Rahanjam, N.; Safaei Nikouei, N.; Ghassemi, A.; Alizadeh Noghani, M.; Boey, A.; Hancock, R.E.W. Identification of an IDR peptide formulation candidate that prevents peptide aggregation and retains immunomodulatory activity. Pept. Sci. 2019, 111, e24077. [Google Scholar] [CrossRef]
- Imura, Y.; Nishida, M.; Matsuzaki, K. Action mechanism of PEGylated magainin 2 analogue peptide. Biochim. Biophys. Acta 2007, 1768, 2578–2585. [Google Scholar] [CrossRef]
- Guiotto, A.; Pozzobon, M.; Canevari, M.; Manganelli, R.; Scarin, M.; Veronese, F.M. PEGylation of the antimicrobial peptide nisin A: Problems and perspectives. Farmaco 2003, 58, 45–50. [Google Scholar] [CrossRef]
- Wu, Z.; Li, X.; Ericksen, B.; de Leeuw, E.; Zou, G.; Zeng, P.; Xie, C.; Li, C.; Lubkowski, J.; Lu, W.Y.; et al. Impact of pro segments on the folding and function of human neutrophil alpha-defensins. J. Mol. Biol. 2007, 368, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Imura, Y.; Nishida, M.; Ogawa, Y.; Takakura, Y.; Matsuzaki, K. Action mechanism of tachyplesin I and effects of PEGylation. Biochim. Biophys. Acta 2007, 1768, 1160–1169. [Google Scholar] [CrossRef]
- Kumar, P.; Takayesu, A.; Abbasi, U.; Kalathottukaren, M.T.; Abbina, S.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptide-polymer conjugates with high activity: Influence of polymer molecular weight and peptide sequence on antimicrobial activity, proteolysis, and biocompatibility. ACS Appl. Mater. Interfaces 2017, 9, 37575–37586. [Google Scholar] [CrossRef] [PubMed]
- Talat, S.; Thiruvikraman, M.; Kumari, S.; Kaur, K.J. Glycosylated analogs of formaecin I and drosocin exhibit differential pattern of antibacterial activity. Glycoconj. J. 2011, 28, 537–555. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yang, N.; Teng, D.; Mao, R.; Hao, Y.; Wang, X.; Wang, J. C-terminal mini-PEGylation of a marine peptide N6 had potent antibacterial and anti-inflammatory properties against Escherichia coli and Salmonella strains in vitro and in vivo. BMC Microbiol. 2022, 22, 128. [Google Scholar] [CrossRef]
- Falciani, C.; Lozzi, L.; Scali, S.; Brunetti, J.; Bracci, L.; Pini, A. Site-specific pegylation of an antimicrobial peptide increases resistance to Pseudomonas aeruginosa elastase. Amino Acids 2014, 46, 1403–1407. [Google Scholar] [CrossRef]
- Morris, C.J.; Beck, K.; Fox, M.A.; Ulaeto, D.; Clark, G.C.; Gumbleton, M. Pegylation of antimicrobial peptides maintains the active peptide conformation, model membrane interactions, and antimicrobial activity while improving lung tissue biocompatibility following airway delivery. Antimicrob. Agents Chemother. 2012, 56, 3298–3308. [Google Scholar] [CrossRef]
- Coltart, D.M.; Royyuru, A.K.; Williams, L.J.; Glunz, P.W.; Sames, D.; Kuduk, S.D.; Schwarz, J.B.; Chen, X.T.; Danishefsky, S.J.; Live, D.H. Principles of mucin architecture: Structural studies on synthetic glycopeptides bearing clustered mono-, di-, tri-, and hexasaccharide glycodomains. J. Am. Chem. Soc. 2002, 124, 9833–9844. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide and Derivatives | Sequence | HPLC (Ret. Time (min)) | MW (Calc.) | MW (Obs.) | Purity (%) |
|---|---|---|---|---|---|
| IDR1018 | VRLIVAVRIWRR-NH2 | 9.1 | 1535.8 | 1535.8 | >95.0 |
| PEG6-IDR1018 | PEG6-VRLIVAVRIWRR-NH2 | 17.9 | 1872.3 | 1872.6 | >95.0 |
| Glc-IDR1018 | T(GlcNAc)VRLIVAVRIWRR-NH2 | 14.3 | 1841.4 | 1841.3 | >92.0 |
| Bacteria | MIC (μg mL−1) | ||
|---|---|---|---|
| IDR1018 | PEG6-IDR1018 | Glc-IDR1018 | |
| S. aureus MRSA0017 | 16 | 32 | 64 |
| P. aeruginosa PA14 | 16 | 64 | 128 |
| E. coli ATTC489 | 16 | 64 | 64 |
| A. baumannii 5015 | 16 | 64 | 64 |
| Bacteria | MBEC (μg mL−1) | ||
|---|---|---|---|
| IDR1018 | PEG6-IDR1018 | Glc-IDR1018 | |
| S. aureus MRSA0017 | 16 | 64 | 64 |
| P. aeruginosa PA14 | 16 | 256 | 256 |
| E. coli ATTC489 | 16 | 128 | 128 |
| A. baumannii 5015 | 16 | 128 | 256 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Etayash, H.; Yip, F.; Hancock, R.E.W. Impacts of PEGylation and Glycosylation on the Biological Properties of Host Defense Peptide IDR1018. Pharmaceutics 2023, 15, 1391. https://doi.org/10.3390/pharmaceutics15051391
Etayash H, Yip F, Hancock REW. Impacts of PEGylation and Glycosylation on the Biological Properties of Host Defense Peptide IDR1018. Pharmaceutics. 2023; 15(5):1391. https://doi.org/10.3390/pharmaceutics15051391
Chicago/Turabian StyleEtayash, Hashem, Fione Yip, and Robert E. W. Hancock. 2023. "Impacts of PEGylation and Glycosylation on the Biological Properties of Host Defense Peptide IDR1018" Pharmaceutics 15, no. 5: 1391. https://doi.org/10.3390/pharmaceutics15051391
APA StyleEtayash, H., Yip, F., & Hancock, R. E. W. (2023). Impacts of PEGylation and Glycosylation on the Biological Properties of Host Defense Peptide IDR1018. Pharmaceutics, 15(5), 1391. https://doi.org/10.3390/pharmaceutics15051391