
CO2 Enrichment in Protected Agriculture: A Systematic Review of Greenhouses, Controlled Environment Systems, and Vertical Farms—Part 2

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification
2.2. Screening
2.3. Analysis
3. Results and Discussion
3.1. Impact of CO2 Enrichment on Crop Physiology and Photosynthesis
3.2. Direct Effect of CO2 Enrichment on Crop Production and Yield
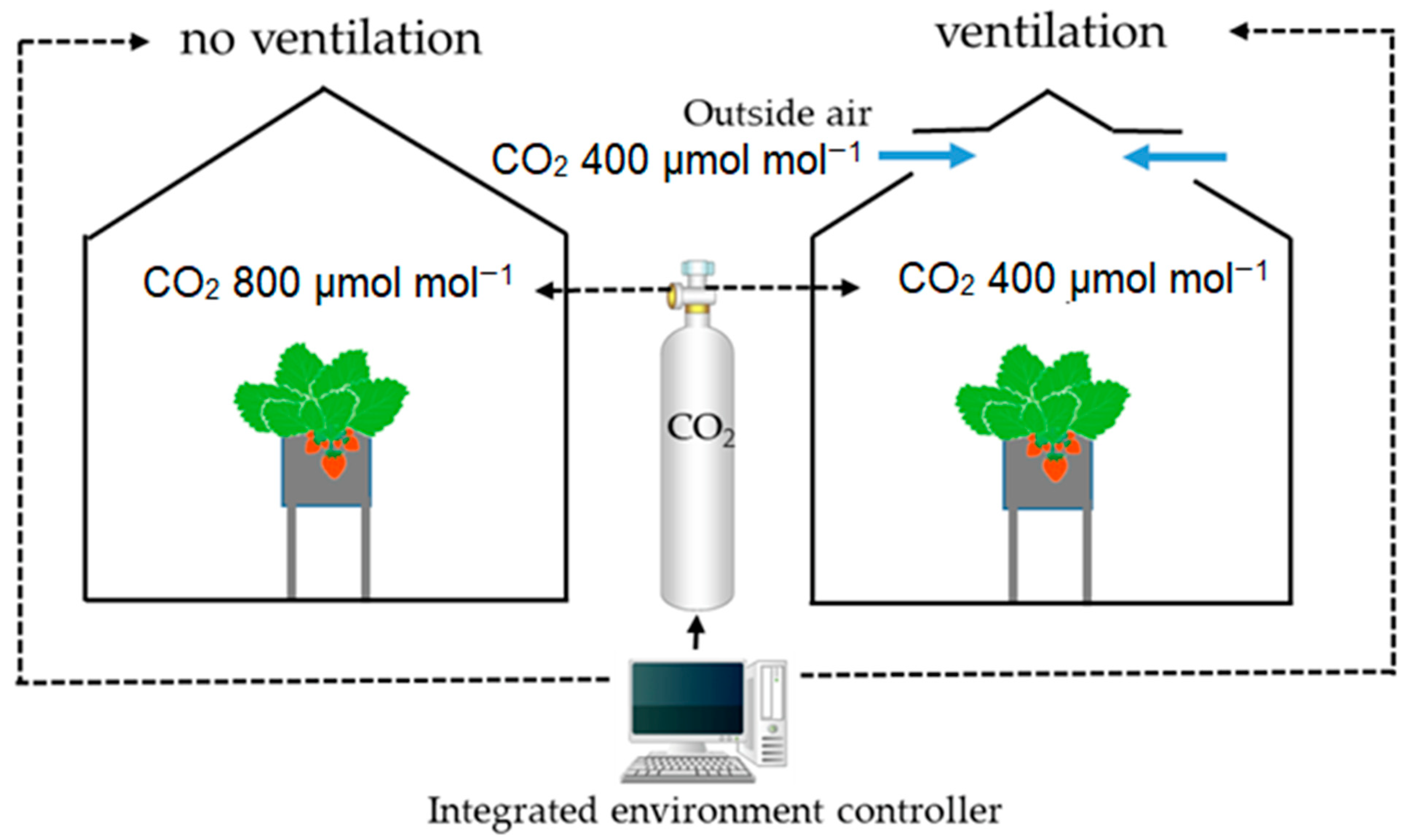
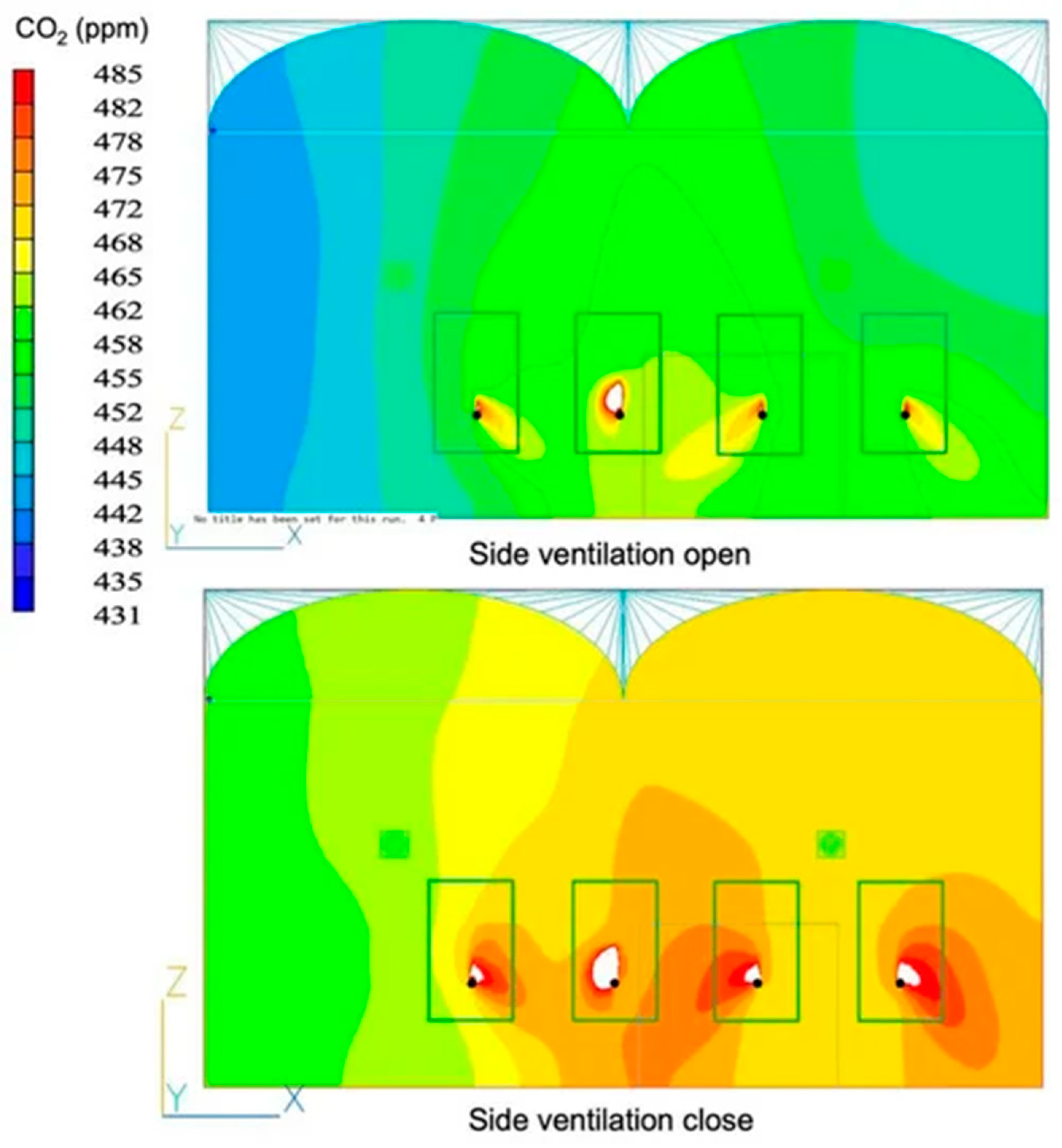
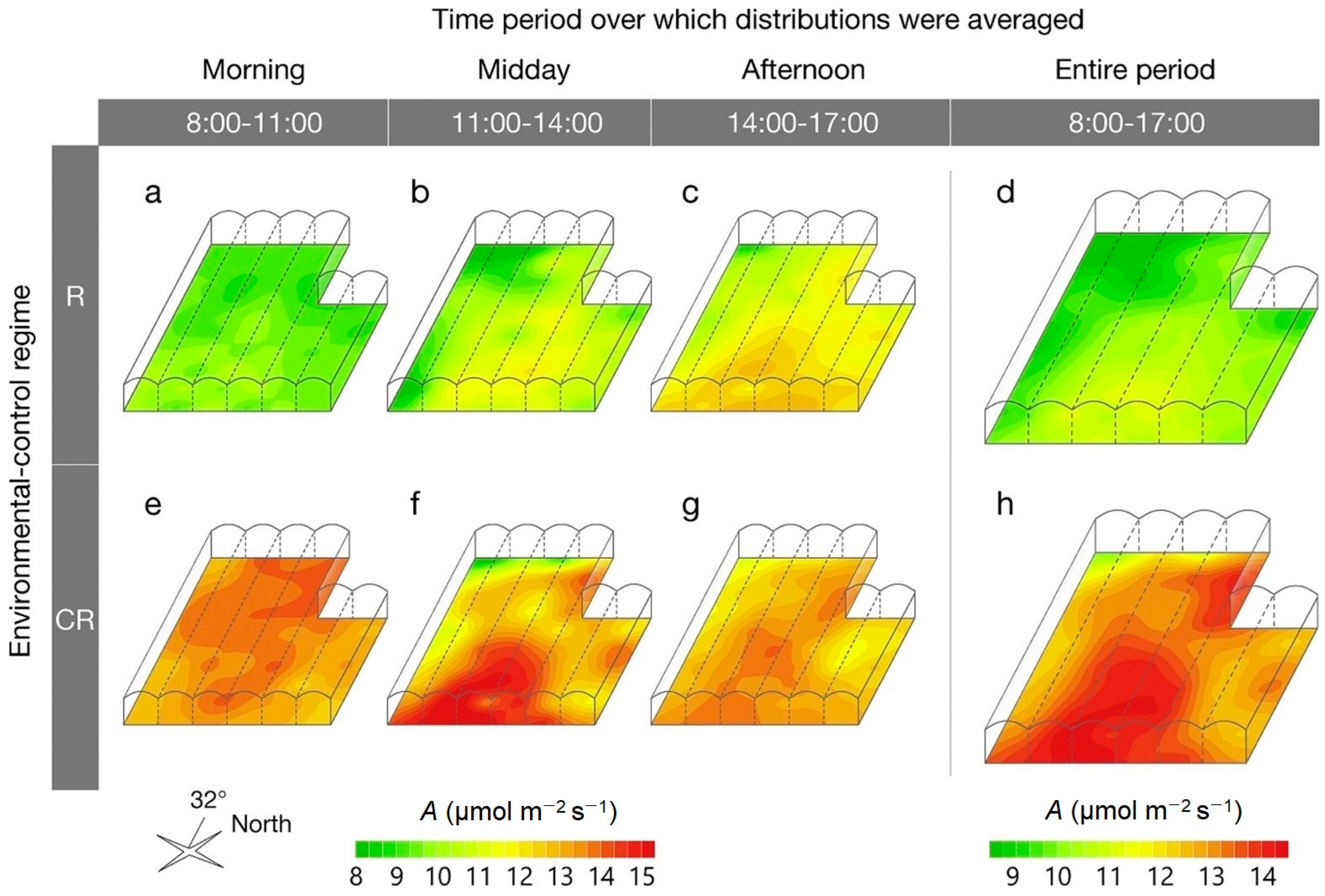
3.3. Application of Modeling and Simulation Techniques to Analyze the Distribution of CO2 and Its Impact on Crops Within Protected Environments
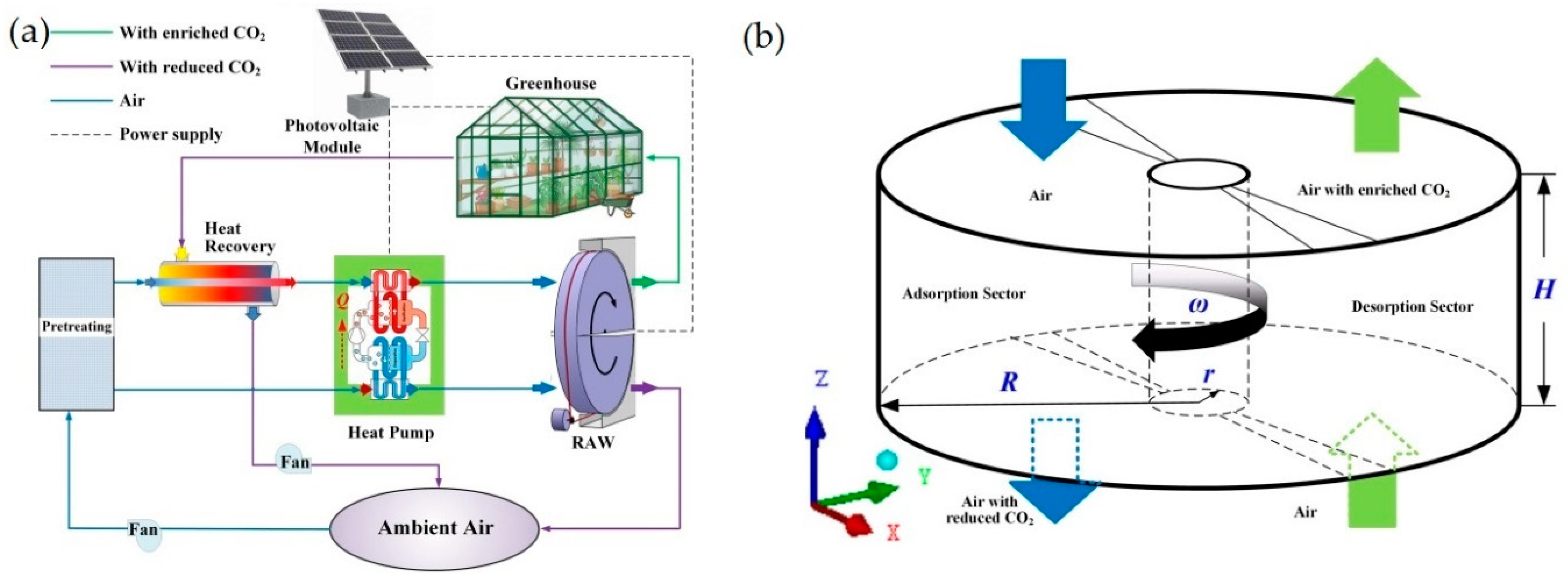
3.4. CO2 Injection and Enrichment Technologies
3.5. Plant Factory with Artificial Lighting (PFAL) and CO2 Enrichment
3.6. Economic Aspects and Energy Efficiency of CO2 Enrichment in Protected Agriculture
3.7. Environmental Aspects and Sustainability of CO2 Enrichment in Protected Agriculture
4. Challenges and Knowledge Gaps
5. Future Research Development Needs
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tahery, D.; Roshandel, R.; Avami, A. An Integrated Dynamic Model for Evaluating the Influence of Ground to Air Heat Transfer System on Heating, Cooling and CO2 Supply in Greenhouses: Considering Crop Transpiration. Renew. Energy 2021, 173, 42–56. [Google Scholar]
- Agency, I.E.; Birol, F. World Energy Outlook 2013; International Energy Agency Paris: Paris, France, 2013; ISBN 9264201300. [Google Scholar]
- Masudi, F. Greenhouses Key to Water and Food Security in UAE, Expert Says. Gulf News. 2019. Available online: https://gulfnews.com/uae/environment/greenhouses-key-to-water-and-food-security-in-uae-expert-says-1.1349562 (accessed on 31 December 2024).
