Abstract
Grazing is very common in the grassland ecosystem, and it has a significant impact on the C stocks and cycle. One of the most important drivers of soil C stocks is functional diversity. However, limited studies have attempted to explore the effects of functional diversity on soil C stocks associated with grazing disturbance. This study was carried out in Hulunbeier grassland, Inner Mongolia, and four grazing intensities (no grazing (NG), light grazing (LG), moderate grazing (MG), and heavy grazing (HG)) were identified. The plant functional traits and important soil properties under different grazing intensities were measured. Functional identity and diversity were calculated based on the measured functional traits. The impacts of functional identity and diversity on soil organic carbon stocks (SOCstocks) were analyzed using a multi-model inference (MMI) approach. Our study showed that the functional diversity effect on soil C stocks varies depending on grazing intensity. We identified that functional richness has a significant impact on SOCstocks in NG. The community weighted mean of leaf area became the best predictor of SOCstocks in LG. As grazing intensified, functional divergence best explained SOCstocks in moderate and heavy grazing sites, and their relationship was positive. The major outcomes of this research could shed light on the mechanics of soil carbon storage.
1. Introduction
Grassland ecosystems occupy approximately 40% of the Earth’s surface and are crucial to supplying livestock productivity. These areas store a considerable amount of carbon and play a vital role in the global carbon cycle [1,2,3,4]. Livestock grazing is a common land use in grassland ecosystems, which is not only the major livelihood of natives but essential for management for sustaining grassland health [3,5]. Since the 1980s, the demand for dairy products has surged with the progressive improvement in living conditions, further intensifying grassland utilization in Inner Mongolia, the primary ecological barrier of north China [5]. It has been reported that, in recent decades, long term overgrazing in grassland areas in Inner Mongolia has caused widespread grassland degradation and even desertification, threatening the region’s ecological security and socioeconomic development [6]. Achieving a better understanding of how grazing affects the key ecosystem properties is critical to maintaining grassland functionality and sustainability [7,8,9].
Soils are the largest terrestrial carbon (C) reservoirs, with about 3200 Pg of C stored as soil organic carbon (SOC) in the top three meters of the world’s soils [10,11]. Monitoring of SOC stocks is essential for sustainable soil management and meeting climate change mitigation targets [12,13,14]. In grassland ecosystems, soil C pools can be determined by plant biomass, the quantity and quality of plant litter, root mass turnover, soil microbial activities, and other environmental factors [1,15,16]. Among these factors, the collective traits of plant communities are key drivers of soil C stocks [17,18]. As functional trait based approaches develop, these functional characteristics of communities can be well depicted by functional diversity, identified as the value, range, distribution, and relative abundance of the functional traits of the organisms that make up an ecosystem [19]. Evidence suggests that examining trait based plant functional diversity can help to capture some ‘hidden’ mechanistic linking between biodiversity and ecosystem functioning, and predict ecosystem properties [20,21]. Functional diversity contains different components, which are related to distinct mechanisms of how diversity influences ecosystem functioning [18]. One of the important components of functional diversity is functional identity, which represents the trait values of the most abundant species and can be determined by the community weighted mean (CWM) trait. According to the mass ratio hypothesis, the contribution of different species to ecosystem functioning is proportional to their biomass. Thus, the trait of the most abundant species will be the main determining factor of ecosystem functioning [22]. For instance, critical functional plant traits of both above and below ground underpin the quantity and quality of litter input, affecting soil C stocks directly [18,23]. Previous studies have reported that functional traits associated with resource acquisitive strategies (e.g., high leaf nitrogen concentration and specific leaf area) are expected to produce more biomass to easily form mineralized litter, thus contributing to soil C accumulation [24,25]. On the other hand, the variety of functional trait values in the community is another important component of functional diversity and can be expressed by a series of indices, e.g., functional richness (FRic), functional evenness (FEve), and functional divergence (FDiv) [26]. These functional diversity indices reflect the niche complementarity hypothesis, suggesting that functionally different species traits, in terms of variations in resource-use strategies, would promote the efficient use of resources by species and enhance ecosystem functioning [27]. The coexistence of species with diverging functional traits could make full use of resources, further produce higher plant biomass, and trigger enhanced C storage through the higher input of litter [28].
A critical question here is which components of functional diversity are the most strongly associated with ecosystem properties. As mentioned in the prior studies, the mass ratio and niche complementarity hypotheses are not mutually exclusive, and different components of functional diversity could show distinct significances in diverse environments [29,30]. It has previously been observed that forest soil C inputs would be higher in the communities with resource acquisitive strategies functional traits in warmer or more humid conditions than that in drier conditions [31]. Most studies have focused on how the relationships between functional diversity and identity within ecosystem properties vary along natural environmental gradients in temperate, tropical, subtropical forests, and agroforests [23,25,32,33], but few scholars have attempted to explore these relationships with grazing disturbance, particularly in temperate grassland.
The temperate grassland in Inner Mongolia, a vital component of the Eurasian Grassland, is renowned for its extensive distribution, high aesthetic, and production values. A long history of grazing is still the main land-use regime in Inner Mongolia grassland [34]. It is now well established, from a variety of studies in the region, that livestock grazing is one of the most important disturbances that affect grassland, and research on the relationship between biodiversity and ecosystem properties mainly emphasizes species diversity but downplays the role of functional diversity [34,35]. The relative importance of functional diversity for explaining the underlying mechanism of the effects of grazing on soil C stock remains uncertain.
