
Antiviral Effect of Antimicrobial Peptoid TM9 and Murine Model of Respiratory Coronavirus Infection

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Synthesis of TM9 Peptoid
2.3. Test of Efficacy and Toxicity of TM9 Peptoid
2.4. Virus
2.5. Plaque Assay
2.6. Generation of the Animal Model for Respiratory Infection
2.7. Statistical Analysis and Graphics
3. Results
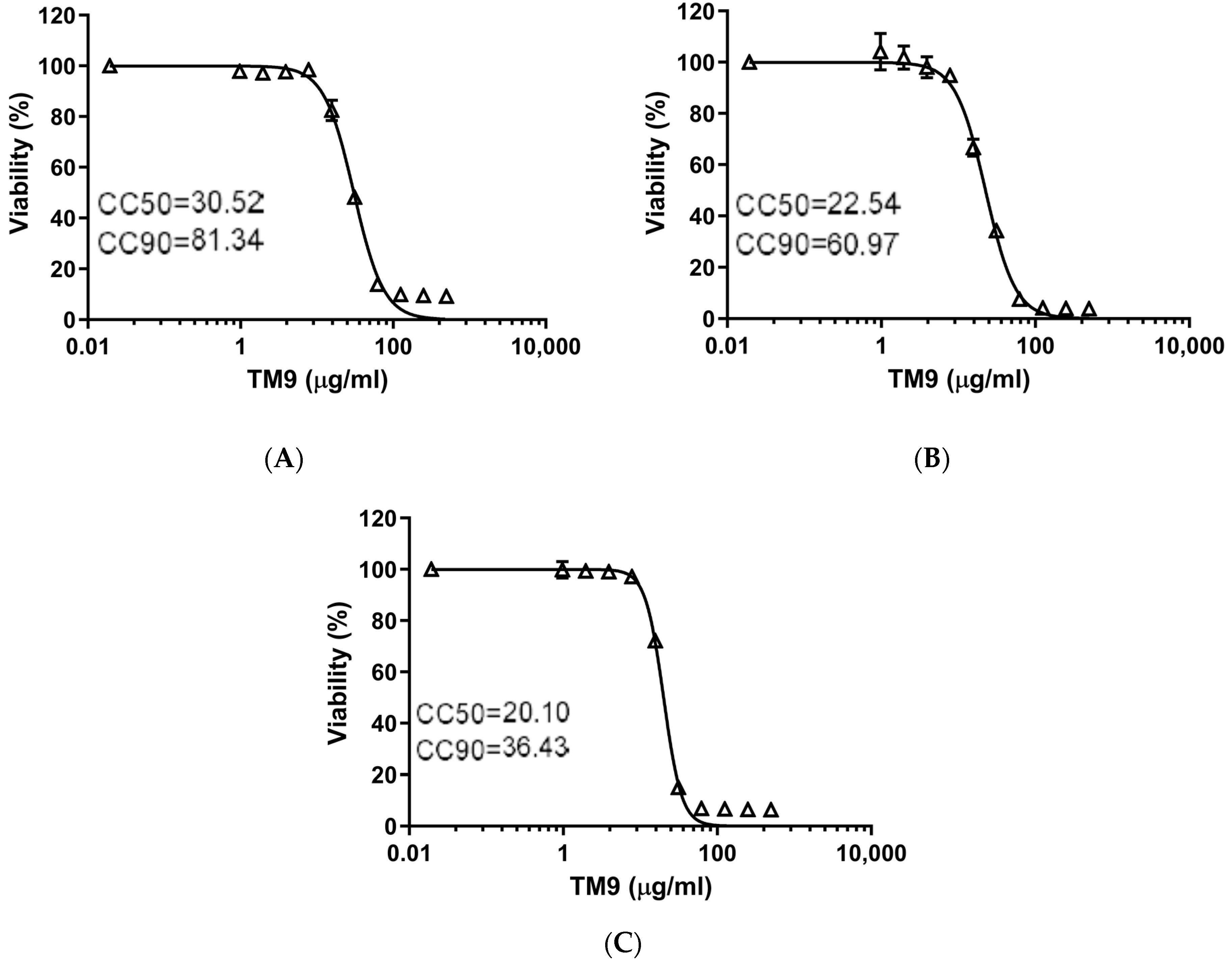
3.1. The Effect of TM9 Peptoid Was Similar on Different Cell Lines but Dependent on the Duration of Exposure
3.2. The Effective Antiviral Concentration of TM9 Was within a Therapeutic Window in L2 Cell Culture
3.3. Respiratory MHV Infection Was Effectively Generated in All Experimental Animals by Aerosol Inoculation Using Madison Chamber
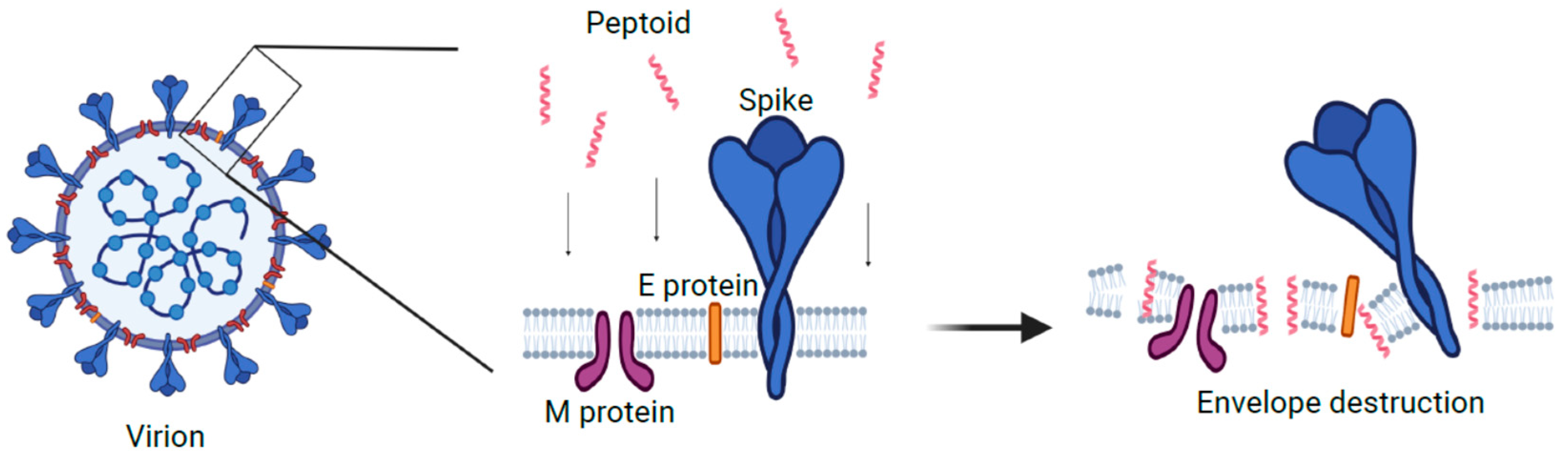
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atzrodt, C.L.; Maknojia, I.; McCarthy, R.D.P.; Oldfield, T.M.; Po, J.; Ta, K.T.L.; Stepp, H.E.; Clements, T.P. A Guide to COVID-19: A global pandemic caused by the novel coronavirus SARS-CoV-2. FEBS J. 2020, 287, 3633–3650. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; He, Y.; Zhou, Y.; Liu, S.; Zheng, B.J.; Jiang, S. The spike protein of SARS-CoV—A target for vaccine and therapeutic development. Nat. Rev. Microbiol. 2009, 7, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Furuta, Y.; Komeno, T.; Nakamura, T. Favipiravir (T-705), a broad spectrum inhibitor of viral RNA polymerase. Proc. Jpn. Acad. Ser. B 2017, 93, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Báez-Santos, Y.M.; St John, S.E.; Mesecar, A.D. The SARS-coronavirus papain-like protease: Structure, function and inhibition by designed antiviral compounds. Antiviral. Res. 2015, 115, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, S.; Shin, D.H.; Kim, M.S. Inhibition of SARS-CoV 3CL protease by flavonoids. J. Enzyme. Inhib. Med. Chem. 2020, 35, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Felgenhauer, U.; Schoen, A.; Gad, H.H.; Hartmann, R.; Schaubmar, A.R.; Failing, K.; Drosten, C.; Weber, F. Inhibition of SARS-CoV-2 by type I and type III interferons. J. Biol. Chem. 2020, 295, 13958–13964. [Google Scholar] [CrossRef] [PubMed]
