Abstract
The increasing usage of chemical control agents, as well as fungicides to manage plant diseases, causes human and environmental health problems. Macroalgae represent a reservoir for a tremendous variety of secondary metabolites that display a wide range of biological activities. However, their anti-phytopathogenic properties are still being studied. The current study was conducted to investigate whether or not the macroalgae Ulva fasciata extract exhibits antifungal and antiviral activities. In this regard, the organic extracts of U. fasciata were tested for their capabilities against tobacco mosaic virus (TMV) and three molecularly identified fungal isolates, Fusarium verticillioides, Alternaria tenuissima, and Botrytis cinerea with accession numbers OP363619, OP363620, and OP363621, respectively. Among the three tested extract concentrations, 100 µg/mL had the best biological activity against B. cinerea and TMV, with 69.26%and 81.25% inhibition rates, respectively. The HPLC analysis of chemical profiling of the extract showed the presence of a number of phenolic and flavonoid compounds widely known to display many biological activities. In this line, the 4-Hydroxybenzoic acid was the highest phenolic compound (12.3 µg/mL) present in the extract, followed by ferulic acid (9.05 µg/mL). The 7-hydroxyflavone (12.45 µg/mL) was the highest flavonoid in the organic extract of U. faciata followed by rutin, which recorded a concentration of 7.62 µg/ mL. The results of this study show that the U. fasciata extract has antiviral and antifungal properties, which makes it a possible source of natural antimicrobial agents.
Keywords:
Ulva fasciata; macroalgae; plant disease; antiviral; antifungal; TMV; HPLC; biocontrol agent 1. Introduction
Plant diseases have resulted in significant crop losses worldwide, further complicating already major issues with food security [1]. Plant viruses are the most dangerous diseases for plants, and once they appear in the field, they quickly spread and result in significant problems for agricultural productivity [2]. The tobacco mosaic virus (TMV) is a well-known plant virus that has a large, negative impact on crop quality and productivity worldwide [3]. In plant pathology, the tobacco mosaic virus (TMV) is one of the top 10 plant viruses [4]. It is also used as a model virus, and Nicotiana glutinosa is used as a host for TMV-local lesions in many biological studies [5]. Egyptian tomatoes are one of the country’s most valuable crops, ranked fifth in production worldwide [6,7]. Tomatoes are economically important in Egypt because they are less expensive than other vegetables with higher nutritional content. Moreover, their demand and consumption are rapidly increasing due to population growth [7]. Fungal plant infections cause damage to different plants, which may cause the loss of crop harvests or, in the most severe cases, the death of the plants [2,8]. Crop quality is also affected negatively by fungal pathogens, which may cause the accumulation of toxins within plant tissues, which leads to harmful health effects for humans and animals [9]. Although many fungal strains are reported to cause serious plant diseases, it was also noted that specific fungal strains are capable of removing toxins from habitats [10]. Fungal pathogens that can spread through the air and soil and cause diseases such as root rot, wilts, and grey mold greatly impact crops, especially tomatoes, and cause production losses [11,12]. Several fungi bring on wilt infections in tomato plants, the most common of which are Fusaria, such as Fusarium oxysporum, F. verticillioides, and F. equiseti [13]. Moreover, Botrytis cinerea and Alternaria tenuissima fungi were associated with different symptoms of tomato grey mold disease and leaf spot [14,15]. PCR markers, for example, are fast and accurate fungus detection methods that can be employed for this purpose. Inter transcript spacer region (ITS) and elongation factor 1-alpha gene (TEF-1) DNA sequences were found to be better genetic markers for identifying closely related Fusarium spp. [16,17] than identification and quantification based on morphological features of fungal pathogens, which took more time and space in the lab to keep cultures. In a study performed by Chehri [18], the isolated Fusaria species from tomato plants were classified into four species based on morphological characteristics and tef1 and tub2 gene sequence data: F. oxysporum, F. redolens, F. proliferatum, and F. verticillioides. Stracquadanio et al. [19] found that symptoms of decay were linked to the production of mycotoxins in tomato fruits infected with F. verticillioides. Meanwhile, Chang et al. [20] molecularly identified seven Fusaria species isolated from tomato Fusarium wilt disease and reported four of them, including F. oxysporum, F. solani, F. subglutinans, and F. verticillioides, were pathogenic to tomato plants.
Applying chemical fungicides to plants or developing transgenic plants are the two most common strategies for eradicating plant diseases. However, synthetic pesticides harm humans and the environment, and transgenic crops have not yet gained widespread acceptance [1,21]. So, finding new antiviral methods safe for the environment is still a priority. Recently, algae have been used as biocontrol agents against plant diseases [8]. Compared to synthetic and semi-synthetic alternatives, fungicides made from seaweed extracts are significantly less harmful [22]. It has been shown that red algal seaweed extracts contain chemicals that kill fungi, which has been the subject of many studies in the past [23]. Many macroalgae are considered promising sources for producing fungicide products [24]. Many previous reports prove that the extracts of many macroalgae, especially red and brown ones exert high antifungal activity against many plant pathogens such as Fusarium moniliforme, Rhizoctonia solani, and Phytophthora cinnamomi [25,26].
Green algae represent the largest algal group found on earth and inhabit different ecosystems, including fresh and marine habitats [27]. They range from unicellular to multi-cellular, microscopic to macroscopic forms. Their thalli vary from free filaments to shaped forms [27]. Green algae are characterized by the production of a wide range of metabolites, including polysaccharides, polyphenols, terpenes, and carotenoids which play many different biological activities such as antimicrobial, antioxidant, and antitumor activities [28]. Ulva is one of the most widely distributed green algal genera known as sea lettuce [29]. Ulva is known to be a good source of food, development of novel drugs and functional foods, and pharmaceutical, in addition to different agricultural applications [30]. It has proven to be a rich source of structurally diverse bioactive compounds with valuable biomedical potential [31]. The famous ulvan product produced exclusively by the Ulva genera is a water-soluble polysaccharide with many biological activities, including anticancer and antimicrobial [32].
Until now, the application of Ulva fasciata extracts as biocontrol agents against plant pathogens, antiviral and antifungal, is still limited. So, in this work, we aimed to analyze the phytochemical compounds of U. fasciata extract using high-performance liquid chromatography (HPLC) as well as evaluate its antiviral activity against tobacco mosaic virus, determine whether or not the extract inhibited the growth of three different types of fungi F. verticillioides, A. tenuissima, and B. cinerea.
