


Novel Anti-Trop2 Nanobodies Disrupt Receptor Dimerization and Inhibit Tumor Cell Growth
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Antibodies
2.2. Phage Biopanning
2.3. Phage ELISA
2.4. Anti-Trop2 Nanobodies Expression and Purification
2.5. SDS-PAGE
2.6. Size-Exclusion Chromatography
2.7. ELISA
2.8. Biolayer Interferometry (BLI)
2.9. Cell-Surface Binding by Flow Cytometry
2.10. Receptor Internalization Assay by Flow Cytometry
2.11. Cell Viability Assay
2.12. RNA Interference Analysis
2.13. Cell Migration Assay
2.14. Epitope Analysis
2.15. Co-Immunoprecipitation (Co-IP)
2.16. AlphaFold 3 Antibody–Antigen Modeling
2.17. Statistical Analysis
3. Result
3.1. Screening and Characterization of Anti-Trop2 Nanobodies
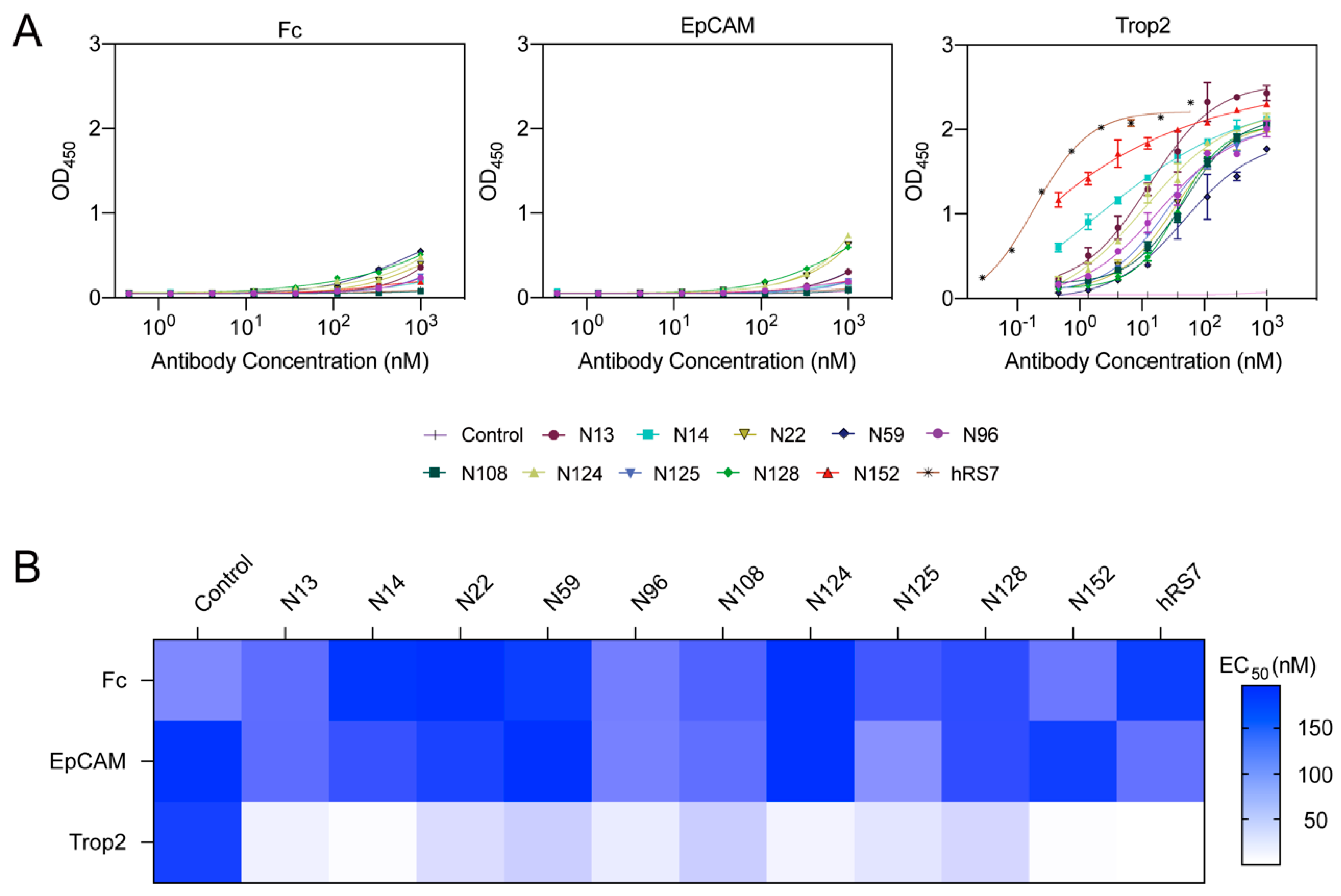
3.2. N14 and N152 Exert Binding Specificity and Activity towards Trop2
3.3. N14 and N152 Induce Receptor Internalization and Inhibit Receptor Recycling in Trop2-Positive Tumor Cells
3.4. N14 and N152 Inhibit Cell Proliferation and Migration
3.5. N14 and N152 Recognize Multiple Domains of Trop2-ECD and Inhibit Trop2 Dimerization
3.6. N14 Binds to the Site on Cis-Interface
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Tsukahara, Y.; Tanaka, M.; Miyajima, A. TROP2 expressed in the trunk of the ureteric duct regulates branching morphogenesis during kidney development. PLoS ONE 2011, 6, e28607. [Google Scholar] [CrossRef] [PubMed]
- Moretto, R.; Germani, M.M.; Giordano, M.; Conca, V.; Proietti, A.; Niccoli, C.; Pietrantonio, F.; Lonardi, S.; Tamburini, E.; Zaniboni, A.; et al. Trop-2 and Nectin-4 immunohistochemical expression in metastatic colorectal cancer: Searching for the right population for drugs’ development. Br. J. Cancer 2023, 128, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chao, Y.; Zhang, C.; Hu, W.; Huang, Y.; Lv, Y.; Liu, B.; Ji, D.; Liu, M.; Yang, B.; et al. TROP2 translation mediated by dual m(6)A/m(7)G RNA modifications promotes bladder cancer development. Cancer Lett. 2023, 566, 216246. [Google Scholar] [CrossRef] [PubMed]
- Cubas, R.; Li, M.; Chen, C.; Yao, Q. Trop2: A possible therapeutic target for late stage epithelial carcinomas. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2009, 1796, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Deng, J.; Yuan, Y.; Chen, W.; Sun, W.; Wang, Y.; Huang, H.; Liang, B.; Ming, T.; Wen, J.; et al. Advances in Trop2-targeted therapy: Novel agents and opportunities beyond breast cancer. Pharmacol. Ther. 2022, 239, 108296. [Google Scholar] [CrossRef]
- Basu, A.; Goldenberg, D.M.; Stein, R. The epithelial/carcinoma antigen EGP-1, recognized by monoclonal antibody RS7-3G11, is phosphorylated on serine 303. Int. J. Cancer 1995, 62, 472–479. [Google Scholar] [CrossRef]
