1. Introduction
Climate change is an important issue for the small island states of the Caribbean due to an inherent climate sensitivity attributable to small size, location in the Atlantic hurricane belt, and other factors [
1]. The Caribbean’s climate is already changing, as seen by rising temperatures, increasingly variable rainfall, increasing drought and flood frequency, rising sea levels, and intensifying storms [
1,
2,
3,
4]. Modelling studies project that, by the end of the current century, the region will experience an increase in intensity of rainfall events; a decrease in total annual rainfall of up to 40% in some places and particularly during the climatologically wettest period of May to October; warming of up to 4 °C, with night-time temperatures increasing at a faster rate than daytime; more intense hurricanes; more frequent floods and droughts; and sea level rise of up to 1 m [
2,
5,
6,
7,
8,
9]. Being a tropical region, the Caribbean is also projected to experience a climate departure characterized by extreme climate shifts far sooner than other regions [
10]. For example, by the mid-2020s it is projected that annual minimum temperatures in some years may be higher than historical maximum temperatures [
10].
Although over 9% of neotropical dry forests occur in the insular Caribbean, approximately 66% of that dry forest cover has been lost and much of what remains is at least somewhat fragmented or degraded [
11]. Tropical dry forests (TDFs) are highly sensitive to shifts in climate, but have been the focus of very few studies on the ecological impacts of climate change [
12]. In these ecosystems, rainfall has a strong influence on seedling survival, species composition, and above-ground biomass, which can be significantly changed or impaired by drought occurrence [
13,
14,
15,
16,
17]. Hurricanes also significantly impact survival of mature vegetation and species composition of mature TDFs. In 2015, Hurricane Patricia made landfall as a category 4 cyclone on the Pacific coast of Mexico. In assessing the damage to the Chamela-Cuixmala TDF, Jimenez-Rodríguez et al. (2018) identified that more severe damage occurred in old growth areas than in secondary forest, and the forest composition changed to favor smaller plants that could regenerate quickly [
18]. Therefore, climate change will likely result in a shift in the forest composition, ecosystem dynamics, and extinction risk of many species in TDFs, which are already the most threatened ecosystems in the world [
12,
19]. Characterizing the impacts of climate change on dry forests and other biodiverse Caribbean ecosystems is a necessary first step for planning and promoting actions aimed at the conservation of the region’s biodiversity, particularly in the minimally studied dry forests of the region [
12,
19,
20].
Despite being a hotspot for biodiversity, the impact of climate change on Caribbean ecosystems has not been extensively studied. This may be due to several reasons including (i) the low importance placed on biodiversity, (ii) data limitations, (iii) the lack of pristine study sites, and (iv) the lack of suitable/well developed methodologies [
21]. Few studies have focused on the region’s terrestrial faunal species [
20]. Bioclimatic modelling, in particular, has been applied only minimally in the insular Caribbean, as the small sizes of the islands limit their accurate representation within many of the existing models [
20]. Additionally, only limited work has previously examined vulnerability to climatic influence (in particular rainfall and temperature) as well as survival potential and extinction risk in important tropical dry forest habitats such as the Hellshire Hills, Jamaica or forests like it. These include studies examining (i) rainfall as a predictor of
Anolis lineatopus hatchling numbers and juvenile growth [
22]; (ii) the link between lizard extinction and temperature [
23]; and (iii) seedling dynamics and climate variability [
24].
Previous studies have shown that climate is a strong modulator of ectotherm populations, which further emphasizes the need to characterize potential impacts of climate change in these sensitive hotspots. Rainfall and relative humidity are important factors in the life cycles of arthropods and small lizards. Low moisture availability can retard the life cycle of the organism by impairing oviposition, juvenile recruitment, growth and development, and physical function, while high relative humidity can increase pathogen infection [
25,
26,
27]. Studies assessing the variability of arthropod populations as a function of rainfall seasonality in Grenada in 1977 showed that arthropod populations were 2.3 times larger and 3.1 times greater in biomass during the wet season (June–December) than during the dry season (January–May) [
28]. The influence of rainfall on arthropod abundance also impacts food availability for lizards, which has been found to be a limitation of juvenile growth in Jamaican
Anolis lineatopus [
22]. Moisture is also critical to anoles for egg laying and development, and has been identified as a determining factor in the geographical variation in morphology of
Anolis lineatopus and
A. grahami [
22,
25,
29,
30]. These factors indicate that rainfall is linked to anole abundance both directly through provision of water and indirectly through its modulation of prey availability. Water availability plays a large part in microhabitat choice in invertebrates and lizards, which tend to occupy shaded areas during dry periods in order to minimize water loss and thermal stress [
31,
32,
33].