2. Materials and Methods
2.1. Sampling Site
Ulva fasciata biomasses were handpicked during the mid-autumn of 2021 at a depth of 0.2 m or less from Ras El Bar coast. Ras El Bar lies in a peninsula on the coast of the Mediterranean Sea, and it is bordered on the western by the Damietta Nile branch at (N 31°30′45″ E 31°49′32″). The Ulva fasciata biomasses were collected straight from the intertidal zone using five quadrants (1 mm) according to the Londo scale [33]. The fresh algal biomass sample size followed the rules of the Egyptian Environmental Affairs Agency (EEAA) assigned for the bio-conservation of the protected areas.
2.2. Preparation of Dried Algal Biomass
The alga was rinsed several times with saline water and distilled water to remove epiphytes and sediments. The alga was identified based on their morphology and anatomy and by using taxonomic references according to Böer and Warnken [34] and Komárek and Zapomělová [35]. After drying the algal biomass in the shade, the sample was cut, ground, and stored in a firmly sealed dark jar.
2.3. Extraction of Ulva fasciata Biomass
In order to extract Ulva fasciata biomass, 25 g of the dried alga biomass was extracted using the Soxhlet extraction method. The solvent used in the extraction process was a mixture of methanol and hexane with (a 1:1) ratio. The extraction steps were repeated several times until most chemical compounds in the alga biomass were extracted. The organic extract was cooled and concentrated using a rotary evaporator at 30–45 °C. The remnant dried residue of the extract was redissolved using 3 mL of DMSO and kept at 4 °C until use. The extraction procedures were repeated using a solvent mixture only to prepare a control sample.
2.4. Viral Source and Antiviral Activity Assays
The strain KH1 of the Egyptian TMV (Acc# MG264131) [3] was inoculated and propagated on tobacco plants under greenhouse conditions. Using the procedures described by Gooding and Hebert [36], the TMV was purified and adjusted to an inoculum concentration of 20 μg/mL using 100 mM sodium phosphate buffer, pH 7.2. The 100 mg/mL stock concentration of U. fasciata extract was prepared by dissolving 100 mg of U. fasciata extract in 1 mL DMSO. After that, the three working solutions with 25 μg/mL, 50 μg/mL, and 100 μg/mL were prepared and used as three different antiviral treatment assays. A DMSO solution at an equivalent concentration was used as a negative control treatment. Antiviral efficacy was measured using the leaf assay method, in which Nicotiana glutinosa was used as a TMV local lesion host. Based on the developed local lesion on the inoculated leaves, the percentage of inhibitory effect was calculated using the formula: [I = (1 − T/C) × 100], where I is the percentage of inhibitory effect, T is the number of local lesions on the treated leaves, and C is the number of local lesions on the control leaves. The antiviral experiment assay was performed in five treatments under greenhouse conditions on N. glutinosa plants at the 5–6 leaf stage. The first treatment was the Mock treatment, in which the N. glutinosa leaves were sprayed with free-extract working DMSO solution and then inoculated with viral inoculation buffer. The second treatment was TMV treatment, in which the N. glutinosa leaves were sprayed with free-extract working DMSO solution and then inoculated with TMV. The third, fourth, and fifth treatments were 25 μg/mL, 50 μg/mL, and 100 μg/mL treatments, in which N. glutinosa leaves were sprayed with 25 μg/mL, 50 μg/mL, and 100 μg/mL of U. fasciata extract, respectively, 24 h before inoculation with TMV. The N. glutinosa leaves were foliar sprayed with a sterilized plastic hand sprayer until runoff occurred, and the leaves seemed to be coated with the U. fasciata extract. As previously described, the N. glutinosa leaves were mechanically inoculated with TMV [37]. Briefly, the upper surface of the N. glutinosa leaves was dusted with carborundum (600-mesh) and then gently inoculated by finger-rubbing with 20 μL TMV inoculum. After 5 min of the viral inoculation, the inoculated leaves were rinsed with sterile distilled water. Each experiment was repeated three times. At four days post-TMV inoculation (dpi), the number of local lesions was photographed and contented.
2.5. Ulva fasciata Extract Antifungal Assay
2.5.1. Isolation of Fungi
Tissues from infected tomato plants were examined for signs of stem wilting, leaf early blight, and tomato fruit postharvest rots. The fungal pathogens were recovered from the infected samples when grown in potato dextrose agar (PDA) media plates. For purification, the single spore culture method is used [38]. Once the fungi were purified, they were transferred to slant tubes and cultivated at room temperature for four days. Microscopic phenotypic identification and genetic characterization of the pure cultures were performed.
2.5.2. Nuclear Ribosomal Internal Transcribed Spacer (ITS) Region Amplification in Fungi
The Rapid Fungal Genomic DNA Isolation Kit (CAT. #: FT71415, NBS Biologicals Ltd., Huntingdon, UK) is used to isolate total DNA from various fungal species in a simple and efficient manner. The ITS region was amplified according to the procedure of White et al. [39]. The ITS PCR reactions were sequenced at Macrogen Company (Seoul, Korea), and the sequences were assembled, and Genbank deposited.
2.5.3. Antifungal Assay
The antifungal activity of Ulva fasciata extract was examined against the three plant pathogenic fungi by the food poison technique [40]. In brief, the extract-treated Petri-plates were created by adding 250, 500, and 1000 µg of U. fasciata extract/10 mL of sterilized potato dextrose agar media (PDA) to achieve the desired final concentrations (25, 50, and 100 µg/mL) compared to the control (without extract treatment). A total of 10 mL of the extract-treated PDA was poured into the plates. After PDA solidification, one disc (0.5 cm in diameter) of each freshly grown tested fungus was placed in the middle of the Petri plate of each treatment. The fungal species were placed in an incubator at 28 °C till the growth of the control fungus filled the plate. All the treatments were repeated three times. Finally, the antifungal activity was achieved by recording the inhibition percentage of fungal hyphae as follows the formula:
2.6. HPLC Analysis
The phenolic and flavonoid compounds were detected and quantified in the U. fasciata biomass using HPLC methods. The characterization condition was previously described in previous works. The phenolic compounds were characterized and quantified using a Quaternary pump and a Zorbax Eclipse Plus C18 column in an Agilent HPLC (1260 Infinity Series, 100 mm × 4.6 mm i.d.). Analytical grade water, 0.2% H3PO4 (v/v), methanol, and acetonitrile were used in gradient elution to achieve separation. A detector with a tunable wavelength of 284 nm was employed. Flavonoid compounds were analyzed using an HPLC fitted with a binary pump (Zorbax Eclipse plus C18 column, 150 mm × 4.6 mm i.d.). To accomplish the separation, a mixture of methanol and water (50:50) comprising 0.5% H3PO4 and flowing at a rate of 0.7 mL/min was used as the eluent. The extract was injected at a volume of 20 µL, the UV detector was adjusted to 273 nm, and the results were compared with standard compounds and integrated. Phenolic and flavonoid compounds were defined by caffeic acid, ferulic acid, chlorogenic acid, 4-hydroxybenzoic acid, salicylic acid, ellagic acid, protocatechuic acid, rutin, quercetin, kaempferol, luteolin, and 7-hydroxyflavone standard compounds, which were purchased from Merck KGaA (Darmstadt, Germany).
