
Increased Cellular Uptake of ApoE3- or c(RGD)-Modified Liposomes for Glioblastoma Therapy Depending on the Target Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Liposomes
2.3. Surface-Modification of Liposomes with Apolipoprotein E3
2.3.1. Thiolation of Apolipoprotein E3
2.3.2. Post-Conjugation of Apolipoprotein E3 to Liposomes
2.4. Characterization of Liposomes
2.5. In Vitro Studies
2.5.1. Cell Culture and Reagents
2.5.2. Cell Staining Assay
2.5.3. Liposomal Uptake Assay
2.5.4. Cytotoxicity Assay
2.6. Statistical Analysis
3. Results and Discussion
3.1. Characterization and Stability of Liposomes
3.2. Expression Levels of LRP1 and Integrin αV in Endothelial and Cancer Cells
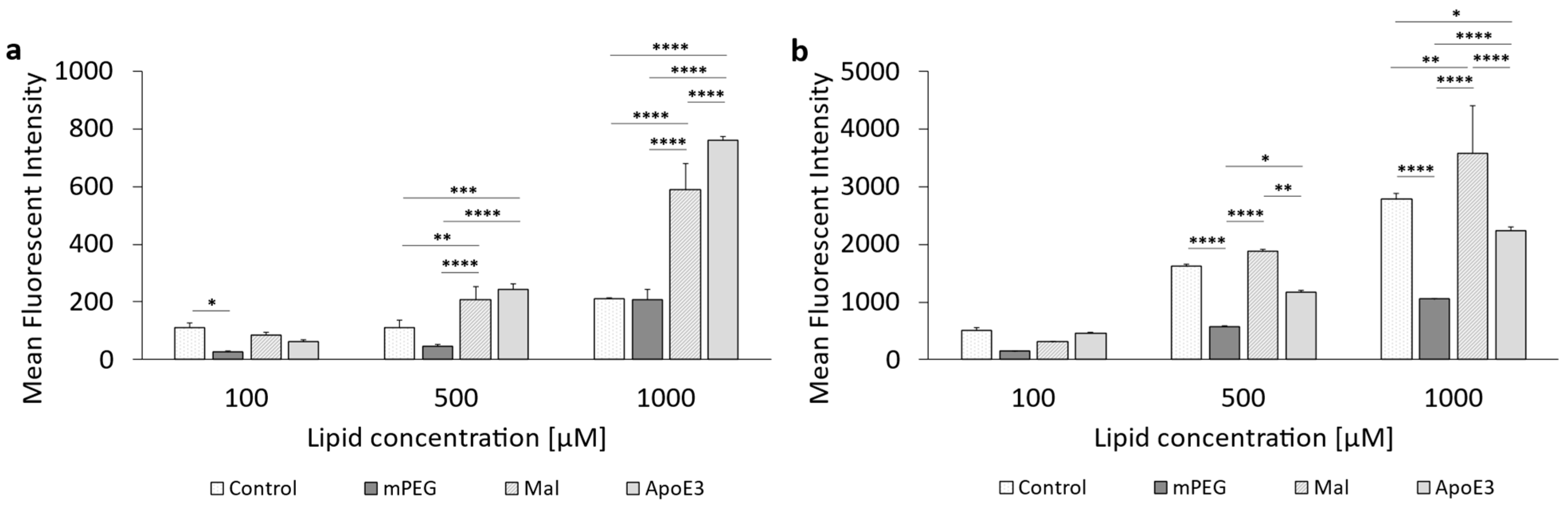
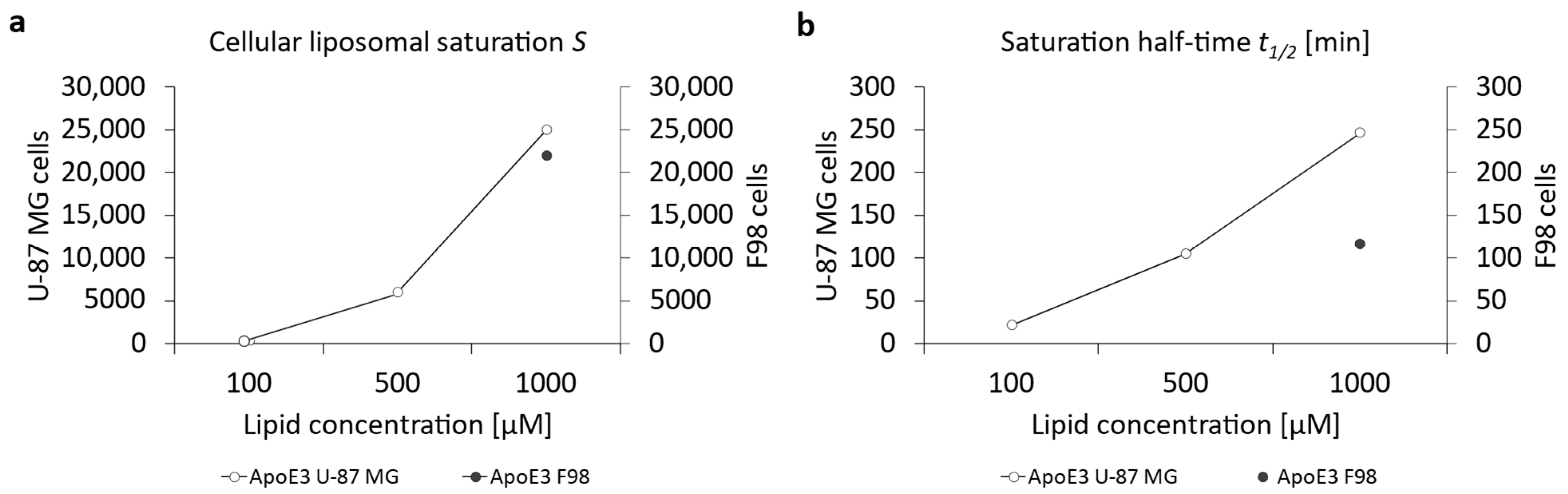
3.3. Cellular Uptake of ApoE3-Modified Liposomes
3.4. Cellular Uptake of c(RGD)-Modified Liposomes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Price, M.; Neff, C.; Cioffi, G.; Waite, K.A.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2015–2019. Neuro Oncol. 2022, 24, v1–v95. [Google Scholar] [CrossRef]
- Grech, N.; Dalli, T.; Mizzi, S.; Meilak, L.; Calleja, N.; Zrinzo, A. Rising Incidence of Glioblastoma Multiforme in a Well-Defined Population. Cureus 2020, 12, e8195. [Google Scholar] [CrossRef]
- Johnson, D.R.; O’Neill, B.P. Glioblastoma Survival in the United States before and during the Temozolomide Era. J. Neurooncol 2012, 107, 359–364. [Google Scholar] [CrossRef]
- Obermeier, B.; Daneman, R.; Ransohoff, R.M. Development, Maintenance and Disruption of the Blood-Brain Barrier. Nat. Med. 2013, 19, 1584–1596. [Google Scholar] [CrossRef]
- Bruinsmann, F.A.; Richter Vaz, G.; de Cristo Soares Alves, A.; Aguirre, T.; Raffin Pohlmann, A.; Stanisçuaski Guterres, S.; Sonvico, F. Nasal Drug Delivery of Anticancer Drugs for the Treatment of Glioblastoma: Preclinical and Clinical Trials. Molecules 2019, 24, 4312. [Google Scholar] [CrossRef]
- Chung, K.; Ullah, I.; Kim, N.; Lim, J.; Shin, J.; Lee, S.C.; Jeon, S.; Kim, S.H.; Kumar, P.; Lee, S.-K. Intranasal Delivery of Cancer-Targeting Doxorubicin-Loaded PLGA Nanoparticles Arrests Glioblastoma Growth. J. Drug Target. 2020, 28, 617–626. [Google Scholar] [CrossRef]
- Saka, R.; Sathe, P.; Khan, W. Brain Local Delivery Strategy. In Brain Targeted Drug Delivery System; Elsevier: Amsterdam, The Netherlands, 2019; pp. 241–286. [Google Scholar] [CrossRef]
- Oller-Salvia, B.; Sánchez-Navarro, M.; Giralt, E.; Teixidó, M. Blood-Brain Barrier Shuttle Peptides: An Emerging Paradigm for Brain Delivery. Chem. Soc. Rev. 2016, 45, 4690–4707. [Google Scholar] [CrossRef]