This research takes the Hulunbeier grassland in Inner Mongolia as the study site and aims to reveal the impact mechanisms of grazing on SOC stocks, using a functional trait based approach. The major objectives of this study are to (i) assess the responses of species diversity and different components of functional diversity to grazing intensity; (ii) assess the responses of soil abiotic properties and soil C stocks to grazing intensity; (iii) identify which components of plant functional diversity are vital for regulating soil C stocks under different grazing intensities. We expected that, by incorporating a functional trait based approach, the findings of this study would give insights into the mechanisms of grazing disturbance of soil C stocks, providing a scientific foundation for determining the optimal grassland grazing intensity and a valuable reference for improving grassland conservation and sustainable soil management.
2. Materials and Methods
2.1. Study Area
This study was performed in the Hulunbeier grassland of the Huihe National Nature Reserve (118°48′–119°43′ E, 48°10′–49°03′ N, 600–750 m a.s.l.), Inner Mongolia Autonomous Region, China (Figure 1). The area is located in the transition zone from the Greater Khingan Range forest to the Hulunbuir grassland and from meadow grassland to typical grassland, with a temperate continental monsoon climate. The mean annual temperature is −2–0 °C, and the mean annual precipitation ranges from 250 mm to 400 mm, 70% of which happens from June to August, concurrent with the growing season.
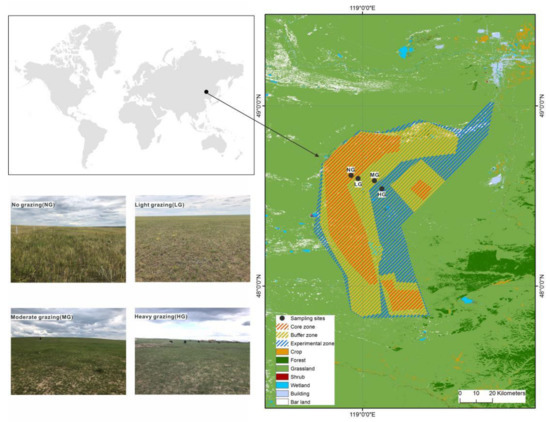
Figure 1.
Geographical location and field photos of the study sites in Hulunbeier grassland of Huihe National Nature Reserve, Inner Mongolia Autonomous Region, China.
The vegetation type of the study sites is meadow grassland, with the dominant species of Stipa baicalensis, Cleistogenes squarrosa, and Leymus chinensis. The soil is classified as dark chestnut soil according to the Chinese Soil Classification System [36]. Study sites have a long history of livestock grazing, which resulted in the degradation of grassland.
2.2. Experimental Design
A field experiment was performed in the Huihe National Nature Reserve during the growing season in July 2020 (Figure 1). National Nature Reserves in China are generally divided into three zones: core, buffer, and experimental zones. The core zones are strictly protected. Around them are the designated buffer zones, where nondestructive scientific investigation is permitted. The experimental zones are located outside the buffer zones, and anthropogenic activities are conducted there. Therefore, disturbance intensity increases with the distance from the core zones. We chose sites to match four grazing intensity categories with consistent topographic characteristics based on the distance to the core zones, vegetation, soil properties, and suggestions from local guides: no grazing (NG), light grazing (LG), moderate grazing (MG), and heavy grazing (HG). We set up NG sites in a 100-hectare fenced flat area that has been fenced to exclude grazing activities since 2009. LG sites were chosen in the buffer zone, where there are occasional cases of secret grazing. MG sites were selected from near experimental zones, and HG sites were chosen in experimental zones where grazing disturbance was more evident. Detailed information about the grazing intensities is shown in Table 1. Three plots of 10 m × 10 m with similar topographic characteristics were chosen randomly for each grazing intensity. The average distance between neighboring 10 m × 10 m plots was 50 m, and three 1 m × 1 m quadrats were randomly selected to survey vegetation and soil samples in each 10 m × 10 m plot.

Table 1.
Detailed information about four grazing intensities.
2.3. Plant and Soil Sampling
All living vascular plants from the quadrats were identified at the species level. Five important plant functional traits, including plant height, leaf area (LA), specific leaf area (SLA), leaf dry matter content (LDMC), and root length (RL), were measured. We chose these functional traits based on their ecological sensitivity to grazing, field operability, and functional linkages to plant growth, development, and resource acquisition [37,38]. Plant functional traits were measured on three individuals of each species with similar size in each 1-m2 quadrat. Measurements of plant functional traits were performed based on the standardized measurement procedure [39].
Soil cores from two soil layers (0–15 and 15–30 cm in depth) were collected using cutting rings (volume, 100 cm3) in each quadrat (three replicates, collected at least 50 cm apart). Soil samples were stored in a refrigerator at 4 °C until further analysis. Soil water content (SWC) was measured using the oven drying method. Soil pH was measured using a pH combination electrode at a soil–water ratio of 1:2.5. Soil total nitrogen (STN) was determined by the Kjeldahl method. Soil organic carbon (SOC) was measured using the Walkley and Black potassium dichromate oxidation method [40].
2.4. Data Analysis
We employed both Simpson’s diversity index and species richness to evaluate species diversity. The details of Simpson’s diversity index calculation are as follows:
where S is the number of species in a sampling unit and pi is the relative abundance of species i.
The community weighted mean (CWM) of each functional trait was calculated using the following formula:
where pi is the relative abundance of species in the community and traiti is the trait value of species i.