- Ju, B.; Zhang, Q.; Ge, J.; Wang, R.; Sun, J.; Ge, X.; Yu, J.; Shan, S.; Zhou, B.; Song, S.; et al. Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 2020, 584, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, W.; Drabek, D.; Okba, N.M.A.; van Haperen, R.; Osterhaus, A.D.M.E.; van Kuppeveld, F.J.M.; Haagmans, B.L.; Grosveld, F.; Bosch, B.-J. A human monoclonal antibody blocking SARS-CoV-2 infection. Nat. Commun. 2020, 11, 2251. [Google Scholar] [CrossRef]
- Abbott, T.R.; Dhamdhere, G.; Liu, Y.; Lin, X.; Goudy, L.; Zeng, L.; Chemparathy, A.; Chmura, S.; Heaton, N.S.; Debs, R.; et al. Development of CRISPR as an Antiviral Strategy to Combat SARS-CoV-2 and Influenza. Cell 2020, 181, 865–876.e12. [Google Scholar] [CrossRef]
- Adamson, C.S.; Chibale, K.; Goss, R.J.M.; Jaspars, M.; Newman, D.J.; Dorrington, R.A. Antiviral drug discovery: Preparing for the next pandemic. Chem. Soc. Rev. 2021, 50, 3647. [Google Scholar] [CrossRef] [PubMed]
- Sztukowska, M.N.; Roky, M.; Demuth, D.R. Peptide and non-peptide mimetics as potential therapeutics targeting oral bacteria and oral biofilms. Mol. Oral Microbiol. 2019, 34, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Pinto, I.B.; Machado, L.S.; Meneguetti, B.T.; Nogueira, M.L.; Carvalho, C.M.E.; Roel, A.R.; Franco, O.L. Utilization of antimicrobial peptides, analogs and mimics in creating antimicrobial surfaces and bio-materials. Biochem. Eng. J. 2019, 150, 107237. [Google Scholar] [CrossRef]
- Panjla, A.; Kaul, G.; Chopra, S.; Titz, A.; Verma, S. Short Peptides and Their Mimetics as Potent Antibacterial Agents and Antibiotic Adjuvants. ACS Chem. Biol. 2021, 16, 2731–2745. [Google Scholar] [CrossRef] [PubMed]
- Baltz, R.H.; Miao, V.; Wrigley, S.K. Natural products to drugs: Daptomycin and related lipopeptide antibiotics. Nat. Prod. Rep. 2005, 22, 717–741. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.D.; Palmer, M. The action mechanism of daptomycin. Bioorg. Med. Chem. 2016, 24, 6253–6268. