
An Integrative Study of Scrophularia takesimensis Nakai in an Ovalbumin-Induced Murine Model of Asthma: The Effect on T Helper 2 Cell Activation


 , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Morphological Observations
2.3. Genome Sequencing and Assembly
2.4. Genome Annotation and Comparative Analysis
2.5. Phylogenetic Analysis
2.6. Preparation of S. takesimensis Extract (STE)
2.7. Animals
2.8. Experimental Procedures
2.9. Histology
2.10. Measurement of T Helper 2 (Th2) Cytokine and Immunoglobulin E (IgE) Levels in Bronchoalveolar Lavage Fluid (BALF)
2.11. Flow Cytometry
2.12. Statistical Analysis
3. Results
3.1. Description and Morphological Characteristics of Scrophularia Takesimensis
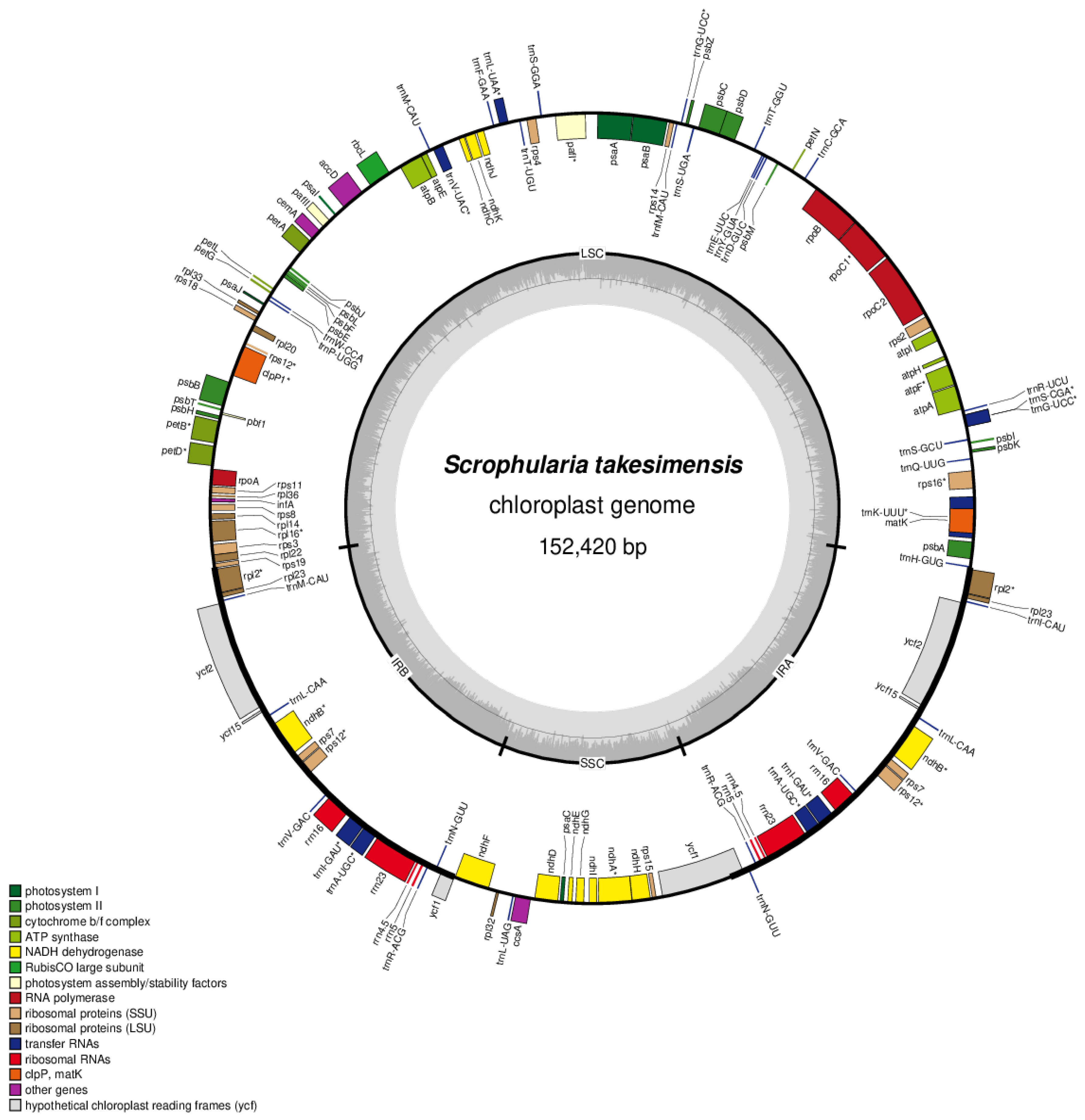
3.2. Chloroplast Genome Organization and Repeat Sequence of S. takesimensis
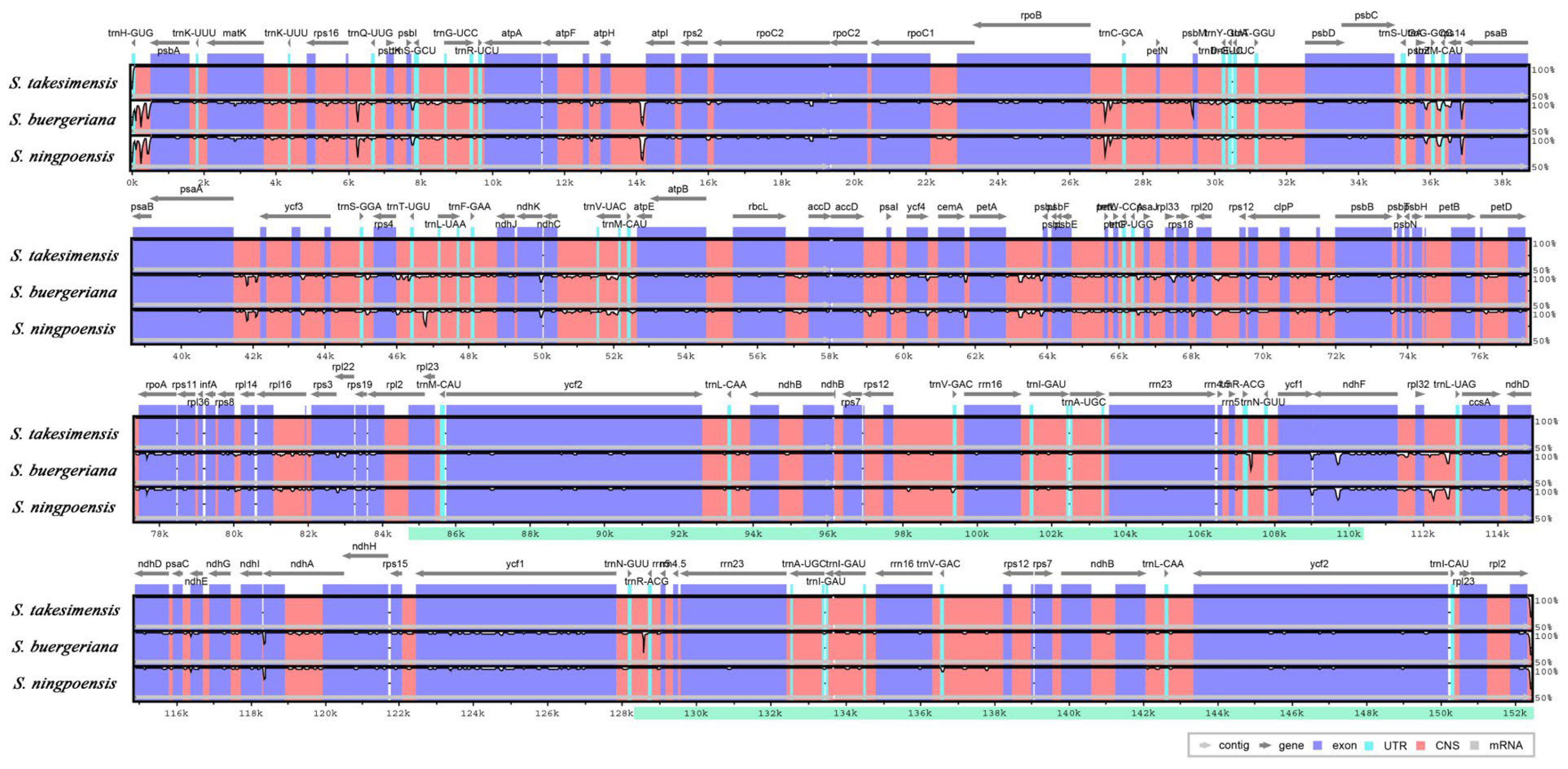
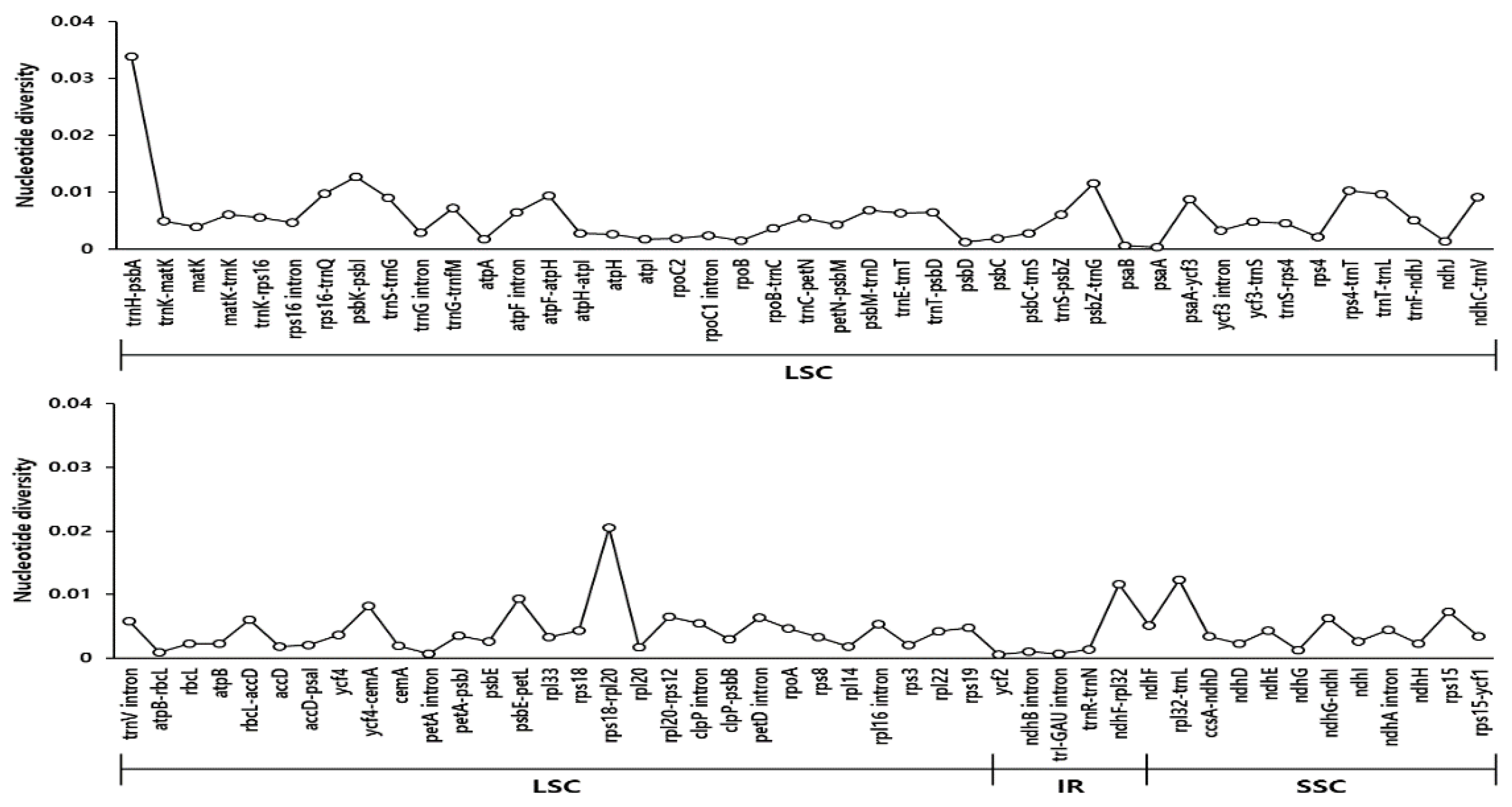
3.3. Comprehensive Comparative Analysis of Scrophularia Chloroplast Genomes
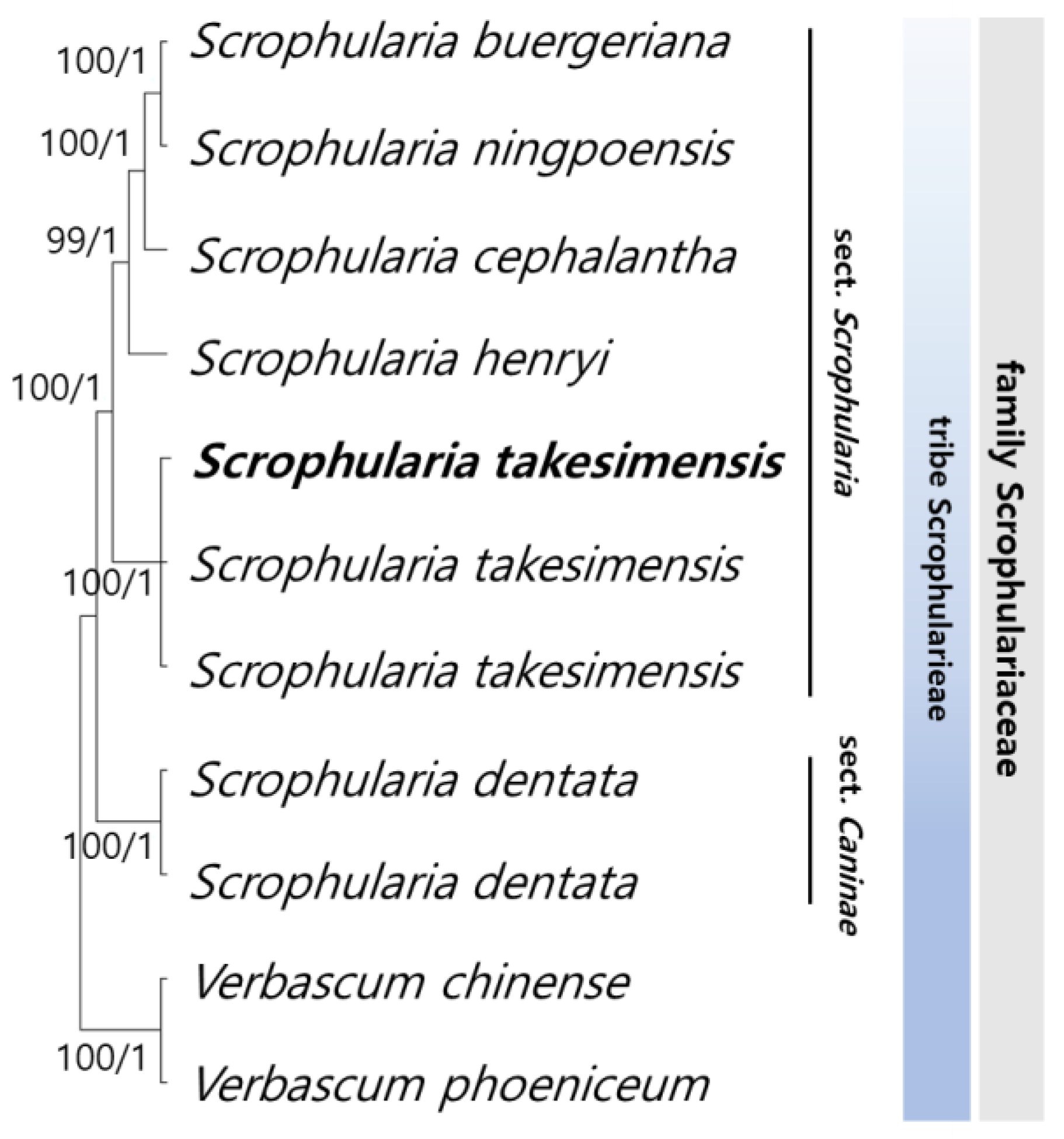
3.4. Phylogenetic Relationships among Scrophularia Genus
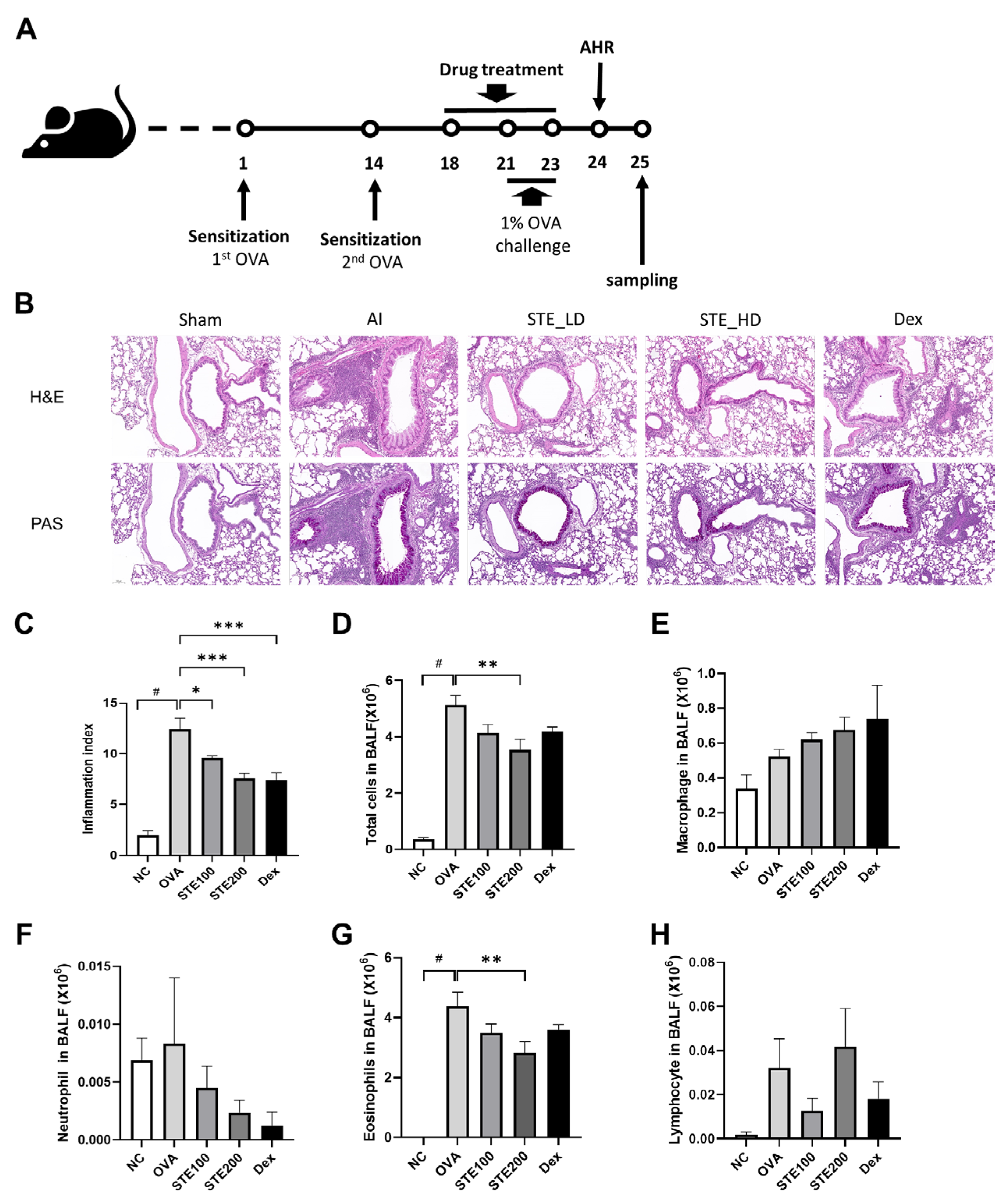