- Vadiee, A.; Martin, V. Energy Management Strategies for Commercial Greenhouses. Appl. Energy 2014, 114, 880–888. [Google Scholar] [CrossRef]
- Vadiee, A.; Martin, V. Energy Management in Horticultural Applications through the Closed Greenhouse Concept, State of the Art. Renew. Sustain. Energy Rev. 2012, 16, 5087–5100. [Google Scholar]
- De Gelder, A.; Dieleman, J.A.; Bot, G.P.A.; Marcelis, L.F.M. An Overview of Climate and Crop Yield in Closed Greenhouses. J. Hortic. Sci. Biotechnol. 2012, 87, 193–202. [Google Scholar] [CrossRef]
- Takeya, S.; Muromachi, S.; Maekawa, T.; Yamamoto, Y.; Mimachi, H.; Kinoshita, T.; Murayama, T.; Umeda, H.; Ahn, D.-H.; Iwasaki, Y. Design of Ecological CO2 Enrichment System for Greenhouse Production Using TBAB + CO2 Semi-Clathrate Hydrate. Energies 2017, 10, 927. [Google Scholar] [CrossRef]
- Heuvelink, E.; Bakker, M.; Marcelis, L.F.M.; Raaphorst, M. Climate and Yield in a Closed Greenhouse. Acta Hortic. 2008, 801, 1083–1092. [Google Scholar]
- Singh, H.; Poudel, M.R.; Dunn, B.L.; Fontanier, C.; Kakani, G. Effect of Greenhouse CO2 Supplementation on Yield and Mineral Element Concentrations of Leafy Greens Grown Using Nutrient Film Technique. Agronomy 2020, 10, 323. [Google Scholar] [CrossRef]
- Chen, D.; Mei, Y.; Liu, Q.; Wu, Y.; Yang, Z. Carbon Dioxide Enrichment Promoted the Growth, Yield, and Light-use Efficiency of Lettuce in a Plant Factory with Artificial Lighting. Agron. J. 2021, 113, 5196–5206. [Google Scholar]
- Roy, J.C.; Pouillard, J.B.; Boulard, T.; Fatnassi, H.; Grisey, A. Experimental and CFD results on the CO2 distribution in a semi closed greenhouse. Acta Hortic. 2014, 1037, 993–1000. [Google Scholar] [CrossRef]
- Zhang, Y.; Yasutake, D.; Hidaka, K.; Kitano, M.; Okayasu, T. CFD Analysis for Evaluating and Optimizing Spatial Distribution of CO2 Concentration in a Strawberry Greenhouse under Different CO2 Enrichment Methods. Comput. Electron. Agric. 2020, 179, 105811. [Google Scholar] [CrossRef]
- Zhang, Y.; Yasutake, D.; Hidaka, K.; Kimura, K.; Okayasu, T.; Kitano, M.; Hirota, T. Eco-Friendly Strategy for CO2 Enrichment Performance in Commercial Greenhouses Based on the CO2 Spatial Distribution and Photosynthesis. Sci. Rep. 2023, 13, 17277. [Google Scholar] [CrossRef] [PubMed]
- Syed, A.M.; Hachem, C. Review of Design Trends in Lighting, Environmental Controls, Carbon Dioxide Supplementation, Passive Design, and Renewable Energy Systems for Agricultural Greenhouses. J. Biosyst. Eng. 2019, 44, 28–36. [Google Scholar] [CrossRef]
- Bailey, B.J. Optimal Control of Carbon Dioxide Enrichment in Tomato Greenhouses. Acta Hortic. 2001, 578, 63–69. [Google Scholar] [CrossRef]
- Li, Y.; Ding, Y.; Li, D.; Miao, Z. Automatic Carbon Dioxide Enrichment Strategies in the Greenhouse: A Review. Biosyst. Eng. 2018, 171, 101–119. [Google Scholar] [CrossRef]
- Wischlitzki, E.; Amler, N.; Hiller, J.; Drexler, H. Psychosocial Risk Management in the Teaching Profession: A Systematic Review. Saf. Health Work 2020, 11, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Kang, Q.; Song, Z. The Current State of Systematic Reviews in Library and Information Studies. Libr. Inf. Sci. Res. 2015, 37, 296–310. [Google Scholar] [CrossRef]
- Scheerder, A.; Van Deursen, A.; Van Dijk, J. Determinants of Internet Skills, Uses and Outcomes. A Systematic Review of the Second-and Third-Level Digital Divide. Telemat. Inform. 2017, 34, 1607–1624. [Google Scholar] [CrossRef]
- Villagran, E.; Espitia, J.J.; Velázquez, F.A.; Rodriguez, J. Solar Dryers: Technical Insights and Bibliometric Trends in Energy Technologies. AgriEngineering 2024, 6, 4041–4063. [Google Scholar] [CrossRef]
- Yao, L.; Zhang, Y.; Zhao, C.; Zhao, F.; Bai, S. The PRISMA 2020 Statement: A System Review of Hospital Preparedness for Bioterrorism Events. Int. J. Environ. Res. Public Health 2022, 19, 16257. [Google Scholar] [CrossRef]
- Espitia, J.J.; Velázquez, F.A.; Rodriguez, J.; Gomez, L.; Baeza, E.; Aguilar-Rodríguez, C.E.; Flores-Velazquez, J.; Villagran, E. Solar Energy Applications in Protected Agriculture: A Technical and Bibliometric Review of Greenhouse Systems and Solar Technologies. Agronomy 2024, 14, 2791. [Google Scholar] [CrossRef]
- Hao, X.; Wang, Q.; Khosla, S. Responses of a Long Greenhouse Tomato Crop to Summer CO2 Enrichment. Can. J. Plant Sci. 2006, 86, 1395–1400. [Google Scholar]
- Zhang, Z.; Yuan, H.; Liu, Y.; Li, J.; Zheng, J.; Sun, S.; Xing, G. Photosynthetic Responses of Tomato to Different Concentrations of CO2 Enrichment in Greenhouse. J. Plant Nutr. Fertil. 2018, 24, 1010–1018. [Google Scholar]
- Nilsen, S.; Hovland, K.; Dons, C.; Sletten, S.P. Effect of CO2 Enrichment on Photosynthesis, Growth and Yield of Tomato. Sci. Hortic. 1983, 20, 1–14. [Google Scholar]
- Ayari, O.; Dorais, M.; Gosselin, A. Daily Variations of Photosynthetic Efficiency of Greenhouse Tomato Plants during Winter and Spring. J. Am. Soc. Hortic. Sci. 2000, 125, 235–241. [Google Scholar] [CrossRef]
- Dannehl, D.; Kläring, H.-P.; Schmidt, U. Light-Mediated Reduction in Photosynthesis in Closed Greenhouses Can Be Compensated for by CO2 Enrichment in Tomato Production. Plants 2021, 10, 2808. [Google Scholar] [CrossRef]
- Tartachnyk, I.I.; Blanke, M.M. Photosynthesis and Transpiration of Tomato and CO2 Fluxes in a Greenhouse under Changing Environmental Conditions in Winter. Ann. Appl. Biol. 2007, 150, 149–156. [Google Scholar]
- Pan, T.; Ding, J.; Qin, G.; Wang, Y.; Xi, L.; Yang, J.; Li, J.; Zhang, J.; Zou, Z. Interaction of Supplementary Light and CO2 Enrichment Improves Growth, Photosynthesis, Yield, and Quality of Tomato in Autumn through Spring Greenhouse Production. HortScience 2019, 54, 246–252. [Google Scholar] [CrossRef]
- Nomura, K.; Saito, M.; Tada, I.; Yasutake, D.; Kimura, K.; Kitano, M. Simulating the Photosynthetic and Annual-Yield Enhancement of a Row-Planted Greenhouse Tomato Canopy Through Diffuse Covering, CO2 Enrichment, and High-Wire Techniques. Horticulturae 2024, 10, 1210. [Google Scholar] [CrossRef]
- Haghighi, M.; Golabdar, S.; Abolghasemi, R.; Kappel, N. CO2 Enrichment Changed N Metabolism of Tomatoes under Salinity Stress. Sci. Hortic. 2022, 305, 111412. [Google Scholar]
- Du, B.; Shukla, M.K.; Yang, X.; Du, T. Enhanced Fruit Yield and Quality of Tomato by Photosynthetic Bacteria and CO2 Enrichment under Reduced Irrigation. Agric. Water Manag. 2023, 277, 108106. [Google Scholar]
- Pan, C.; Ahammed, G.J.; Li, X.; Shi, K. Elevated CO2 Improves Photosynthesis under High Temperature by Attenuating the Functional Limitations to Energy Fluxes, Electron Transport and Redox Homeostasis in Tomato Leaves. Front. Plant Sci. 2018, 9, 1739. [Google Scholar]
- Dorais, M.; Ayari, O.; Samson, G.; Gosselin, A. Does Carbohydrate Accumulation Affect the Photosynthetic Efficiency of Tomato Leaves? Acta Hortic. 2001, 554, 251–260. [Google Scholar]
- Zheng, S.; Yang, L.; Zheng, H.; Wu, J.; Zhou, Z.; Tian, J. Identification of Hub Genes and Physiological Effects of Overexpressing the Photosynthesis-Related Gene Soly720 in Tomato under High-CO2 Conditions. Int. J. Mol. Sci. 2024, 25, 757. [Google Scholar] [CrossRef]
