
Preparation of Nanoparticles Loaded with Membrane-Impermeable Peptide AC3-I and Its Protective Effect on Myocardial Ischemia and Reperfusion


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Reagents
2.3. Preparation of Curcumin Albumin Nanoparticles
2.4. Preparation of AC3-I@NPs
2.5. Morphology, Particle Size, and UV-Vis Spectral Analysis of AC3-I@NPs
2.6. Release of CCM in AC3-I@NPs
2.7. Stability of AC3-I@NPs in Solution
2.8. Isolation and Culture of Adult Rat Cardiac Myocytes
2.9. Hypoxia–Reoxygenation (H/R) of Cardiac Myocytes
2.10. PI/Hoechst Staining
2.11. Measurement of Lactate Dehydrogenase (LDH)
2.12. Electrical Field Stimulation in Primary Adult Rat Cardiomyocyte
2.13. Western Blot
2.14. Langendorff-Isolated Adult Rat Heart Perfusion
2.15. Experimental Groups and Treatments
2.16. Detection of Biochemical Indexes
2.17. Statistical Analysis
3. Results
3.1. Characterization of AC3-I@NPs
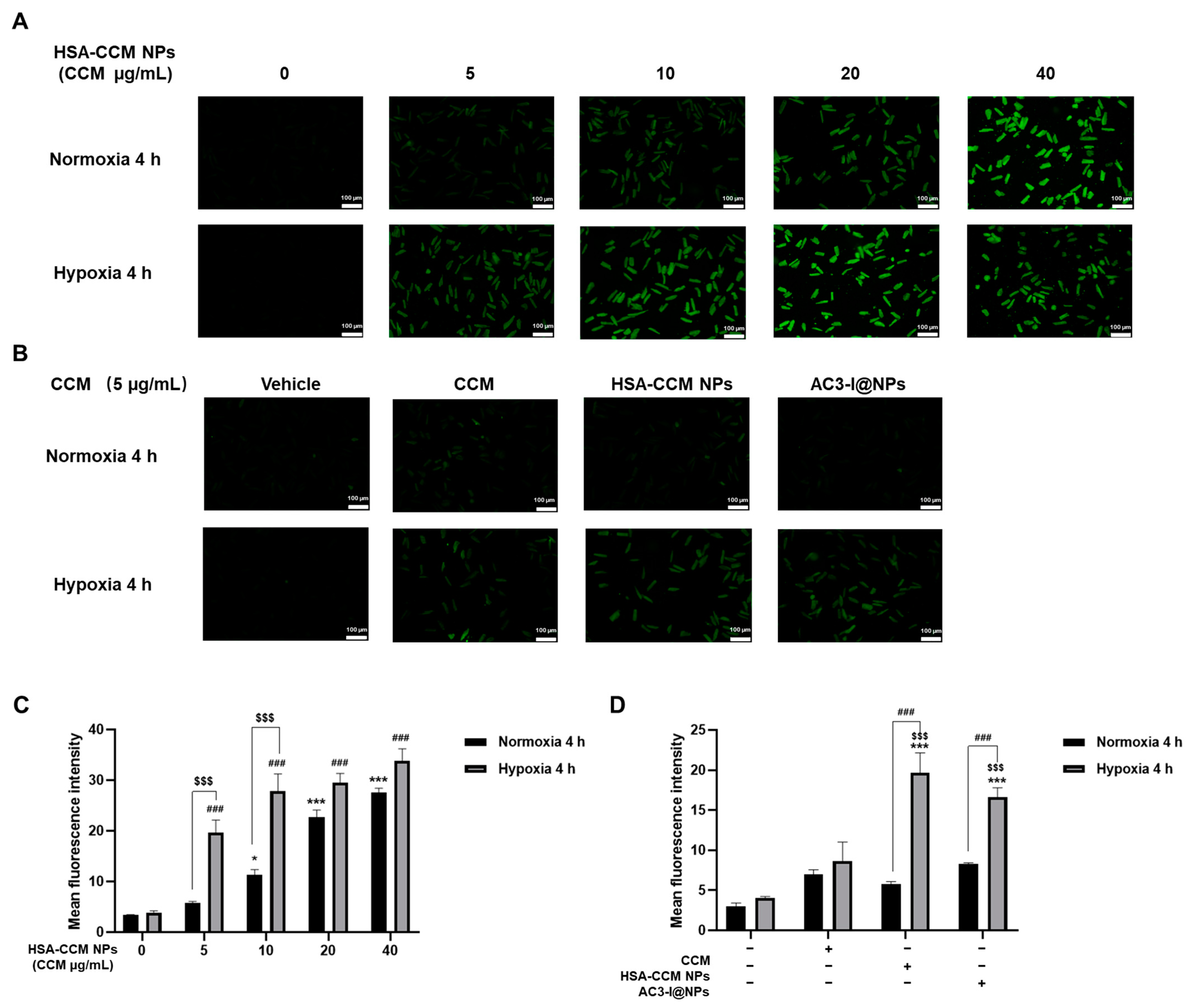
3.2. Uptake of AC3-I@NPs in Cultured Rat Cardiomyocytes
3.3. Biocompatibility of AC3-I@NPs
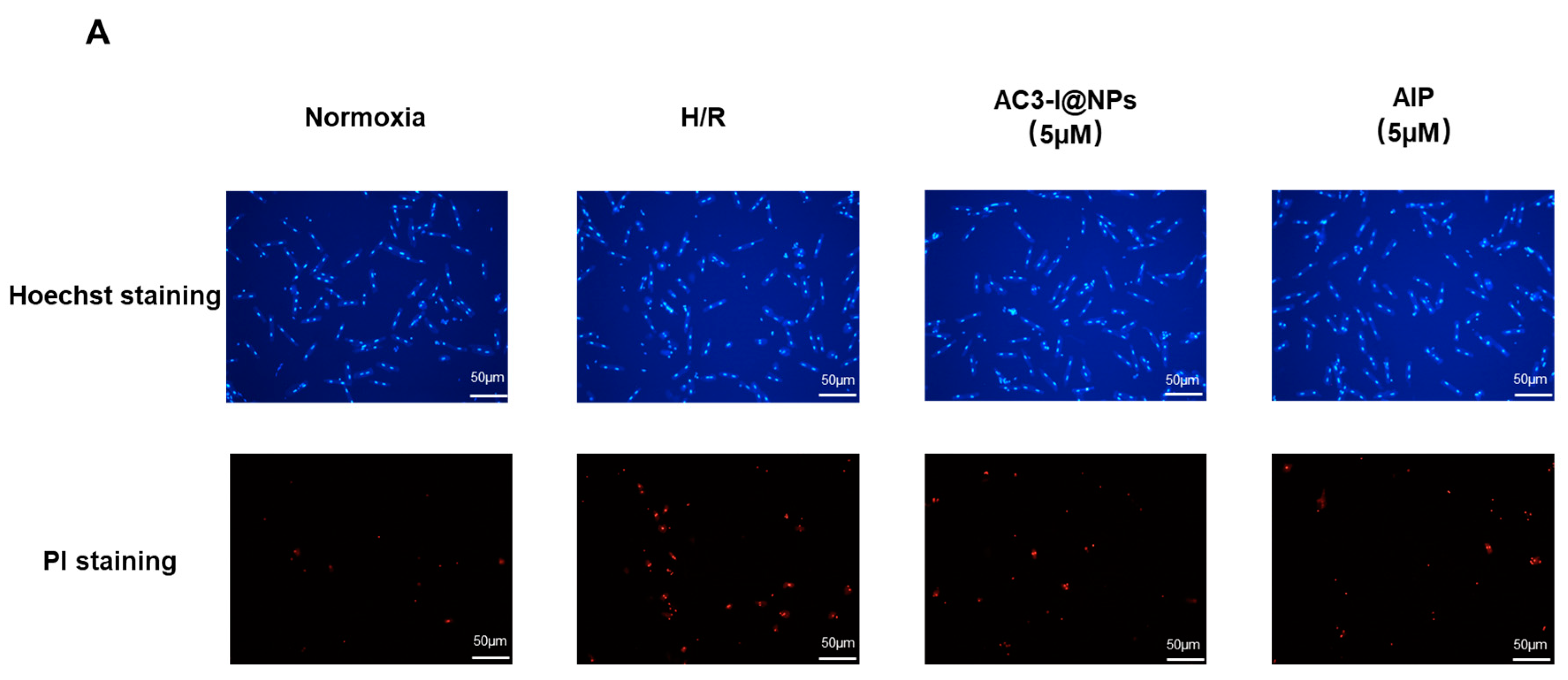
3.4. AC3-I@NPs Reduced Cell Death Caused by Hypoxia and Reoxygenation
3.5. AC3-I@NPs Alleviated the Cell Damage Caused by Hypoxia and Reoxygenation
3.6. AC3-I@NPs Reduced p-PLB Levels Evoked by Hypoxia–Reoxygenation and Electrical Field Stimulation in Adult Rat Cardiac Myocytes
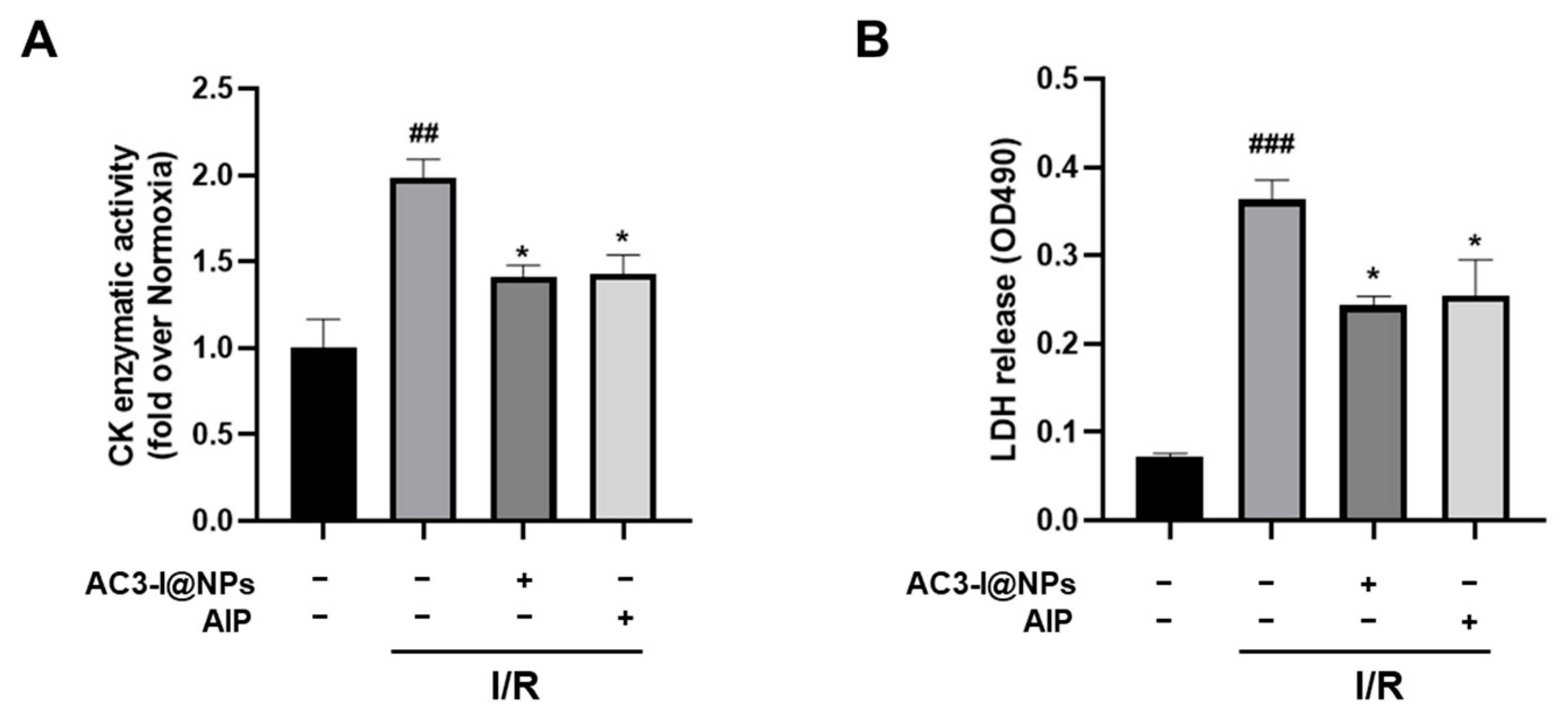
3.7. AC3-I@NPs Alleviated Levels of Lactate Dehydrogenase and Creatine Kinase in Efflux in Isolated Working Hearts after Ischemia–Reperfusion Injury
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ARCMs | Adult rat cardiomyocytes |