Functional diversity was expressed using three indices, which were calculated based on the 5 measured trait values together. Functional richness (FRic) represents the volume of multidimensional space occupied by all species in a community. Functional evenness (FEve) depicts the distribution and relative abundance of species across the functional space. Functional divergence (FDiv) quantifies the proportion of total abundance supported by species with the most extreme trait values within a community and reflects the degree of niche differentiation. These three indices together provide a comprehensive description of community functional diversity [41]. Calculations of CWM, FRic, FEve, and FDiv were conducted in R 4.0.2 using the ‘dbFD’ function within the FD package.
We performed a principal components analysis (PCA) using the CWMs of the five traits to refine the CWM traits into a few principal components to describe most of the total variance and represent a comprehensive functional identity in subsequent analyses.
We created a trait space in which species are ordered according to their trait values, and the distances between species reflect their trait similarity to illustrate the distribution characteristics of species in functional space. We used principal coordinates analysis (PCoA) to build functional spaces by ordinating the distance matrices in the software CANOCO 5 software.
For each soil sample, the SOC stocks were calculated as:
where i is the number of soil layers, Di is the soil depth (cm), Bi is the soil bulk density (g cm−3), and Ci is the C concentration (g kg−1) of soil layer i.
where SOCstocks(NG) and SOCstocks(LG, MG, HG) are the 0–30 cm soil organic carbon stocks of NG, LG, MG, and HG sites, formulated as (3).
PCA was applied to examine the relationships between CWM of functional traits, functional diversity indices, soil carbon stocks and soil properties. The analysis was conducted using the software CANOCO 5.
2.5. Statistical Analysis
We conducted a Shapiro–Wilk normality analysis to test the normal distribution of the data. The data was transformed to a normalized pattern if necessary. A multi-model inference (MMI) approach was used to examine the importance of each predictor and to obtain model averaged results. The MMI approach has been used in many areas of ecology and evolution, and we identified that it would be the best method to provide a more robust means of obtaining parameter estimates (both point and uncertainty estimates) by making inferences from a weighted combination of results from all the models in the set. The MMI approach effectively makes more stable predictions compared to traditional approaches, e.g., stepwise methods [32,42]. However, this method is limited when dealing with data sets with missing data, as deleting cases containing missing values can severely affect the results of model selection [42]. We built separate models for each grazing intensity using this method. In each case, the response variable is the total SOCstocks of 0–30 cm soil layer, and the predictors are functional identity and functional diversity indices. For the response variable, all possible models were fitted and ranked according to their AICC value (Akaike’s information criterion corrected by sample size). The relative importance of the predictors was calculated by summing Akaike’s weights of each candidate model. The analyses were performed using R 4.0.2 with “MuMIn” packages.
To determine the effects of grazing intensity on CWM of functional traits, functional diversity indices, SOCstocks, and soil properties, a one way analysis of variance (ANOVA) followed by Tukey’s HSD test was used to detect significant differences among grazing intensities.
3. Results
3.1. Responses of Taxonomic and Functional Indices
There were significant effects of grazing intensity on the taxonomic diversity index (Figure 2). Both species richness (p < 0.001) and the Simpson’s index (p < 0.001) were significantly lower in MG and HG than those in NG and LG (Figure 2a,b). Functional indices respond differently to grazing intensity. Grazing significantly affected FRic (p < 0.01) and FDiv (p < 0.001) (Figure 2c,e). Especially, FRic and FDiv were significantly lower in HG than that in NG. FEve showed no significant differences among grazing intensities (Figure 2d). The CWM of five plant functional traits (plant height, LA, SLA, LDMC, and RL) were examined in the study. The CWM of plant height (p < 0.001) and LDMC (p < 0.001) decreased significantly as grazing intensified (Figure 2f,i), while the CWM of SLA showed an opposite trend (p < 0.001), with the maximum in HD (Figure 2h). The CWM of LA (p < 0.01) and RL (p < 0.05) fluctuated among grazing intensities, with the minimum in MG and the maximum in LG and NG, respectively (Figure 2g,j).
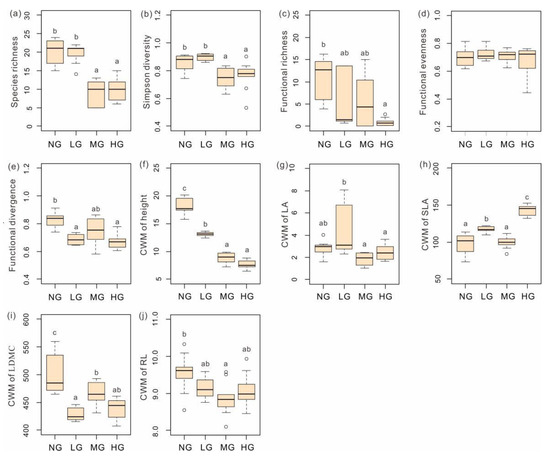
Figure 2.
(a) Species richness, (b) Simpson’s diversity, (c) functional richness, (d) functional evenness, (e) functional divergence, (f) the community weighted mean (CWM) of height, (g) CWM of leaf area (LA), (h) CWM of specific leaf area (SLA), (i) CWM of leaf dry matter content (LDMC), and (j) CWM of root length (RL) in NG (n = 9), LG (n = 9), MG (n = 9) and HG (n = 9) sites. For each variable, the middle horizontal line in the box plot represents the median, and the upper and lower short lines represent the maximum and minimum values. Alphabet indicates the significant differences among the grazing intensities (Tukey’s HSD test, p < 0.01).