- Boondum, S.; Chulaka, P.; Kaewsorn, P.; Nukaya, T.; Takagaki, M.; Yamori, W. Carbon Dioxide (CO2) Enrichment in Greenhouse Enhanced Growth and Productivity of Tomato (Solanum lycopersicum L.) during Winter. Acta Hortic. 2018, 1245, 61–64. [Google Scholar]
- Porras, M.E.; Lorenzo, P.; Medrano, E.; Sánchez-González, M.J.; Otálora-Alcón, G.; Piñero, M.C.; Del Amor, F.M.; Sánchez-Guerrero, M.C. Photosynthetic Acclimation to Elevated CO2 Concentration in a Sweet Pepper (Capsicum Annuum) Crop under Mediterranean Greenhouse Conditions: Influence of the Nitrogen Source and Salinity. Funct. Plant Biol. 2017, 44, 573–586. [Google Scholar] [PubMed]
- Porras, M.E.; Medrano, E.; Lorenzo, P.; Sánchez-González, M.J.; Baeza, E.J.; Sánchez-Guerrero, M.C. Sweet Pepper Acclimation to Variable CO2 Supply in a Mediterranean Greenhouse. Acta Hortic. 2015, 1170, 797–804. [Google Scholar]
- Rezende, F.C.; Frizzone, J.A.; de Oliveira, R.F.; Pereira, A.S. CO2 and Irrigation in Relation to Yield and Water Use of the Bell Pepper Crop. Sci. Agric. 2003, 60, 7–12. [Google Scholar]
- Jang, Y.; Mun, B.; Do, K.; Um, Y.; Chun, C. Effects of Photosynthetic Photon Flux and Carbon Dioxide Concentration on the Photosynthesis and Growth of Grafted Pepper Transplants during Healing and Acclimatization. Hortic. Environ. Biotechnol. 2014, 55, 387–396. [Google Scholar]
- Yasutake, D.; Miyauchi, K.; Mori, M.; Kitano, M.; Ino, A.; Takahashi, A. Multiple Effects of CO2 Concentration and Humidity on Leaf Gas Exchanges of Sweet Pepper in the Morning and Afternoon. Environ. Control Biol. 2016, 54, 177–181. [Google Scholar]
- Vafiadis, D.T.; Papamanthos, C.; Ntinas, G.K.; Nikita-Martzopoulou, C. Influence of CO2 Enrichment in Greenhouses on Pepper Plant (Capsicum annuum L.) Yield under High Temperature Conditions. Acta Hortic. 2012, 952, 749–754. [Google Scholar]
- Koo, J.K.; Hwang, H.S.; Hwang, J.H.; Park, E.W.; Yu, J.; Yun, J.H.; Hwang, S.Y.; Choi, H.E.; Hwang, S.J. Supplemental Lighting and CO2 Enrichment on the Growth, Fruit Quality, and Yield of Cucumber. Hortic. Environ. Biotechnol. 2024, 66, 77–85. [Google Scholar]
- Li, X.; Chu, W.; Dong, J.; Duan, Z. An Improved High-Performance Liquid Chromatographic Method for the Determination of Soluble Sugars in Root Exudates of Greenhouse Cucumber Grown under CO2 Enrichment. J. Am. Soc. Hortic. Sci. 2014, 139, 356–363. [Google Scholar]
- Namizaki, H.; Iwasaki, Y.; Wang, R. Effects of Elevated CO2 Levels on the Growth and Yield of Summer-Grown Cucumbers Cultivated under Different Day and Night Temperatures. Agronomy 2022, 12, 1872. [Google Scholar] [CrossRef]
- Wei, M.; Xing, Y.; Wang, X.; Ma, H. CO2 Environment in Solar Greenhouse in Northern China and Effects of CO2 Enrichment on Photosynthetic Rate and Dry Matter Production of Cucumber. Acta Hortic. 2002, 633, 115–120. [Google Scholar]
- Fukuda, N.; Kanesaka, N.; Okushima, L.; Sase, S. Effects of Over-Night Lighting by Red Light Emitting Diodes on the Growth and Photosynthesis of Lettuce under CO2 Enrichment. Acta Hortic. 2019, 1296, 605–612. [Google Scholar]
- Li, P.P.; Hu, Y.G.; Zhao, Y.G.; Ying, X.J.; Mao, H.P. Comprehensive Model on the Effect of CO2 Enrichment on Lettuce Photosynthesis in Greenhouse. Trans. CSAE 2001, 17, 75–79. [Google Scholar]
- Osman, M.; Qaryouti, M.; Alharbi, S.; Alghamdi, B.; Al-Soqeer, A.; Alharbi, A.; Almutairi, K.; Abdelaziz, M.E. Impact of CO2 Enrichment on Growth, Yield and Fruit Quality of F1 Hybrid Strawberry Grown under Controlled Greenhouse Condition. Horticulturae 2024, 10, 941. [Google Scholar] [CrossRef]
- Han, X.; Sun, Y.; Chen, J.; Wang, Z.; Qi, H.; Liu, Y.; Liu, Y. Effects of CO2 Enrichment on Carbon Assimilation, Yield and Quality of Oriental Melon Cultivated in a Solar Greenhouse. Horticulturae 2023, 9, 561. [Google Scholar] [CrossRef]
- Li, X.; Zhao, J.; Shang, M.; Song, H.; Zhang, J.; Xu, X.; Zheng, S.; Hou, L.; Li, M.; Xing, G. Physiological and Molecular Basis of Promoting Leaf Growth in Strawberry (Fragaria ananassa Duch.) by CO2 Enrichment. Biotechnol. Biotechnol. Equip. 2020, 34, 905–917. [Google Scholar] [CrossRef]
- Zhou, Y.; Mahmoud Ali, H.S.; Xi, J.; Yao, D.; Zhang, H.; Li, X.; Yu, K.; Zhao, F. Response of Photosynthetic Characteristics and Yield of Grape to Different CO2 Concentrations in a Greenhouse. Front. Plant Sci. 2024, 15, 1378749. [Google Scholar]
- Jung, D.H.; Hwang, I.; Son, J.E. Three-Dimensional Estimation of Greenhouse-Grown Mango Photosynthesis with Different CO2 Enrichment Heights. Hortic. Environ. Biotechnol. 2022, 63, 823–834. [Google Scholar] [CrossRef]
- Wada, Y.; Soeno, T.; Inaba, Y. Effects of Light and Temperature on Photosynthetic Enhancement by High CO2 Concentration of Strawberry Cultivar Tochiotome Leaves under Forcing or Half-Forcing Culture. Jpn. J. Crop Sci. 2010, 79, 192–197. [Google Scholar] [CrossRef]
- Tagawa, A.; Ehara, M.; Ito, Y.; Araki, T.; Ozaki, Y.; Shishido, Y. Effects of CO2 Enrichment on Yield, Photosynthetic Rate, Translocation and Distribution of Photoassimilates in Strawberry ‘Sagahonoka’. Agronomy 2022, 12, 473. [Google Scholar] [CrossRef]
- Mavrogianopoulos, G.N.; Spanakis, J.; Tsikalas, P. Effect of Carbon Dioxide Enrichment and Salinity on Photosynthesis and Yield in Melon. Sci. Hortic. 1999, 79, 51–63. [Google Scholar] [CrossRef]
- Hong, T.; Cai, Z.; Li, R.; Liu, J.; Li, J.; Wang, Z.; Zhang, Z. Effects of Water and Nitrogen Coupling on Watermelon Growth, Photosynthesis and Yield under CO2 Enrichment. Agric. Water Manag. 2022, 259, 107229. [Google Scholar] [CrossRef]
- Boutaleb Joutei, A.; Salager, J.L.; Sonie, L.; van Impe, G.; Lebrun, P. Effects of Elevated CO2 on the Physiology and Yield Parameters of Bean. Agronomie 2003, 23, 327–334. [Google Scholar]
- Villagran, E.; Bojacá, C. Analysis of the Microclimatic Behavior of a Greenhouse Used to Produce Carnation (Dianthus caryophyllus L.). Ornam. Hortic. 2020, 26, 109–204. [Google Scholar] [CrossRef]
- Villagran, E.; Bojacá, C. Experimental Evaluation of the Thermal and Hygrometric Behavior of a Colombian Greenhouse Used for the Production of Roses (Rosa spp.). Ornam. Hortic. 2020, 26, 205–291. [Google Scholar] [CrossRef]
- Cho, A.R.; Chung, S.W.; Kim, Y.J. CO2 Enrichment with Light Control Improves Photosynthetic Response and Flower Quality in Phalaenopsis Queen Beer ‘Mantefon’. Acta Hortic. 2019, 1296, 645–650. [Google Scholar] [CrossRef]
- Xu, S.; Zhu, X.; Li, C.; Ye, Q. Effects of CO2 Enrichment on Photosynthesis and Growth in Gerbera Jamesonii. Sci. Hortic. 2014, 177, 77–84. [Google Scholar] [CrossRef]
- Atkins, I.K.; Boldt, J.K. Photosynthetic Responses of Greenhouse Ornamentals to Interaction of Irradiance, Carbon Dioxide Concentration, and Temperature. J. Am. Soc. Hortic. Sci. 2022, 147, 82–94. [Google Scholar] [CrossRef]
- Jiao, J.; Grodzinski, B. Environmental Influences on Photosynthesis and Carbon Export in Greenhouse Roses during Development of the Flowering Shoot. J. Am. Soc. Hortic. Sci. 1998, 123, 1081–1088. [Google Scholar]