3.5. Effects of S. takesimensis Extract (STE) on Lung Histopathological Change and Number of Inflammatory Cells in the Bronchoalveolar Lavage Fluid (BALF) of an Ovalbumin (OVA)-Induced Asthmatic Animal Model
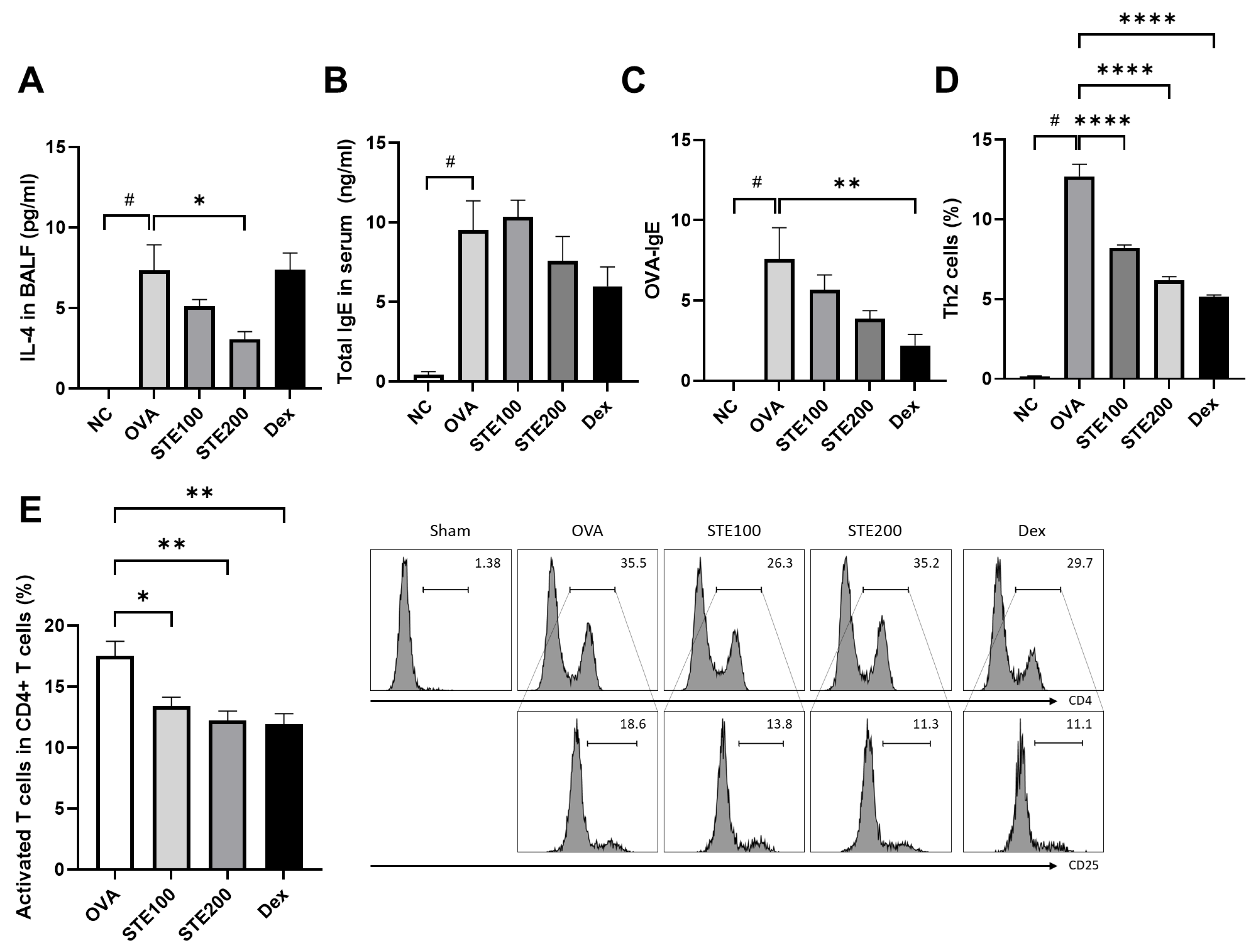
3.6. Effects of S. takesimensis Extract (STE) on Interleukin-4 (IL-4) and Immunoglobulin E (IgE) Levels in Bronchoalveolar Lavage Fluid (BALF) of Asthmatic Animal Model
3.7. Effects of S. takesimensis Extract (STE) on CD4+ T Cell (Helper T Cell) Activation in the Lung Tissues of Asthmatic Animal Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yamazaki, T. Scrophulariaceae. In Flora