- Berridge, M.J. Calcium signalling and cell proliferation. BioEssays 1995, 17, 491–500. [Google Scholar] [CrossRef]
- Cubas, R.; Zhang, S.; Li, M.; Chen, C.; Yao, Q. Trop2 expression contributes to tumor pathogenesis by activating the ERK MAPK pathway. Mol. Cancer 2010, 9, 253. [Google Scholar] [CrossRef]
- Lupu, V.D.; Kaznacheyeva, E.; Krishna, U.M.; Falck, J.R.; Bezprozvanny, I. Functional coupling of phosphatidylinositol 4,5-bisphosphate to inositol 1,4,5-trisphosphate receptor. J. Biol. Chem. 1998, 273, 14067–14070. [Google Scholar] [CrossRef]
- Stoyanova, T.; Goldstein, A.S.; Cai, H.; Drake, J.M.; Huang, J.; Witte, O.N. Regulated proteolysis of Trop2 drives epithelial hyperplasia and stem cell self-renewal via β-catenin signaling. Genes Dev. 2012, 26, 2271–2285. [Google Scholar] [CrossRef]
- Nakatsukasa, M.; Kawasaki, S.; Yamasaki, K.; Fukuoka, H.; Matsuda, A.; Tsujikawa, M.; Tanioka, H.; Nagata-Takaoka, M.; Hamuro, J.; Kinoshita, S. Tumor-associated calcium signal transducer 2 is required for the proper subcellular localization of claudin 1 and 7: Implications in the pathogenesis of gelatinous drop-like corneal dystrophy. Am. J. Pathol. 2010, 177, 1344–1355. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Lv, A.; Deng, Q.; Zhang, G.; Hu, X.; Cui, H. TROP2 promotes the proliferation and metastasis of glioblastoma cells by activating the JAK2/STAT3 signaling pathway. Oncol. Rep. 2019, 41, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Wu, Y.Y.; Wu, J.Y.; Lin, T.C.; Wu, C.T.; Chang, Y.L.; Jou, Y.S.; Hong, T.M.; Yang, P.C. TROP2 is epigenetically inactivated and modulates IGF-1R signalling in lung adenocarcinoma. EMBO Mol. Med. 2012, 4, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Tang, Q.; Jia, L.; Chen, Y.; Lin, L.; Kuai, X.; Gong, A.; Feng, Z. TROP2 increases growth and metastasis of human oral squamous cell carcinoma through activation of the PI3K/Akt signaling pathway. Int. J. Mol. Med. 2019, 44, 2161–2170. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhu, X.; Zhao, L.; Li, X.; Cheng, D.; Feng, K. Tumor-associated calcium signal transducer 2 regulates neovascularization of non-small-cell lung cancer via activating ERK1/2 signaling pathway. Tumour Biol. 2017, 39, 1010428317694324. [Google Scholar] [CrossRef]
- Iwamoto, S.; Mori, Y.; Yamashita, T.; Ojima, K.; Akita, K.; Togano, S.; Kushiyama, S.; Yashiro, M.; Yatera, Y.; Yamaguchi, T.; et al. Trophoblast cell surface antigen-2 phosphorylation triggered by binding of galectin-3 drives metastasis through down-regulation of E-cadherin. J. Biol. Chem. 2023, 299, 104971. [Google Scholar] [CrossRef]