- Urban, L.; Barthélémy, L.; Bearez, P.; Pyrrha, P. Effect of Elevated CO2 on Photosynthesis and Chlorophyll Fluorescence of Rose Plants Grown at High Temperature and High Photosynthetic Photon Flux Density. Photosynthetica 2001, 39, 275–281. [Google Scholar] [CrossRef]
- Baille, M.; Romero-Aranda, R.; Baille, A. Gas-Exchange Responses of Rose Plants to CO2 Enrichment and Light. J. Hortic. Sci. 1996, 71, 945–956. [Google Scholar] [CrossRef]
- Salinas-Velandia, D.A.; Romero-Perdomo, F.; Numa-Vergel, S.; Villagrán, E.; Donado-Godoy, P.; Galindo-Pacheco, J.R. Insights into Circular Horticulture: Knowledge Diffusion, Resource Circulation, One Health Approach, and Greenhouse Technologies. Int. J. Environ. Res. Public Health 2022, 19, 12053. [Google Scholar] [CrossRef]
- Villagrán, E.; Flores-Velazquez, J.; Akrami, M.; Bojacá, C. Microclimatic Evaluation of Five Types of Colombian Greenhouses Using Geostatistical Techniques. Sensors 2022, 22, 3925. [Google Scholar] [CrossRef] [PubMed]
- Villagran, E.; Bojacá, C.; Akrami, M. Contribution to the Sustainability of Agricultural Production in Greenhouses Built on Slope Soils: A Numerical Study of the Microclimatic Behavior of a Typical Colombian Structure. Sustainability 2021, 13, 4748. [Google Scholar] [CrossRef]
- Vafiadis, D.; Martzopoulou, A.; Denizopoulou, A.; Fragos, V.P. Plant Growth in Greenhouses Using CO2 Enrichment at High Temperatures with a Passive Solar System. Acta Hortic. 2019, 1296, 417–424. [Google Scholar]
- Higashide, T. Tomatoes: Cultivation, Varieties and Nutrition; Nova Science Publishers, Incorporated: Hauppauge, NY, USA, 2013; ISBN 1624179762. [Google Scholar]
- Huber, B.M.; Louws, F.J.; Hernández, R. Impact of Different Daily Light Integrals and Carbon Dioxide Concentrations on the Growth, Morphology, and Production Efficiency of Tomato Seedlings. Front. Plant Sci. 2021, 12, 615853. [Google Scholar]
- Hidaka, K.; Nakahara, S.; Yasutake, D.; Zhang, Y.; Okayasu, T.; Dan, K.; Kitano, M.; Sone, K. Crop-Local CO2 Enrichment Improves Strawberry Yield and Fuel Use Efficiency in Protected Cultivations. Sci. Hortic. 2022, 301, 111104. [Google Scholar]
- Zhu, S.; Xu, W. Ecophysiological Effects of CO2 Enrichment on Bottle Gour in Plastic House. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2002, 13, 429–432. [Google Scholar]
- Baba, M.Y.; Maroto, J.V.; San Batoutista, A.; Pascual, B.; Lopez, S.; Baixauli, C. Agronomic Response of Sweet Pepper (Capsicum annuum L.) to CO2 Enrichment in Greenhouses with Static Ventilation. Acta Hortic. 2006, 719, 521–528. [Google Scholar] [CrossRef]
- Li, X.; Kang, S.; Li, F.; Zhang, X.; Huo, Z.; Ding, R.; Tong, L.; Du, T.; Li, S. Light Supplement and Carbon Dioxide Enrichment Affect Yield and Quality of Off-season Pepper. Agron. J. 2017, 109, 2107–2118. [Google Scholar] [CrossRef]
- Song, H.; Wu, P.; Lu, X.; Wang, B.; Song, T.; Lu, Q.; Li, M.; Xu, X. Comparative Physiological and Transcriptomic Analyses Reveal the Mechanisms of CO2 Enrichment in Promoting the Growth and Quality in Lactuca sativa. PLoS ONE 2023, 18, e0278159. [Google Scholar] [CrossRef]
- Tamagi, J.T.; Boas, M.A.V.; Sampaio, S.C.; Damasceno, S.; Guimarães, V.F. Effect of CO2 Applied to Lettuce Crop (Lactuca sativa L.) through Irrigation Water. IRRIGA 2007, 12, 492–504. [Google Scholar] [CrossRef]
- Holley, J.; Mattson, N.; Ashenafi, E.; Nyman, M. The Impact of CO2 Enrichment on Biomass, Carotenoids, Xanthophyll, and Mineral Content of Lettuce (Lactuca sativa L.). Horticulturae 2022, 8, 820. [Google Scholar] [CrossRef]
- Shi, S.; Wang, X.; Li, H.; Song, J.; He, X.; Yang, Z. Species-Dependent Response of Brassica chinensis L. to Elevated CO2 Gradients Influences Uptake and Utilization of Soil Nitrogen, Phosphorus and Potassium. Agronomy 2024, 14, 1684. [Google Scholar] [CrossRef]
- Savé i Montserrat, R.; de Herralde Traveria, F.; Codina Mahrer, C.; Sánchez Molino, F.J.; Biel Loscos, C. Effects of Atmospheric Carbon Dioxide Fertilization on Biomass and Secondary Metabolites of Some Plant Species with Pharmacological Interes under Greenhouse Conditions. Afinidad 2007, 64, 237–241. [Google Scholar]
- Tisserat, B. Influence of Ultra-High Carbon Dioxide Levels on Growth and Morphogenesis of Lamiaceae Species in Soil. J. Herbs. Spices Med. Plants 2002, 9, 81–89. [Google Scholar] [CrossRef]
- Reinert, R.A.; Ho, M.C. Vegetative Growth of Soybean as Affected by Elevated Carbon Dioxide and Ozone. Environ. Pollut. 1995, 89, 89–96. [Google Scholar] [CrossRef]
- Mousseau, M. Effects of Elevated CO2 on Growth, Photosynthesis and Respiration of Sweet Chestnut (Castanea sativa Mill.). Vegetatio 1993, 104, 413–419. [Google Scholar] [CrossRef]
- Olivo, N.; Martinez, C.A.; Oliva, M.A. The Photosynthetic Response to Elevated CO2 in High Altitude Potato Species (Solanum curtilobum). Photosynthetica 2002, 40, 309–313. [Google Scholar]
- Mortensen, L.M. Nitrogen Oxides Produced during CO2 Enrichment: I. Effects on Different Greenhouse Plants. New Phytol. 1985, 101, 103–108. [Google Scholar] [CrossRef]
- Mortensen, L.M. Effect of Carbon Dioxide Concentration on Biomass Production and Partitioning in Betula Pubescens Ehrh. Seedlings at Different Ozone and Temperature Regimes. Environ. Pollut. 1995, 87, 337–343. [Google Scholar] [CrossRef]
- Van de Staaij, J.W.M.; Lenssen, G.M.; Stroetenga, M.; Rozema, J. The Combined Effects of Elevated CO2 Levels and UV-B Radiation on Growth Characteristics of Elymus Athericus (=E. pycnanathus). Vegetatio 1993, 104, 433–439. [Google Scholar] [CrossRef]
- Flores-Velázquez, J.; Rojano, F.; Aguilar-Rodríguez, C.E.; Villagran, E.; Villarreal-Guerrero, F. Greenhouse Thermal Effectiveness to Produce Tomatoes Assessed by a Temperature-Based Index. Agronomy 2022, 12, 1158. [Google Scholar] [CrossRef]
- Villagrán, E.; Flores-Velazquez, J.; Bojacá, C.; Akrami, M. Evaluation of the Microclimate in a Traditional Colombian Greenhouse Used for Cut Flower Production. Agronomy 2021, 11, 1330. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, J.; Duan, J.; Wang, M.; Dong, Z. Greenhouse CO2 Control Based on Improved Genetic Algorithm and Fuzzy Neural Network. In Proceedings of the 2018 2nd IEEE Advanced Information Management, Communicates, Electronic and Automation Control Conference (IMCEC), Xi’an, China, 25–27 May 2018; IEEE: New York, NY, USA, 2018; pp. 1537–1540. [Google Scholar]
- Zhang, R.; Xiang, M.; Li, P.; Bai, B. Decision-Making on Greenhouse CO2 Control Quantity Based on Information Fusion. Trans. Chin. Soc. Agric. Mach. 2009, 40, 175–178. [Google Scholar]
- Lee, D.H.; Lee, K.-S.; Choi, C.-H.; Kim, H.-J.; Chung, S.-O.; Cho, Y.-J. Prediction of CO2 Emission from Soil for Optimal Greenhouse Control. In Proceedings of the 2011 ASABE Annual International Meeting, Louisville, KY, USA, 7–10 August 2011; American Society of Agricultural and Biological Engineers: St. Joseph, MI, USA, 2011; p. 1. [Google Scholar] [CrossRef]
- Draoui, B.; Bounaama, F.; Boulard, T.; Bibi-Triki, N. In-Situ Modelisation of a Greenhouse Climate Including Sensible Heat, Water Vapour and CO2 Balances. In Proceedings of the EPJ Web of Conferences; EDP Sciences: les Ulis, France, 2013; Volume 45, p. 1023. [Google Scholar]