of Japan; Iwatsuki, K., Yamazaki, T., Boufford, D.E., Ohba, H., Eds.; Kodansha: Tokyo, Japan, 1993; Volume IIIa, pp. 326–331. [Google Scholar]
- Hong, D.Y.; Yang, H.B.; Jin, C.L.; Fischer, M.A.; Holmgren, N.H.; Mill, R.R. Scrophulariaceae. In Flora of China; Wu, Z.Y., Raven, P.H., Eds.; Science Press: Beijing China; Missori Botanical Garden Press: St. Louis, MO, USA, 1998; Volume 18, pp. 11–20. [Google Scholar]
- Choi, H.K. Scrophularia. In The Genera of Vascular Plants of Korea; Park, C.W., Ed.; Academy Publishing Co.: Seoul, Republic of Korea, 2018; pp. 1148–1150. [Google Scholar]
- Kim, J.K.; Kim, Y.H.; Lee, H.H.; Lim, S.S.; Park, K.W. Effect of Scrophularia buergeriana extract on the degranulation of mast cells and ear swelling induced by dinitrofluorobenzene in mice. Inflammation 2012, 35, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Korea Food and Drug Administration. The Korean Pharmacopoeia, 12th ed.; The KFDA Notification: Seoul, Republic of Korea, 2023. [Google Scholar]
- Korea Institute of Oriental Medicine. Defining Dictionary for Medicinal Herbs. Available online: https://oasis.kiom.re.kr/herblib/main.do (accessed on 10 October 2023).