Tropical arthropods and lizards also exhibit high thermal sensitivity that is not minimized by their tolerance of warm habitats. Warmer temperatures alter the rate of development for many arthropods, which can result in fluctuating population size due to changes in breeding patterns, survival, and food availability [
27,
34,
35]. There is, however, a temperature threshold for growth and development, which may be different for adults and nymphs of the same species [
36]. Experiments conducted on
Nilaparvata lugens, one of the most potent rice pests globally, showed that adults consistently had higher critical thermal maximum (Ctmax), upper lethal temperature (ULT), and heat coma temperature (HCT) than nymphs [
37]. Given recent global temperature trends, particularly in the tropics where the species is most prevalent, nymphs are currently very close to exceeding their optimal temperatures [
37]. For tropical lizards, studies of thermal vulnerability have found that living in a warm habitat does not remove the possibility of intolerance to high temperatures, given their sensitivity to extreme heating due to their thermoconforming nature [
30,
38]. Thermal responses are more dependent on general behavior of the species, as basking species living in open habitats tend to maintain high body and operative temperatures relative to shaded habitat non-basking species, which are often active at lower temperatures that are also closer to ambient temperatures [
30,
38]. Also, the relatively narrow annual temperature range to which tropical groups are accustomed adds to their vulnerability to overheating and reduced activity under substantive rises in ambient temperature outside of their preferred temperature ranges, particularly during summer months [
30,
38].
Historical relationships between ectotherms and climate show that climate change will lead to evolution in physiology, shifts in climate niches, migration, range expansion or contraction, and alteration of trophic interactions and predator-prey relationships [
30,
34,
39,
40,
41,
42]. Some tropical species do, however, have natural physiological responses that confer adaptive capacity, such as the heat-hardening capacity in lizards that allows for survival in extreme temperatures [
40,
43]. It must be noted, however, that heat hardening varies geographically and does not negate negative impacts of extreme heat on locomotor function [
43].
The overall aim of this study is to investigate the potential role of climate in determining faunal abundance in a small dry forest located in Hellshire Hills, Jamaica. We characterize the climate of the Hellshire Hills dry forest, and investigate whether there are discernible links among climate, arthropod, and lizard abundance, and whether the relationships found can be used to develop a simple bioclimatic model representative of the region. The validated simple model is then used to infer information about the impact of projected future changes in the climate of the Caribbean on the biodiversity of the region. Given the dearth of studies examining climatic impacts on regional fauna, this will be a useful addition to regional conservation efforts.
3. Results
3.1. Climate of the Hellshire Hills
The Hellshire Hills climatology was determined using data obtained from the weather station installed at the site (
Figure 2). The climatology suggests a pattern of mean temperatures that are coolest in the northern hemisphere winter months (minimum in December) and warmest in the summer months (peak in August). The Hellshire Hills exhibit an overall warmer climate than both Norman Manley and the Jamaican average, with wider variation between maximum and minimum temperatures throughout the year. The region is generally hot all year round, with comparatively little monthly variation (
Figure 2A). The annual ranges for mean, maximum and minimum temperatures for the Hellshire Hills region were only 3.3, 3.4, and 3.2 °C respectively. Daily temperature range was typically smaller during the warmer summer months than it was during colder periods. The available data also indicated that the maximum mean daytime temperature for the Hellshire Hills varied between 31 and 33 °C, and was consistently high throughout the year. In comparison, minimum night-time temperature varied from a low of 21 °C in December-March to as high as 25 °C in June, while mean temperatures varied between 24 and 28 °C, with a peak in August. The diurnal temperature variation appeared to be greater for Hellshire Hills than Norman Manley for the same period, with generally cooler nights and warmer days. A maximum summer daytime temperature of 39 °C was recorded in the Hellshire Hills on 31 July 2012, and a minimum winter night-time temperature of 17 °C was recorded on 15 December 2010. In the summer of 2011, 13 days between June and August exceeded 35 °C. This rose to 25 days in the summer of 2012, indicating an increase in the number of hot days.