- Li, T.; Ji, Y.; Zhang, M.; Sha, S.; Li, M. Universality of an Improved Photosynthesis Prediction Model Based on PSO-SVM at All Growth Stages of Tomato. Int. J. Agric. Biol. Eng. 2017, 10, 63–73. [Google Scholar]
- Li, T.; Ji, Y.; Zhang, M.; Sha, S.; Jiang, Y. Tomato Photosynthetic Rate Prediction Models under Interaction of CO2 Enrichments and Soil Moistures. Trans. Chin. Soc. Agric. Mach. 2015, 46, 208–214. [Google Scholar]
- Bailey, B.J.; Chalabi, Z.S.; Aikman, D.P.; Cockshull, K.E. Improved Strategies for Controlling CO2 Enrichment in Tomato Greenhouses. Acta Hortic. 1996, 443, 155–162. [Google Scholar]
- Nurmalisa, M.; Tokairin, T.; Kumazaki, T.; Takayama, K.; Inoue, T. CO2 Distribution under CO2 Enrichment Using Computational Fluid Dynamics Considering Photosynthesis in a Tomato Greenhouse. Appl. Sci. 2022, 12, 7756. [Google Scholar] [CrossRef]
- Yildiz, I.; Stombaugh, D.P. Heat Pump Cooling and Greenhouse Microclimates in Open and Confined Greenhouse Systems. Acta Hortic. 2006, 719, 255–262. [Google Scholar]
- Zhang, Y.; Yasutake, D.; Hidaka, K.; Okayasu, T.; Kitano, M.; Hirota, T. Crop-Localised CO2 Enrichment Improves the Microclimate, Photosynthetic Distribution and Energy Utilisation Efficiency in a Greenhouse. J. Clean. Prod. 2022, 371, 133465. [Google Scholar]
- Kimura, K.; Yasutake, D.; Koikawa, K.; Kitano, M. Spatiotemporal Variability of Leaf Photosynthesis and Its Linkage with Microclimates across an Environment-Controlled Greenhouse. Biosyst. Eng. 2020, 195, 97–115. [Google Scholar] [CrossRef]
- Moon, T.; Choi, H.-Y.; Jung, D.-H.; Chang, S.-H.; Son, J.-E. Prediction of CO2 Concentration via Long Short-Term Memory Using Environmental Factors in Greenhouses. Korean J. Hortic. Sci. Technol. 2020, 38, 201–209. [Google Scholar]
- Kim, S.-H.; Fisher, P.R.; Lieth, J.H. Analysis and Modeling of Gas Exchange Processes in Scaevola Aemula. Sci. Hortic. 2007, 114, 170–176. [Google Scholar] [CrossRef]
- Jung, D.H.; Hwang, I.; Shin, J.; Son, J.E. Analysis of Leaf Photosynthetic Rates of Hydroponically-Grown Paprika (Capsicum Annuum L.) Plants According to Vertical Position with Multivariable Photosynthesis Models. Hortic. Environ. Biotechnol. 2021, 62, 41–51. [Google Scholar] [CrossRef]
- Shin, J.; Hwang, I.; Kim, D.; Kim, J.; Kim, J.H.; Son, J.E. Waning Advantages of CO2 Enrichment on Photosynthesis and Productivity Due to Accelerated Phase Transition and Source-Sink Imbalance in Sweet Pepper. Sci. Hortic. 2022, 301, 111130. [Google Scholar]
- Effat, M.B.; Shafey, H.M.; Nassib, A.M. Solar Greenhouses Can Be Promising Candidate for CO 2 Capture and Utilization: Mathematical Modeling. Int. J. Energy Environ. Eng. 2015, 6, 295–308. [Google Scholar]
- Cooper, P.I.; Fuller, R.J. A Transient Model of the Interaction between Crop, Environment and Greenhouse Structure for Predicting Crop Yield and Energy Consumption. J. Agric. Eng. Res. 1983, 28, 401–417. [Google Scholar] [CrossRef]
- Jin, C.; Du, S.; Wang, Y.; Condon, J.; Lin, X.; Zhang, Y. Carbon Dioxide Enrichment by Composting in Greenhouses and Its Effect on Vegetable Production. J. Plant Nutr. Soil Sci. 2009, 172, 418–424. [Google Scholar] [CrossRef]
- Liu, L.; Sun, C.; He, X.; Liu, X.; Wu, H.; Liu, M.; Tang, C.; Zhang, Y. The Secondary Compost Products Enhances Soil Suppressive Capacity against Bacterial Wilt of Tomato Caused by Ralstonia Solanacearum. Eur. J. Soil Biol. 2016, 75, 70–78. [Google Scholar] [CrossRef]
- Karim, M.F.; Hao, P.; Nordin, N.H.B.; Qiu, C.; Zeeshan, M.; Khan, A.A.; Wu, F.; Shamsi, I.H. CO2 Enrichment Using CRAM Fermentation Improves Growth, Physiological Traits and Yield of Cherry Tomato (Solanum lycopersicum L.). Saudi J. Biol. Sci. 2020, 27, 1041–1048. [Google Scholar] [CrossRef]
- Poudel, M.; Dunn, B. Greenhouse Carbon Dioxide Supplementation [Superseded]; Oklahoma State University: Stillwater, OK, USA, 2017. [Google Scholar]
- Sun, Q.; Cui, S.; Song, Y.; Yang, Z.; Dong, Q.; Sun, S.; Wu, H. Impact of Illumination and Temperature Performance of Blanket-inside Solar Greenhouse and CO2 Enrichment on Cucumber Growth and Development. Agric. Sci. Technol. 2016, 17, 1757. [Google Scholar]
- Chalabi, Z.S.; Biro, A.; Bailey, B.J.; Aikman, D.P.; Cockshull, K.E. Optimal Control Strategies for Carbon Dioxide Enrichment in Greenhouse Tomato Crops. Part II: Using the Exhaust Gases of Natural Gas Fired Boilers. Biosyst. Eng. 2002, 81, 323–332. [Google Scholar] [CrossRef]
- Moreno, J.V.R.; Pinna-Hernández, M.G.; Molina, J.A.S.; Fernández, M.D.F.; Hernández, J.C.L.; Fernández, F.G.A. Carbon Capture from Biomass Flue Gases for CO2 Enrichment in Greenhouses. Heliyon 2024, 10, e23274. [Google Scholar] [CrossRef]
- Umeda, H.; Ahn, D.-H.; Iwasaki, Y.; Matsuo, S.; Takeya, S. A Cooling and CO2 Enrichment System for Greenhouse Production Using CO2 Clathrate Hydrate. Eng. Agric. Environ. Food 2015, 8, 307–312. [Google Scholar] [CrossRef]
- Jaffrin, A.; Bentounes, N.; Joan, A.M.; Makhlouf, S. Landfill Biogas for Heating Greenhouses and Providing Carbon Dioxide Supplement for Plant Growth. Biosyst. Eng. 2003, 86, 113–123. [Google Scholar] [CrossRef]
- Zhang, S.; Bi, X.T.; Clift, R. Life Cycle Analysis of a Biogas-Centred Integrated Dairy Farm-Greenhouse System in British Columbia. Process Saf. Environ. Prot. 2015, 93, 18–30. [Google Scholar] [CrossRef]
- Roy, Y.; Lefsrud, M.; Orsat, V.; Filion, F.; Bouchard, J.; Nguyen, Q.; Dion, L.-M.; Glover, A.; Madadian, E.; Lee, C.P. Biomass Combustion for Greenhouse Carbon Dioxide Enrichment. Biomass Bioenergy 2014, 66, 186–196. [Google Scholar] [CrossRef]
- Snchez-Molina, J.A.; Group, J.V.R.M.M.B.R.; Group, F.G.A.M.M.B.R. Development of a Biomass-Based System for Nocturnal Temperature and Diurnal CO2 Concentration Control in Greenhouses. Biomass Bioenergy 2015, 67, 60–71. [Google Scholar] [CrossRef]
- Monteroa, J.I.; Stanghellini, C.; Castilla, N. Greenhouse Technology for Sustainable Production in Mild Winter Climate Areas: Trends and Needs. Acta Hortic. 2009, 807, 33–44. [Google Scholar] [CrossRef]
- Stanghellini, C.; Incrocci, L.; Gázquez, J.C.; Dimauro, B. Carbon Dioxide Concentration in Mediterranean Greenhouses: How Much Lost Production? Acta Hortic. 2007, 801, 1541–1550. [Google Scholar] [CrossRef]
- Villagrán, E.A.; Bojacá, C.R. Effects of Surrounding Objects on the Thermal Performance of Passively Ventilated Greenhouses. J. Agric. Eng. 2019, 856, 20–27. [Google Scholar] [CrossRef]
- Chalabi, Z.S.; Biro, A.; Bailey, B.J.; Aikman, D.P.; Cockshull, K.E. ERRATUM-Optimal Control Strategies for Carbon Dioxide Enrichment in Greenhouse Tomato Crops Part 1: Using Pure Carbon Dioxide. Biosyst. Eng. 2002, 1, 127. [Google Scholar]
- Kuroyanagi, T.; Yasuba, K.; Higashide, T.; Iwasaki, Y.; Takaichi, M. Efficiency of Carbon Dioxide Enrichment in an Unventilated Greenhouse. Biosyst. Eng. 2014, 119, 58–68. [Google Scholar] [CrossRef]