- Jiang, H.; Qin, X.; Wang, Q.; Xu, Q.; Wang, J.; Wu, Y.; Chen, W.; Wang, C.; Zhang, T.; Xing, D.; et al. Application of carbohydrates in approved small molecule drugs: A review. Eur. J. Med. Chem. 2021, 223, 113633. [Google Scholar] [CrossRef]
- Ma, Q.; Jiang, H.; Ma, L.; Zhao, G.; Xu, Q.; Guo, D.; He, N.; Liu, H.; Meng, Z.; Liu, J.; et al. The moonlighting function of glycolytic enzyme enolase-1 promotes choline phospholipid metabolism and tumor cell proliferation. Proc. Natl. Acad. Sci. USA 2023, 120, e2209435120. [Google Scholar] [CrossRef]
- Liu, X.; Ma, L.; Li, J.; Sun, L.; Yang, Y.; Liu, T.; Xing, D.; Yan, S.; Zhang, M. Trop2-targeted therapies in solid tumors: Advances and future directions. Theranostics 2024, 14, 3674–3692. [Google Scholar] [CrossRef]
- Petrylak, D.P.; Tagawa, S.T.; Jain, R.K.; Bupathi, M.; Balar, A.V.; Rezazadeh, A.; George, S.; Palmbos, P.L.; Nordquist, L.T.; Davis, N.B.; et al. Primary analysis of TROPHY-U-01 cohort 2, a phase 2 study of sacituzumab govitecan (SG) in platinum (PT)-ineligible patients (pts) with metastatic urothelial cancer (mUC) that progressed after prior checkpoint inhibitor (CPI) therapy. J. Clin. Oncol. 2023, 41, 520. [Google Scholar] [CrossRef]
- Rugo, H.S.; Bardia, A.; Marmé, F.; Cortes, J.; Schmid, P.; Loirat, D.; Tredan, O.; Ciruelos, E.; Dalenc, F.; Pardo, P.G.; et al. Primary results from TROPiCS-02: A randomized phase 3 study of sacituzumab govitecan (SG) versus treatment of physician’s choice (TPC) in patients (Pts) with hormone receptor–positive/HER2-negative (HR+/HER2-) advanced breast cancer. J. Clin. Oncol. 2022, 40 (Suppl. S17), LBA1001-LBA. [Google Scholar] [CrossRef]
- Tagawa, S.T.; Balar, A.V.; Petrylak, D.P.; Kalebasty, A.R.; Loriot, Y.; Fléchon, A.; Jain, R.K.; Agarwal, N.; Bupathi, M.; Barthelemy, P.; et al. TROPHY-U-01: A Phase II Open-Label Study of Sacituzumab Govitecan in Patients With Metastatic Urothelial Carcinoma Progressing After Platinum-Based Chemotherapy and Checkpoint Inhibitors. J. Clin. Oncol. 2021, 39, 2474–2485. [Google Scholar] [CrossRef] [PubMed]
- Tagawa, S.T.; Faltas, B.M.; Lam, E.T.; Saylor, P.J.; Bardia, A.; Hajdenberg, J.; Morgans, A.K.; Lim, E.A.; Kalinsky, K.; Simpson, P.S.; et al. Sacituzumab govitecan (IMMU-132) in patients with previously treated metastatic urothelial cancer (mUC): Results from a phase I/II study. J. Clin. Oncol. 2019, 37 (Suppl. S7), 354. [Google Scholar] [CrossRef]