2.7. Data Analysis
The results obtained were statistically analyzed using one-way ANOVA. Comparisons among mean values obtained were made using Tukey’s HSD at the 0.05 level of probability.
3. Results
3.1. Antiviral Activity Assay of U. fasciata Extract
The antiviral efficacy of U. fasciata extract against TMV was evaluated on N. glutinosa under greenhouse-controlled conditions. The leaf assay assessed the protective activity by counting the number of local lesions on inoculated leaves four days after the TMV challenge. Compared to untreated N. glutinosa leaves, U. fasciata extract-treated tissues 24 h before TMV inoculation showed a considerable reduction in local lesion numbers compared to untreated ones at different concentrations (Figure 1). The leaves of mock-treated plants exhibited no local lesions. The highest antiviral activity, with a rate of inhibition of 81.25%, was seen when 100 µg/mL of U. fasciata extract was applied (Figure 2). At 50 and 25 µg/mL doses, the inhibition rates were determined to be 72.08 and 57.92%, respectively (Figure 2). According to the findings, an extract of U. fasciata can potentially protect tomato plants against a TMV infection. Therefore, it can be used as a biocontrol agent to inhibit and/or prevent TMV infection.

Figure 1.
A photograph of the signs of disease on N. glutinosa leaves infected with the tobacco mosaic virus at 4 days post-inoculation with different concentrations of U. fasciata extract. Mock: N. glutinosa leaves sprayed with free-extract working DMSO solution and then inoculated with viral inoculation buffer; TMV: N. glutinosa leaves sprayed with free-extract working DMSO solution and then inoculated with TMV; U. fasciata: N. glutinosa leaves were sprayed with 25 μg/mL, 50 μg/mL, and 100 μg/mL of U. fasciata extract, respectively, 24 h before inoculation with TMV.
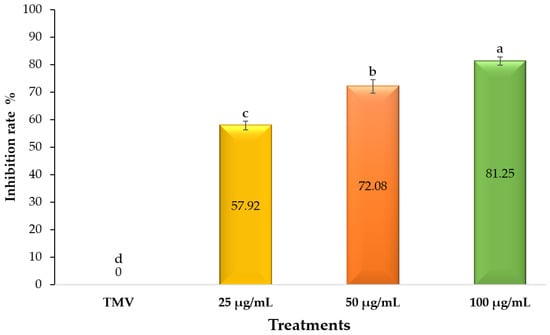
Figure 2.
A histogram showing the inhibition rates of different concentrations of U. fasciata extract on N. glutinosa leaves infected with tobacco mosaic virus at 4 days post-inoculation. TMV: N. glutinosa leaves sprayed with free-extract working DMSO solution and then inoculated with TMV; U. fasciata: N. glutinosa leaves were sprayed with 25 μg/mL, 50 μg/mL, and 100 μg/mL of U. fasciata extract, respectively, 24 h before inoculation with TMV.
3.2. Ulva fasciata Extract Activity against the Fungi
3.2.1. Fungal Pathogens used and Initial Morphological Identification
The isolation and purification trails from tomato plant parts revealed three fungal species. The initial phenotypic characters showed that they were A. tenuissima, F. verticillioides, and B. cinerea.
3.2.2. ITS Molecular Identification
The ITS blasted sequence results confirmed the phenotypic characterization. All the sequence data were deposited in Genbank (NCBI) under the scientific names A. tenuissima isolate At-12, F. verticillioides isolate Fv-11, and B. cinerea isolate Bc-13 and have accessioned with numbers OP363620, OP363619, and OP363621, respectively. The phylogenetic tree of all the isolates was drawn with aligned sequences obtained from the Genbank nucleotide site, as presented in Figure 3.

Figure 3.
Phylogenetic tree of the three fungal isolates identified in this study by ITS region amplified primers compared with sequences retrieved from the Genbank. The three characterized isolates were A. tenuissima isolate At-12 (OP363620), F. verticillioides isolate Fv-11 (OP363619), and B. cinerea isolate Bc-13 (OP363621).
3.2.3. Ulva fasciata Extract Antifungal Activity
The organic extract of Ulva fasciata displayed different antifungal activities against the three fungi used in this study. The results of the antifungal activity of the green alga organic extract are clearly shown in Table 1 and Figure 4. The highest antifungal efficacy of the extract was detected against B. cinerea using 100 and 50 µg/mL of the extract, with values of 69.26 and 59.26%, respectively. The extract causes a good fungal inhibition against F. verticillioides and A. tenuissima, with a percent of mycelial inhibition reaching 51.85 and 55.93% using 100 µg/mL of the extract, respectively.

Table 1.
Antifungal activity of the Ulva fasciata extract.
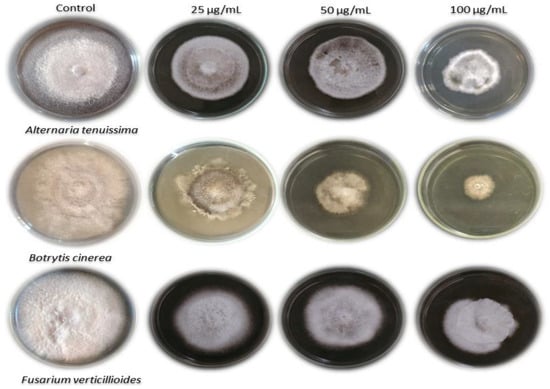
Figure 4.
