
How Much Can We See from a UAV-Mounted Regular Camera? Remote Sensing-Based Estimation of Forest Attributes in South American Native Forests

 , ,
, , 
Abstract
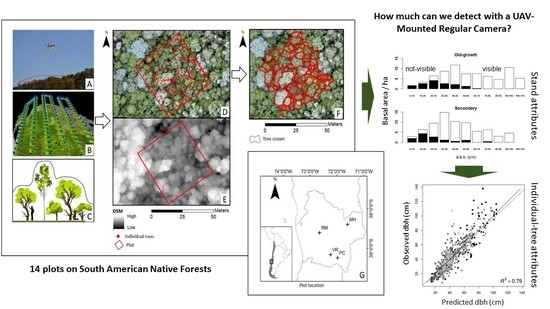
:
1. Introduction
2. Materials and Methods
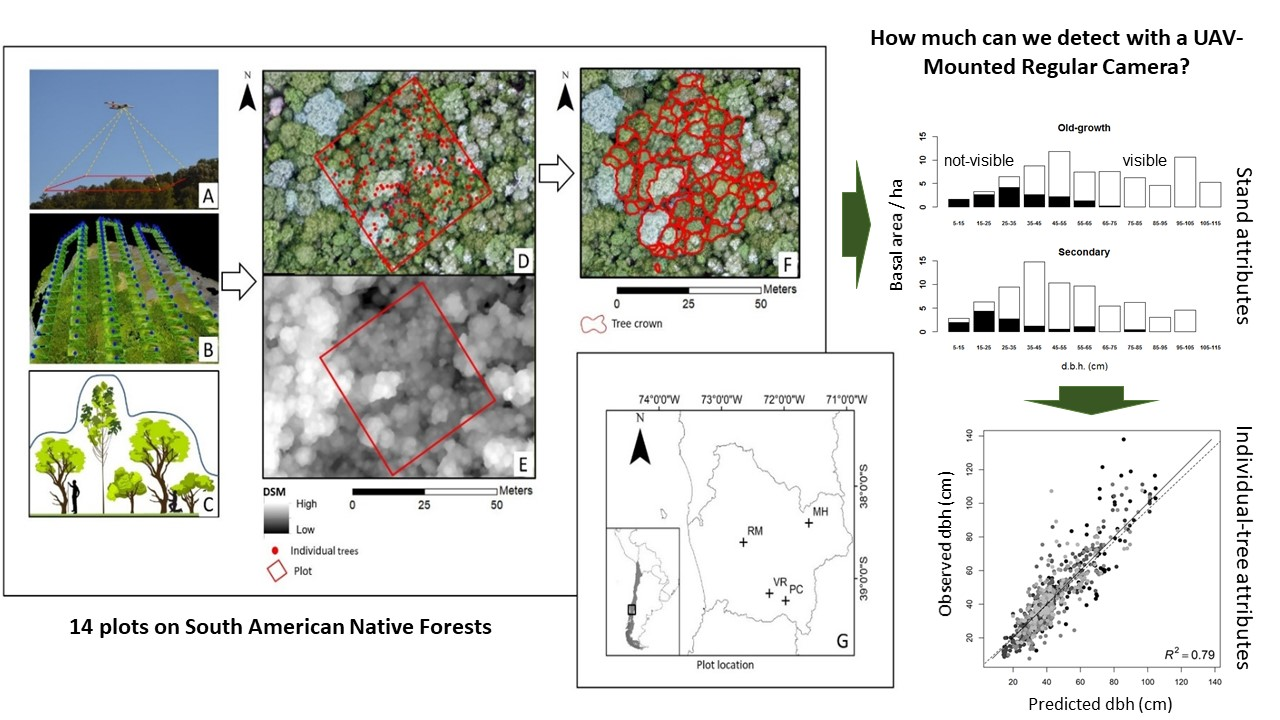