- Garon, E.; Johnson, M.; Lisberg, A.; Spira, A.; Yamamoto, N.; Heist, R.; Sands, J.; Yoh, K.; Meric-Bernstam, F.; Kitazono, S.; et al. MA03.02 TROPION-PanTumor01: Updated Results From the NSCLC Cohort of the Phase 1 Study of Datopotamab Deruxtecan in Solid Tumors. J. Thorac. Oncol. 2021, 16, S892–S893. [Google Scholar] [CrossRef]
- Bardia, A.; Messersmith, W.; Kio, E.; Berlin, J.; Vahdat, L.; Masters, G.; Moroose, R.; Santin, A.; Kalinsky, K.; Picozzi, V.; et al. Sacituzumab govitecan, a Trop-2-directed antibody-drug conjugate, for patients with epithelial cancer: Final safety and efficacy results from the phase I/II IMMU-132-01 basket trial. Ann. Oncol. 2021, 32, 746–756. [Google Scholar] [CrossRef]
- Kalinsky, K.; Diamond, J.; Vahdat, L.; Tolaney, S.; Juric, D.; O’SHaughnessy, J.; Moroose, R.; Mayer, I.; Abramson, V.; Goldenberg, D.; et al. Sacituzumab govitecan in previously treated hormone receptor-positive/HER2-negative metastatic breast cancer: Final results from a phase I/II, single-arm, basket trial. Ann. Oncol. 2020, 31, 1709–1718. [Google Scholar] [CrossRef]
- Bardia, A.; Mayer, I.A.; Vahdat, L.T.; Tolaney, S.M.; Isakoff, S.J.; Diamond, J.R.; O’Shaughnessy, J.; Moroose, R.L.; Santin, A.D.; Abramson, V.G.; et al. Sacituzumab Govitecan-hziy in Refractory Metastatic Triple-Negative Breast Cancer. N. Engl. J. Med. 2019, 380, 741–751. [Google Scholar] [CrossRef]
- Bardia, A.; Hurvitz, S.A.; Tolaney, S.M.; Loirat, D.; Punie, K.; Oliveira, M.; Brufsky, A.; Sardesai, S.D.; Kalinsky, K.; Zelnak, A.B.; et al. Sacituzumab Govitecan in Metastatic Triple-Negative Breast Cancer. N. Engl. J. Med. 2021, 384, 1529–1541. [Google Scholar] [CrossRef]
- Krop, I.; Juric, D.; Shimizu, T.; Tolcher, A.; Spira, A.; Mukohara, T.; Lisberg, A.E.; Kogawa, T.; Papadopoulos, K.P.; Hamilton, E.; et al. Abstract GS1-05: Datopotamab deruxtecan in advanced/metastatic HER2- breast cancer: Results from the phase 1 TROPION-PanTumor01 study. Cancer Res. 2022, 82 (Suppl. S4), GS1-05. [Google Scholar] [CrossRef]
- Shaffer, C. Trop2 deal heats up antibody–drug conjugate space in cancer. Nat. Biotechnol. 2021, 39, 128–130. [Google Scholar] [CrossRef]
- Coates, J.T.; Sun, S.; Leshchiner, I.; Thimmiah, N.; Martin, E.E.; McLoughlin, D.; Danysh, B.P.; Slowik, K.; Jacobs, R.A.; Rhrissorrakrai, A.; et al. Parallel Genomic Alterations of Antigen and Payload Targets Mediate Polyclonal Acquired Clinical Resistance to Sacituzumab Govitecan in Triple-Negative Breast Cancer. Cancer Discov. 2021, 11, 2436–2445. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, J.; Deng, J.; Zhao, J.; Zhao, G.; Zhang, T.; Jiang, H.; Liang, B.; Xing, D.; Wang, J. Targeting Trop2 in solid tumors: A look into structures and novel epitopes. Front. Immunol. 2023, 14, 1332489. [Google Scholar] [CrossRef] [PubMed]