| CCM | Curcumin |
| CK | Creatine kinase |
| CaMKII | Ca2+/calmodulin-dependent protein kinase II |
| EFS | Electric field stimulation |
| H/R | Hypoxia–reoxygenation |
| HSA | Human serum albumin |
| I/R | Ischemia–reperfusion |
| LDH | Lactate dehydrogenase |
| MI | Myocardial ischemia |
| PBS | Phosphate-buffered saline |
| PLB | Phospholamban |
| ROS | Reactive oxygen species |
| RES | Reticular endothelial system |
| Tris | Tris(hydroxymethyl)aminomethane |
References
- Ibáñez, B.; Heusch, G.; Ovize, M.; Van de Werf, F. Evolving therapies for myocardial ischemia/reperfusion injury. J. Am. Coll. Cardiol. 2015, 65, 1454–1471. [Google Scholar] [CrossRef]
- Heusch, G. Myocardial ischaemia-reperfusion injury and cardioprotection in perspective. Nat. Rev. Cardiol. 2020, 17, 773–789. [Google Scholar] [CrossRef] [PubMed]
- Heusch, G.; Gersh, B.J. The pathophysiology of acute myocardial infarction and strategies of protection beyond reperfusion: A continual challenge. Eur. Heart J. 2017, 38, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.W.; Iyer, K.S.; Hool, L.C. The potential for nanotechnology to improve delivery of therapy to the acute ischemic heart. Nanomedicine 2016, 11, 817–832. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.T.; Huang, Z.H.; Xu, T.Z.; Sun, A.J.; Ge, J.B. New diagnostic and therapeutic strategies for myocardial infarction via nanomaterials. EBioMedicine 2022, 78, 103968. [Google Scholar] [CrossRef] [PubMed]
- Maier, L.S.; Bers, D.M. Role of Ca2+/calmodulin-dependent protein kinase (CaMK) in excitation-contraction coupling in the heart. Cardiovasc. Res. 2007, 73, 631–640. [Google Scholar] [CrossRef]