A principal component analysis (PCA) was conducted to compress the variation of CWM traits into two principal components, which jointly represented 73.43% of the total variation (Table 2). The first principal component (CWM1) captured 48.14% of the total variation, exhibiting positive correlations with plant height, LDMC, and RL, and negative correlations with SLA. The second principal component (CWM2), which captured 25.29% of the total variation, mainly reflected the variation in the LA.
Table 2.
Loadings for axes1 and 2 according to principal component analysis (PCA) built using CWM of functional traits.
3.2. Responses of Soil Abiotic Properties and Soil Organic Carbon Stocks
Soil water content was affected by grazing intensity for both soil layers, with a significant decrease from MG to HG (p < 0.001). Grazing increased soil pH, but the change was significant only for the 0–15 cm soil layer (p < 0.001). SOC and soil TN showed similar trends; both of them decreased significantly from LG to MG (p < 0.001) and decreased significantly from the upper to the deeper soil layers. Soil C:N ratio did not show a significant difference among grazing intensities, with a slightly higher value in NG than in the other sites (Table 3).
Table 3.
Soil properties of different soil layers among the grazing intensities.
SOCstocks changed significantly as grazing intensified (p < 0.001), primarily exhibiting an acute decrease from LG to MG. Compared with NG, SOCstocks of the 0–30 cm soil layer decreased by 53.32% in HG (Figure 3).
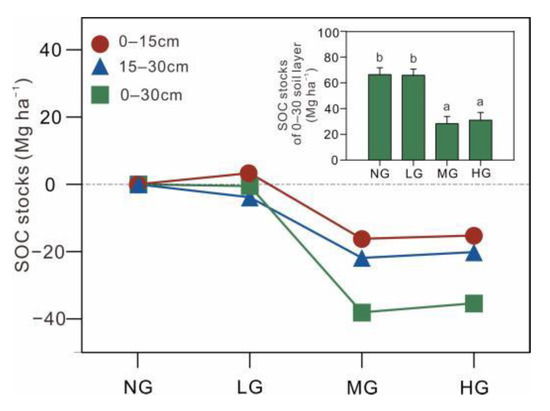
Figure 3.
The ∆ soil organic carbon stocks and soil organic carbon stocks (SOCstocks) of 0–30 cm soil layer under different grazing intensities. Alphabet indicates the significant difference among different grazing intensities (Tukey’s HSD test, p < 0.05).
3.3. Relationships between Functional Diversity Indices and Soil Organic Carbon Stocks
The result of PCA showed that differences in soil properties and functional diversity indices were pronounced among different grazing intensity sites (Figure 4). There was a positive correlation between FRic and TN and SOC concentration. FDiv showed a positive relationship with SWC. SLA was positively correlated with SBD and soil pH, and negatively correlated with soil nutrients, which indicates that heavy grazing supported species with high SLA traits. LDMC and RL showed a close relationship with SWC, and plant height was correlated with soil TN and SOC.
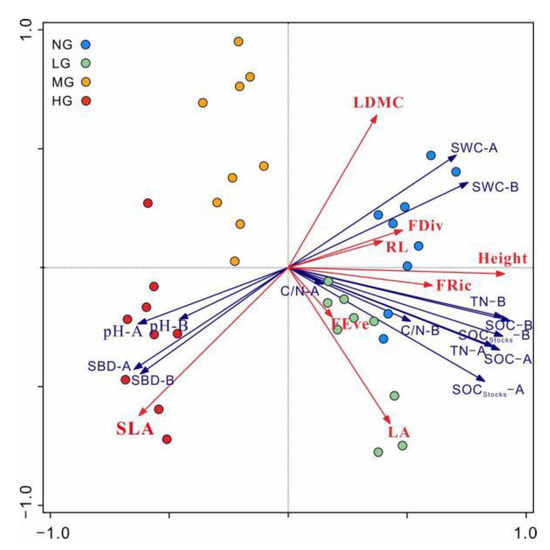
Figure 4.
Relationships between functional diversity indices and soil properties based on PCA. FRic, functional richness; FEve, functional evenness; FDiv, functional divergence; SLA, specific leaf area; LA, leaf area; LDMC, leaf dry matter content; RL, root length; SWC, soil water content; SOC, soil organic carbon concentration; TN, soil total nitrogen concentration; SOCstocks, soil organic carbon stocks; C/N, soil C to N ratio; SBD, soil bulk density; -A, 0–15 cm soil layer; -B, 15–30 cm soil layer.
Assessment of the effects of the functional diversity indices on SOCstocks by MMI suggested that the role of functional diversity indices in affecting SOCstocks were distinct under different grazing intensities (Table 4). In NG, there was a positive relationship between FRic and SOCstocks. The highest weight of FRic indicated that FRic was the major factor affecting SOCstocks in NG (Figure 5). For LG, the relative importance of CWM2 was 0.471, a larger value than the other indices, indicating that CWM2 was the main index related to SOCstocks. FDiv was of relatively higher importance than other indices in MG and HG and showed a positive relationship with SOCstocks. In addition, CWM1 also exhibited relative importance in affecting SOCstocks in MG and HG.
Table 4.
The two best predictive models of SOCstocks based on the multi-models inference approach, with model selection according to their corrected Akaike information criterion (AICc).
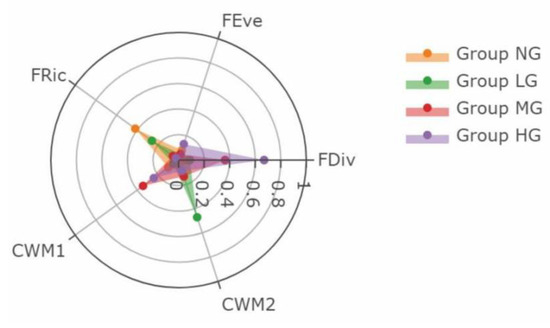
Figure 5.