- Kamble, P.R.; Patkar, S.R.; Breed, A.A.; Pathak, B.R. N-glycosylation status of Trop2 impacts its surface density, interaction with claudin-7 and exosomal release. Arch. Biochem. Biophys. 2021, 714, 109084. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Zhang, H.; Jiang, M.; Chai, Y.; Qi, J.; Gao, G.F.; Tan, S. Structural insights into the cis and trans assembly of human trophoblast cell surface antigen 2. iScience 2021, 24, 103190. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Perrone, E.; Zeybek, B.; Bellone, S.; Tymon-Rosario, J.; Altwerger, G.; Menderes, G.; Feinberg, J.; Haines, K.; Karger, M.E.M.; et al. In vitro and in vivo activity of sacituzumab govitecan, an antibody-drug conjugate targeting trophoblast cell-surface antigen 2 (Trop-2) in uterine serous carcinoma. Gynecol. Oncol. 2020, 156, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Truong, A.; Feng, N.; Sayegh, D.; Mak, B.; O’Reilly, K.; Fung, S.W.; Ceric, N.; Hahn, S.; Pereira, D.; Findlay, H.; et al. AR47A6.4.2, a functional naked monoclonal antibody targeting Trop-2, demonstrates in vivo efficacy in human pancreatic, colon, breast and prostate cancer models. Mol. Cancer Ther. 2007, 6 (Suppl. S11), PR-12. [Google Scholar]
- Trerotola, M.; Guerra, E.; Ali, Z.; Aloisi, A.L.; Ceci, M.; Simeone, P.; Acciarito, A.; Zanna, P.; Vacca, G.; D’AMore, A.; et al. Trop-2 cleavage by ADAM10 is an activator switch for cancer growth and metastasis. Neoplasia 2021, 23, 415–428. [Google Scholar] [CrossRef]
- Alberti, S.; Miotti, S.; Stella, M.; Klein, C.E.; Fornaro, M.; Menard, S.; Colnaghi, M.I. Biochemical characterization of Trop-2, a cell surface molecule expressed by human carcinomas: Formal proof that the monoclonal antibodies T16 and MOv-16 recognize Trop-2. Hybridoma 1992, 11, 539–545. [Google Scholar] [CrossRef]
- Ikeda, M.; Yamaguchi, M.; Kato, K.; Nakamura, K.; Shiina, S.; Ichikawa-Ando, T.; Misaka, H.; Myojo, K.; Nakamura, K.; Sugimoto, Y.; et al. Pr1E11, a novel anti-TROP-2 antibody isolated by adenovirus-based antibody screening, recognizes a unique epitope. Biochem. Biophys. Res. Commun. 2015, 458, 877–882. [Google Scholar] [CrossRef]
- Kaufmann, R.; Hainzl, A.; Sterry, W.; Alberti, S.; Klein, C.E. In vivo targeting of integrin receptors in human skin xenografts by intravenously applied antibodies. Arch. Dermatol. Res. 1994, 286, 6–11. [Google Scholar] [CrossRef]