The activity of Ulva fasciata extract against the growth of A. tenuissima, B. cinerea, and F. verticillioides.
3.3. HPLC Analysis of the Phenolic and Flavonoid Compounds
The HPLC chromatograms of phenolic and flavonoid compounds in the Ulva fasciata extract are clearly in Figure 5. The chemical characterization of the Ulva fasciata extract showed a number of phenolic compounds with the highest concentration recorded by 4-hydroxybenzoic acid (12.33 µg/mL). Similarly, the chemical characterization of the offered number of flavonoid compounds in the alga extract. The highest flavonoid compound recorded in the alga extract was 7-hydroxyflavone, with a concentration of 12.45 µg/ mL (Table 2).
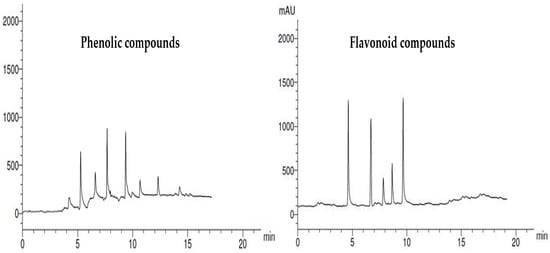
Figure 5.
HPLC chromatograms of phenolic and flavonoid compounds identified in Ulva fasciata extract.
Table 2.
The concentration of phenolic and flavonoid compounds present in the extract of Ulva fasciata using HPLC.
4. Discussion
One of the most significant challenges facing global food security is the prevalence of plant diseases [41]. They cause farmers to lose billions of dollars each year [42]. Using resistant plant cultivars or large amounts of fungicides is necessary for disease control in plants, but both approaches pose risks to human and environmental health [43]. Moreover, surface water pollution from pesticides is a major contributor to environmental and ecological problems [44]. Instead of using dangerous pesticides, biocontrol agents could be employed to manage plant diseases [45]. Algal extracts are reported as a sustainable and environmentally friendly alternative to chemical control agents because they contain a wide range of secondary metabolites that are good for the plant’s health [46]. These metabolites may boost the plant’s systemic resistance and limit the growth of pathogens. The current study assessed the effectiveness of an extract from Ulva fasciata against three different fungal pathogens and a viral infection caused by the tobacco mosaic virus (TMV). High-performance liquid chromatography (HPLC) was also used to determine the main phytochemical parts of the U. fasciata extract.
Many aquatic organisms are capable of producing a vast number of allelopathic chemicals [47]. The production and release of such allelopathic chemicals could be regarded as a defense mechanism of the aquatic macroalgae against other phototrophic organisms competing for light and nutrients [48]. Among the allelopathic chemical, many previous studies have demonstrated that marine macroalgae produce many bioactive chemicals [23,24,46,49,50]. Sea lettuce “Ulva” is among the most famous green macroalgae used in feeding, especially in coastal countries. In this study, the identity of the green seaweed was verified morphologically and using the microscopic examination of its edges. Similarly, Ulva represents a good source for many metabolites that display different bioactivities, including antimicrobial, anticancer, antioxidant, and immunomodulating activities [51,52,53,54,55].
The phytochemical profile of the extract shows that it contains bioactive phenolic and flavonoid compounds. The results obtained in this work revealed the presence of high phenolic content in the extract of U. fasciata. This finding is consistent with many previous studies [23,56]. Farasat et al. [57] confirmed the presence of high phenolic content in the extracts of four Ulva species collected from the Arabian Gulf. A high phenolic content was identified in the ethanolic extract of the green seaweed Caulerpa racemosa [58]. Similarly, the organic extract of U. fasciata showed a number of flavonoid compounds Like Rutin, Quercetin, and Kaempferol. El-Mesallamy et al. [59] proved the presence of content of flavonoid compounds in the extract of U. lactuca. The phenolic and flavonoid compounds displayed many biological activities, including anticancer, antimicrobial, anti-inflammatory, and immunomodulatory functions [60]. The antifungal activity displayed by the extract of U. fasciata against the three-plant pathogens may be attributed to the synergetic effect of different bioactive compounds present in the organic extract of U. fasciata.
The utilization of helpful microorganisms as a secure and ethical method of preventing viral plant diseases has attracted much attention in recent years [61,62]. However, the application of algae for the biological control of viral plant diseases is still limited, and how they might operate against viral infections is unknown. In the current study, the inhibitory effect of U. fasciata extract against TMV ranged from 57.92% at a dose of 25 µg/mL to 81.25% at a dose of 100 µg/mL, when applied on N. glutinosa tissues. Based on these results, we think that U. fasciata extract has elicitor compounds that either help the immune system fight TMV better or stop it from spreading. Antiviral agents were reported to stop the spread of viruses in one of two ways: either by stopping viral replication directly or by boosting the host’s immune system that activates SAR to fight the infection [63]. Many different polyphenols, such as caffeic acid, ellagic acid, catechins, chlorogenic acid, gallic acid, quercetin, ferulic acid, and myricetin, have been shown to have benefits against bacteria, viruses, inflammation, cancer, and oxidative stress [64,65]. It has been documented that using 500 µg/mL of exogenous caffeic acid stifles the development of several Fusarium and Saccharomyces species [66]. Ferulic acid is a typical phytochemical found in leaves and seeds that are both free and covalently attached to lignin, hydroxy fatty acids, polysaccharides, glycoproteins, and polyamines. Ferulic acid stiffens cell walls and is an antioxidant, antibacterial, antiviral, anticarcinogenic, and chemical that changes how enzymes work [67]. It was reported that kaempferol plays a significant role in the auxin-dependent defense response in CMVsat-infected Arabidopsis plant [68]. Moreover, luteolin, an antiviral bioflavonoid, showed extracellular virucidal activity against Japanese encephalitis virus replication [69]. Due to its inexpensive cost and inhibitory action on plant viruses, quercetin offers great potential for controlling plant virus infections. It was observed that 1 mol/L quercetin inhibited the proliferation of cucumber necrosis virus, turnip crinkle virus, and TMV in Nicotiana benthamiana up to 90% [70,71]. Protocatechuic acid is a compound found in plants that are high in phenolics. Many in vitro studies have shown anti-inflammatory, antimicrobial, and antioxidant effects [72]. Salicylic acid (SA) is a phenolic compound produced by various plant species at different levels that plays an important role in inducing resistance to biotic and abiotic stresses. The exogenous application of SA increased the expression of defense-related genes and antioxidant enzyme activities and enhanced plant resistance to pathogens, including viruses [73]. In light of this, we believe that such detected polyphenolic compounds in the extract could be utilized as elicitor molecules to promote SAR and boost resistance to TMV infection [74]. It can thus be applied as an alternative to chemical pesticides and to develop sustainable, eco-friendly antiviral agents.