The relative importance of the five variables (FRic, FEve, FDiv, CWM1, and CWM2), expressed by the sum of Akaike’s weights for all possible models. The importance values range between 0 and 1, where values closer to one indicate higher importance of the variable.
4. Discussion
4.1. Effects of Functional Diversity on Soil Carbon Stocks under Different Grazing Intensities
Our study examined grassland biodiversity and soil C stocks with different grazing intensities. The results revealed that severe grazing exerted a negative impact on species diversity, functional diversity, and soil C stocks in the Hulunbeier grassland. Our conclusion is in accordance with what has been suggested by other studies that showed global grasslands are significantly negatively impacted by grazing and that grassland under high grazing intensity suffer from carbon storage reduction, biodiversity loss, and key ecosystem properties damage [43,44].
Previous studies have shown that various ecosystem processes and functioning can be driven by plant functional diversity [45,46]. In our study, the trait based approach was used to explain the effect of functional diversity on soil C stocks. This study demonstrated that the relative importance of different components of functional diversity in affecting soil C stocks shifted among different grazing intensities. These results corroborate the findings of the previous work in tropical and temperate forests concerning land-use intensities [45,47].
In NG sites, FRic played a more important role in affecting soil C stocks than the other functional indices and showed the maximum value. FRic represents the functional space occupied by a community, including both the aboveground parts and the root systems [32]. Plant communities occupying more functional space could lead to fuller resource exploitation [48], which promotes plant growth and underpins the quality and quantity of litter input, and further has direct influences on SOC stocks. In addition, a related study carried out in China reported that plant communities with high root abundance occupied more underground functional space, which could enhance soil stabilization and reduce the erosion of soil C stocks [48]. Therefore, it can be concluded that a community with higher FRic can maintain SOC stocks better in grassland without grazing disturbance.
For light grazing sites (LG), the relative importance of FRic weakened, and CWM2 became the best predictor of SOC stocks, with a positive relationship. CWM describes traits of the dominant species in a community, following the “mass-ratio hypothesis”. According to the results of the PCA that extracted the main information of CWM traits into two PCA axes (CWM1 and CWM2), CWM2 was mainly positively correlated to LA. Several studies have found that LA plays an important role in leaf energy capture and water balance [49,50], and larger leaves require more dry mass investment per unit leaf area than smaller ones [51,52], which affects the amount of C in aboveground biomass and further influences C incorporated into the soil [53]. This means that grassland under light grazing had led to the local dominance of the larger LA plants associated with more C storage in soil.
As grazing intensified, FDiv best explained SOC stocks in MG and HG sites, and the relationship was positive. A higher value of FDiv suggested that a plant community has more abundant species with more extreme functional traits in a functional space [32,41]. On the one hand, the results supported the “niche complementarity hypothesis” that a plant community with diverging values of functional traits should lead to fuller resource exploitation, resulting in enhanced C stocks in plant biomass and soil. On the other hand, results suggested the role of some species with specific functional traits associated with C storage under heavy grazing. Based on PCoA results (Figure S1), we found that some species (e.g., Astragalus scaberrimus in HG) with more extreme functional traits showed high functional dissimilarity from other species assemblage in the functional space. A. scaberrimus are typical members of the leguminous family and have a unique capacity for nitrogen fixation, which could be associated with its contribution to SOC stocks. Similar results from previous studies demonstrated that species driven idiosyncratic effects are important in improving SOC stocks in most ecosystems [41,54].
CWM1 was also important for predicting SOC stocks in MG and HG sites, and the relationship was negative. CWM1 is positively correlated to plant height and LDMC, while negatively correlated with SLA. This implied that the dominance of high SLA and lower LDMC and height was positively associated with SOC stocks in MG and HG sites. Prior studies demonstrated that functional traits of high SLA and low LDMC were associated with high rates of resource acquisitive strategies, which could shed more biomass in the form of litter and promote carbon loss through leaf decomposition, thus contributing directly to the accumulation of soil C inputs [25,33,51]. Our results align with previous studies that demonstrated that underlined plant trait composition was an important driver of ecosystem functions. [33,53].
4.2. Implications for Sustainable Grassland Management Strategies
The management of grassland degradation caused by grazing disturbance is one of the most challenging problems faced today [55]. Understanding how grazing affects ecosystem functioning is critical to promoting sustainable development [45,46]. This research could be helpful to determine the mechanisms of soil C sequestration and help guide grassland conservation measurements.
Grazing disturbance should be controlled under a certain level of moderate grazing. As shown in our study, light grazing maintained a high level of species diversity, soil N content, and C stocks, while intensified grazing led to a significant decrease in functional and species diversity and soil nutrients and C stocks, indicating an increase in ecosystem fragility, and the ecosystem being less buffered against species loss in MG in the face of further disturbance.
Species diversity and species richness have commonly been used for monitoring ecosystem degradation. In the present study, fewer species changes with more functional diversity (e.g., FRic and FDiv) changed as the grazing intensified from NG to LG, implying that FD indicators could be more sensitive than species diversity indicators in reflecting the effect of disturbance. Given that multiple biodiversity indicators and ecosystem properties are becoming new important indicators reflecting the sustainability of ecosystems [45,56], we propose that integrating FD indices in monitoring programs can be an effective method to identify ecosystem sensitivity and prioritize management actions to avoid biodiversity loss and damage to ecosystem functioning caused by grazing disturbance.