- Sánchez-Guerrero, M.C.; Lorenzo, P.; Medrano, E.; Castilla, N.; Soriano, T.; Baille, A. Effect of Variable CO2 Enrichment on Greenhouse Production in Mild Winter Climates. Agric. For. Meteorol. 2005, 132, 244–252. [Google Scholar] [CrossRef]
- Qasem, N.A.A.; Ben-Mansour, R. Assessment of Appropriate Geometry for Thermally Efficient CO2 Adsorption Beds. Appl. Sci. 2022, 12, 5726. [Google Scholar] [CrossRef]
- Qasem, N.A.A. Physical Adsorption of Carbon Dioxide Capture by Novel Materials. Ph.D. Thesis, King Fahd University of Petroleum and Minerals, Dhahran, Saudi Arabia, 2017. [Google Scholar]
- Herraiz, L.; Palfi, E.; Sánchez Fernández, E.; Lucquiaud, M. Rotary Adsorption: Selective Recycling of CO2 in Combined Cycle Gas Turbine Power Plants. Front. Energy Res. 2020, 8, 482708. [Google Scholar] [CrossRef]
- Tang, C.; Gao, X.; Shao, Y.; Wang, L.; Liu, K.; Gao, R.; Che, D. Investigation on the Rotary Regenerative Adsorption Wheel in a New Strategy for CO2 Enrichment in Greenhouse. Appl. Therm. Eng. 2022, 205, 118043. [Google Scholar]
- Bao, J.; Zhao, J.; Bi, X.T. CO2 Adsorption and Desorption for CO2 Enrichment at Low-Concentrations Using Zeolite 13X. Chemie Ing. Tech. 2023, 95, 143–150. [Google Scholar]
- Kozai, T.; Kubota, C.; Takagaki, M.; Maruo, T. Greenhouse Environment Control Technologies for Improving the Sustainability of Food Production. Acta Hortic. 2014, 1107, 1–14. [Google Scholar]
- Boulard, T.; Roy, J.-C.; Pouillard, J.-B.; Fatnassi, H.; Grisey, A. Modelling of Micrometeorology, Canopy Transpiration and Photosynthesis in a Closed Greenhouse Using Computational Fluid Dynamics. Biosyst. Eng. 2017, 158, 110–133. [Google Scholar]
- Xu, Z.; Jiang, Y.; Zhou, G. Response and Adaptation of Photosynthesis, Respiration, and Antioxidant Systems to Elevated CO2 with Environmental Stress in Plants. Front. Plant Sci. 2015, 6, 701. [Google Scholar]
- Baker, J.T.; Lascano, R.J.; Yates, C.; Gitz III, D.C. Nighttime CO2 Enrichment Did Not Increase Leaf Area or Shoot Biomass in Cotton Seedlings. Agric. For. Meteorol. 2022, 320, 108931. [Google Scholar]
- Vermeulen, P. Alternative Sources of CO2 for the Greenhouse Horticulture. In Proceedings of the Proc. 2nd Int. Symposium Energy Challenges Mechanics (ISECM2), Brisbane, Australia, 17 August 2014; pp. 19–21. [Google Scholar]
- Dong, J.; Gruda, N.; Lam, S.K.; Li, X.; Duan, Z. Effects of Elevated CO2 on Nutritional Quality of Vegetables: A Review. Front. Plant Sci. 2018, 9, 924. [Google Scholar]
- Neethirajan, S.; Jayas, D.S.; Sadistap, S. Carbon Dioxide (CO2) Sensors for the Agri-Food Industry—A Review. Food Bioprocess Technol. 2009, 2, 115–121. [Google Scholar]
- Wang, J.; Niu, X.; Zheng, L.; Zheng, C.; Wang, Y. Wireless Mid-Infrared Spectroscopy Sensor Network for Automatic Carbon Dioxide Fertilization in a Greenhouse Environment. Sensors 2016, 16, 1941. [Google Scholar] [CrossRef]
- Ali, A.; Shah, G.A.; Farooq, M.O.; Ghani, U. Technologies and Challenges in Developing Machine-to-Machine Applications: A Survey. J. Netw. Comput. Appl. 2017, 83, 124–139. [Google Scholar]
- Kim, H.T.; Kim, C.H.; Choi, E.G.; Jin, B.O.; Yoon, Y.C.; Kim, H.T. The Effect of Thermal Conditions on CO2 Distribution in a Greenhouse. Trop. Agric. Res. 2015, 26. [Google Scholar] [CrossRef]
- Demirbas, M.; Arora, A.; Mittal, V.; Kulathumani, V. A Fault-Local Self-Stabilizing Clustering Service for Wireless Ad Hoc Networks. IEEE Trans. Parallel Distrib. Syst. 2006, 17, 912–922. [Google Scholar]
- Taylor, C.J.; Aerts, J.-M. Control of Nonlinear Biological Systems by Non-Minimal State Variable Feedback. Stat. Biosci. 2014, 6, 290–313. [Google Scholar]
- Pasgianos, G.D.; Arvanitis, K.G.; Polycarpou, P.; Sigrimis, N. A Nonlinear Feedback Technique for Greenhouse Environmental Control. Comput. Electron. Agric. 2003, 40, 153–177. [Google Scholar]
- Graamans, L.; Baeza, E.; van den Dobbelsteen, A.; Tsafaras, I.; Stanghellini, C. Plant Factories versus Greenhouses: Comparison of Resource Use Efficiency. Agric. Syst. 2018. [Google Scholar] [CrossRef]
- Gu, X.; Goto, E. Evaluation of Plant Canopy Microclimates with Realistic Plants in Plant Factories with Artificial Light Using a Computational Fluid Dynamics Model. Build. Environ. 2024, 264, 111876. [Google Scholar]
- Duggan-Jones, D.I.; Nichols, M.A. Determining the Optimum Temperature, Irradiation and CO2 Enrichment on the Growth of Lettuce and Cabbage Seedlings in Plant Factories. In Proceedings of the XXIX International Horticultural Congress on Horticulture: Sustaining Lives, Livelihoods and Landscapes (IHC2014), Brisbane, Australia, 17 August 2014; Volume 1107, pp. 187–194. [Google Scholar]
- Kitaya, Y.; Shibuya, T.; Yoshida, M.; Kiyota, M. Effects of Air Velocity on Photosynthesis of Plant Canopies under Elevated CO2 Levels in a Plant Culture System. Adv. Sp. Res. 2004, 34, 1466–1469. [Google Scholar]
- Perone, C.; Orsino, M.; La Fianza, G.; Brunetti, L.; Giametta, F.; Catalano, P. CO2 Use and Energy Efficiency in Closed Plant Production System by Means of Mini-Air Handling Unit. In Proceedings of the International Conference on Safety, Health and Welfare in Agriculture and Agro-food Systems; Springer: Berlin/Heidelberg, Germany, 2020; pp. 506–515. [Google Scholar]
- Shiina, T.; Hosokawa, D.; Roy, P.; Nakamura, N.; Thammawong, M.; Orikasa, T. Life Cycle Inventory Analysis of Leafy Vegetables Grown in Two Types of Plant Factories. In Proceedings of the XXVIII International Horticultural Congress on Science and Horticulture for People (IHC2010): International Symposium on 919, Lisbon, Portugal, 22–27 August 2010; pp. 115–122. [Google Scholar]
- Becker, C.; Kläring, H.-P. CO2 Enrichment Can Produce High Red Leaf Lettuce Yield While Increasing Most Flavonoid Glycoside and Some Caffeic Acid Derivative Concentrations. Food Chem. 2016, 199, 736–745. [Google Scholar]
- Chowdhury, M.; Kiraga, S.; Islam, M.N.; Ali, M.; Reza, M.N.; Lee, W.-H.; Chung, S.-O. Effects of Temperature, Relative Humidity, and Carbon Dioxide Concentration on Growth and Glucosinolate Content of Kale Grown in a Plant Factory. Foods 2021, 10, 1524. [Google Scholar] [CrossRef]
- Lee, Y.B.; Park, M.H. Effects of CO2 Concentration, Light Intensity and Nutrient Level on Growth of Leaf Lettuce in a Plant Factory. In Proceedings of the International Symposium on Growing Media and Hydroponics 548, Macedonia, Greece, 31 August–6 September 1999; pp. 377–384. [Google Scholar]
- Ikeda, A.; Nakayama, S.; Kitaya, Y.; Yabuki, K. Effects of Photoperiod, CO2 Concentration, and Light Intensity on Growth and Net Photosynthetic Rates of Lettuce and Turnip. Acta Hortic. 1987, 229, 273–282. [Google Scholar]
- Kennebeck, E.J.; Meng, Q. Mustard ‘Amara’Benefits from Superelevated CO2 While Adapting to Far-Red Light Over Time. HortScience 2024, 59, 139–145. [Google Scholar] [CrossRef]
- Kubota, C. Concepts and Background of Photoautotrophic Micropropagation. In Progress in Biotechnology; Elsevier: Amsterdam, The Netherlands, 2001; Volume 18, pp. 325–334. ISBN 0921-0423. [Google Scholar]
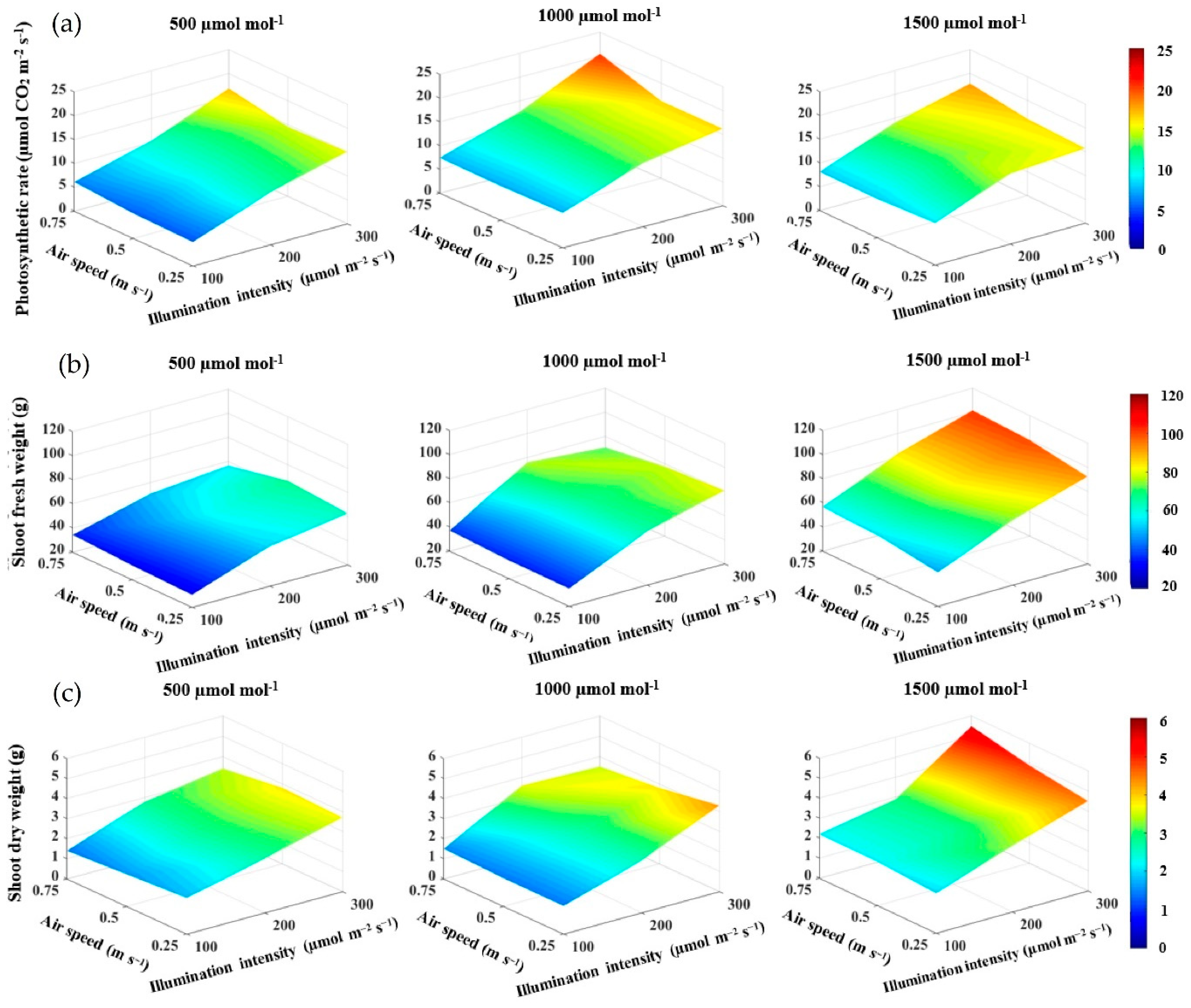