- Stepan, L.P.; Trueblood, E.S.; Hale, K.; Babcook, J.; Borges, L.; Sutherland, C.L. Expression of Trop2 cell surface glycoprotein in normal and tumor tissues: Potential implications as a cancer therapeutic target. J. Histochem. Cytochem. 2011, 59, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Trerotola, M.; Cantanelli, P.; Guerra, E.; Tripaldi, R.; Aloisi, A.L.; Bonasera, V.; Lattanzio, R.; de Lange, R.; Weidle, U.H.; Piantelli, M.; et al. Upregulation of Trop-2 quantitatively stimulates human cancer growth. Oncogene 2013, 32, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Guerra, E.; Trerotola, M.; Relli, V.; Lattanzio, R.; Ceci, M.; Boujnah, K.; Pantalone, L.; Di Pietro, R.; Iezzi, M.; Tinari, N.; et al. The 2EF Antibody Targets a Unique N-Terminal Epitope of Trop-2 and Enhances the In Vivo Activity of the Cancer-Selective 2G10 Antibody. Cancers 2023, 15, 3721. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Luan, L.; Liu, X.; Jiang, D.; Deng, J.; Xu, J.; Yuan, Y.; Xing, J.; Chen, B.; Xing, D.; et al. A novel nanobody-based HER2-targeting antibody exhibits potent synergistic antitumor efficacy in trastuzumab-resistant cancer cells. Front. Immunol. 2023, 14, 1292839. [Google Scholar] [CrossRef]
- Verhaar, E.R.; Woodham, A.W.; Ploegh, H.L. Nanobodies in cancer. Semin. Immunol. 2021, 52, 101425. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, Y.; Lin, J.; Wu, S.; Lv, H.; Ji, X.; Wang, S. Identification and Characterization of Specific Nanobodies against Trop-2 for Tumor Targeting. Int. J. Mol. Sci. 2022, 23, 7942. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, L.; Liu, X.; Zhang, J.; Yin, Y.; Luan, L.; Jiang, D.; Yang, X.; Li, L.; Xiong, H.; et al. A potent synthetic nanobody with broad-spectrum activity neutralizes SARS-CoV-2 virus and the Omicron variant BA.1 through a unique binding mode. J. Nanobiotechnol. 2022, 20, 411. [Google Scholar] [CrossRef]
- Szala, S.; Froehlich, M.; Scollon, M.; Kasai, Y.; Steplewski, Z.; Koprowski, H.; Linnenbach, A.J. Molecular cloning of cDNA for the carcinoma-associated antigen GA733-2. Proc. Natl. Acad. Sci. USA 1990, 87, 3542–3546. [Google Scholar] [CrossRef]
- Balzar, M.; Winter, M.; de Boer, C.; Litvinov, S. The biology of the 17–1A antigen (Ep-CAM). J. Mol. Med. 1999, 77, 699–712. [Google Scholar] [CrossRef]
- Stein, R.; Govindan, S.V.; Mattes, M.J.; Shih, L.B.; Griffiths, G.L.; Hansen, H.J.; Goldenberg, D.M. Targeting human cancer xenografts with monoclonal antibodies labeled using radioiodinated, diethylenetriaminepentaacetic acid-appended peptides. Clin. Cancer Res. 1999, 5 (Suppl. S10), 3079s–3087s. [Google Scholar]