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.G.; Dullaghan, E.; Mookherjee, N.; Glavas, N.; Waldbrook, M.; Thompson, A.; Wang, A.; Lee, K.; Doria, S.; Hamill, P.; et al. An anti-infective peptide that selectively modulates the innateimmune response. Nat. Biotechnol. 2007, 25, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Chongsiriwatana, N.P.; Patch, J.A.; Czyzewski, A.M.; Dohm, M.T.; Ivankin, A.; Gidalevitz, D.; Zuckermann, R.N.; Barron, A.E. Peptoids that mimic the structure, function, and mechanism of helical antimicrobial peptides. Proc. Natl. Acad. Sci. USA 2008, 105, 2794–2799. [Google Scholar] [CrossRef]
- Bals, R.; Weiner, D.J.; Moscioni, A.D.; Meegalla, R.L.; Wilson, J.M. Augmentation of Innate Host Defense by Expression of a Cathelicidin Antimicrobial Peptide. Infect. Immun. 1999, 67, 6084–6089. [Google Scholar] [CrossRef]
- Gudmundsson, G.H.; Agerberth, B.; Odeberg, J.; Bergman, T.; Olsson, B.; Salcedo, R. The Human Gene FALL39 and Processing of the Cathelin Precursor to the Antibacterial Peptide LL-37 in Granulocytes. JBIC J. Biol. Inorg. Chem. 1996, 238, 325–332. [Google Scholar] [CrossRef]
- Johansson, J.; Gudmundsson, G.H.; Rottenberg, M.E.; Berndt, K.D.; Agerberth, B. Conformation-dependent Antibacterial Activity of the Naturally Occurring Human Peptide LL-37. J. Biol. Chem. 1998, 273, 3718–3724. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Weiss, T.M.; Lehrer, R.I.; Huang, H.W. Crystallization of Antimicrobial Pores in Membranes: Magainin and Protegrin. Biophys. J. 2000, 79, 2002–2009. [Google Scholar] [CrossRef] [PubMed]
- Beyond Structural Models for the Mode of Action: How Natural Antimicrobial Peptides Affect Lipid Transport|Elsevier Enhanced Reader [Internet]. Available online: https://reader.elsevier.com/reader/sd/pii/S0021979720311413?token=88D9A9CD0BF4811511B3484D660C84A4F3DDB01A89765BB4AD1CFE9DF7E97A2C14D99A233184B79EF110BE250B1685A1&originRegion=us-east-1&originCreation=20221230222938 (accessed on 30 December 2022).
- Nyembe, P.L.; Ntombela, T.; Makatini, M.M. Review: Structure-Activity Relationship of Antimicrobial Peptoids. Pharmaceutics 2023, 15, 1506. [Google Scholar] [CrossRef] [PubMed]