The functional diversity effect on soil C stocks differed among different grazing intensities, indicating that protection mechanisms should be considered separately in future monitoring and modeling programs addressing functional diversity and identity effects on soil C stocks. However, due to the variation of plant species compositions and environmental characteristics, generalizing the findings of this study should be carried out with caution when comparing different ecosystems. Furthermore, this study evaluated data for only one year, and grassland ecosystem properties may vary with time. These changes cannot be fully understood due to the complexity of the ecosystem. Therefore, the long term monitoring of grassland under different grazing intensities is required for future research.
5. Conclusions
Our study clearly showed that species richness, the Simpson’s index, functional richness, and functional divergence under moderate or heavy grazing intensity were significantly lower than those under no grazing intensity. The CWM of plant height and the leaf dry matter content LDMC decreased, whereas the CWM of SLA increased as grazing intensified. SOC stocks under moderate or heavy grazing intensity were significantly lower than those under light or no grazing intensity. Our results pointed out that the functional diversity effect on soil C stocks differed among different grazing intensities. Under no grazing conditions, FRic played an important role in affecting SOC stocks. For light grazing sites, CWM2 (mostly represented by leaf area) became the best predictor of SOC stocks. As grazing intensified, FDiv mainly regulated SOC stocks under moderate and heavy grazing sites. This research could be helpful to determine the mechanisms of soil C sequestration and help guide grassland conservation measurements.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/su14084376/s1, Figure S1: The PCoA of the first two axes showing the distribution of the species in the functional space of NG, LG, MG and HG. Plant species names are indicated using the first three letters of both the genus and species name. Full names can be found in Table S1; Table S1: Full names of plant species names in Figure S1.
Author Contributions
Writing—original draft preparation, Z.S.; investigation, J.D. and B.S.; methodology and analysis, J.M. and Y.Z.; writing—review and editing, J.Z., Z.D. and Z.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by The National Key Research and Development Program of China (grant numbers 2020YFC1806305).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Acknowledgments
This study was supported by the National Environmental Protection Hulunber Forest-Steppe Ecotone Observation and Research Station. We thanked Qiang Wo and Zhijian Wu for their considerable assistance in the field investigation. Thanks are given to anonymous reviewers for their excellent suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tian, L.; Bai, Y.; Wang, W.; Qu, G.; Deng, Z.; Li, R.; Zhao, J. Warm- and cold- season grazing affect plant diversity and soil carbon and nitrogen sequestration differently in Tibetan alpine swamp meadows. Plant Soil 2020, 458, 151–164. [Google Scholar] [CrossRef]
- Mipam, T.D.; Chen, S.Y.; Liu, J.Q.; Miehe, G.; Tian, L.M. Short-term yak-grazing alters plant-soil stoichiometric relations in an alpine meadow on the eastern Tibetan Plateau. Plant Soil 2019, 458, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Talore, D.G.; Tesfamariam, E.H.; Hassen, A.; du Toit, J.C.; Klumpp, K.; Soussana, J.-F. Long-term impacts of season of grazing on soil carbon sequestration and selected soil properties in the arid Eastern Cape, South Africa. Plant Soil 2016, 397, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Mcsherry, M.; Ritchie, M.E. Effects of grazing on grassland soil carbon: A global review. Glob. Chang. Biol. 2013, 19, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Li, X.B.; Liu, S.Y.; Li, X.; Lyu, X.; Dang, D.L.; Dou, H.H. Ecosystem services under different grazing intensities in typical grasslands in Inner Mongolia and their relationships. Glob. Ecol. Conserv. 2021, 26, e01526. [Google Scholar] [CrossRef]
- Fan, F.; Liang, C.Z.; Tang, Y.K.; Harker-Schuch, I.; Porter, J.R. Effects and relationships of grazing intensity on multiple ecosystem services in the Inner Mongolian steppe. Sci. Total Environ. 2019, 675, 642–650. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Lauenroth, W.K. Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol. Monogr. 1993, 63, 327–366. [Google Scholar] [CrossRef]
- Abdalla, M.; Hastings, A.; Chadwick, D.R.; Jones, D.L.; Evans, C.D.; Jones, M.; Rees, R.; Smith, P. Critical review of the impacts of grazing intensity on soil organic carbon storage and other soil quality indicators in extensively managed grasslands. Agric. Ecosyst. Environ. 2018, 253, 62–81. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Q.; Epstein, H.; Li, G.Y. Grazing exclusion did not affect soil properties in alpine meadows in the Tibetan permafrost region. Ecol. Eng. 2019, 147, 105657. [Google Scholar] [CrossRef]