- Alam, M.I.; Beg, S.; Samad, A.; Baboota, S.; Kohli, K.; Ali, J.; Ahuja, A.; Akbar, M. Strategy for Effective Brain Drug Delivery. Eur. J. Pharm. Sci. 2010, 40, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Bernacki, J.; Dobrowolska, A.; Nierwińska, K.; Małecki, A. Physiology and Pharmacological Role of the Blood-Brain Barrier. Pharmacol. Rep. 2008, 60, 600–622. [Google Scholar] [PubMed]
- Abbott, N.J.; Patabendige, A.A.K.; Dolman, D.E.M.; Yusof, S.R.; Begley, D.J. Structure and Function of the Blood-Brain Barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Molecular Biology of the Blood-Brain Barrier. Mol. Biotechnol. 2005, 30, 57–70. [Google Scholar] [CrossRef]
- Lei, F.; Fan, W.; Li, X.K.; Wang, S.; Hai, L.; Wu, Y. Design, Synthesis and Preliminary Bio-Evaluation of Glucose–Cholesterol Derivatives as Ligands for Brain Targeting Liposomes. Chin. Chem. Lett. 2011, 22, 831–834. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Drug and Gene Delivery to the Brain: The Vascular Route. Neuron 2002, 36, 555–558. [Google Scholar] [CrossRef]
- Abbott, N.J.; Rönnbäck, L.; Hansson, E. Astrocyte–Endothelial Interactions at the Blood–Brain Barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef]
- Andersen, O.M.; Willnow, T.E. Lipoprotein Receptors in Alzheimer’s Disease. Trends Neurosci. 2006, 29, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Deane, R.; Du Yan, S.; Submamaryan, R.K.; LaRue, B.; Jovanovic, S.; Hogg, E.; Welch, D.; Manness, L.; Lin, C.; Yu, J.; et al. RAGE Mediates Amyloid-β Peptide Transport across the Blood-Brain Barrier and Accumulation in Brain. Nat. Med. 2003, 9, 907–913. [Google Scholar] [CrossRef]
- Hauser, P.S.; Narayanaswami, V.; Ryan, R.O. Apolipoprotein E: From Lipid Transport to Neurobiology. Prog. Lipid Res. 2011, 50, 62–74. [Google Scholar] [CrossRef]
- Wagner, S.; Zensi, A.; Wien, S.L.; Tschickardt, S.E.; Maier, W.; Vogel, T.; Worek, F.; Pietrzik, C.U.; Kreuter, J.; von Briesen, H. Uptake Mechanism of ApoE-Modified Nanoparticles on Brain Capillary Endothelial Cells as a Blood-Brain Barrier Model. PLoS ONE 2012, 7, e0032568. [Google Scholar] [CrossRef]
- Bayly, G.R. Lipids and Disorders of Lipoprotein Metabolism. In Clinical Biochemistry: Metabolic and Clinical Aspects; Elsevier: Amsterdam, The Netherlands, 2014; pp. 702–736. [Google Scholar]