- Stein, R.; Basu, A.; Chen, S.; Shih, L.B.; Goldenberg, D.M. Specificity and properties of MAb RS7-3G11 and the antigen defined by this pancarcinoma monoclonal antibody. Int. J. Cancer 1993, 55, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Govindan, S.V.; Stein, R.; Qu, Z.; Chen, S.; Andrews, P.; Ma, H.; Hansen, H.J.; Griffiths, G.L.; Horak, I.D.; Goldenberg, D.M. Preclinical therapy of breast cancer with a radioiodinated humanized anti-EGP-1 monoclonal antibody: Advantage of a residualizing iodine radiolabel. Breast Cancer Res. Treat. 2004, 84, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Pavšič, M. Trop2 Forms a Stable Dimer with Significant Structural Differences within the Membrane-Distal Region as Compared to EpCAM. Int. J. Mol. Sci. 2021, 22, 10640. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.S.; Balaya, R.D.A.; Parate, S.S.; Dagamajalu, S.; Prasad, T.S.K.; Shetty, R.; Raju, R. An assembly of TROP2-mediated signaling events. J. Cell Commun. Signal. 2023, 17, 1105–1111. [Google Scholar] [CrossRef]
- Abazari, M.A.; Soltani, M.; Kashkooli, F.M. Targeted nano-sized drug delivery to heterogeneous solid tumor microvasculatures: Implications for immunoliposomes exhibiting bystander killing effect. Phys. Fluids 2023, 35, 011905. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Bordeau, B.M.; Balthasar, J.P. Use of Payload Binding Selectivity Enhancers to Improve Therapeutic Index of Maytansinoid–Antibody–Drug Conjugates. Mol. Cancer Ther. 2023, 22, 1332–1342. [Google Scholar] [CrossRef]
- Bordeau, B.M.; Nguyen, T.D.; Polli, J.R.; Chen, P.; Balthasar, J.P. Payload-Binding Fab Fragments Increase the Therapeutic Index of MMAE Antibody-Drug Conjugates. Mol. Cancer Ther. 2023, 22, 459–470. [Google Scholar] [CrossRef]
- van Rij, C.M.; Frielink, C.; Goldenberg, D.M.; Sharkey, R.M.; Lütje, S.; McBride, W.J.; Oyen, W.J.; Boerman, O.C. Pretargeted Radioimmunotherapy of Prostate Cancer with an Anti-TROP-2×Anti-HSG Bispecific Antibody and a 177Lu-Labeled Peptide. Cancer Biother. Radiopharm. 2014, 29, 323–329. [Google Scholar] [CrossRef]
- Chen, W.; Li, M.; Younis, M.H.; Barnhart, T.E.; Jiang, D.; Sun, T.; Lang, J.M.; Engle, J.W.; Zhou, M.; Cai, W. ImmunoPET of trophoblast cell-surface antigen 2 (Trop-2) expression in pancreatic cancer. Eur. J. Nucl. Med. 2022, 49, 861–870. [Google Scholar] [CrossRef]
- van Rij, C.M.; Frielink, C.; Goldenberg, D.M.; Sharkey, R.M.; Franssen, G.M.; Lütje, S.; McBride, W.J.; Oyen, W.J.G.; Boerman, O.C. Pretargeted immunoPET of prostate cancer with an anti-TROP-2 x anti-HSG bispecific antibody in mice with PC3 xenografts. Mol. Imaging Biol. 2015, 17, 94–101. [Google Scholar] [CrossRef] [PubMed]