The present investigation represents a trial to study the antifungal effect of the organic extract of U. fasciata against common pathogenic fungal species. The fungal species were isolated from tomato plants and molecularly identified as A. tenuissima, F. verticillioides, and B. cinerea. The isolated A. tenuissima and B. cinerea fungi have been associated with tomato symptoms as described before, causing leaf spot and grey mold diseases [14,15]. F. verticillioides is known to infect maize and a few reports. Mwangi et al. [75] have mentioned that this pathogen is associated with tomato stem diseases or is a vascular wilt pathogen of tomatoes. The same was reported by Akbar et al. [76] for F. equiseti as an emerging wilt pathogen. Furthermore, another study mentions that apart from F. oxysporum f.sp. lycopersici, Fusarium incarnatum-equiseti species complex, and F. graminearum produce mild wilt symptoms in tomatoes [77]. The extract of U. fasciata causes considerable mycelial inhibition in B. cinerea, while F. verticillioides and A. tenuissima show good antifungal activity against Rhizoctonia solani. These results are consistent with the limited number of previous reports that studied the antifungal effect of Ulva extracts. Lotfi et al. [78] reported a high antifungal activity of U. lactuca against Macrophomina phaseolina and F. oxysporum. Selim et al. [79] have demonstrated a fungicidal effect of U. lactuca against F. solani, Rhizoctonia solani, Sclerotinia sclerotiorum, Alternaria solani, Phytophthora infestans, and B. cinerea. Meanwhile, the antimicrobial activities of methanol, ethanol and acetone extracts of U. fasciata and lactuca species were reported against Bacillus subtilis, Staphylococcus aureus, Escherichia coli, S. typhi, Klebsiella pneumonia, and Candida albicans [80]. In another study conducted by Kosanic et al. [81], the U. lactuca acetone extract showed weak minimum fungal inhibitory concentration against Aspergillus flavus, A. fumigatus, Penicillium purpurescens, and P. verrucosum. Otherwise, to explain the antifungal activity, the present work paid considerable attention to the phytochemical characterization of the extract. Furthermore, we need further studies on the role of phenolic compounds in controlling plant viruses and fungi.
5. Conclusions
The findings from this study demonstrated that the Ulva fasciata organic extract possessed promising antiviral and antifungal properties. The HPLC analysis showed that 4-hydroxybenzoic acid was the highest phenolic compound, followed by ferulic acid, while 7-hydroxyflavone was the highest flavonoid in the organic extract of U. faciata, followed by rutin. At a concentration of 100 µg/mL, the U. fasciata extract was most effective at inhibiting TMV infection as well as B. cinerea growth by 81.25 and 69.26%, respectively. So, as far as we know, the results of this study may be the first to show that the seaweed U. fasciata can suppress plant viruses and fight fungi. The promising result in this work makes the tested green alga a promising source for producing antiviral and antifungal agents to control plant diseases.
Author Contributions
Conceptualization, E.H.E.-B., S.I.B. and A.A.; methodology, E.H.E.-B., A.-N.A.A.-M., A.A., I.I.T., A.E.-N.S. and S.I.B.; software, S.I.B. and A.A.; validation, M.M.E., A.E.-N.S. and E.H.E.-B.; formal analysis, A.A.H. and S.I.B.; investigation, A.A.; resources, E.H.E.-B.; data curation, M.M.E.; writing—original draft preparation, E.H.E.-B.; writing—review and editing, S.I.B. and A.A.; visualization, A.A.A.; supervision, A.A.A.-A.; project administration, F.O.A. and A.A.A.-A.; funding acquisition, F.O.A. and A.A.A.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This research project was supported by a grant from the Researchers Supporting Project number (RSP-2021/114), King Saud University, Riyadh, Saudi Arabia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to extend their appreciation to the Researchers Supporting Project number (RSP-2021/114), King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abdelkhalek, A.; Hafez, E. Plant Viral Diseases in Egypt and Their Control. In Cottage Industry of Biocontrol Agents and Their Applications; Springer: Berlin/Heidelberg, Germany, 2020; pp. 403–421. [Google Scholar]
- Abdelkhalek, A.; Behiry, S.I.; Al-Askar, A.A. Bacillus velezensis PEA1 Inhibits Fusarium oxysporum Growth and Induces Systemic Resistance to Cucumber Mosaic Virus. Agronomy 2020, 10, 1312. [Google Scholar] [CrossRef]
- Abdelkhalek, A. Expression of tomato pathogenesis related genes in response to Tobacco mosaic virus. JAPS J. Anim. Plant Sci. 2019, 29, 1596–1602. [Google Scholar]
- Scholthof, K.G.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalek, A.; Salem, M.Z.M.; Ali, H.M.; Kordy, A.M.; Salem, A.Z.M.; Behiry, S.I. Antiviral, antifungal, and insecticidal activities of Eucalyptus bark extract: HPLC analysis of polyphenolic compounds. Microb. Pathog. 2020, 147, 104383. [Google Scholar] [CrossRef] [PubMed]
- Heflish, A.A.; Abdelkhalek, A.; Al-Askar, A.A.; Behiry, S.I. Protective and Curative Effects of Trichoderma asperelloides Ta41 on Tomato Root Rot Caused by Rhizoctonia solani Rs33. Agronomy 2021, 11, 1162. [Google Scholar] [CrossRef]
- Oraby, S.A.M.; Radwan, R.I.M.; Ibrahim, S.A.; Hefny, R.M. Economic Study of Tomato Crop in Egypt WITH Referening to the Situation in North Sinai. Sinai J. Appl. Sci. 2017, 6, 223–234. [Google Scholar]