- Mulik, R.S.; Mönkkönen, J.; Juvonen, R.O.; Mahadik, K.R.; Paradkar, A.R. ApoE3 Mediated Poly(Butyl) Cyanoacrylate Nanoparticles Containing Curcumin: Study of Enhanced Activity of Curcumin against Beta Amyloid Induced Cytotoxicity Using in Vitro Cell Culture Model. Mol. Pharm. 2010, 7, 815–825. [Google Scholar] [CrossRef]
- Athalye, M.; Teli, D.; Chorawala, M.; Sharma, A.; Patel, R.; Dua, K.; Singh, S.K.; Gupta, G.; Patel, M. Apolipoprotein E3 Functionalized Lipid-Drug Conjugated Nanoparticles of Levetiracetam for Enhanced Delivery to the Brain: In-Vitro Cell Line Studies and in-Vivo Study. Int. J. Biol. Macromol. 2024, 254, 127799. [Google Scholar] [CrossRef]
- Kreuter, J.; Shamenkov, D.; Petrov, V.; Ramge, P.; Cychutek, K.; Koch-Brandt, C.; Alyautdin, R. Apolipoprotein-Mediated Transport of Nanoparticle-Bound Drugs across the Blood-Brain Barrier. J. Drug Target. 2002, 10, 317–325. [Google Scholar] [CrossRef]
- Michaelis, K.; Hoffmann, M.M.; Dreis, S.; Herbert, E.; Alyautdin, R.N.; Michaelis, M.; Kreuter, J.; Langer, K. Covalent Linkage of Apolipoprotein E to Albumin Nanoparticles Strongly Enhances Drug Transport into the Brain. J. Pharm. Exp. Ther. 2006, 317, 1246–1253. [Google Scholar] [CrossRef]
- Neves, A.R.; Queiroz, J.F.; Weksler, B.; Romero, I.A.; Couraud, P.-O.; Reis, S. Solid Lipid Nanoparticles as a Vehicle for Brain-Targeted Drug Delivery: Two New Strategies of Functionalization with Apolipoprotein E. Nanotechnology 2015, 26, 495103. [Google Scholar] [CrossRef]
- Neves, A.R.; Queiroz, J.F.; Lima, S.A.C.; Reis, S. Apo E-Functionalization of Solid Lipid Nanoparticles Enhances Brain Drug Delivery: Uptake Mechanism and Transport Pathways. Bioconjug. Chem. 2017, 28, 995–1004. [Google Scholar] [CrossRef]
- Dal Magro, R.; Ornaghi, F.; Cambianica, I.; Beretta, S.; Re, F.; Musicanti, C.; Rigolio, R.; Donzelli, E.; Canta, A.; Ballarini, E.; et al. ApoE-Modified Solid Lipid Nanoparticles: A Feasible Strategy to Cross the Blood-Brain Barrier. J. Control Release 2017, 249, 103–110. [Google Scholar] [CrossRef]
- Petri, B.; Bootz, A.; Khalansky, A.; Hekmatara, T.; Müller, R.; Uhl, R.; Kreuter, J.; Gelperina, S. Chemotherapy of Brain Tumour Using Doxorubicin Bound to Surfactant-Coated Poly(Butyl Cyanoacrylate) Nanoparticles: Revisiting the Role of Surfactants. J. Control. Release 2007, 117, 51–58. [Google Scholar] [CrossRef]
- Kreuter, J.; Hekmatara, T.; Dreis, S.; Vogel, T.; Gelperina, S.; Langer, K. Covalent Attachment of Apolipoprotein A-I and Apolipoprotein B-100 to Albumin Nanoparticles Enables Drug Transport into the Brain. J. Control Release 2007, 118, 54–58. [Google Scholar] [CrossRef]