- Chongsiriwatana, N.P.; Lin, J.S.; Kapoor, R.; Wetzler, M.; Rea, J.A.C.; Didwania, M.K.; Contag, C.H.; Barron, A.E. Intracellular biomass flocculation as a key mechanism of rapid bacterial killing by cationic, amphipathic antimicrobial peptides and peptoids. Sci. Rep. 2017, 7, 16718. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.M.; Simon, R.J.; Zuckermann, R.N.; Kerr, J.M.; Moos, W.H. Comparison of the proteolytic susceptibilities of homologous L-amino acid, D-amino acid, and N-substituted glycine peptide and peptoid oligomers. Drug Dev. Res. 1995, 35, 20–32. [Google Scholar] [CrossRef]
- Benjamin, A.B.; Moule, M.G.; Didwania, M.K.; Hardy, J.; Saenkham-Huntsinger, P.; Sule, P.; Nielsen, J.E.; Lin, J.S.; Contag, C.H.; Barron, A.E.; et al. Efficacy of cathelicidin-mimetic antimicrobial peptoids against Staphylococcus aureus. Microbiol. Spectr. 2022, 10, e00534-22. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.E.; Alford, M.A.; Yung, D.B.Y.; Molchanova, N.; Fortkort, J.A.; Lin, J.S.; Diamond, G.; Hancock, R.E.W.; Jenssen, H.; Pletzer, D.; et al. Self-Assembly of Antimicrobial Peptoids Impacts Their Biological Effects on ESKAPE Bacterial Pathogens. ACS Infect. Dis. 2022, 8, 533–545. [Google Scholar] [CrossRef]
- Efficacy of Cathelicidin-Mimetic Antimicrobial Peptoids against Staphylococcus aureus [Internet]. Available online: https://journals.asm.org/doi/epub/10.1128/spectrum.00534-22 (accessed on 29 December 2022).
- Diamond, G.; Molchanova, N.; Herlan, C.; Fortkort, J.A.; Lin, J.S.; Figgins, E.; Bopp, N.; Ryan, L.K.; Chung, D.; Adcock, R.S.; et al. Potent Antiviral Activity against HSV-1 and SARS-CoV-2 by Antimicrobial Peptoids. Pharmaceuticals 2021, 14, 304. [Google Scholar] [CrossRef] [PubMed]
- Tate, P.M.; Mastrodomenico, V.; Cunha, C.; McClure, J.; Barron, A.E.; Diamond, G.; Mounce, B.C.; Kirshenbaum, K. Peptidomimetic Oligomers Targeting Membrane Phosphatidylserine Exhibit Broad Antiviral Activity. ACS Infect. Dis. 2023, 9, 1508–1522. [Google Scholar] [CrossRef]
- Current ICTV Taxonomy Release|ICTV [Internet]. Available online: https://ictv.global/taxonomy (accessed on 18 January 2023).