- Ahmed, H.A.; Tong, Y.; Li, L.; Sahari, S.Q.; Almogahed, A.M.; Cheng, R. Integrative Effects of CO2 Concentration, Illumination Intensity and Air Speed on the Growth, Gas Exchange and Light Use Efficiency of Lettuce Plants Grown under Artificial Lighting. Horticulturae 2022, 8, 270. [Google Scholar] [CrossRef]
- Jung, D.H.; Kim, D.; Yoon, H.I.; Moon, T.W.; Park, K.S.; Son, J.E. Modeling the Canopy Photosynthetic Rate of Romaine Lettuce (Lactuca sativa L.) Grown in a Plant Factory at Varying CO 2 Concentrations and Growth Stages. Hortic. Environ. Biotechnol. 2016, 57, 487–492. [Google Scholar] [CrossRef]
- Tsafaras, I.; de Zwart, H.F.; Voogt, W.; Campen, J.B.; van der Heide, H.; Al Harbi, A.; Al Assaf, K.; Qaryouti, M.; Abdelaziz, M.E. Light Use Efficiency of Various Greenhouse Systems in Arid Environment. Acta Hortic. 2023, 1377, 283–290. [Google Scholar] [CrossRef]
- Calvet, J.-C.; Soussana, J.-F. Modelling CO2-Enrichment Effects Using an Interactive Vegetation SVAT Scheme. Agric. For. Meteorol. 2001, 108, 129–152. [Google Scholar] [CrossRef]
- Chen, W.-H.; You, F. MPC for the Indoor Climate Control and Energy Optimization of a Building-Integrated Rooftop Greenhouse Systems. IFAC-Pap. 2024, 58, 164–169. [Google Scholar] [CrossRef]
- Martzopoulou, A.; Vafiadis, D.; Fragos, V.P. Relative Humidity Variation in Greenhouses as Affected by CO2 Enrichment at High Temperatures Resulting in Energy Saving. Acta Hortic. 2020, 1296, 141–148. [Google Scholar] [CrossRef]
- Deckmyn, G.; Caeyenberghs, E.; Ceulemans, R. Reduced UV-B in Greenhouses Decreases White Clover Response to Enhance CO2. Environ. Exp. Bot. 2001, 46, 109–117. [Google Scholar] [CrossRef]
- Villagrán, E.; Romero-Perdomo, F.; Numa-Vergel, S.; Galindo-Pacheco, J.R.; Salinas-Velandia, D.A. Life Cycle Assessment in Protected Agriculture: Where Are We Now, and Where Should We Go Next? Horticulturae 2024, 10, 15. [Google Scholar] [CrossRef]
- Gruda, N.; Bisbis, M.; Tanny, J. Impacts of Protected Vegetable Cultivation on Climate Change and Adaptation Strategies for Cleaner Production—A Review. J. Clean. Prod. 2019, 225, 324–339. [Google Scholar] [CrossRef]
- Weidner, T.; Yang, A.; Forster, F.; Hamm, M.W. Regional Conditions Shape the Food–Energy–Land Nexus of Low-Carbon Indoor Farming. Nat. Food 2022, 3, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Song, G.; Liu, X.; Batlle-Bayer, L.; Fullana-i-Palmer, P. Life Cycle Climate Performance of Urban Plant Factory versus Rural Greenhouse under China’s Power-Grid Decarbonization: Considering Short-Lived Methane and Nitrous Oxide Emissions. Carbon Footprints 2024, 3, 10. [Google Scholar]
- Wang, X.; Dou, Z.; Shi, X.; Zou, C.; Liu, D.; Wang, Z.; Guan, X.; Sun, Y.; Wu, G.; Zhang, B. Innovative Management Programme Reduces Environmental Impacts in Chinese Vegetable Production. Nat. Food 2021, 2, 47–53. [Google Scholar]
- Gong, W.; Wang, C.; Fan, Z.; Xu, Y. Drivers of the Peaking and Decoupling between CO2 Emissions and Economic Growth around 2030 in China. Environ. Sci. Pollut. Res. 2022, 29, 3864–3878. [Google Scholar]
- Alvarez, R.A.; Pacala, S.W.; Winebrake, J.J.; Chameides, W.L.; Hamburg, S.P. Greater Focus Needed on Methane Leakage from Natural Gas Infrastructure. Proc. Natl. Acad. Sci. 2012, 109, 6435–6440. [Google Scholar] [CrossRef] [PubMed]
- Eaton, M.; Shelford, T.; Cole, M.; Mattson, N. Modeling Resource Consumption and Carbon Emissions Associated with Lettuce Production in Plant Factories. J. Clean. Prod. 2023, 384, 135569. [Google Scholar]
- Stacey, N.; Fox, J.; Hildebrandt, D. Reduction in Greenhouse Water Usage through Inlet CO2 Enrichment. AIChE J. 2018, 64, 2324–2328. [Google Scholar]
- Frantz, J.M. Elevating Carbon Dioxide in a Commercial Greenhouse Reduced Overall Fuel Carbon Consumption and Production Cost When Used in Combination with Cool Temperatures for Lettuce Production. Horttechnology 2011, 21, 647–651. [Google Scholar]
- Gavazzi, M.; Seiler, J.; Aust, W.; Zedaker, S. The Influence of Elevated Carbon Dioxide and Water Availability on Herbaceous Weed Development and Growth of Transplanted Loblolly Pine (Pinus Taeda). Environ. Exp. Bot. 2000, 44, 185–194. [Google Scholar]
- Suzuki, K.; Yamakawa, T.; Itoh, S.; Ohishi, N.; Kiriiwa, Y. Determination of Fossil-Derived Carbon Ratio of Tomato Plants in Greenhouse with CO2 Enrichment Using Biobased Content Method. Sci. Hortic. 2024, 327, 112818. [Google Scholar] [CrossRef]
- Zhang, M.; Li, T.; Ji, Y.; Sha, S. Optimization of CO2 Enrichment Strategy Based on BPNN for Tomato Plants in Greenhouse. Trans. Chinese Soc. Agric. Mach. 2015, 46, 239–245. [Google Scholar]
- Yi, Z.; Li, S.; Liang, Y.; Zhao, H.; Hou, L.; Yu, S.; Ahammed, G.J. Effects of Exogenous Spermidine and Elevated CO 2 on Physiological and Biochemical Changes in Tomato Plants under Iso-Osmotic Salt Stress. J. Plant Growth Regul. 2018, 37, 1222–1234. [Google Scholar] [CrossRef]
- Ghiat, I.; Mahmood, F.; Govindan, R.; Al-Ansari, T. CO2 Utilisation in Agricultural Greenhouses: A Novel ‘Plant to Plant’Approach Driven by Bioenergy with Carbon Capture Systems within the Energy, Water and Food Nexus. Energy Convers. Manag. 2021, 228, 113668. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, T.; Ma, J. Plant Factory: A New Method for Reducing Carbon Emissions. In Proceedings of the AIP Conference Proceedings; AIP Publishing: Melville, NY, USA, 2017; Volume 1820. [Google Scholar]
- Chen, H.; Dong, X.; Lei, J.; Zhang, N.; Wang, Q.; Shi, Z.; Yang, J. Life Cycle Assessment of Carbon Capture by an Intelligent Vertical Plant Factory within an Industrial Park. Sustainability 2024, 16, 697. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villagran, E.; Espitia, J.J.; Amado, G.; Rodriguez, J.; Gomez, L.; Velasquez, J.F.; Gil, R.; Baeza, E.; Aguilar, C.E.; Akrami, M.; et al. CO2 Enrichment in Protected Agriculture: A Systematic Review of Greenhouses, Controlled Environment Systems, and Vertical Farms—Part 2. Sustainability 2025, 17, 2809. https://doi.org/10.3390/su17072809
Villagran E, Espitia JJ, Amado G, Rodriguez J, Gomez L, Velasquez JF, Gil R, Baeza E, Aguilar CE, Akrami M, et al. CO2 Enrichment in Protected Agriculture: A Systematic Review of Greenhouses, Controlled Environment Systems, and Vertical Farms—Part 2. Sustainability. 2025; 17(7):2809. https://doi.org/10.3390/su17072809
Chicago/Turabian StyleVillagran, Edwin, John Javier Espitia, Gina Amado, Jader Rodriguez, Luisa Gomez, Jorge Flores Velasquez, Rodrigo Gil, Esteban Baeza, Cruz Ernesto Aguilar, Mohammad Akrami, and et al. 2025. "CO2 Enrichment in Protected Agriculture: A Systematic Review of Greenhouses, Controlled Environment Systems, and Vertical Farms—Part 2" Sustainability 17, no. 7: 2809. https://doi.org/10.3390/su17072809
APA StyleVillagran, E., Espitia, J. J., Amado, G., Rodriguez, J., Gomez, L., Velasquez, J. F., Gil, R., Baeza, E., Aguilar, C. E., Akrami, M., & Arias, L. A. (2025). CO2 Enrichment in Protected Agriculture: A Systematic Review of Greenhouses, Controlled Environment Systems, and Vertical Farms—Part 2. Sustainability, 17(7), 2809. https://doi.org/10.3390/su17072809