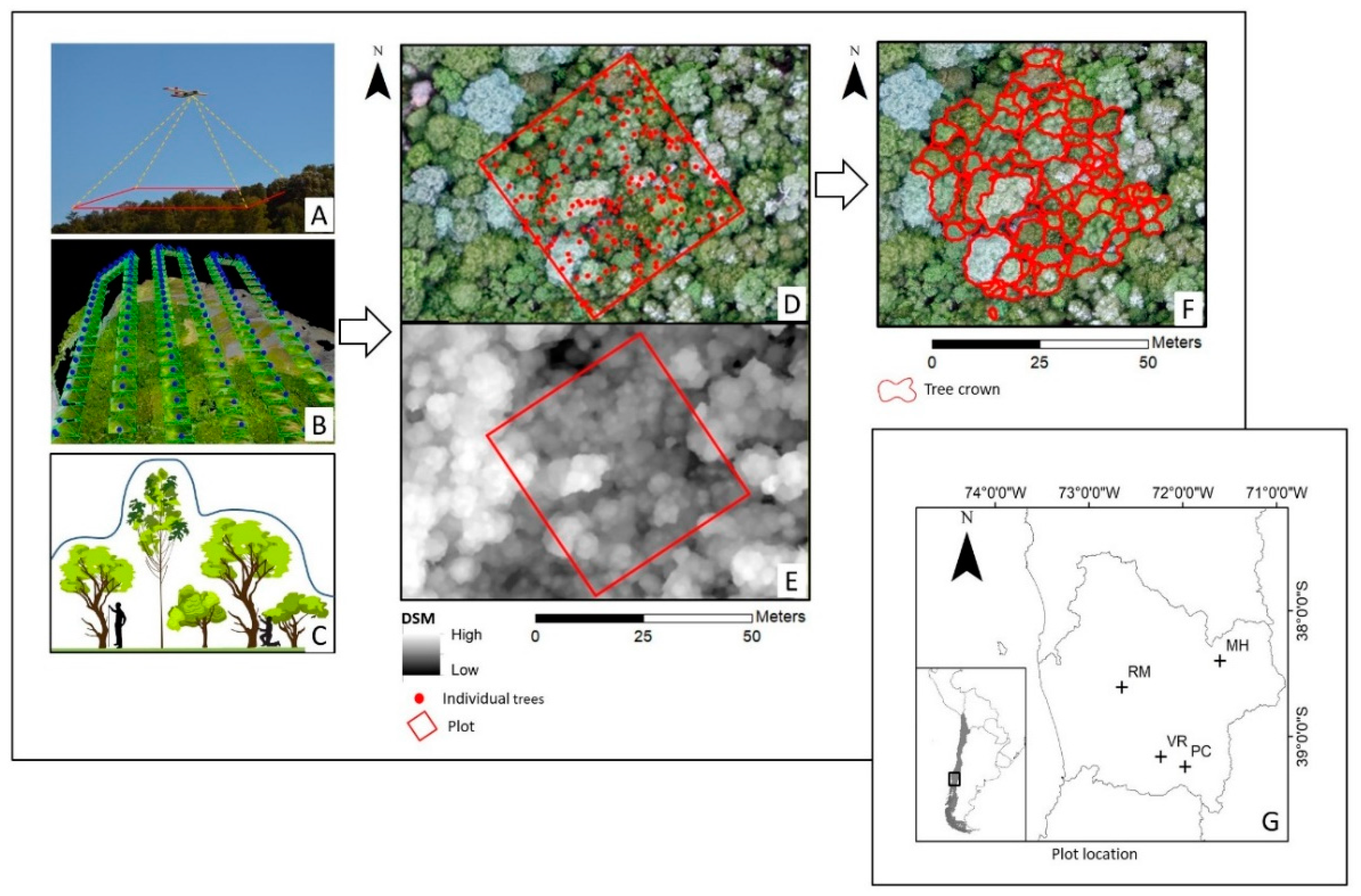
2.1. Study Sites
2.2. UAV Data Acquisition and Image Processing