- Körner, R.; Majjouti, M.; Alcazar, M.; Mahabir, E. Of Mice and Men: The Coronavirus MHV and Mouse Models as a Translational Approach to Understand SARS-CoV-2. Viruses 2020, 12, 880. [Google Scholar] [CrossRef]
- De Albuquerque, N.; Baig, E.; Ma, X.; Zhang, J.; He, W.; Rowe, A.; Habal, M.; Liu, M.; Shalev, I.; Downey, G.P.; et al. MurineHepatitis Virus Strain 1 Produces a Clinically Relevant Model of Severe Acute Respiratory Syndrome in A/J Mice. J. Virol. 2006, 80, 10382–10394. [Google Scholar] [CrossRef] [PubMed]
- Featherstone, A.B.; Brown, A.C.; Chitlapilly Dass, S. Murine Hepatitis Virus, a Biosafety Level 2 Model for SARS-CoV-2, Can Remain Viable on Meat and Meat Packaging Materials for at Least 48 Hours. Microbiol. Spectr. 2022, 10, e01862-22. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.-Z.; Bartczak, A.; Zhang, J.; Khattar, R.; Chen, L.; Liu, M.F.; Edwards, A.; Levy, G.; McGilvray, I.D. Proteasome Inhibition In Vivo Promotes Survival in a Lethal Murine Model of Severe Acute Respiratory Syndrome. J. Virol. 2010, 84, 12419–12428. [Google Scholar] [CrossRef]
- Yang, Z.; Du, J.; Chen, G.; Zhao, J.; Yang, X.; Su, L.; Cheng, G.; Tang, H. Coronavirus MHV-A59 infects the lung and causes severe pneumonia in C57BL/6 mice. Virol. Sin. 2014, 29, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Kutter, J.S.; de Meulder, D.; Bestebroer, T.M.; Lexmond, P.; Mulders, A.; Richard, M.; Fouchier, R.A.M.; Herfst, S. SARS-CoV and SARS-CoV-2 are transmitted through the air between ferrets over more than one meter distance. Nat. Commun. 2021, 12, 1653. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-C.; Tu, H.-C.; Kuo, H.-Y.; Shao, P.-L.; Gong, Y.-N.; Weng, H.-Y.; Shu, H.-Y.; Kuo, C.-H.; Kuo, P.-H.; Chien, C.-H.; et al. Outbreak investigation in a COVID-19 designated hospital: The combination of phylogenetic analysis and field epidemiology study suggesting airborne transmission. J. Microbiol. Immunol. Infect. 2023, 56, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Rothfels, K.H.; Axelrad, A.A.; Siminovich, L.; Mclulloch, E.A.; Parker, R.C. The origin of altered cell lines from mouse, monkey and man as indicated by chromosome and transplantation studies. Can. Cancer Conf. 1959, 3, 189–214. [Google Scholar]
- Connolly, M.D.; Xuan, S.; Molchanova, N.; Zuckermann, R.N. Chapter 8—Submonomer synthesis of sequence defined peptoids with diverse side-chains. In Methods in Enzymology, 1st ed.; Pyle, A.M., Christianson, D.W., Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: Cambridge, MA, USA, 2021; Volume 656, pp. 241–270. [Google Scholar]
- Das Sarma, J.; Scheen, E.; Seo, S.H.; Koval, M.; Weiss, S.R. Enhanced green fluorescent protein expression may be used to monitor murine coronavirus spread in vitro and in the mouse central nervous system. J. Neurovirol. 2002, 8, 381–391. [Google Scholar] [CrossRef]
- Xhindoli, D.; Pacor, S.; Benincasa, M.; Scocchi, M.; Gennaro, R.; Tossi, A. The human cathelicidin LL-37—A pore-forming antibacterial peptide and host-cell modulator. Biochim. Biophys. Acta BBA Biomembr. 2016, 1858, 546–566. [Google Scholar] [CrossRef]