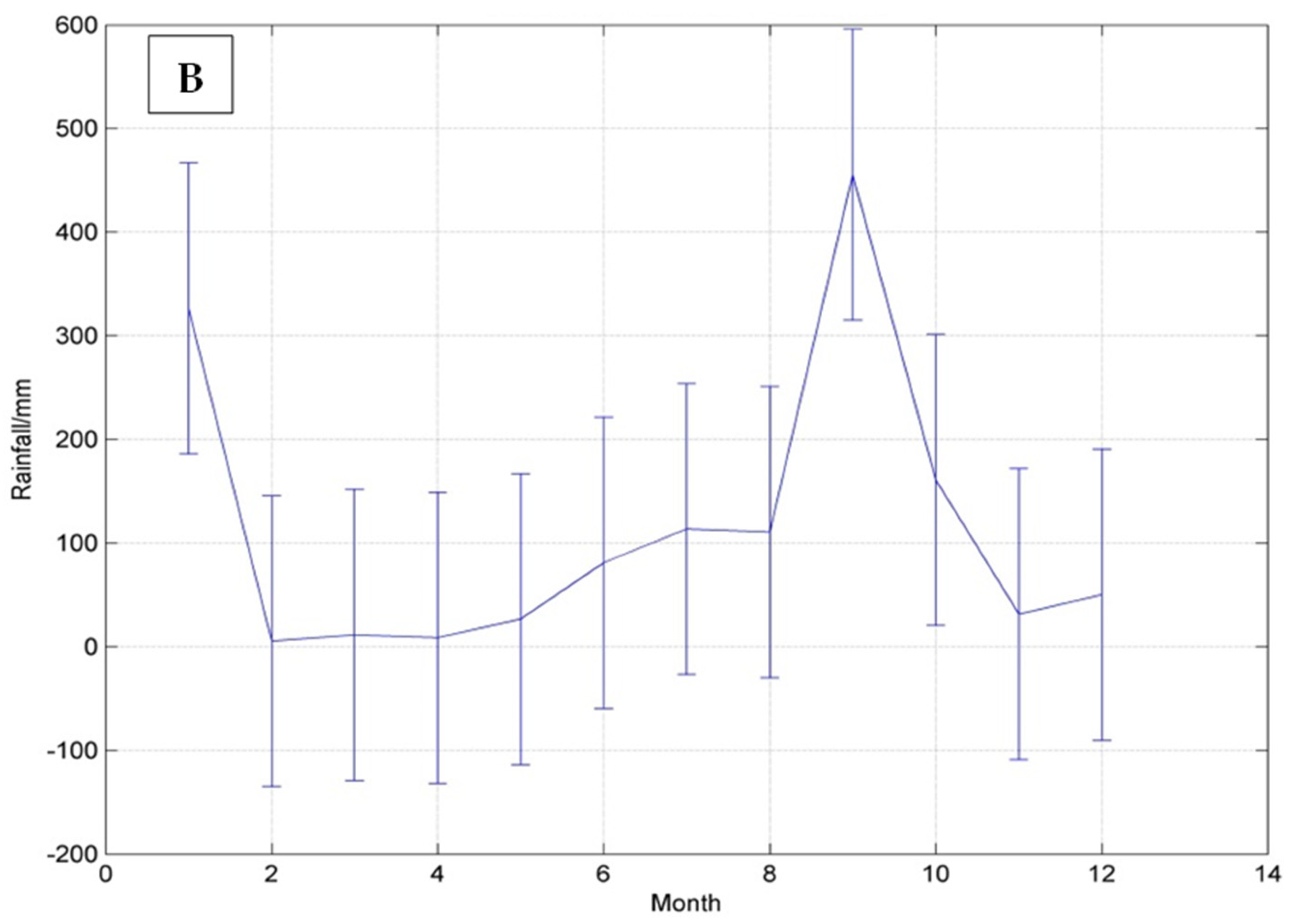
The site is by comparison very dry when compared with rainfall received at Norman Manley or the island average. Year by year analysis of the HH station data suggested a general mirroring of Jamaica’s bimodal rainfall pattern, i.e., with a rainy season between May and November interrupted by a short dry spell in August. However, the peak in May was not distinct in all three years of data, resulting in the derived climatology displaying a more unimodal distribution over the rainfall season (
Figure 2B). The early months (February to March) were also particularly dry, with almost no rainfall registered during the period (
Figure 2B). Typically, January–February represents the peak of the Hellshire Hills dry season.
Figure 2B also shows that nine months of the year received approximately 100 mm or less of rainfall. However, we note the influence of the drought of 2010, which may have skewed the data, and the influence of Tropical Storm Nicole and Hurricane Sandy in 2010 and 2012, respectively. Notwithstanding, the general pattern that emerges is of a region with very little rainfall except for a few months of the year.
Projections from the PRECIS RCM for the grid box over the Hellshire Hills indicate that the region is projected to warm by between 2.8 °C (B2) and 4.3 °C (A2) (
Table 2). Warming is most pronounced during the summer months (June–July) under both scenarios, reaching as high as 5.2 °C under the A2 scenario and 3.0 °C under the B2 scenario. The traditionally cooler months between November and February show smaller magnitude warming (2–3 °C). Model projections also suggest an increase in the frequency of very warm days and a corresponding decrease in the frequency of very cool days irrespective of scenario. Projected percentage changes in rainfall indicate between 23% and 40% less rainfall in the annual mean occurring in the Hellshire Hills under the B2 and A2 scenarios (respectively) by the end of the century. That is, an already significantly dry region will become even drier. Drying is most significant in the summer months under both scenarios.
In general, present-day seasonality will still be retained by the end of the century, i.e., the summer months will still be the warmest and wettest periods in the far future, while February through March will remain the driest and coolest months. However, there will be an increase in annual temperature range of 2.5–3 °C along with the possibility that under the more severe emissions scenario rain in January may be comparable to peak rain received in September due to the drastic reduction in the latter month’s totals. In tandem the temperature and rainfall projections present an end of century profile which suggests that the Hellshire Hills will become even warmer and drier. Importantly, the changes projected are larger than the range of variability currently seen, as revealed by the trends in the 17-year present-day dataset. This suggests ‘unprecedented’ changes to the climatic envelope of the Hellshire Hills.
3.2. Variation in Abundance and Climate Linkages
A significant linear trend in abundance was not identified for any taxon over the 17-year period under investigation. Notwithstanding, for 14 of the 22 species, the trend in abundance over time is slightly negative. The taxa with the largest magnitude slope (whether positive or negative) were stink bugs (
Pangaeus bilineatus +6.19) and beetles (
Branchus jamaicensis −5.05, Chrysomelidae −75.01). This is consistent with previously identified significant negative slopes for temperature variables and positive slopes for rainfall [
71]. Interannual variability is also evident in almost all the species. This is very reminiscent of the rainfall trends (annual and seasonal) for which periodicity analysis revealed a dominant three-year signal that may also be linked to the variability evident in annual abundance. This suggests that rainfall is a significant influencing factor [
71].