- van Rij, C.M.; Lütje, S.; Frielink, C.; Sharkey, R.M.; Goldenberg, D.M.; Franssen, G.M.; McBride, W.J.; Rossi, E.A.; Oyen, W.J.G.; Boerman, O.C. Pretargeted immuno-PET and radioimmunotherapy of prostate cancer with an anti-TROP-2 x anti-HSG bispecific antibody. Eur. J. Nucl. Med. 2013, 40, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Lütje, S.; Rijpkema, M.; Goldenberg, D.M.; van Rij, C.M.; Sharkey, R.M.; McBride, W.J.; Franssen, G.M.; Frielink, C.; Helfrich, W.; Oyen, W.J.; et al. Pretargeted dual-modality immuno-SPECT and near-infrared fluorescence imaging for image-guided surgery of prostate cancer. Cancer Res. 2014, 74, 6216–6223. [Google Scholar] [CrossRef] [PubMed]
- D’HUyvetter, M.; Vincke, C.; Xavier, C.; Aerts, A.; Impens, N.; Baatout, S.; De Raeve, H.; Muyldermans, S.; Caveliers, V.; Devoogdt, N.; et al. Targeted radionuclide therapy with A 177Lu-labeled anti-HER2 nanobody. Theranostics 2014, 4, 708–720. [Google Scholar] [CrossRef]
- Gainkam, L.O.T.; Huang, L.; Caveliers, V.; Keyaerts, M.; Hernot, S.; Vaneycken, I.; Vanhove, C.; Revets, H.; De Baetselier, P.; Lahoutte, T. Comparison of the biodistribution and tumor targeting of two 99mTc-labeled anti-EGFR nanobodies in mice, using pinhole SPECT/micro-CT. J. Nucl. Med. 2008, 49, 788–795. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kon (1/Ms) | Kdis (1/s) | KD (nM) | |
|---|---|---|---|
| N13 | (4.45 ± 0.05) × 103 | (2.34 ± 0.08) × 10−4 | 52.6 ± 1.79 |
| N14 | (1.14 ± 0.04) × 104 | (2.03 ± 0.09) × 10−4 | 18.0 ± 0.80 |
| N22 | (1.44 ± 0.02) × 104 | (2.12 ± 0.03) × 10−3 | 147.3 ± 3.02 |
| N59 | (1.29 ± 0.06) × 104 | (2.74 ± 0.10) × 10−3 | 213.0 ± 12.6 |
| N96 | (1.28 ± 0.03) × 104 | (1.62 ± 0.05) × 10−3 | 126.6 ± 5.25 |
| N108 | (4.27 ± 0.07) × 103 | (5.93 ± 0.12) × 10−4 | 138.7 ± 3.52 |
| N124 | (1.10 ± 0.03) × 104 | (1.28 ± 0.05) × 10−3 | 115.6 ± 6.07 |
| N125 | N/A | N/A | N/A |
| N128 | (4.35 ± 0.13) × 103 | (4.21 ± 0.22) × 10−4 | 96.8 ± 5.70 |
| N152 | (1.69 ± 0.04) × 104 | (1.95 ± 0.09) × 10−3 | 115.6 ± 5.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, J.; Geng, Z.; Luan, L.; Jiang, D.; Lu, J.; Zhang, H.; Chen, B.; Liu, X.; Xing, D. Novel Anti-Trop2 Nanobodies Disrupt Receptor Dimerization and Inhibit Tumor Cell Growth. Pharmaceutics 2024, 16, 1255. https://doi.org/10.3390/pharmaceutics16101255
Deng J, Geng Z, Luan L, Jiang D, Lu J, Zhang H, Chen B, Liu X, Xing D. Novel Anti-Trop2 Nanobodies Disrupt Receptor Dimerization and Inhibit Tumor Cell Growth. Pharmaceutics. 2024; 16(10):1255. https://doi.org/10.3390/pharmaceutics16101255
Chicago/Turabian StyleDeng, Junwen, Zhongmin Geng, Linli Luan, Dingwen Jiang, Jian Lu, Hanzhong Zhang, Bingguan Chen, Xinlin Liu, and Dongming Xing. 2024. "Novel Anti-Trop2 Nanobodies Disrupt Receptor Dimerization and Inhibit Tumor Cell Growth" Pharmaceutics 16, no. 10: 1255. https://doi.org/10.3390/pharmaceutics16101255