2.3. Statistical Analysis
3. Results
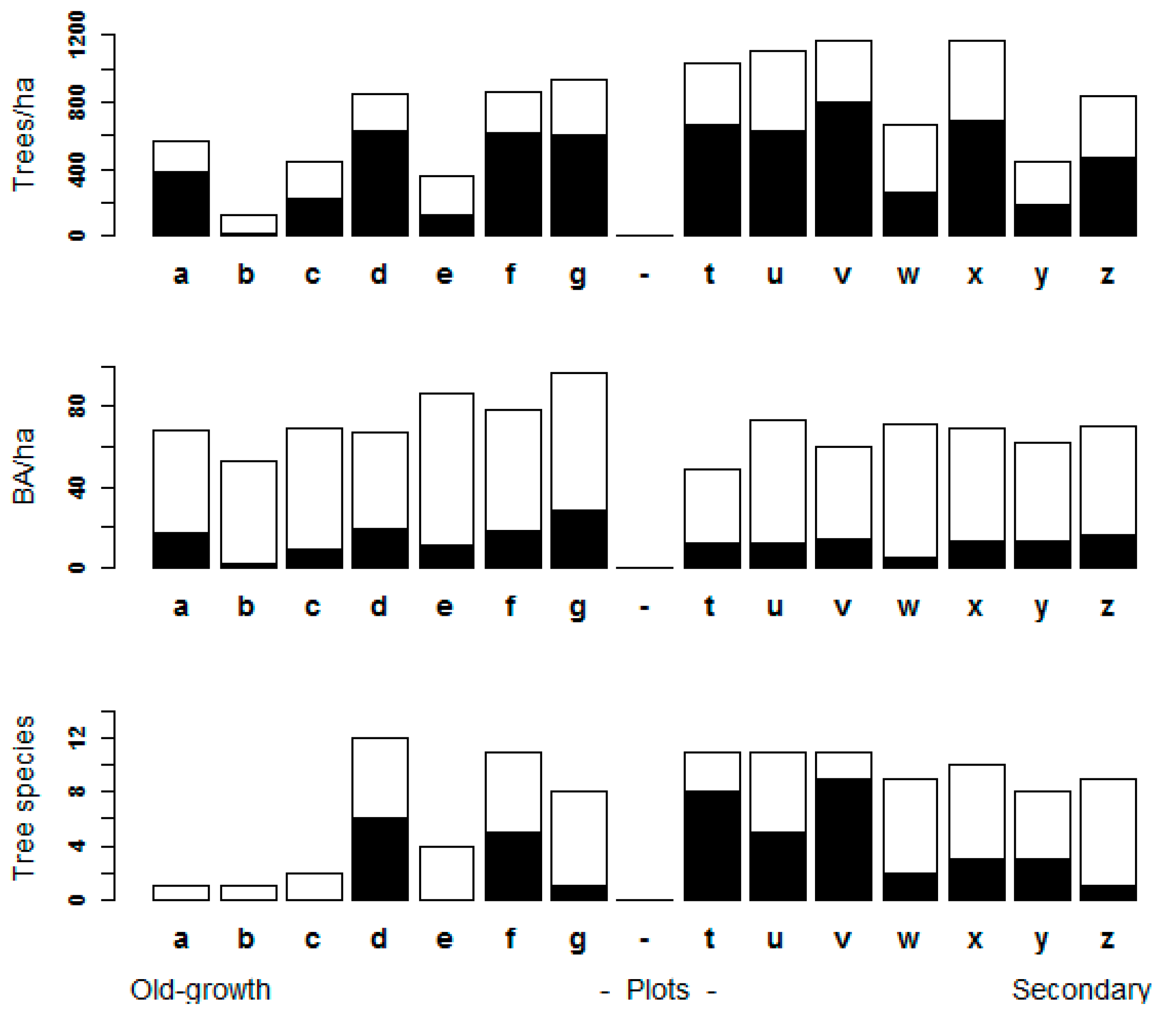