The following observations are therefore noted from abundance data in both control and experimental plots: (i) there is no significant linear trend, (ii) there is strong interannual variability, and (iii) there are common linkages with rainfall across numerous taxa, in particular for lizards.
All trap areas showed similar trends in overall captures throughout the study period, and experimental plots had consistently higher capture rates than control plots. Correlations were generally high and significant (i) between control grids; (ii) between experimental grids, and (iii) between the total captures for the control and experimental plots combined. This suggests that, in general, the factors (e.g., climate) that are driving variability of the trapping at one control or experimental plot are common to the other control or experimental plot.
3.3. Model Creation
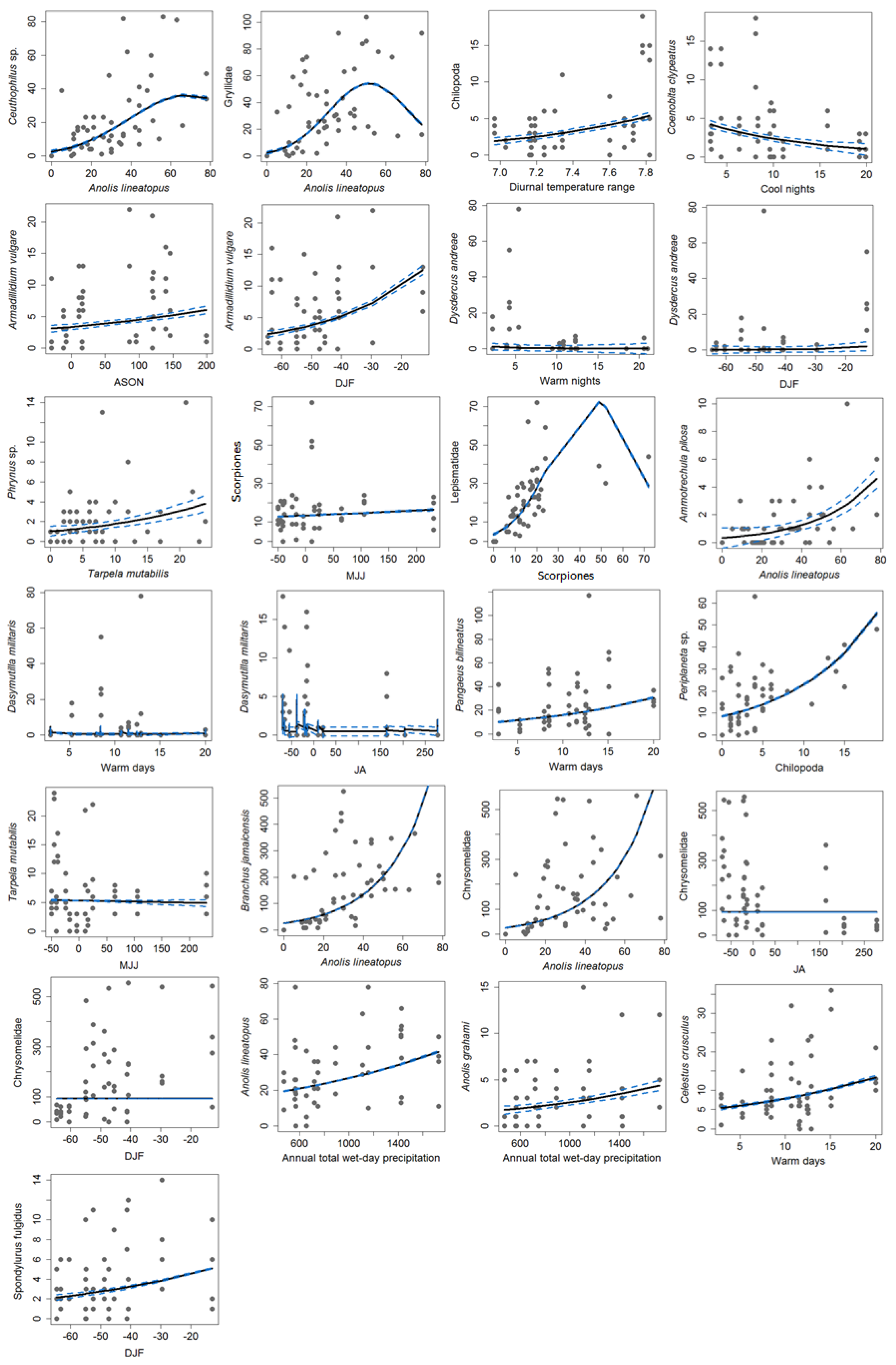
Final taxon models were obtained for 20 of the 22 taxa (
Figure 3;
Table 3). Significant models were not obtained for springtails (Collembola) and Jamaican blue-tailed galliwasps (
Celestus duquesneyi). Three taxa, namely crickets and silverfish (
Ceuthophilus sp., Gryllidae, Lepismatidae), were found to have non-linear (polynomial) relationships with their predictors, but all others were found to be linear. It is noted that 16 of 20 models retained only one predictor. The models had exclusively either only climate or another species as predictors, except for leaf beetles (Chrysomelidae), which had both Jamaican gray anoles (
Anolis lineatopus) and a climate variable (rainfall during the mid-summer dry spell (JA) and dry season (DJF)) as predictors (
Figure 3;
Table 3). The predominant predictors were climate indices, more so rainfall than temperature. Thirteen of the 20 models included a climate variable as a predictor, with 12 having only climate predictors (
Figure 3;
Table 3). The rainfall predictors retained by the models were total annual rainfall and seasonal rainfall (
Figure 3;
Table 3). Among the seasonal rainfall variables retained, the dry season (DJF) was most frequently identified as a significant predictor, and the late wet season (ASON) identified least frequently (
Figure 3;
Table 3). Three of the four lizard taxa for which relationships were obtained were strongly related to rainfall, particularly total annual rainfall and dry season rainfall (DJF) (
Figure 3;
Table 3).