- McNamara, M.G.; Mason, W.P. Antiangiogenic Therapies in Glioblastoma Multiforme. Expert. Rev. Anticancer. Ther. 2012, 12, 643–654. [Google Scholar] [CrossRef]
- Lebelt, A.; Dziecioł, J.; Guzińska-Ustymowicz, K.; Lemancewicz, D.; Zimnoch, L.; Czykier, E. Angiogenesis in Gliomas. Folia Histochem. Cytobiol. 2008, 46, 69–72. [Google Scholar] [CrossRef]
- Hardee, M.E.; Zagzag, D. Mechanisms of Glioma-Associated Neovascularization. Am. J. Pathol. 2012, 181, 1126–1141. [Google Scholar] [CrossRef]
- Jo, J.; Schiff, D.; Purow, B. Angiogenic Inhibition in High-Grade Gliomas: Past, Present and Future. Expert. Rev. Neurother. 2012, 12, 733–747. [Google Scholar] [CrossRef]
- Brodbelt, A.; Greenberg, D.; Winters, T.; Williams, M.; Vernon, S.; Collins, V.P.; (UK) National Cancer Information Network Brain Tumour Group. Glioblastoma in England: 2007–2011. Eur. J. Cancer 2015, 51, 533–542. [Google Scholar] [CrossRef]
- Omuro, A.; DeAngelis, L.M. Glioblastoma and Other Malignant Gliomas: A Clinical Review. JAMA 2013, 310, 1842–1850. [Google Scholar] [CrossRef]
- Wait, S.D.; Prabhu, R.S.; Burri, S.H.; Atkins, T.G.; Asher, A.L. Polymeric Drug Delivery for the Treatment of Glioblastoma. Neuro Oncol. 2015, 17 (Suppl. S2), ii9–ii23. [Google Scholar] [CrossRef]
- Jin, H.; Varner, J. Integrins: Roles in Cancer Development and as Treatment Targets. Br. J. Cancer 2004, 90, 561–565. [Google Scholar] [CrossRef]
- Dubey, P.K.; Singodia, D.; Verma, R.K.; Vyas, S.P. RGD Modified Albumin Nanospheres for Tumour Vasculature Targeting. J. Pharm. Pharmacol. 2011, 63, 33–40. [Google Scholar] [CrossRef]
- Sheikh, A.; Alhakamy, N.A.; Md, S.; Kesharwani, P. Recent Progress of RGD Modified Liposomes as Multistage Rocket Against Cancer. Front. Pharmacol. 2022, 12, 803304. [Google Scholar] [CrossRef]
- Hosseinikhah, S.M.; Vahdat-Lasemi, F.; Farhoudi, L.; Gupta, G.; Kesharwani, P.; Sahebkar, A. RGD-Decorated Nanoparticles: Therapeutic Potential beyond Cancer. J. Drug Deliv. Sci. Technol. 2024, 98, 105924. [Google Scholar] [CrossRef]
- Liang, X.; Wang, H.; Grice, J.E.; Li, L.; Liu, X.; Xu, Z.P.; Roberts, M.S. Physiologically Based Pharmacokinetic Model for Long-Circulating Inorganic Nanoparticles. Nano Lett. 2016, 16, 939–945. [Google Scholar] [CrossRef]
- Huang, H.; Feng, W.; Chen, Y.; Shi, J. Inorganic Nanoparticles in Clinical Trials and Translations. Nano Today 2020, 35, 100972. [Google Scholar] [CrossRef]
- Liu, Q.; Kim, Y.J.; Im, G.B.; Zhu, J.; Wu, Y.; Liu, Y.; Bhang, S.H. Inorganic Nanoparticles Applied as Functional Therapeutics. Adv. Funct. Mater. 2021, 31, 2008171. [Google Scholar] [CrossRef]