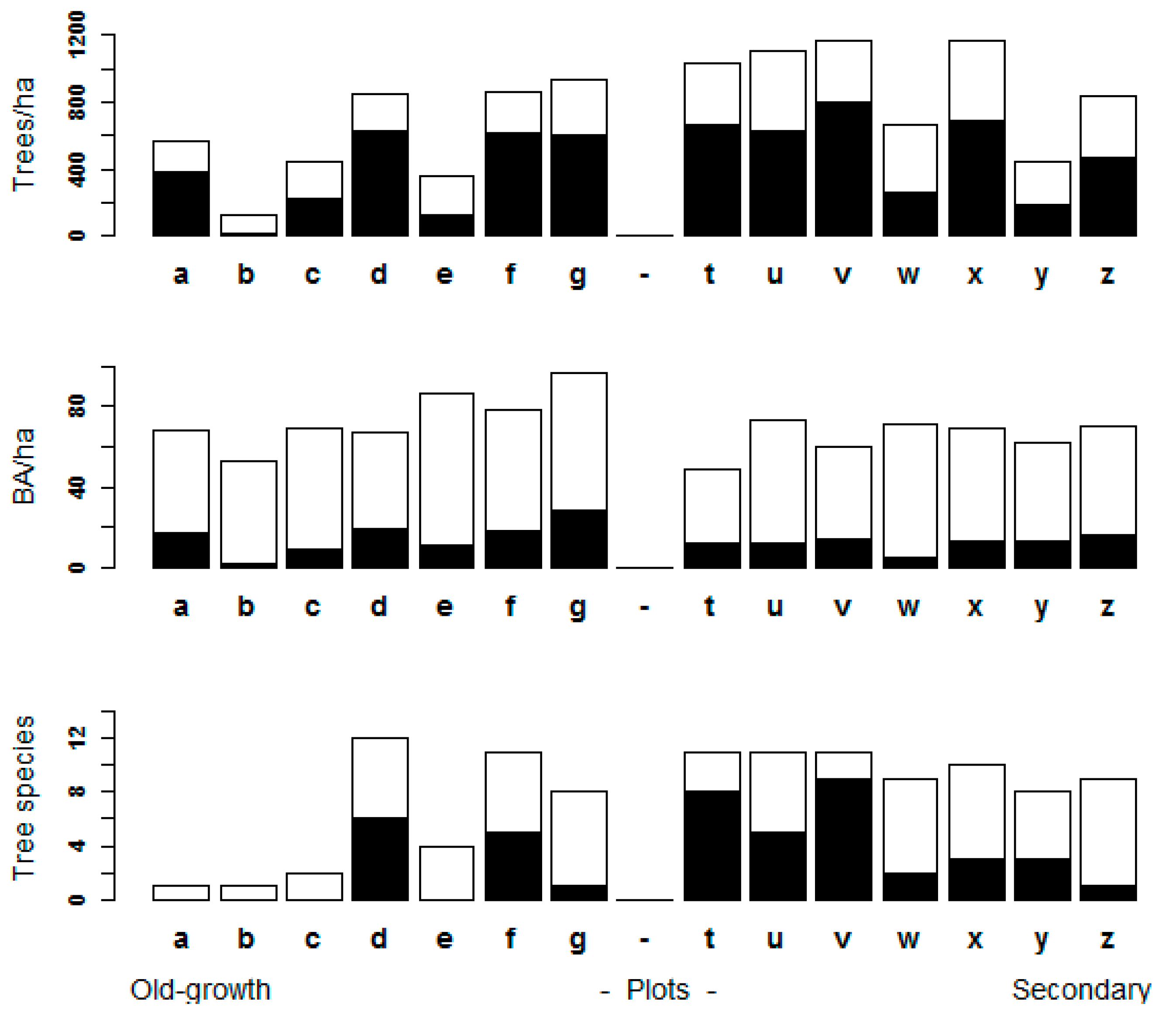
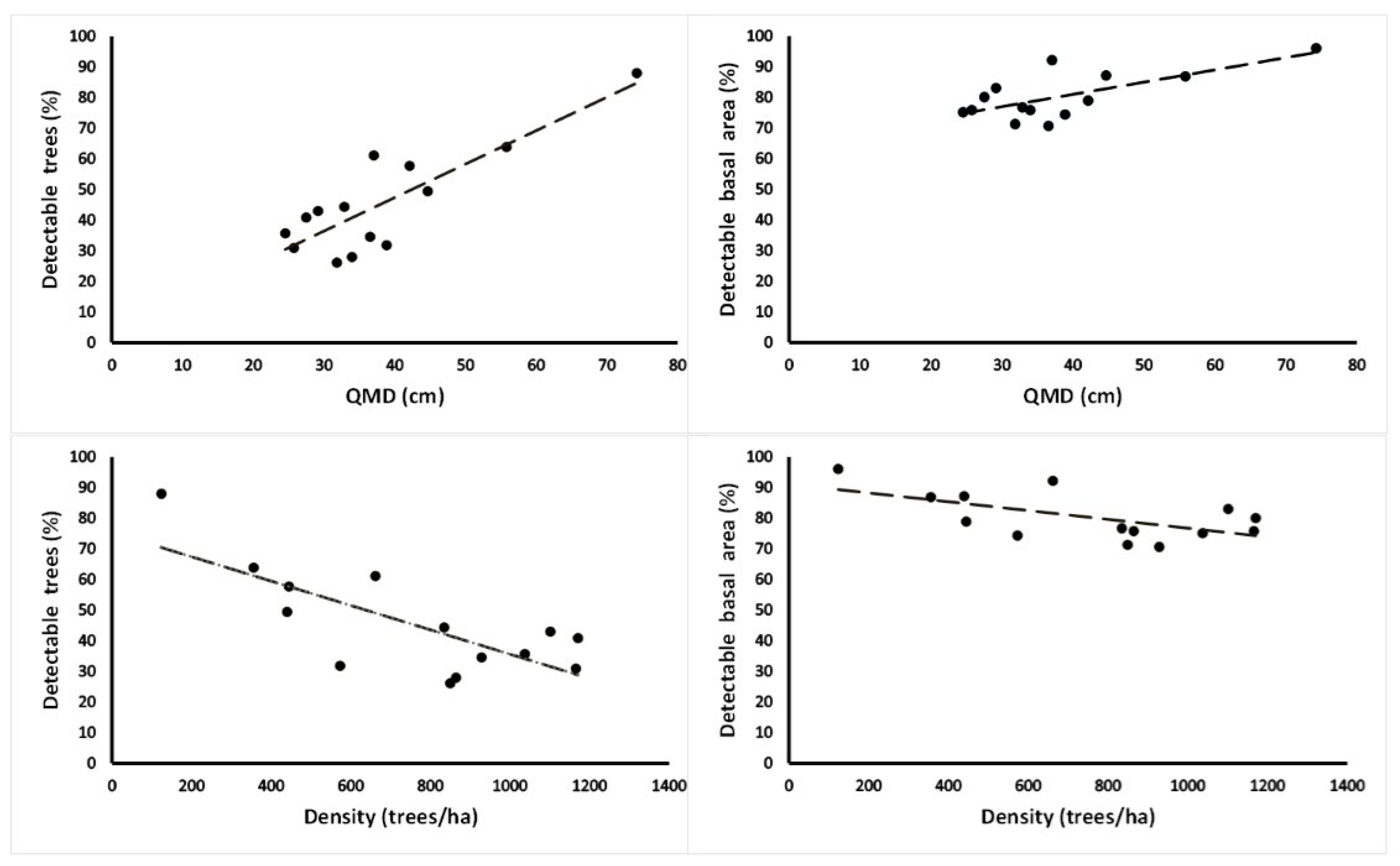
3.1. Assessment of Forest