- Schiedung, M.; Tregurtha, C.S.; Beare, M.H.; Thomas, S.M.; Don, A. Deep soil flipping increases carbon stocks of New Zealand grasslands. Glob. Chang. Biol. 2019, 25, 2296–2309. [Google Scholar] [CrossRef] [Green Version]
- Batjes, N.H. Harmonized soil property values for broad-scale modelling (WISE30sec) with estimates of global soil carbon stocks. Geoderma 2016, 269, 61–68. [Google Scholar] [CrossRef]
- Palandrani, C.; Alberti, G. Tree derived soil carbon is enhanced by tree species richness and functional diversity. Plant Soil 2020, 446, 457–469. [Google Scholar] [CrossRef]
- Cubasch, U.; Wuebbles, D.; Chen, D.; Facchini, M.C.; Frame, D.; Mahowald, N.; Winther, J. Introduction. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Eds.; Cambridge University Press: Cambridge, UK, 2013; pp. 121–157. [Google Scholar]
- Chabala, L.M.; Angombe, S.; Amelung, W.; Lark, R.M. The effect of water deficit and livestock stocking density on soil organic carbon stocks in Namibia. Geoderma 2022, 407, 115522. [Google Scholar] [CrossRef]
- Hassan, N.; Li, X.F.; Wang, J.Y.; Zhu, H.; Nummi, P.; Wang, D.; Finke, D.; Zhong, Z. Effects of grazing on C:N:P stoichiometry attenuate from soils to plants and insect herbivores in a semi-arid grassland. Oecologia 2021, 195, 785–795. [Google Scholar] [CrossRef]
- Wang, L.; Delgado-Baquerizo, M.; Wang, D.L.; Isbell, F.; Liu, J.; Feng, C.; Liu, J.; Zhong, Z.; Zhu, H.; Yuan, X.; et al. Diversifying livestock promotes multidiversity and multifunctionality in managed grasslands. Proc. Natl. Acad. Sci. USA 2019, 116, 6187–6192. [Google Scholar] [CrossRef] [Green Version]
- Catovsky, S.; Bradford, M.A.; Hector, A. Biodiversity and ecosystem productivity: Implications for carbon storage. Oikos 2002, 97, 443–448. [Google Scholar] [CrossRef]
- Conti, G.; Díaz, S. Plant functional diversity and carbon storage–an empirical test in semi-arid forest ecosystems. J. Ecol. 2013, 101, 18–28. [Google Scholar] [CrossRef]
- Díaz, S.; Lavorel, S.; de Bello, F.; Quetier, F.; Grigulis, K.; Robson, T.M. Incorporating plant functional diversity effects in ecosystem service assessments. Proc. Natl. Acad. Sci. USA 2007, 104, 20684–20689. [Google Scholar] [CrossRef] [Green Version]
- Funk, J.L.; Larson, J.E.; Ames, G.M.; Butterfield, B.J.; Cavender-Bares, J.; Firn, J.; Laughlin, D.C.; Sutton-Grier, A.E.; Williams, L.; Wright, J. Revisiting the Holy Grail: Using plant functional traits to understand ecological processes. Biol. Rev. 2017, 92, 1156–1173. [Google Scholar] [CrossRef]
- McLean, M.; Auber, A.; Graham, N.A.J.; Houk, P.; Villeger, S.; Violle, C.; Thuiller, W.; Wilson, S.K.; Mouillot, D. Trait structure and redundancy determine sensitivity to disturbance in marine fish communities. Glob. Chang. Biol. 2019, 25, 3424–3437. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Buchanan, S.W.; Mafa-Attoye, T.; Dunfield, K.; Thevathasan, N.V.; Isaac, M.E. The role of plant functional traits and diversity in soil carbon dynamics within riparian agroforests. J. Environ. Qual. 2021, 51, 33–43. [Google Scholar] [CrossRef]
- Faucon, M.P.; Houben, D.; Lambers, H. Plant functional traits: Soil and ecosystem services. Trends Plant Sci. 2017, 22, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Hisano, M.; Taylor, A.R.; Chen, H.Y.H. The effects of functional diversity and identity (acquisitive versus conservative strategies) on soil carbon stocks are dependent on environmental contexts. For. Ecol. Manag. 2022, 503, 119820. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Villeger, S.; Mason, N.W.H.; Mouillot, D. Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Tilman, D.; Knops, J.; Wedin, D.; Reich, P.; Ritchie, M.; Siemann, E. The influence of functional diversity and composition on ecosystem processes. Science 1997, 277, 1300–1302. [Google Scholar] [CrossRef] [Green Version]
- Scherer-Lorenzen, M.; Bonilla, J.L.; Potvin, C. Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment. Oikos 2007, 116, 2108–2124. [Google Scholar] [CrossRef]
- Mensah, S.; Veldtman, R.; Assogbadjo, A.E.; Kakai, R.G.; Seifert, T. Tree species diversity promotes aboveground carbon storage through functional diversity and functional dominance. Ecol. Evol. 2016, 6, 7546–7557. [Google Scholar] [CrossRef]
- Zirbel, C.R.; Bassett, T.; Grman, E.; Brudvig, L.A. Plant functional traits and environmental conditions shape community assembly and ecosystem functioning during restoration. J. Appl. Ecol. 2017, 54, 1070–1079. [Google Scholar] [CrossRef] [Green Version]
- De Deyn, G.B.; Cornelissen, J.H.C.; Bardgett, R.D. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol. Lett. 2008, 11, 516–531. [Google Scholar] [CrossRef]
- Hou, J.; Zhu, H.X.; Fu, B.J.; Lu, Y.H.; Zhou, J. Functional traits explain seasonal variation effects of plant communities on soil erosion in semiarid grasslands in the Loess Plateau of China. Catena 2020, 194, 104743. [Google Scholar] [CrossRef]
- Asanok, L.; Taweesuk, R.; Kamyo, T. Plant functional diversity is linked to carbon storage in deciduous dipterocarp forest edges in Northern Thailand. Sustainability 2021, 13, 11416. [Google Scholar] [CrossRef]