- Chen, X.; Chen, Y.; Zou, L.; Zhang, X.; Dong, Y.; Tang, J.; McClements, D.J.; Liu, W. Plant-Based Nanoparticles Prepared from Proteins and Phospholipids Consisting of a Core-Multilayer-Shell Structure: Fabrication, Stability, and Foamability. J. Agric. Food Chem. 2019, 67, 6574–6584. [Google Scholar] [CrossRef]
- Gan, Z.; Lokugamage, M.P.; Hatit, M.Z.C.; Loughrey, D.; Paunovska, K.; Sato, M.; Cristian, A.; Dahlman, J.E. Nanoparticles Containing Constrained Phospholipids Deliver MRNA to Liver Immune Cells in Vivo without Targeting Ligands. Bioeng. Transl. Med. 2020, 5, e10161. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Q.; Li, Z.; Li, F.; Wu, D.; Fan, M.; Zheng, A.; Huang, B.; Gan, L.; Zhao, Y.; et al. Hydrophobicity-Adaptive Nanogels for Programmed Anticancer Drug Delivery. Nano Lett. 2018, 18, 7909–7918. [Google Scholar] [CrossRef]
- Cao, Z.; Zhou, X.; Wang, G. Selective Release of Hydrophobic and Hydrophilic Cargos from Multi-Stimuli-Responsive Nanogels. ACS Appl. Mater. Interfaces 2016, 8, 28888–28896. [Google Scholar] [CrossRef]
- Karpenko, L.I.; Apartsin, E.K.; Dudko, S.G.; Starostina, E.V.; Kaplina, O.N.; Antonets, D.V.; Volosnikova, E.A.; Zaitsev, B.N.; Bakulina, A.Y.; Venyaminova, A.G.; et al. Cationic Polymers for the Delivery of the Ebola Dna Vaccine Encoding Artificial T-Cell Immunogen. Vaccines 2020, 8, 718. [Google Scholar] [CrossRef]
- Zhang, M.; Hong, Y.; Chen, W.; Wang, C. Polymers for DNA Vaccine Delivery. ACS Biomater. Sci. Eng. 2017, 3, 108–125. [Google Scholar] [CrossRef]
- Maillard, D.; Kumar, S.K.; Fragneaud, B.; Kysar, J.W.; Rungta, A.; Benicewicz, B.C.; Deng, H.; Brinson, L.C.; Douglas, J.F. Mechanical Properties of Thin Glassy Polymer Films Filled with Spherical Polymer-Grafted Nanoparticles. Nano Lett. 2012, 12, 3909–3914. [Google Scholar] [CrossRef]
- He, H.; Lu, Y.; Qi, J.; Zhu, Q.; Chen, Z.; Wu, W. Adapting Liposomes for Oral Drug Delivery. Acta Pharm. Sin. B 2019, 9, 36–48. [Google Scholar] [CrossRef]
- Wang, Q.; Qin, X.; Fang, J.; Sun, X. Nanomedicines for the Treatment of Rheumatoid Arthritis: State of Art and Potential Therapeutic Strategies. Acta Pharm. Sin. B 2021, 11, 1158–1174. [Google Scholar] [CrossRef]
- Dai, T.; Jiang, K.; Lu, W. Liposomes and Lipid Disks Traverse the BBB and BBTB as Intact Forms as Revealed by Two-Step Förster Resonance Energy Transfer Imaging. Acta Pharm. Sin. B 2018, 8, 261–271. [Google Scholar] [CrossRef]