- Wang, K.-r.; Zhang, B.-z.; Zhang, W.; Yan, J.-x.; Li, J.; Wang, R. Antitumor effects, cell selectivity and structure–activity relationship of a novel antimicrobial peptide polybia-MPI. Peptides 2008, 29, 963–968. [Google Scholar] [CrossRef]
- Öhlinger, K.; Kolesnik, T.; Meindl, C.; Gallé, B.; Absenger-Novak, M.; Kolb-Lenz, D.; Frhlich, E. Air-liquid interface culture changes surface properties of A549 cells. Toxicol. In Vitro 2019, 60, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Falciani, C.; Zevolini, F.; Brunetti, J.; Riolo, G.; Gracia, R.; Marradi, M.; Loinaz, I.; Ziemann, C.; Cossío, U.; Llop, J.; et al. Antimicrobial Peptide-Loaded Nanoparticles as Inhalation Therapy for Pseudomonas aeruginosa Infections. Int. J. Nanomed. 2020, 15, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chang, R.Y.K.; Britton, W.J.; Chan, H.-K. Advances in the development of antimicrobial peptides and proteins for inhaled therapy. Adv. Drug Deliv. Rev. 2022, 180, 114066. [Google Scholar] [CrossRef] [PubMed]
- Bacalum, M.; Radu, M. Cationic Antimicrobial Peptides Cytotoxicity on Mammalian Cells: An Analysis Using Therapeutic Index Integrative Concept. Int. J. Pept. Res. Ther. 2015, 21, 47–55. [Google Scholar] [CrossRef]
- Savarin, C.; Bergmann, C.C. Viral-induced suppression of self-reactive T cells: Lessons from neurotropic coronavirus-induced demyelination. J. Neuroimmunol. 2017, 308, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Paidas, M.J.; Mohamed, A.B.; Norenberg, M.D.; Saad, A.; Barry, A.F.; Colon, C.; Kenyon, N.S.; Jayakumar, A.R. Multi-Organ Histopathological Changes in a Mouse Hepatitis Virus Model of COVID-19. Viruses 2021, 13, 1703. [Google Scholar] [CrossRef]
- Gong, H.H.; Worley, M.J.; Carver, K.A.; Goldstein, D.R.; Deng, J.C. Neutrophils drive pulmonary vascular leakage in MHV-1 infection of susceptible A/J mice. Front. Immunol. 2023, 13, 1089064. Available online: https://www.frontiersin.org/articles/10.3389/fimmu.2022.1089064 (accessed on 26 July 2023). [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebedev, M.; Benjamin, A.B.; Kumar, S.; Molchanova, N.; Lin, J.S.; Koster, K.J.; Leibowitz, J.L.; Barron, A.E.; Cirillo, J.D. Antiviral Effect of Antimicrobial Peptoid TM9 and Murine Model of Respiratory Coronavirus Infection. Pharmaceutics 2024, 16, 464. https://doi.org/10.3390/pharmaceutics16040464
Lebedev M, Benjamin AB, Kumar S, Molchanova N, Lin JS, Koster KJ, Leibowitz JL, Barron AE, Cirillo JD. Antiviral Effect of Antimicrobial Peptoid TM9 and Murine Model of Respiratory Coronavirus Infection. Pharmaceutics. 2024; 16(4):464. https://doi.org/10.3390/pharmaceutics16040464
Chicago/Turabian StyleLebedev, Maxim, Aaron B. Benjamin, Sathish Kumar, Natalia Molchanova, Jennifer S. Lin, Kent J. Koster, Julian L. Leibowitz, Annelise E. Barron, and Jeffrey D. Cirillo. 2024. "Antiviral Effect of Antimicrobial Peptoid TM9 and Murine Model of Respiratory Coronavirus Infection" Pharmaceutics 16, no. 4: 464. https://doi.org/10.3390/pharmaceutics16040464