Extreme temperature indices emerged as the temperature predictors retained, i.e., as opposed to mean temperatures (
Figure 3;
Table 3). The percentage of days when maximum or minimum temperature exceeded the 90th percentile (TX90P and TN90P) explained five of the six temperature related taxa (
Table 3). Diurnal temperature range (DTR) was the other temperature variable retained for one taxon, centipedes (Chilopoda).
Seven taxa were predicted solely by abundance of another species (
Figure 3;
Table 3). In all cases, the species predictors were themselves all predicted solely by a climate variable, suggesting an indirect climate influence. Predator-prey association was apparent in the eight taxa that had abundance of other species as predictors. The largest explained variance among all models occurred in velvet ants (
Dasymutilla militaris) (mR
2 = 59%, cR
2 = 68.6%) predicted by TX90P and rainfall during JA. Lowest explained variance for any model (mR
2 = 8.1%, cR
2 = 8.1%) was for stink bugs (
Pangeus sp.) predicted by TX90P. The three polynomial models yielded among the highest explained variances of all models with cR
2 ranging from 47.7% for crickets (Gryllidae) to 65.5% silverfish (Lepismatidae).
It is noted that the influence of temperature was most often positive. In cases of apparent predator-prey relationships, prey species were positively influenced by their predictors up to a point, after which a further increase in predator abundance became a negative influence, and predators increased in abundance with an increase in their prey.
3.4. The HH Model
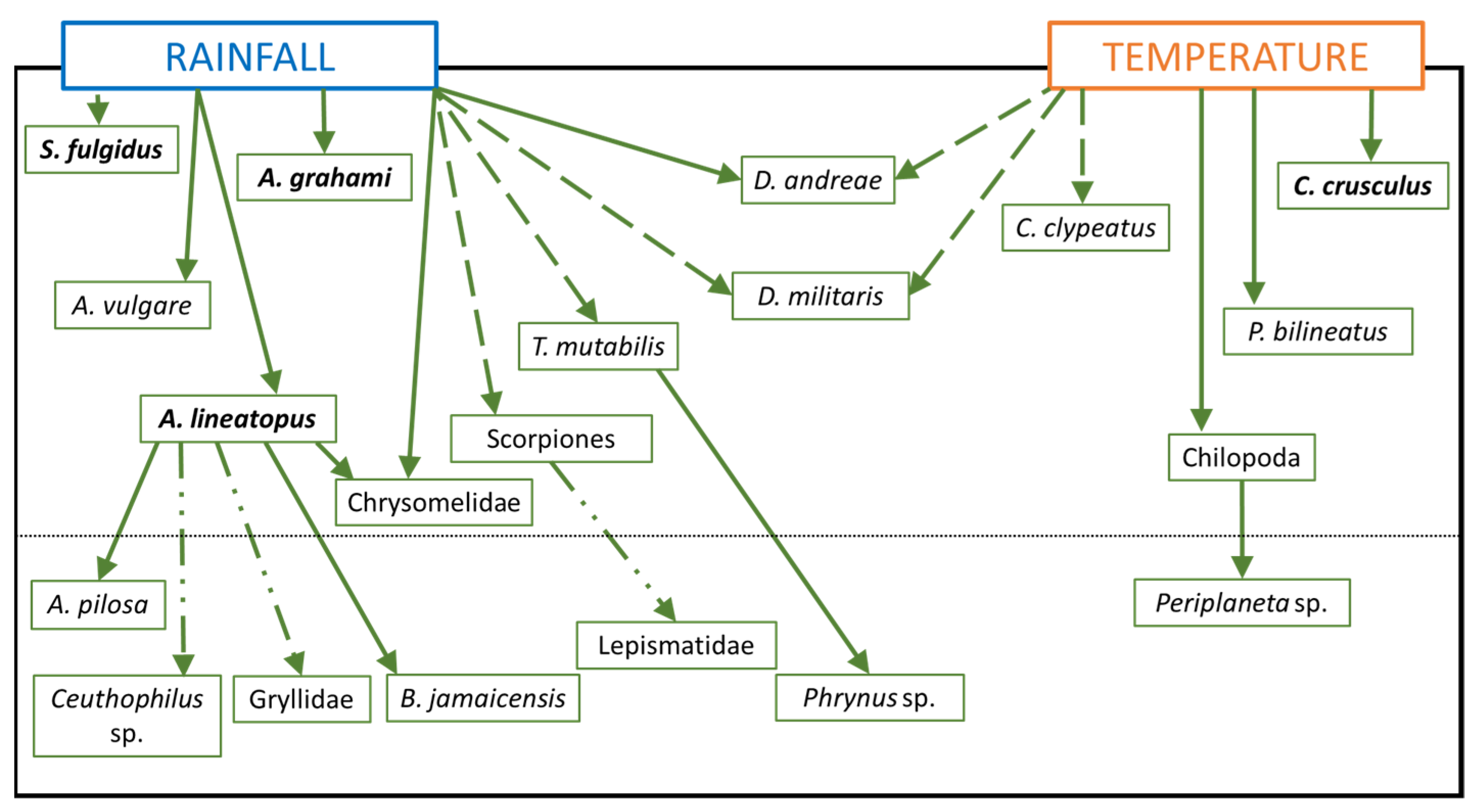
Using the relationships identified from the individual models, a simple schematic diagram compiling all taxon models for the Hellshire Hills (the HH model) was created (
Figure 4). In the figure, the predictor-predictand relationships of the individual models are represented by the arrow directions, i.e., from predictors to predictand. It is not, however, implied that the relationship suggested by each arrow explains the same amount of variability.