- Anderson, M.E.; Brown, J.H.; Bers, D.M. CaMKII in myocardial hypertrophy and heart failure. J. Mol. Cell Cardiol. 2011, 51, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Erickson, J.R.; Joiner, M.L.; Guan, X.; Kutschke, W.; Yang, J.; Oddis, C.V.; Bartlett, R.K.; Lowe, J.S.; O’Donnell, S.E.; Aykin-Burns, N.; et al. A dynamic pathway for calcium-independent activation of CaMKII by methionine oxidation. Cell 2008, 133, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Luczak, E.D.; Anderson, M.E. CaMKII oxidative activation and the pathogenesis of cardiac disease. J. Mol. Cell Cardiol. 2014, 73, 112–116. [Google Scholar] [CrossRef]
- Erickson, J.R.; Pereira, L.; Wang, L.; Han, G.; Ferguson, A.; Dao, K.; Copeland, R.J.; Despa, F.; Hart, G.W.; Ripplinger, C.M.; et al. Diabetic hyperglycaemia activates CaMKII and arrhythmias by O-linked glycosylation. Nature 2013, 502, 372–376. [Google Scholar] [CrossRef]
- Feng, N.; Anderson, M.E. CaMKII is a nodal signal for multiple programmed cell death pathways in heart. J. Mol. Cell Cardiol. 2017, 103, 102–109. [Google Scholar] [CrossRef]
- Zhu, W.; Woo, A.Y.; Yang, D.; Cheng, H.; Crow, M.T.; Xiao, R.P. Activation of CaMKIIdeltaC is a common intermediate of diverse death stimuli-induced heart muscle cell apoptosis. J. Biol. Chem. 2007, 282, 10833–10839. [Google Scholar] [CrossRef]