- Elsharkawy, M.M.; El-Okkiah, S.; Elsadany, A.Y.; Bedier, M.Y.; Omara, R.I.; Behiry, S.I.; Hassan, S.; Abdelkhalek, A. Systemic resistance induction of tomato plants against tomato mosaic virus by microalgae. Egypt. J. Biol. Pest Control 2022, 32, 1–7. [Google Scholar] [CrossRef]
- Mansour, M.M.A.; Salem, M.Z.M.; Khamis, M.H.; Ali, H.M. Natural durability of Citharexylum spinosum and Morus alba woods against three mold fungi. BioResources 2015, 10, 5330–5344. [Google Scholar] [CrossRef]
- Abd El-Rahim, W.M.; Khalil, W.K.B.; Eshak, M.G. Genotoxicity studies on the removal of a direct textile dye by a fungal strain, in vivo, using micronucleus and RAPD-PCR techniques on male rats. J. Appl. Toxicol. 2008, 28, 484–490. [Google Scholar] [CrossRef]
- Behiry, S.I.; Soliman, S.A.; Al-Askar, A.A.; Alotibi, F.O.; Basile, A.; Abdelkhalek, A.; Elsharkawy, M.M.; Salem, M.Z.M.; Hafez, E.E.; Heflish, A.A. Plantago lagopus extract as a green fungicide induces systemic resistance against Rhizoctonia root rot disease in tomato plants. Front. Plant Sci. 2022, 13, 2818. [Google Scholar] [CrossRef]
- Roux, F.; Voisin, D.; Badet, T.; Balagué, C.; Barlet, X.; Huard-Chauveau, C.; Roby, D.; Raffaele, S. Resistance to phytopathogens e tutti quanti: Placing plant quantitative disease resistance on the map. Mol. Plant Pathol. 2014, 15, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Rozlianah, F.S.; Sariah, M. Characterization of Malaysian isolates of Fusarium from tomato and pathogenicity testing. Res. J. Microbiol. 2010, 5, 926–932. [Google Scholar]
- Agamy, R.; Alamri, S.; Moustafa, M.F.M.; Hashem, M. Management of tomato leaf spot caused by Alternaria tenuissima Wiltshire using salicylic acid and Agrileen. Int. J. Agric. Biol. 2013, 15, 266–272. [Google Scholar]
- Sarven, M.S.; Hao, Q.; Deng, J.; Yang, F.; Wang, G.; Xiao, Y.; Xiao, X. Biological control of tomato gray mold caused by Botrytis cinerea with the entomopathogenic fungus Metarhizium anisopliae. Pathogens 2020, 9, 213. [Google Scholar] [CrossRef]
- Aslam, S.; Tahir, A.; Aslam, M.F.; Alam, M.W.; Shedayi, A.A.; Sadia, S. Recent advances in molecular techniques for the identification of phytopathogenic fungi—A mini review. J. Plant Interact. 2017, 12, 493–504. [Google Scholar] [CrossRef]
- Gazis, R.; Rehner, S.; Chaverri, P. Species delimitation in fungal endophyte diversity studies and its implications in ecological and biogeographic inferences. Mol. Ecol. 2011, 20, 3001–3013. [Google Scholar] [CrossRef]
- Chehri, K. Molecular identification of pathogenic Fusarium species, the causal agents of tomato wilt in western Iran. J. Plant Prot. Res. 2016, 56, 143–148. [Google Scholar] [CrossRef]
- Stracquadanio, C.; Luz, C.; La Spada, F.; Meca, G.; Cacciola, S.O. Inhibition of mycotoxigenic fungi in different vegetable matrices by extracts of Trichoderma species. J. Fungi 2021, 7, 445. [Google Scholar] [CrossRef]
- CHANG, Y.; Bin, D.U.; Ling, W.; Pei, J.I.; XIE, Y.; LI, X.; LI, Z.; WANG, J. A study on the pathogen species and physiological races of tomato Fusarium wilt in Shanxi, China. J. Integr. Agric. 2018, 17, 1380–1390. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A.A.; Elbeaino, T.; Moawad, H.; El-Gendi, H. Protective and Curative Activities of Paenibacillus polymyxa against Zucchini yellow mosaic virus Infestation in Squash Plants. Biology 2022, 11, 1150. [Google Scholar] [CrossRef]
- Baloch, G.N.; Tariq, S.; Ehteshamul-Haque, S.; Athar, M.; Sultana, V.; Ara, J. Management of root diseases of eggplant and watermelon with the application of asafoetida and seaweeds. J. Appl. Bot. Food Qual. 2013, 86, 138–142. [Google Scholar]
- Pourakbar, L.; Moghaddam, S.S.; Enshasy, H.A.; El Sayyed, R.Z. Antifungal activity of the extract of a macroalgae, Gracilariopsis persica, against four plant pathogenic fungi. Plants 2021, 10, 1781. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, Y.S.; Alamri, S.A.; Alrumman, S.A.; Hashem, M.; Taher, M.A.; Baka, Z.A. In Vitro and In Vivo Biocontrol of Tomato Fusarium Wilt by Extracts from Brown, Red, and Green Macroalgae. Agriculture 2022, 12, 345. [Google Scholar] [CrossRef]
- Khan, A.M.; Naz, S.; Abid, M. Evaluation of marine red alga Melanothamnus afaqhusainii against Meloidogyne incognita, fungus and as fertilizing potential on okra. Pak. J. Nematol 2016, 34, 91–100. [Google Scholar]
- Jiménez, E.; Dorta, F.; Medina, C.; Ramírez, A.; Ramírez, I.; Peña-Cortés, H. Anti-phytopathogenic activities of macro-algae extracts. Mar. Drugs 2011, 9, 739–756. [Google Scholar] [CrossRef]
- Umen, J.G. Green algae and the origins of multicellularity in the plant kingdom. Cold Spring Harb. Perspect. Biol. 2014, 6, a016170. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Hayden, H.S.; Blomster, J.; Maggs, C.A.; Silva, P.C.; Stanhope, M.J.; Waaland, J.R. Linnaeus was right all along: Ulva and Enteromorpha are not distinct genera. Eur. J. Phycol. 2003, 38, 277–294. [Google Scholar] [CrossRef]
- Koyanagi, S.; Tanigawa, N.; Nakagawa, H.; Soeda, S.; Shimeno, H. Oversulfation of fucoidan enhances its anti-angiogenic and antitumor activities. Biochem. Pharmacol. 2003, 65, 173–179. [Google Scholar] [CrossRef]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; de Sabry, D.A.; Câmara, R.B.G.; Nobre, L.T.D.B.; Costa, M.S.S.P.; Almeida-Lima, J.; Farias, E.H.C. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef]