- Barenholz, Y. Liposome Application: Problems and Prospects. Curr. Opin. Colloid. Interface Sci. 2001, 6, 66–77. [Google Scholar] [CrossRef]
- Hartl, N.; Gabold, B.; Uhl, P.; Kromer, A.; Xiao, X.; Fricker, G.; Mier, W.; Liu, R.; Merkel, O.M. ApoE—Functionalization of Nanoparticles for Targeted Brain Delivery—A Feasible Method for Polyplexes? Drug Deliv. Transl. Res. 2024, 14, 1660–1677. [Google Scholar] [CrossRef]
- Zhang, P.; Cao, F.; Zhang, J.; Tan, Y.; Yao, S. Temozolomide and Chloroquine Co-Loaded Mesoporous Silica Nanoparticles Are Effective against Glioma. Heliyon 2023, 9, e18490. [Google Scholar] [CrossRef]
- Sahel, D.K.; Goswami, S.G.; Jatyan, R.; Tharmatt, A.; Singh, V.; Dalela, M.; Mohanty, S.; Mittal, A.; Ramalingam, S.; Chitkara, D. CRGD-Modified Hybrid Lipopolymeric Nanoplexes for Gene Editing in the Posterior Segment of the Eye. Int. J. Biol. Macromol. 2024, 271, 132426. [Google Scholar] [CrossRef]
- Northrop, B.H.; Frayne, S.H.; Choudhary, U. Thiol–Maleimide “Click” Chemistry: Evaluating the Influence of Solvent, Initiator, and Thiol on the Reaction Mechanism, Kinetics, and Selectivity. Polym. Chem. 2015, 6, 3415–3430. [Google Scholar] [CrossRef]
- Ashraf, S.; Hassan Said, A.; Hartmann, R.; Assmann, M.; Feliu, N.; Lenz, P.; Parak, W.J. Analyse Quantitativer Partikelaufnahme von Zellen Über Verschiedene Messmethoden. Angewandte Chem. 2020, 132, 5478–5494. [Google Scholar] [CrossRef]
- Yamamoto, M.; Ikeda, K.; Ohshima, K.; Tsugu, H.; Kimura, H.; Tomonaga, M. Increased Expression of Low Density Lipoprotein Receptor-Related Protein/Alpha2-Macroglobulin Receptor in Human Malignant Astrocytomas. Cancer Res. 1997, 57, 2799–2805. [Google Scholar] [PubMed]
- Yamamoto, M.; Ikeda, K.; Ohshima, K.; Tsugu, H.; Kimura, H.; Tomonaga, M. Expression and Cellular Localization of Low-Density Lipoprotein Receptor-Related Protein/Alpha 2-Macroglobulin Receptor in Human Glioblastoma in Vivo. Brain Tumor Pathol. 1998, 15, 23–30. [Google Scholar] [CrossRef]
- Maletínská, L.; Blakely, E.A.; Bjornstad, K.A.; Deen, D.F.; Knoff, L.J.; Forte, T.M. Human Glioblastoma Cell Lines: Levels of Low-Density Lipoprotein Receptor and Low-Density Lipoprotein Receptor-Related Protein. Cancer Res. 2000, 60, 2300–2303. [Google Scholar] [PubMed]
- N. R., S.; Behera, M.M.; Naik, S.K.; Das, S.K.; Gopan, S.; Ghosh, A.; Sahu, R.N.; Patra, S.; Purkait, S. Elevated Expression of Cholesterol Transporter LRP-1 Is Crucially Implicated in the Pathobiology of Glioblastoma. Front. Neurol. 2022, 13, 1003730. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.E.; Zahid, M. Cell Penetrating Peptides, Novel Vectors for Gene Therapy. Pharmaceutics 2020, 12, 225. [Google Scholar] [CrossRef] [PubMed]