- Pellicena, P.; Schulman, H. CaMKII inhibitors: From research tools to therapeutic agents. Front. Pharmacol. 2014, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.H.; Samal, A.B.; Lee, M.; Vlach, J.; Novikov, N.; Niedziela-Majka, A.; Feng, J.Y.; Koltun, D.O.; Brendza, K.M.; Kwon, H.J.; et al. The KN-93 Molecule Inhibits Calcium/Calmodulin-Dependent Protein Kinase II (CaMKII) Activity by Binding to Ca(2+)/CaM. J. Mol. Biol. 2019, 431, 1440–1459. [Google Scholar] [CrossRef]
- Gao, Y.; Davies, S.P.; Augustin, M.; Woodward, A.; Patel, U.A.; Kovelman, R.; Harvey, K.J. A broad activity screen in support of a chemogenomic map for kinase signalling research and drug discovery. Biochem. J. 2013, 451, 313–328. [Google Scholar] [CrossRef]
- Anderson, M.E.; Braun, A.P.; Wu, Y.; Lu, T.; Wu, Y.; Schulman, H.; Sung, R.J. KN-93, an inhibitor of multifunctional Ca2+/calmodulin-dependent protein kinase, decreases early afterdepolarizations in rabbit heart. J. Pharmacol. Exp. Ther. 1998, 287, 996–1006. [Google Scholar]
- Liu, Y.; Liu, Y.; Huang, X.; Zhang, J.; Yang, L. Protective effects and mechanism of curcumin on myocardial injury induced by coronary microembolization. J. Cell. Biochem. 2019, 120, 5695–5703. [Google Scholar] [CrossRef]
- Saleh, T.; Tooba, S.; Seyed, A.S. Redox responsive curcumin-loaded human serum albumin nanoparticles: Preparation, characterization and in vitro evaluation. Int. J. Biol. Macromol. 2018, 114, 759–766. [Google Scholar] [CrossRef]
- Chelu, M.; Adina, M.M. Advanced biomedical applications of multifunctional natural and synthetic biomaterials. Processes 2023, 11, 2696. [Google Scholar] [CrossRef]
- Kim, J.; Cao, L.; Shvartsman, D.; Silva, E.A.; Mooney, D.J. Targeted delivery of nanoparticles to ischemic muscle for imaging and therapeutic angiogenesis. Nano Lett. 2011, 11, 694–700. [Google Scholar] [CrossRef]