- Al-Malki, A.L. In vitro cytotoxicity and pro-apoptotic activity of phycocyanin nanoparticles from Ulva lactuca (Chlorophyta) algae. Saudi J. Biol. Sci. 2020, 27, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Londo, G. The decimal scale for relevés of permanent quadrats. In Sampling Methods and Taxon Analysis in Vegetation Science: Releve Surveys,‘Vegetationsaufnahmen’, Floristic Analysis of Plant Communities; Junk: Pretoria, South Africa, 1984; pp. 45–49. [Google Scholar]
- Böer, B.; Warnken, J. Flora of the Jubail Marine Wildlife Sanctuary, Saudi Arabia. A Mar. Wildl. Sanctuary Arab. Gulf. Environ. Res. Conserv. Follow. 1991, 290–301. [Google Scholar]
- Komárek, J.; Zapomělová, E. Planktic morphospecies of the cyanobacterial genus Anabaena= subg. Dolichospermum–1. part: Coiled types. Fottea 2007, 7, 1–31. [Google Scholar] [CrossRef]
- Gooding, G.V., Jr.; Hebert, T.T. A simple technique for purification of tobacco mosaic virus in large quantities. Phytopathology 1967, 57, 1285. [Google Scholar]
- Abdelkhalek, A.; Ismail, I.A.I.A.; Dessoky, E.S.E.S.; El-Hallous, E.I.E.I.; Hafez, E. A tomato kinesin-like protein is associated with Tobacco mosaic virus infection. Biotechnol. Biotechnol. Equip. 2019, 33, 1424–1433. [Google Scholar] [CrossRef]
- Dhingra, O.D.; Sinclair, J.B. Basic Plant Pathology Methods; CRC Press: Boca Raton, FL, USA, 2017; ISBN 1315138131. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guid. Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Schimitz, H. Poisoned food technique Industrial and Engineering Chemistry Analyst. Ed 1930, 2, 361. [Google Scholar]
- Strange, R.N.; Scott, P.R. Plant disease: A threat to global food security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef]
- Omar, A.Z.; Hamdy, E.; Hamed, E.A.; Hafez, E.; Abdelkhalek, A. The curative activity of some arylidene dihydropyrimidine hydrazone against Tobacco mosaic virus infestation. J. Saudi Chem. Soc. 2022, 26, 101504. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A.A.; Behiry, S.I. Bacillus licheniformis strain POT1 mediated polyphenol biosynthetic pathways genes activation and systemic resistance in potato plants against Alfalfa mosaic virus. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.-Q. Heavy Metals and Pesticides Toxicity in Agricultural Soil and Plants: Ecological Risks and Human Health Implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- El-Gendi, H.; Al-Askar, A.A.; Király, L.; Samy, M.A.; Moawad, H.; Abdelkhalek, A. Foliar Applications of Bacillus subtilis HA1 Culture Filtrate Enhance Tomato Growth and Induce Systemic Resistance against Tobacco mosaic virus Infection. Horticulturae 2022, 8, 301. [Google Scholar] [CrossRef]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Gross, E.M.; Hilt, S.; Lombardo, P.; Mulderij, G. Searching for allelopathic effects of submerged macrophytes on phytoplankton—state of the art and open questions. In Shallow Lakes in a Changing World; Netherlands Institute of Ecology: Nieuwersluis, The Netherands, 2007; pp. 77–88. [Google Scholar]
- Mulderij, G.; Mooij, W.M.; Smolders, A.J.P.; Van Donk, E. Allelopathic inhibition of phytoplankton by exudates from Stratiotes aloides. Aquat. Bot. 2005, 82, 284–296. [Google Scholar] [CrossRef]
- Mofeed, J.; Deyab, M.; Mohamed, A.; Moustafa, M.; Negm, S.; El-Bilawy, E. Antimicrobial activities of three seaweeds extract against some human viral and bacterial pathogens. Biocell 2022, 46, 247. [Google Scholar] [CrossRef]
- Mishra, B.; Tiwari, A.; Mahmoud, A.E.D. Microalgal potential for sustainable aquaculture applications: Bioremediation, biocontrol, aquafeed. Clean Technol. Environ. Policy 2022, 1–13. [Google Scholar] [CrossRef]
- Abd El-Baky, H.H.; El-Baz, F.K.; El-Baroty, G.S. Natural preservative ingredient from marine alga Ulva lactuca L. Int. J. food Sci. Technol. 2009, 44, 1688–1695. [Google Scholar] [CrossRef]
- Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, C.J.; Telser, J. Role of oxygen radicals in DNA damage and cancer incidence. Mol. Cell. Biochem. 2004, 266, 37–56. [Google Scholar] [CrossRef]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef]
- Peasura, N.; Laohakunjit, N.; Kerdchoechuen, O.; Vongsawasdi, P.; Chao, L.K. Assessment of biochemical and immunomodulatory activity of sulphated polysaccharides from Ulva intestinalis. Int. J. Biol. Macromol. 2016, 91, 269–277. [Google Scholar] [CrossRef]
- Kidgell, J.T.; Magnusson, M.; de Nys, R.; Glasson, C.R.K. Ulvan: A systematic review of extraction, composition and function. Algal Res. 2019, 39, 101422. [Google Scholar] [CrossRef]
- Gunathilake, T.; Akanbi, T.O.; Suleria, H.A.R.; Nalder, T.D.; Francis, D.S.; Barrow, C.J. Seaweed phenolics as natural antioxidants, aquafeed additives, veterinary treatments and cross-linkers for microencapsulation. Mar. Drugs 2022, 20, 445. [Google Scholar] [CrossRef] [PubMed]
- Farasat, M.; Khavari-Nejad, R.-A.; Nabavi, S.M.B.; Namjooyan, F. Antioxidant activity, total phenolics and flavonoid contents of some edible green seaweeds from northern coasts of the Persian Gulf. Iran. J. Pharm. Res. 2014, 13, 163. [Google Scholar] [PubMed]