The HH model is simple, comprehensive, and climate-based. Although it captures climate-species and species-species relationships, it is guided solely by statistical relationships and does not capture any feedback mechanisms. The HH model has only climate variables (rainfall and temperature) as inputs, which are then used to estimate taxon abundance. It is this latter property of the HH model that facilitates a determination of how changes in climate may potentially influence the abundance of the species present in the Hellshire Hills.
3.5. Model Validation
The models varied in performance. However, they generally capture magnitudes and patterns. Magnitudes and trends in abundance during the independent period were also captured reasonably by most models. However, these predictions tended to be more accurate in the last two to three years (2011–2013) than in the initial year (2010). Six models showed little to no predicted trend during the independent period, namely crickets, centipedes, hermit crabs, red cotton stainers, boxy-whip scorpions, and cockroaches (Ceuthophilus sp., Chilopoda, Coenobita clypeatus, Dysdercus andreae, Phrynus sp., and Periplaneta sp.), despite trends being evident in observed data. Population size was not necessarily a hindrance to prediction, as is evident in the accuracy of predictions for solpugids (Ammotrechula pilosa) and Jamaican skinks (Spondylurus fulgidus) during the independent period.
Mean squared error, root mean squared error, and mean absolute error (see
Table 4) were lowest in models for which trends seemed to be best captured. RMSE estimates for these models all fell below 2.0. These were followed by eight models that had RMSE values below 10.0. High error estimates for remaining models, particularly beetles (
Branchus jamaicensis and Chrysomelidae), indicate divergence between observed and predicted abundance.
3.6. Model Projections
Projected mean change factors estimated for each taxon under each scenario for the end of the century varied (
Table 5), i.e., they indicated a decrease or no change in many taxa, but also an increase in others, with some clear differences dependent on the scenario considered.
Under the A2 scenario, eight taxa are projected to decrease, including three of the five lizards assessed. These groups are pill bugs, red cotton stainers, solpugids, beetles, gray anoles, turquoise anoles, and skinks (Armadillidium vulgare, Dysdercus andreae, Ammotrechula pilosa, Branchus jamaicensis, Chrysomelidae, Anolis lineatopus, Anolis grahami, and Spondylurus fulgidus). It is notable that this group consists entirely of species predicted by total or seasonal rainfall, or species predicted by other species that were in turn predicted by rainfall. Four taxa are projected to experience no discernible change in abundance, three of which were predicted by temperature indices. Eight taxa were projected to increase in abundance. These groups were crickets, scorpions, silverfish, velvet ants, stink bugs, beetles, and common galliwasps (Ceuthophilus sp., Gryllidae, Scorpiones, Lepismatidae, Dasymutilla militaris, Pangaeus bilineatus, Tarpela mutabilis, and Celestus crusculus). The magnitudes of the changes are, however, on a much smaller scale than the losses estimated for the eight taxa that decreased. The predictors also represent a ‘mixed bag’ including rainfall, temperature and other species. Of most interest was the fact that two of the taxa that increased (cricket taxa Ceuthophilus sp. and Gryllidae) are predicted by gray anoles (Anolis lineatopus), for which abundance decreased under the significant drying. The latter is known to be a predator of the former.