- Ho, Y.T.; Poinard, B.; Kah, J.C. Nanoparticle drug delivery systems and their use in cardiac tissue therapy. Nanomedicine 2016, 11, 693–714. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Eckle, T. Ischemia and reperfusion—From mechanism to translation. Nat. Med. 2011, 17, 1391–1401. [Google Scholar] [CrossRef]
- Li, D.; Wang, X.; Huang, Q.; Li, S.; Zhou, Y.; Li, Z. Cardioprotection of CAPE-oNO(2) against myocardial ischemia/reperfusion induced ROS generation via regulating the SIRT1/eNOS/NF-κB pathway in vivo and in vitro. Redox Biol. 2018, 15, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [PubMed]
- Backs, J.; Backs, T.; Neef, S.; Kreusser, M.M.; Lehmann, L.H.; Patrick, D.M.; Grueter, C.E.; Qi, X.; Richardson, J.A.; Hill, J.A.; et al. The delta isoform of CaM kinase II is required for pathological cardiac hypertrophy and remodeling after pressure overload. Proc. Natl. Acad. Sci. USA 2009, 106, 2342–2347. [Google Scholar] [CrossRef] [PubMed]
- Kreusser, M.M.; Lehmann, L.H.; Wolf, N.; Keranov, S.; Jungmann, A.; Gröne, H.J.; Müller, O.J.; Katus, H.A.; Backs, J. Inducible cardiomyocyte-specific deletion of CaM kinase II protects from pressure overload-induced heart failure. Basic. Res. Cardiol. 2016, 111, 65. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From kitchen to clinic. Biochem. Pharmacol. 2008, 75, 787–809. [Google Scholar] [CrossRef]
- Elzoghby, A.O.; Samy, W.M.; Elgindy, N.A. Protein-based nanocarriers as promising drug and gene delivery systems. J. Control Release 2012, 161, 38–49. [Google Scholar] [CrossRef]