- Lee, H.J.; Kim, H.L.; Lee, D.R.; Choi, B.K.; Yang, S.H. Scrophulariae Radix: An Overview of Its Biological Activities and Nutraceutical and Pharmaceutical Applications. Molecules 2021, 26, 5250. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.D.; Oh, B.U. A taxonomic study of Korean Scrophularia L. (Scrophulariaceae) based on morphological characters. Kor. J. Plant Res. 2013, 26, 271–283. [Google Scholar] [CrossRef]
- Lim, Y.; Na, S.T.; Lee, S.J.; Cho, K.H.; Shin, H. Spatial distribution patterns and implications for conservation of Scrophularia takesimensis (Scrophulariaceae), an endangered endemic species on Ulleung Island, Korea. J. Plant Biol. 2008, 51, 213–220. [Google Scholar] [CrossRef]
- Park, J.; Kim, M.; Park, K.-R. Genetic variation in endangered Scrophularia takesimensis (Scrophulariaceae) from Ulleung Island. Bot. Stud. 2010, 51, 371–376. [Google Scholar]
- Ma, S.; Lim, Y.S.; Na, S.T.; Lee, J.; Shin, H.C. Genetic structure and population differentiation of endangered Scrophularia takesimensis (Scrophulariaceae) in Ulleung Island, Korea. Korean J. Plant Taxon. 2011, 41, 182–3193. [Google Scholar] [CrossRef]
- Choi, H.J.; Jang, H.; Isagi, Y.; Oh, B.U. Distribution and conservation status of the critically endangered Scrophularia takesimensis, a plant endemic to Ulleung Island, Republic of Korea. Oryx 2012, 46, 399–402. [Google Scholar] [CrossRef][Green Version]
- Gil, H.Y.; Maki, M.; Pimenova, E.A.; Taran, A.; Kim, S.C. Origin of the critically endangered endemic species Scrophularia takesimensis (Scrophulariaceae) on Ulleung Island, Korea: Implications for conservation. J. Plant Res. 2020, 133, 765–782. [Google Scholar] [CrossRef]
- Jansen, R.K.; Cai, Z.; Raubeson, L.A.; Daniell, H.C.; Depamphilis, W.; Leebens-Mack, J.; Muller, K.F.; Guisinger-Bellian, M.; Haberle, R.C.; Hansen, A.K.; et al. Analysis of 81 genes from 64 plastid genomes resolves relationships in angiosperms and identifies genome-scale evolutionary patterns. Proc. Natl. Acad. Sci. USA 2007, 104, 19369–19374. [Google Scholar] [CrossRef]
- Parks, M.; Cronn, R.; Liston, A. Increasing phylogenetic resolution at low taxonomic levels using massively parallel sequencing of chloroplast genomes. BMC Biol. 2009, 7, 84. [Google Scholar] [CrossRef]
- Jang, H.; Nam, G.; Park, M. Characterization of the complete chloroplast genome of Scrophularia cephalantha endemic to Korea. Mitochondrial DNA B Resour. 2021, 6, 3179–3180. [Google Scholar] [CrossRef]
- Ni, L.; Zhao, Z.; Dorge, G.; Ma, M. The complete chloroplast genome of Ye-Xing-Ba (Scrophularia dentata; Scrophulariaceae), an Alpine Tibetan Herb. PLoS ONE. 2016, 11, e0158488. [Google Scholar] [CrossRef]
- Guo, L.; Wang, X.; Wang, R.; Li, P. Characterization and comparative analysis of chloroplast genomes of medicinal herb Scrophularia ningpoensis and Its common adulterants (Scrophulariaceae). Int. J. Mol. Sci. 2023, 24, 10034. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E.J.; Lee, K.Y.; Kim, S.H.; Sung, S.H.; Kim, Y.C. Cognitive-enhancing and antioxidant activities of iridoid glycosides from Scrophularia buergeriana in scopolamine-treated mice. Eur. J. Pharmacol. 2008, 588, 78–84. [Google Scholar] [CrossRef]
- Lee, E.J.; Kim, S.R.; Kim, J.; Kim, Y.C. Hepatoprotective phenylpropanoids from Scrophularia buergeriana roots against CCl(4)-induced toxicity: Action mechanism and structure-activity relationship. Planta Med. 2002, 68, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Sung, S.H.; Jang, Y.P.; Markelonis, G.J.; Oh, T.H.; Kim, Y.C. E-p-methoxycinnamic acid protects cultured neuronal cells against neurotoxicity induced by glutamate. Br. J. Pharmacol. 2002, 135, 1281–1291. [Google Scholar] [CrossRef]
- Kim, S.J.; Park, J.S.; Myung, N.Y.; Moon, P.D.; Choi, I.Y.; An, H.J.; Kim, N.H.; Na, H.J.; Kim, D.H.; Kim, M.C.; et al. Scrophularia buergeriana regulates cytokine production in vitro. Immunopharmacol. Immunotoxicol. 2009, 31, 246–252. [Google Scholar] [CrossRef]