Under the B2 scenario, an increase was projected for 11 taxa, including all lizards. These were pill bugs, red cotton stainers, scorpions, silverfish, cockroaches, leaf beetles, greay anoles, turquoise anoles, common galliwasps, and skinks (Armadillidium vulgare, Dysdercus andreae, Scorpiones, Lepismatidae, Periplaneta sp., Branchus jamaicensis, Chrysomelidae, Anolis lineatopus, Anolis grahami, Celestus crusculus, and Spondylurus fulgidus). Interestingly, all but two of these 11 taxa are predicted by rainfall directly or indirectly. The suggestion is that the taxa modelled may be sensitive to the magnitude of change, particularly with respect to rainfall. Decreased abundance was projected for nine arthropods and no lizards. These taxa were crickets, centipedes, hermit crabs, boxy whip scorpions, solpugids, velvet ants, stink bugs, and beetles (Ceuthophilus sp., Gryllidae, Chilopoda, Coenobita clypeatus, Phrynus sp., Ammotrechula pilosa, Dasymutilla militaris, Pangaeus bilineatus and Tarpela mutabilis).
In comparing the projections for the two scenarios, eight taxa shifted from a pattern of increase or no change under A2 to one of decrease under B2. The reverse was noted in seven. For taxa that increased under both scenarios, that change was larger under the B2 scenario.
A2 and B2 generally were both dry scenarios, with A2 indicating severe drying. There appears to be sensitivity to the magnitude of drying, particularly among lizard taxa and those taxa predicted by the lizards. The effect of increased temperatures is not entirely evident from the models created.
4. Discussion
The assessment of the historical and future climate of the Hellshire Hills indicates a warming and drying dry forest. Results of this study support the idea of climate sensitivity among the fauna of the site and provide indications of how that sensitivity is being manifested. For example, (i) some of the strongest correlations were with seasonal totals for rainfall and derived temperature indices rather than for annual totals; (ii) in general, covariation tended to be stronger with rainfall (especially for the lizard species), and (iii) not all taxa exhibited a direct climate-species link but indirect climate linkages were suggested, i.e., even though some species did not exhibit a strong correlation with climate, they were significantly correlated with another species that did. The simple HH model offers insight into the equilibrium among species as aspects of their habitat change over time.
The models produced predominantly used rainfall variables as predictors, but also included temperature predictors for a number of taxa. Dry season rainfall was identified as the predictor of Jamaican skinks (
Spondylurus fulgidus, formerly
Mabuya mabouya). This can feasibly be linked to the strong wet-dry seasonality of skink reproduction, particularly given that birth of young and peak testis size have been known to occur in at least three other species of tropical skinks (
Mabuya frenata,
M. heathi and
M. nigropunctata) in the late dry season or early wet season [
72]. Number of warm days was identified as a predictor of common galliwasps (
Celestus crusculus), indicating that warmer ambient temperatures are preferable for maintaining optimal body temperature as has been seen in other actively thermoregulating anguids [
73].
Five arthropod taxa were predicted by gray anoles (
Anolis lineatopus). These were of the orders Solpugida (solpugids), Orthoptera (crickets), and Coleoptera (beetles). Whereas crickets and beetles are known prey choices of regional anoles [
22,
74,
75], solpugids may be linked to anoles as prey or competition, as has been seen in previous studies of interactions between anoles and arachnids [
76,
77]. Solpugids have been found to be consumed by lizards such as side-blotched lizards and geckos [
78,
79] and are also known to prey on lizards and larger prey [
80], but no studies directly showing interaction between solpugids and West Indian anoles were found during this study. Two additional likely predator-prey interactions were noted, in which boxy-whip scorpions (
Phrynus sp.) was predicted by beetles (
Tarpela mutabilis) and scorpions (Scorpiones) predicted silverfish (Lepismatidae). Though these interactions have not been directly studied in the region,
Phrynus sp. and Scorpiones have been found to opportunistically prey on a wide range of arthropods (and vertebrates), particularly under food stress [
76,
80,
81,
82,
83]. In both instances, the predictor was one that was modulated by rainfall.
While temperature is a contributing factor, seasonal rainfall and soil moisture have been found in previous studies to be particularly critical to the survival of pill bugs (
Armadillidium vulgare) [
84,
85]. Prior studies assessing
A. vulgare have noted desiccation during periods of drought due to reduced moisture in leaf litter, and strong linkages between population density and summer rainfall of the previous year [
84].
Combinations of temperature and rainfall predictors were identified for red cotton stainers and velvet ants (
Dysdercus andreae and
Dasymutilla militaris). The red cotton stainer,
Dysdercus andreae, was predicted by a negative association with number of warm nights and a positive association with dry season rainfall. However, higher rainfall has been linked to lower abundance in
D. andreae [
86] and higher temperature to greater growth and reproduction in a similar species,
D. fasciatus [
87]. It should be noted, therefore, that the climate influences found by the model were accurate, but in the opposite direction of those previously identified. The velvet ant
Dasymutilla militaris was predicted by rainfall during the midsummer dry spell (JA) and number of warm days, both inverse linkages. This temperature relationship is expected in species of Mutillidae, as they are most active during the early morning and late afternoon when temperatures are cooler, and least active during warmer hours, indicating that warming would be detrimental to feeding and reproduction and may impact patterns of behavior [
88]. Similarly, it has been argued that dry conditions are more favorable to velvet ants, though this has not been proven [
89].