- Gong, G.; Xu, Y.; Zhou, Y.; Meng, Z.; Ren, G.; Zhao, Y.; Zhang, X.; Wu, J.; Hu, Y. Molecular switch for the assembly of lipophilic drug incorporated plasma protein nanoparticles and in vivo image. Biomacromolecules 2012, 13, 23–28. [Google Scholar] [CrossRef]
- Nguyen, M.M.; Carlini, A.S.; Chien, M.P.; Sonnenberg, S.; Luo, C.; Braden, R.L.; Osborn, K.G.; Li, Y.; Gianneschi, N.C.; Christman, K.L. Enzyme-responsive nanoparticles for targeted accumulation and prolonged retention in heart tissue after myocardial infarction. Adv. Mater. 2015, 27, 5547. [Google Scholar] [CrossRef]
- Liu, Q.; Sun, J.; Zhang, L.; Xu, Y.; Wu, B.; Cao, J. The Agonist of Inward Rectifier Potassium Channel (IK1) Attenuates Rat Reperfusion Arrhythmias Linked to CaMKII Signaling. Int. Heart J. 2021, 62, 1348–1357. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Niu, Y.; Zhang, W.; Wang, K.; Liu, T.; Zhu, W. Preparation of Nanoparticles Loaded with Membrane-Impermeable Peptide AC3-I and Its Protective Effect on Myocardial Ischemia and Reperfusion. Pharmaceutics 2024, 16, 416. https://doi.org/10.3390/pharmaceutics16030416
Liu Y, Niu Y, Zhang W, Wang K, Liu T, Zhu W. Preparation of Nanoparticles Loaded with Membrane-Impermeable Peptide AC3-I and Its Protective Effect on Myocardial Ischemia and Reperfusion. Pharmaceutics. 2024; 16(3):416. https://doi.org/10.3390/pharmaceutics16030416
Chicago/Turabian StyleLiu, Yi, Yingyi Niu, Wenjie Zhang, Kaikai Wang, Tianqing Liu, and Weizhong Zhu. 2024. "Preparation of Nanoparticles Loaded with Membrane-Impermeable Peptide AC3-I and Its Protective Effect on Myocardial Ischemia and Reperfusion" Pharmaceutics 16, no. 3: 416. https://doi.org/10.3390/pharmaceutics16030416