- Danhier, F.; Pourcelle, V.; Marchand-Brynaert, J.; Jérôme, C.; Feron, O.; Préat, V. Targeting of Tumor Endothelium by RGD-Grafted PLGA-Nanoparticles. In Methods in Enzymology; Academic Press Inc.: Cambridge, MA, USA, 2012; Volume 508, pp. 157–175. [Google Scholar] [CrossRef]
- Ducassou, A.; Uro-Coste, E.; Verrelle, P.; Filleron, T.; Benouaich-Amiel, A.; Lubrano, V.; Sol, J.C.; Delisle, M.B.; Favre, G.; Ken, S.; et al. Avβ3 Integrin and Fibroblast Growth Factor Receptor 1 (FGFR1): Prognostic Factors in a Phase I-II Clinical Trial Associating Continuous Administration of Tipifarnib with Radiotherapy for Patients with Newly Diagnosed Glioblastoma. Eur. J. Cancer 2013, 49, 2161–2169. [Google Scholar] [CrossRef]
- Marzolo, M.P.; Yuseff, M.I.; Retamal, C.; Donoso, M.; Ezquer, F.; Farfán, P.; Li, Y.; Bu, G. Differential Distribution of Low-Density Lipoprotein-Receptor-Related Protein (LRP) and Megalin in Polarized Epithelial Cells Is Determined by Their Cytoplasmic Domains. Traffic 2003, 4, 273–288. [Google Scholar] [CrossRef]
- Donoso, M.; Cancino, J.; Lee, J.; Van Kerkhof, P.; Retamal, C.; Bu, G.; Gonzalez, A.; Cáceres, A.; Marzolo, M.-P. Polarized Traffic of LRP1 Involves AP1B and SNX17 Operating on Y-Dependent Sorting Motifs in Different Pathways. Mol. Biol. Cell 2009, 20, 481–497. [Google Scholar] [CrossRef]
- Van Kerkhof, P.; Lee, J.; McCormick, L.; Tetrault, E.; Lu, W.; Schoenfish, M.; Oorschot, V.; Strous, G.J.; Klumperman, J.; Bu, G. Sorting Nexin 17 Facilitates LRP Recycling in the Early Endosome. EMBO J. 2005, 24, 2851–2861. [Google Scholar] [CrossRef]
- Topal, G.R.; Mészáros, M.; Porkoláb, G.; Szecskó, A.; Polgár, T.F.; Siklós, L.; Deli, M.A.; Veszelka, S.; Bozkir, A. ApoE-Targeting Increases the Transfer of Solid Lipid Nanoparticles with Donepezil Cargo across a Culture Model of the Blood–Brain Barrier. Pharmaceutics 2021, 13, 38. [Google Scholar] [CrossRef]
- Ismail, M.; Yang, W.; Li, Y.; Chai, T.; Zhang, D.; Du, Q.; Muhammad, P.; Hanif, S.; Zheng, M.; Shi, B. Targeted Liposomes for Combined Delivery of Artesunate and Temozolomide to Resistant Glioblastoma. Biomaterials 2022, 287, 121608. [Google Scholar] [CrossRef]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in Cancer: Biological Implications and Therapeutic Opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar] [CrossRef]