Four taxa were predicted solely by temperature variables. Diurnal temperature range was identified as a predictor of the centipedes (Chilopoda), which thereby predicted a known prey species of the group, cockroaches (
Periplaneta sp.) [
90,
91]. Though centipedes in some parts of the Caribbean region have been relatively well identified [
92,
93], the interactions of centipedes with their environments are not generally well known. However, in a study of preferences of 12 species of Chilopoda in a river valley that varied in habitat types, xerothermic conditions were most frequently chosen over wetter and cooler habitat types [
94]. Given the highly significant importance of air and soil temperatures to Chilopoda [
95], it follows that diurnal temperature range is a reasonable influence of abundance. In very cold conditions, it is typical of the burrowing stinkbug (
Pangaeus bilineatus) to occupy deeper layers of soil in suspended development [
96]. The increase in temperature associated with the transition from winter towards summer months, however, results in individuals increasingly occupying soil closer to the surface and becoming reproductively viable [
96], indicating that an increase in the number of warm days as seasons change has strong influence on activity of
P. bilineatus. Assessments of a similar Cydnidae species,
Cyrtomenus bergi, found temperature to be a significant limiting factor to development [
97]. In the study, a temperature of 26 °C was found to be optimal for growth, and immature developmental stages were more resilient under extreme temperatures, provided that temperatures did not exceed 31 °C [
97]. Haemolymph composition and alkalinity of the terrestrial hermit crab (
Coenobita clypeatus) is temperature regulated [
98]. A decrease in temperature is associated with an increase in alkalinity and decrease in the partial pressure of carbon dioxide of haemolymph, and also results in reduced activity, as is characteristic of ectotherms [
98], indicating that an increase in the number of cool nights can feasibly have a negative effect on hermit crab abundance as the model suggests. Larval development of tropical
Coenobita sp. may also be affected by changes in temperature [
99].
Bioclimatic modelling has been used minimally in the Caribbean to investigate climate change impacts such as vegetative distribution in Trinidad [
20], geographic range shifts of orchids and vectors of parasites in Colombia [
100,
101], and climate suitability for coffee in Puerto Rico [
102]. The results of this study agree with previous studies, showing that tropical organisms are vulnerable to climatic shifts, will vary in their responses depending on taxon specific tolerance, and are likely to decrease in abundance or distribution due to climate change. Notwithstanding their utility, bioclimatic modelling has not widely been undertaken for small scales such as insular tropical dry forest ecosystems, but is more frequently implemented for continental or large island scales. Accordingly, the application of modelling techniques to smaller systems such as islands and small forested areas, which are often centres of endemism and thus high conservation priorities, is urgently needed [
20].
The species models’ individual abilities reflected varied performances using the metrics of validation, i.e., RSME, patterns of prediction for the training and independent period, and explained variability. The validation process suggested that the models generally captured magnitudes and patterns, particularly for the training period, with notable peaks being represented in most cases. Magnitudes and trends in abundance during the independent period tended, however, to be less reasonably captured by most models. Where an indirect climate link was being captured by a model, the potential for divergence from observed values increased due to the propagation of errors. Nonetheless, the presence of some skill for each model suggested that the simple HH model can be used for the indicative purpose of determining the influence of future climate on faunal abundance in the Hellshire Hills and other dry forests on the small islands of the Caribbean region.
When future climate scenarios were used in the simple HH model, changes in end of century faunal abundance were strongly dependent on the future climate scenario. Given that many of the relationships identified were closely linked to seasonal rainfall totals, it seems that there is a sensitivity in both arthropods and lizards to the magnitude of change in rainfall, which was different (particularly for the dry season) under the two scenarios.
The implications of these results are not only evident for the Hellshire Hills, but can be extrapolated to other small dry forests in the region. Firstly, there is an indisputable link between Caribbean dry forest fauna and climatic change, whether through direct modulation by rainfall and temperature, or through indirect effects among predators and prey. These linkages are expected to be non-linear, and driven by patterns of change rather than magnitude. Secondly, studies on these climatic influences must be specific to these forests and the taxa that occupy them. In providing a simple methodology, the HH model illustrates that this is entirely achievable once there are ecological and climatological datasets available. It allows for expansion of the pool of climate predictors used to determine with even greater clarity the exact nature of the climate influence in any area. It also emphasizes the value of existing ecological and climatological datasets across the insular Caribbean, and the necessity of improving our understanding of terrestrial and marine ecosystems through ongoing field assessments. Thirdly, climate conservation will become critical to small islands as climate change continues to progress. Evaluation of extinction risks due to climate change will be a necessary component of any ecological conservation program in the Caribbean, as will climate change mitigation and adaptation measures.
This study has demonstrated that, even where they are well preserved, dry forest ecosystems are not immune to the influence of climate change. There is, therefore, a clear need for mainstreaming biodiversity monitoring and implementing local targeted climate mitigation and adaptation measures. The creation and enforcement of protected areas and conservation zones become even more relevant for vulnerable areas like the Hellshire Hills [
44]. Additionally, there is a need for greater research into dry forest carbon storage capacity and ecosystem services, as well as identification of appropriate management tools and strategies based on unique characteristics of particular dry forests [
19].
To improve the techniques developed here, knowledge gaps must be filled and data acquired for both relatively undisturbed and degraded habitats. This will facilitate an evaluation of the future climate risks to the fauna of these ecosystems, and will require the collaboration of environmental organisations and climate scientists across the region.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