- Li, Z.; Wang, B.; Zhang, Q.; Qu, Y.; Xu, H.; Li, G. Preparation and antioxidant property of extract and semipurified fractions of Caulerpa racemosa. J. Appl. Phycol. 2012, 24, 1527–1536. [Google Scholar] [CrossRef]
- Elmosallamy, A.M.D.; Amer, T.N.; Mohamed, S.Z.; Ali, Y.M.; Hussein, S.A.A. Phytochemical Constituents of Ulva lactuca and Supplementation to Improve the Nile Tilapia (Oreochromis Niloticus) Haemato-Biochemical Status. Egypt. J. Chem. 2021, 64, 2663–2670. [Google Scholar] [CrossRef]
- Guardia, T.; Rotelli, A.E.; Juarez, A.O.; Pelzer, L.E. Anti-inflammatory properties of plant flavonoids. Effects of rutin, quercetin and hesperidin on adjuvant arthritis in rat. Il Farm. 2001, 56, 683–687. [Google Scholar] [CrossRef]
- Shahzad, G.-R.; Passera, A.; Maldera, G.; Casati, P.; Marcello, I.; Bianco, P.A. Biocontrol Potential of Endophytic Plant-Growth-Promoting Bacteria against Phytopathogenic Viruses: Molecular Interaction with the Host Plant and Comparison with Chitosan. Int. J. Mol. Sci. 2022, 23, 6990. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Aseel, D.G.; Király, L.; Künstler, A.; Moawad, H.; Al-Askar, A.A. Induction of Systemic Resistance to Tobacco mosaic virus in Tomato through Foliar Application of Bacillus amyloliquefaciens Strain TBorg1 Culture Filtrate. Viruses 2022, 14, 1830. [Google Scholar] [CrossRef] [PubMed]
- Waziri, H.M.A. Plants as antiviral agents. J. Plant Pathol. Microbiol. 2015, 6, 1. [Google Scholar] [CrossRef]
- Mani, J.S.; Johnson, J.B.; Steel, J.C.; Broszczak, D.A.; Neilsen, P.M.; Walsh, K.B.; Naiker, M. Natural product-derived phytochemicals as potential agents against coronaviruses: A review. Virus Res. 2020, 284, 197989. [Google Scholar] [CrossRef]
- Shaygannia, E.; Bahmani, M.; Zamanzad, B.; Rafieian-Kopaei, M. A review study on Punica granatum L. J. Evid. Based. Complementary Altern. Med. 2016, 21, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Davidson, P.M.; Taylor, T.M.; Schmidt, S.E. Chemical preservatives and natural antimicrobial compounds. In Food Microbiology: Fundamentals and Frontiers, 4th ed.; ASM Press: Washington, DC, USA, 2014; pp. 765–801. [Google Scholar]
- Kumar, N.; Pruthi, V. Potential applications of ferulic acid from natural sources. Biotechnol. Rep. 2014, 4, 86–93. [Google Scholar] [CrossRef]
- Likić, S.; Šola, I.; Ludwig-Müller, J.; Rusak, G. Involvement of kaempferol in the defence response of virus infected Arabidopsis thaliana. Eur. J. Plant Pathol. 2014, 138, 257–271. [Google Scholar] [CrossRef]
- Fan, W.; Qian, S.; Qian, P.; Li, X. Antiviral activity of luteolin against Japanese encephalitis virus. Virus Res. 2016, 220, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.Y.-L.; Stork, J.; Nagy, P.D. A key role for heat shock protein 70 in the localization and insertion of tombusvirus replication proteins to intracellular membranes. J. Virol. 2009, 83, 3276–3287. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hao, K.; Yu, F.; Shen, L.; Wang, F.; Yang, J.; Su, C. Field application of nanoliposomes delivered quercetin by inhibiting specific hsp70 gene expression against plant virus disease. J. Nanobiotechnol. 2022, 20, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mahfuz, S.; Mun, H.-S.; Dilawar, M.A.; Ampode, K.M.B.; Yang, C.-J. Potential Role of Protocatechuic Acid as Natural Feed Additives in Farm Animal Production. Animals 2022, 12, 741. [Google Scholar] [CrossRef]
- Li, T.; Huang, Y.; Xu, Z.-S.; Wang, F.; Xiong, A.-S. Salicylic acid-induced differential resistance to the Tomato yellow leaf curl virus among resistant and susceptible tomato cultivars. BMC Plant Biol. 2019, 19, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalek, A.; Al-Askar, A.A.; Alsubaie, M.M.; Behiry, S.I. First Report of Protective Activity of Paronychia argentea Extract against Tobacco Mosaic Virus Infection. Plants 2021, 10, 2435. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, M.W.; Muiru, W.M.; Kimenju, J.W. Characterisation of Fusarium species infecting tomato in Mwea West Sub-county, Kirinyaga County, Kenya. Can. J. Plant Pathol. 2021, 43, 56–61. [Google Scholar] [CrossRef]
- Akbar, A.; Hussain, S.; Ali, G.S. Germplasm evaluation of tomato for resistance to the emerging wilt pathogen Fusarium equiseti. J. Agric. Stud 2018, 5, 174. [Google Scholar] [CrossRef]
- Akbar, A.; Hussain, S.; Ullah, K.; Fahim, M.; Ali, G.S. Detection, virulence and genetic diversity of Fusarium species infecting tomato in Northern Pakistan. PLoS ONE 2018, 13, e0203613. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, A.; Kottb, M.; Elsayed, A.; Shafik, H. Antifungal activity of some Mediterranean seaweed against Macrophomina phaseolina and Fusarium oxysporum in Vitro. Alfarama J. Basic Appl. Sci. 2021, 2, 81–96. [Google Scholar] [CrossRef]
- Selim, R.E.; Ahmed, S.M.; El-Zemity, S.R.; Ramses, S.S.; Moustafa, Y.T.A. Antifungal activity and seasonal variation of green alga (Ulva lactuca) extracts. Asian J. Agric. Food Sci. 2015, 3, 419–427. [Google Scholar]
- Osman, M.E.H.; Abushady, A.M.; Elshobary, M.E. In vitro screening of antimicrobial activity of extracts of some macroalgae collected from Abu-Qir bay Alexandria, Egypt. African J. Biotechnol. 2010, 9, 7203–7208. [Google Scholar]
- Kosanić, M.; Ranković, B.; Stanojković, T. Biological activities of two macroalgae from Adriatic coast of Montenegro. Saudi J. Biol. Sci. 2015, 22, 390–397. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).