- Jung, T.Y.; Lee, A.Y.; Song, J.H.; Lee, M.Y.; Lim, J.O.; Lee, S.J.; Ko, J.W.; Shin, N.R.; Kim, J.C.; Shin, I.S.; et al. Scrophularia koraiensis Nakai Attenuates Allergic Airway Inflammation via Suppression of NF-κB and Enhancement of Nrf2/HO-1 Signaling. Antioxidants 2020, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Lee, A.Y.; Song, J.H.; Yang, S.; Park, I.; Lim, J.O.; Jung, T.Y.; Ko, J.W.; Kim, J.C.; Lim, K.S.; et al. Scrophularia buergeriana attenuates allergic inflammation by reducing NF-κB activation. Phytomedicine 2020, 67, 153159. [Google Scholar] [CrossRef]
- Allen, G.; Flores-Vergara, M.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat. Protoc. 2006, 1, 2320. [Google Scholar]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. Gigascience 2012, 1, 18. [Google Scholar] [CrossRef]
- Delcher, A.L.; Salzberg, S.L.; Phillippy, A.M. Using MUMmer to identify similar regions in large sequence sets. Curr. Protoc. Bioinform. 2003, 10, 10.3. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [PubMed]
- Carver, T.; Berriman, M.; Tivey, A.; Patel, C.; Bohme, U.; Barrell, B.G.; Parkhill, J.; Rajandream, M.A. Artemis and ACT: Viewing, annotating and comparing sequences stored in a relational database. Bioinformatics 2008, 24, 2672–2676. [Google Scholar] [CrossRef]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Michael, D.; Gurusaran, M.; Santhosh, R.; Hussain, M.K.; Satheesh, S.N.; Suhan, S.; Sivaranjan, P.; Jaiswal, A.; Sekar, K. RepEx: A web server to extract sequence repeats from protein and DNA sequences. Comput. Biol. Chem. 2019, 78, 424–430. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Beier, S.; Thiel, T.; Munch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v 1.4.2 Molecular Evolution, Phylogenetics and Epidemiology. Univ. Edinburgh, Inst, Evol. Biol. Edinburgh. 2014. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 1 January 2023).
- Shin, I.S.; Park, J.W.; Shin, N.R.; Jeon, C.M.; Kwon, O.K.; Kim, J.S.; Kim, J.C.; Oh, S.R.; Ahn, K.S. Melatonin reduces airway inflammation in ovalbumin-induced asthma. Immunobiology 2014, 219, 901–908. [Google Scholar] [CrossRef]
- Kim, J.S.; Son, Y.; Bae, M.J.; Lee, S.S.; Park, S.H.; Lee, H.J.; Lee, S.I.; Lee, C.G.; Kim, S.D.; Jo, W.S.; et al. Continuous Exposure to Low-Dose-Rate Gamma Irradiation Reduces Airway Inflammation in Ovalbumin-Induced Asthma. PLoS ONE 2015, 10, e0143403. [Google Scholar] [CrossRef]
- Kang, S.; Kim, H.Y.; Lee, A.Y.; Kim, H.S.; Park, J.H.; Moon, B.C.; Nam, H.H.; Chae, S.W.; Jung, B.; Moon, C.; et al. Camellia sinensis (L.) Kuntze Extract Attenuates Ovalbumin-Induced Allergic Asthma by Regulating Airway Inflammation and Mucus Hypersecretion. Pharmaceutics 2023, 15, 2355. [Google Scholar] [CrossRef]
- Nam, H.H.; Lee, J.H.; Ryu, S.M.; Lee, S.; Yang, S.; Noh, P.; Moon, B.C.; Kim, J.S.; Seo, Y.-S. Gekko gecko extract attenuates airway inflammation and mucus hypersecretion in a murine model of ovalbumin-induced asthma. J. Ethnopharmacol. 2022, 282, 114574. [Google Scholar] [CrossRef]
- Ernst, M.; Saslis-Lagoudakis, C.H.; Grace, O.M.; Nilsson, N.; Simonsen, H.T.; Horn, J.W.; Rønsted, N. Evolutionary prediction of medicinal properties in the genus Euphorbia L. Sci. Rep. 2016, 6, 30531. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Spandidos, D.A.; Tsatsakis, A.; Margina, D.; Izotov, B.N.; Yang, S.H. Neuroprotective effects of Scrophularia buergeriana extract against glutamate-induced toxicity in SH-SY5Y cells. Int. J. Mol. Med. 2019, 43, 2144–2152. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E.J.; Ma, C.J.; Lee, K.Y.; Kim, S.H.; Sung, S.H.; Kim, Y.C. KD-501, a standardized extract of Scrophularia buergeriana has both cognitive-enhancing and antioxidant activities in mice given scopolamine. J. Ethnopharmacol. 2009, 121, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Xie, W.; Xu, Q.; Liang, T.; Xu, X.; Sun, G.; Sun, X. Neuroprotective Effects of Radix Scrophulariae on Cerebral Ischemia and Reperfusion Injury via MAPK Pathways. Molecules 2018, 23, 2401. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.H.; Baek, J.M.; Erkhembaatar, M.; Kim, M.S.; Yoon, K.H.; Oh, J.; Lee, M.S. Harpagoside Inhibits RANKL-Induced Osteoclastogenesis via Syk-Btk-PLCγ2-Ca(2+) Signaling Pathway and Prevents Inflammation-Mediated Bone Loss. J. Nat. Prod. 2015, 78, 2167–2174. [Google Scholar] [CrossRef] [PubMed]