Attributes Observed from a UAV in Old-Growth and Secondary Forests
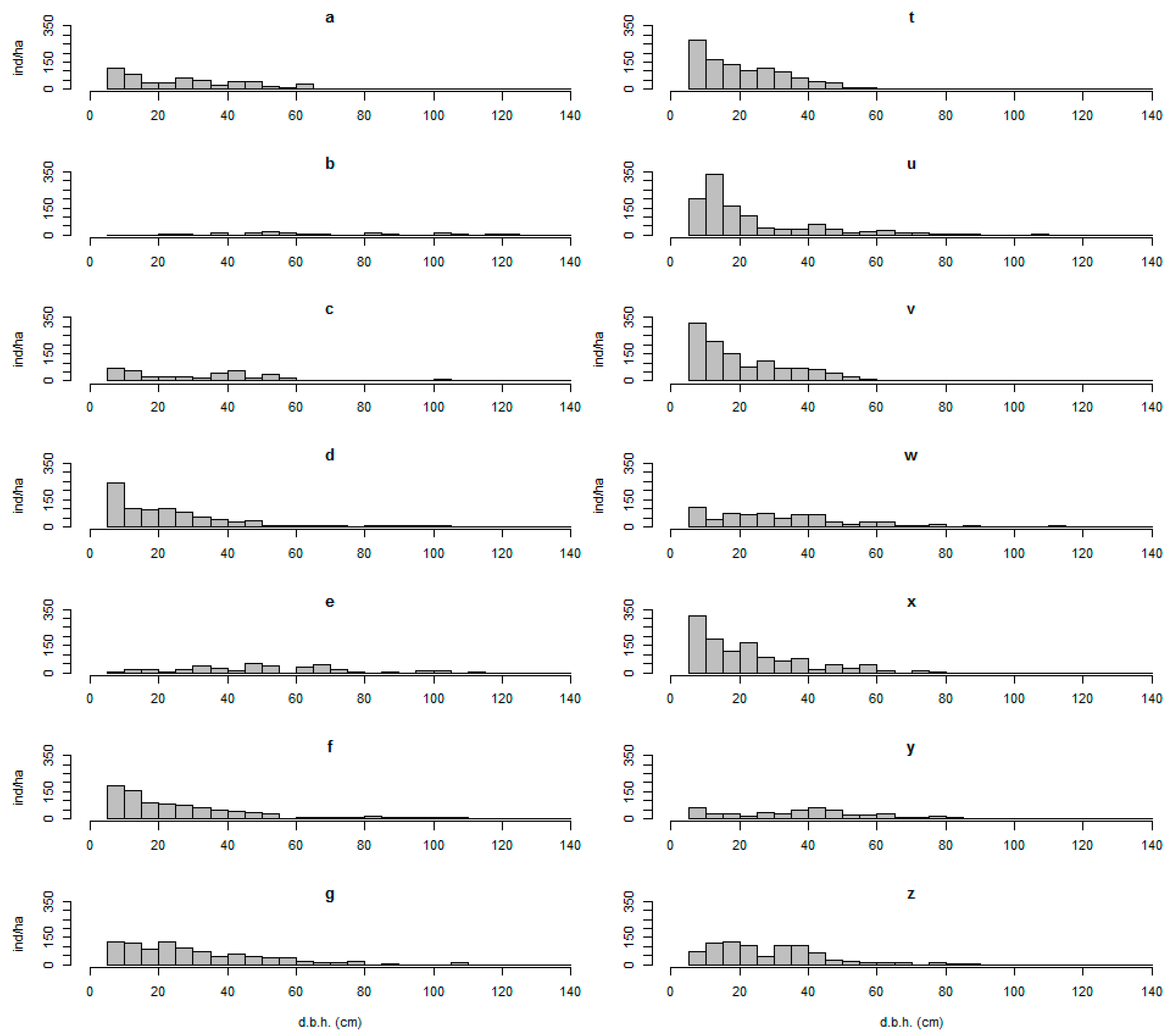