- Yang, Z.P.; Baoyin, T.; Minggagud, H.; Sun, H.P.; Li, F.Y. Recovery succession drives the convergence, and grazing versus fencing drives the divergence of plant and soil N/P stoichiometry in a semiarid steppe of Inner Mongolia. Plant Soil 2017, 420, 303–314. [Google Scholar] [CrossRef]
- Sasaki, T.; Okubo, S.; Okayasu, T.; Jamsran, U.; Ohkuro, T.; Takeuchi, K. Two–phase functional redundancy in plant communities along a grazing gradient in Mongolian rangelands. Ecology 2009, 90, 2598–2608. [Google Scholar] [CrossRef] [Green Version]
- Soil Taxonomy Research Group of Nanjing Institue of Soil Sciences. Chinese Soil Taxonomy Retrieval, 3rd ed.; University of Science and Technology of China Press: Hefei, China, 2001. [Google Scholar]
- Garnier, E.; Navas, M.L. A trait-based approach to comparative functional plant ecology: Concepts, methods and applications for agroecology. A review. Agron. Sustain. Dev. 2012, 32, 365–399. [Google Scholar] [CrossRef] [Green Version]
- Wesuls, D.; Oldeland, J.; Dray, S. Disentangling plant trait responses to livestock grazing from spatio-temporal variation: The partial RLQ approach. J. Veg. Sci. 2012, 23, 98–113. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; Page, A.L., Ed.; American Society of Agronomy: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Mouillot, D.; Graham, N.A.J.; Villeger, S.; Mason, N.W.H.; Bellwood, D.R. A functional approach reveals community responses to disturbances. Trends Ecol. Evol. 2013, 28, 167–177. [Google Scholar] [CrossRef]
- Grueber, C.E.; Nakagawa, S.; Laws, R.J.; Jamieson, I.G. Multimodel inference in ecology and evolution: Challenges and solutions. J. Evol. Biol. 2013, 24, 699–711. [Google Scholar] [CrossRef]
- Ren, H.; Eviner, V.T.; Gui, W.; Wilson, G.W.T.; Cobb, A.B.; Yang, G.; Zhang, Y.; Hu, S.; Bai, Y. Livestock grazing regulates ecosystem multifunctionality in semi-arid grassland. Funct. Ecol. 2018, 32, 2790–2800. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.Q.; He, Z.W. Effects of grazing patterns on grassland biomass and soil environments in China: A meta-analysis. PLoS ONE 2019, 14, e0215223. [Google Scholar] [CrossRef]
- Chillo, V.; Vazquez, D.P.; Amoroso, M.M.; Bennett, E.M. Land-use intensity indirectly affects ecosystem services mainly through plant functional identity in a temperate forest. Funct. Ecol. 2018, 32, 1390–1399. [Google Scholar] [CrossRef] [Green Version]
- Dovrat, G.; Meron, E.; Shachak, M.; Golodets, C.; Osem, Y. The relative contributions of functional diversity and functional identity to ecosystem function in water-limited environments. J. Veg. Sci. 2019, 30, 427–437. [Google Scholar] [CrossRef]
- Pereira, K.M.G.; Cordeiro, N.G.; Terra, M.C.N.S.; Pyles, M.; Cabacinha, C.D.; De Mello, J.M.; Berg, E.V.D. Protection status as determinant of carbon stock drivers in cerrado sensu stricto. J. Plant. Ecol. 2020, 13, 361–368. [Google Scholar] [CrossRef]
- Ghestem, M.; Cao, K.F.; Ma, W.Z.; Rowe, N.P.; Leclerc, R.; Gadenne, C.; Stokes, A. A framework for identifying plant species to be used as ‘ecological engineers’ for fixing soil on unstable slopes. PLoS ONE 2014, 9, e95876. [Google Scholar] [CrossRef]
- Hou, Z.F.; Lv, G.H.; Jiang, L.M. Functional diversity can predict ecosystem functions better than dominant species: The case of desert plants in the Ebinur Lake Basin. Sustainability 2021, 13, 2858. [Google Scholar] [CrossRef]
- Milla, R.; Reich, P.B. The scaling of leaf area and mass: The cost of light interception increases with leaf size. Proc. R. Soc. B 2007, 274, 2109–2114. [Google Scholar] [CrossRef] [Green Version]
- Shipley, B. Structured interspecific determinants of specific leaf area in 34 species of herbaceous. Funct. Ecol. 1995, 9, 312–319. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Portsmuth, A.; Tobias, M. Leaf size modifies support biomass distribution among stems, petioles and mid-ribs in temperate plants. New Phytol. 2006, 171, 91–104. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K. How fundamental plant functional trait relationships scale-up to trade-offs and synergies in ecosystem services. J. Ecol. 2012, 100, 128–140. [Google Scholar] [CrossRef]
- Potvin, C.; Mancilla, L.; Buchmann, N.; Monteza, J.; Moore, T.; Murphy, M.; Oelmann, Y.; Scherer-Lorenzen, M.; Turner, B.; Wilcke, W.; et al. An ecosystem approach to biodiversity effects: Carbon pools in a tropical tree plantation. For. Ecol. Manag. 2011, 261, 1614–1624. [Google Scholar] [CrossRef]
- Wang, S.Z.; Fan, J.W.; Li, Y.Z.; Huang, L. Effects of grazing exclusion on biomass growth and species diversity among various grassland types of the Tibetan Plateau. Sustainability 2019, 11, 1705. [Google Scholar] [CrossRef] [Green Version]
- Allan, E.; Manning, P.; Alt, F.; Binkenstein, J.; Blaser, S.; Blüthgen, N.; Böhm, S.; Grassein, F.; Hölzel, N.; Klaus, V.H.; et al. Land use intensification alters ecosystem multifunctionality via loss of biodiversity and changes to functional composition. Ecol. Lett. 2015, 18, 834–843. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).