- Che, D.; Cao, J.; Liu, R.; Wang, J.; Hou, Y.; Zhang, T.; Wang, N. Harpagoside-induced anaphylactic reaction in an IgE-independent manner both in vitro and in vivo. Immunopharmacol. Immunotoxicol. 2018, 40, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Deng, Y.; Chen, N.; Zhang, X.; Huang, T. Infrared-assisted extraction followed by high performance liquid chromatography to determine angoroside C, cinnamic acid, and harpagoside content in Scrophularia ningpoensis. BMC Complement. Altern. Med. 2019, 19, 130. [Google Scholar] [CrossRef] [PubMed]
- Axmann, S.; Hummel, K.; Nöbauer, K.; Razzazi-Fazeli, E.; Zitterl-Eglseer, K. Pharmacokinetics of harpagoside in horses after intragastric administration of a Devil’s claw (Harpagophytum procumbens) extract. J. Vet. Pharmacol. Ther. 2019, 42, 37–44. [Google Scholar] [CrossRef]
- van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | S. takesimensis | S. buergeriana | S. ningpoensis |
|---|---|---|---|
| Habitat | Seashore | Forest | Forest |
| Height | ~1.5 m | ~1.8 m | ~1.5 m |
| Root | Simple and stout | Fusiform to conical | Fusiform to conical |
| Stem | Wingless, glabrous | Wingless, glabrous | Slightly winged, mostly white crisped hairy |
| Leaf shape | Ovate | Ovate | Mostly ovate |
| Leaf apex | Acute | Acute | Acute |
| Leaf margins | Serrate almost without spinose tooth | Serrate with spinose tooth | Serrate with spinose tooth |
| Leaf base | Cuneate to rounded | Cuneate to rounded | Cuneate to rounded |
| Inflorescence shape | Panicle-like cymes | Spike-like cymes | Panicle-like cymes |
| Inflorescence position | Terminal and axillary | Mostly terminal | Terminal and axillary |
| Flower color | Red | Yellowish green | Red |
| Species | S. takesimensis | S. buergeriana | S. ningpoensis |
|---|---|---|---|
| Accession number | OQ580987 | NC_031437 | NC_053823 |
| Total cp genome size (bp) | 152,420 | 153,631 | 153,175 |
| Large single copy (LSC) region (bp) | 83,526 | 84,454 | 84,255 |
| Inverted repeat (IR) region (bp) | 25,478 | 25,624 | 25,490 |
| Small single copy (SSC) region (bp) | 17,938 | 17,929 | 17,938 |
| Total number of genes (unique) | 114 | 114 | 114 |
| Protein-coding gene (unique) | 80 | 80 | 80 |
| rRNA (unique) | 4 | 4 | 4 |
| tRNA (unique) | 30 | 30 | 30 |
| GC content (%) | 38 | 38 | 38 |
| LSC (%) | 36 | 36 | 36 |
| IR (%) | 43 | 43 | 43 |
| SSC (%) | 32 | 32 | 32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, Y.-S.; Song, J.-H.; Kim, H.S.; Nam, H.H.; Yang, S.; Choi, G.; Chae, S.-W.; Lee, J.; Jung, B.; Kim, J.-S.; et al. An Integrative Study of Scrophularia takesimensis Nakai in an Ovalbumin-Induced Murine Model of Asthma: The Effect on T Helper 2 Cell Activation. Pharmaceutics 2024, 16, 529. https://doi.org/10.3390/pharmaceutics16040529
Seo Y-S, Song J-H, Kim HS, Nam HH, Yang S, Choi G, Chae S-W, Lee J, Jung B, Kim J-S, et al. An Integrative Study of Scrophularia takesimensis Nakai in an Ovalbumin-Induced Murine Model of Asthma: The Effect on T Helper 2 Cell Activation. Pharmaceutics. 2024; 16(4):529. https://doi.org/10.3390/pharmaceutics16040529
Chicago/Turabian StyleSeo, Yun-Soo, Jun-Ho Song, Hyo Seon Kim, Hyeon Hwa Nam, Sungyu Yang, Goya Choi, Sung-Wook Chae, Jeongmin Lee, Bokyung Jung, Joong-Sun Kim, and et al. 2024. "An Integrative Study of Scrophularia takesimensis Nakai in an Ovalbumin-Induced Murine Model of Asthma: The Effect on T Helper 2 Cell Activation" Pharmaceutics 16, no. 4: 529. https://doi.org/10.3390/pharmaceutics16040529
APA StyleSeo, Y.-S., Song, J.-H., Kim, H. S., Nam, H. H., Yang, S., Choi, G., Chae, S.-W., Lee, J., Jung, B., Kim, J.-S., & Park, I. (2024). An Integrative Study of Scrophularia takesimensis Nakai in an Ovalbumin-Induced Murine Model of Asthma: The Effect on T Helper 2 Cell Activation. Pharmaceutics, 16(4), 529. https://doi.org/10.3390/pharmaceutics16040529