- Hersel, U.; Dahmen, C.; Kessler, H. RGD Modified Polymers: Biomaterials for Stimulated Cell Adhesion and Beyond. Biomaterials 2003, 24, 4385–4415. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Deng, J.; Zhao, Y.; Tao, T. Cyclic RGD Peptide-Modified Liposomal Drug Delivery System: Enhanced Cellular Uptake in Vitro and Improved Pharmacokinetics in Rats. Int. J. Nanomed. 2012, 7, 3803–3811. [Google Scholar] [CrossRef] [PubMed]
- Dou, X.; Nomoto, T.; Takemoto, H.; Matsui, M.; Tomoda, K.; Nishiyama, N. Effect of Multiple Cyclic RGD Peptides on Tumor Accumulation and Intratumoral Distribution of IRDye 700DX-Conjugated Polymers. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Chin, D.; Hagl, S.; Hoehn, A.; Huebbe, P.; Pallauf, K.; Grune, T.; Frank, J.; Eckert, G.P.; Rimbach, G. Adenosine Triphosphate Concentrations Are Higher in the Brain of APOE3- Compared to APOE4-Targeted Replacement Mice and Can Be Modulated by Curcumin. Genes. Nutr. 2014, 9, 397. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Nomenclature | Control | mPEG | Mal | ApoE3 | c(RGD) |
|---|---|---|---|---|---|
| Components | Molar Ratio [%] | ||||
| DPPC | 59.9 | 54.9 | 54.8 | 54.8 | 49.9 |
| Cholesterol | 40 | 40 | 40 | 40 | 40 |
| DSPE-PEG2k | - | 5 | 5 | 5 | 5 |
| DSPE-PEG5k-Mal | - | - | 0.1 | - | - |
| DSPE-PEG5k-Mal-ApoE3 | - | - | - | 0.1 | - |
| DSPE-PEG5k-c(RGDyk) | - | - | - | - | 5 |
| DiD | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| Liposomes | Z-Average [nm] | PdI |
|---|---|---|
| Control | 110.8 ± 2.0 | 0.106 ± 0.020 |
| mPEG | 108.5 ± 1.9 | 0.065 ± 0.028 |
| Mal | 103.8 ± 0.7 | 0.136 ± 0.014 |
| ApoE3 | 125.1 ± 1.1 | 0.195 ± 0.012 |
| c(RGD) | 99.6 ± 2.3 | 0.143 ± 0.037 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lubitz, L.J.; Haffner, M.P.; Rieger, H.; Leneweit, G. Increased Cellular Uptake of ApoE3- or c(RGD)-Modified Liposomes for Glioblastoma Therapy Depending on the Target Cells. Pharmaceutics 2024, 16, 1112. https://doi.org/10.3390/pharmaceutics16091112
Lubitz LJ, Haffner MP, Rieger H, Leneweit G. Increased Cellular Uptake of ApoE3- or c(RGD)-Modified Liposomes for Glioblastoma Therapy Depending on the Target Cells. Pharmaceutics. 2024; 16(9):1112. https://doi.org/10.3390/pharmaceutics16091112
Chicago/Turabian StyleLubitz, Larissa J., Moritz P. Haffner, Harden Rieger, and Gero Leneweit. 2024. "Increased Cellular Uptake of ApoE3- or c(RGD)-Modified Liposomes for Glioblastoma Therapy Depending on the Target Cells" Pharmaceutics 16, no. 9: 1112. https://doi.org/10.3390/pharmaceutics16091112