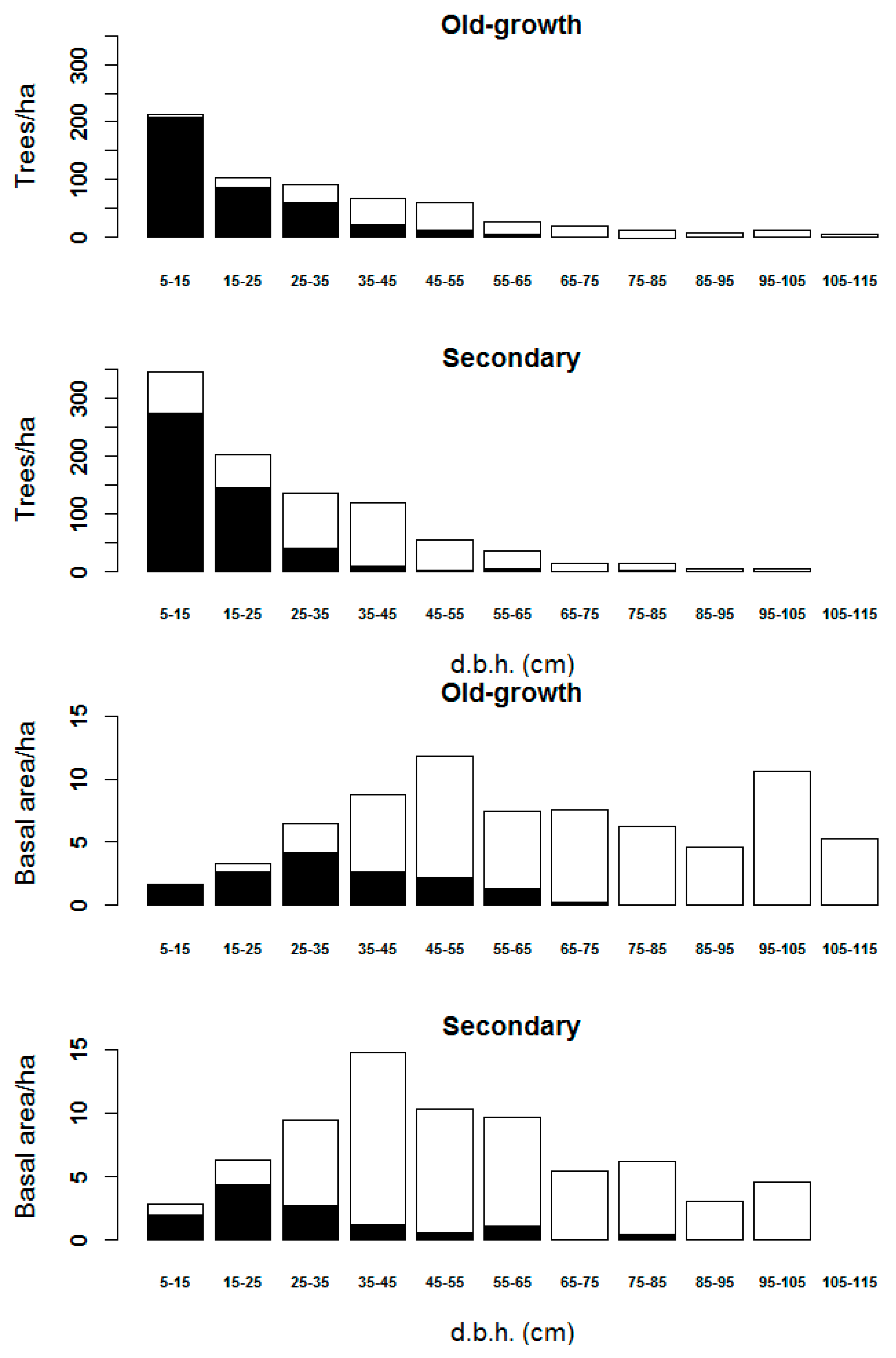
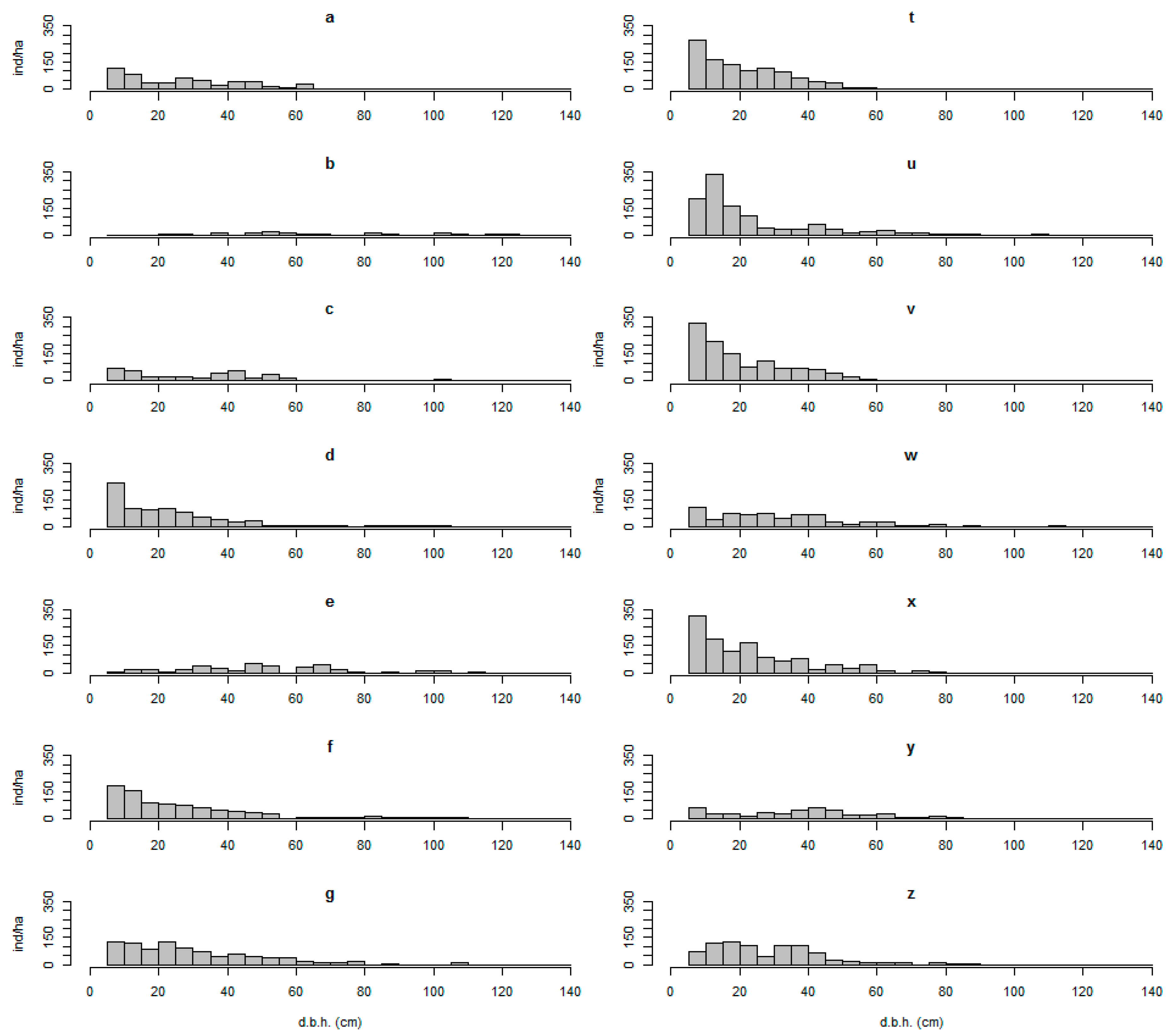
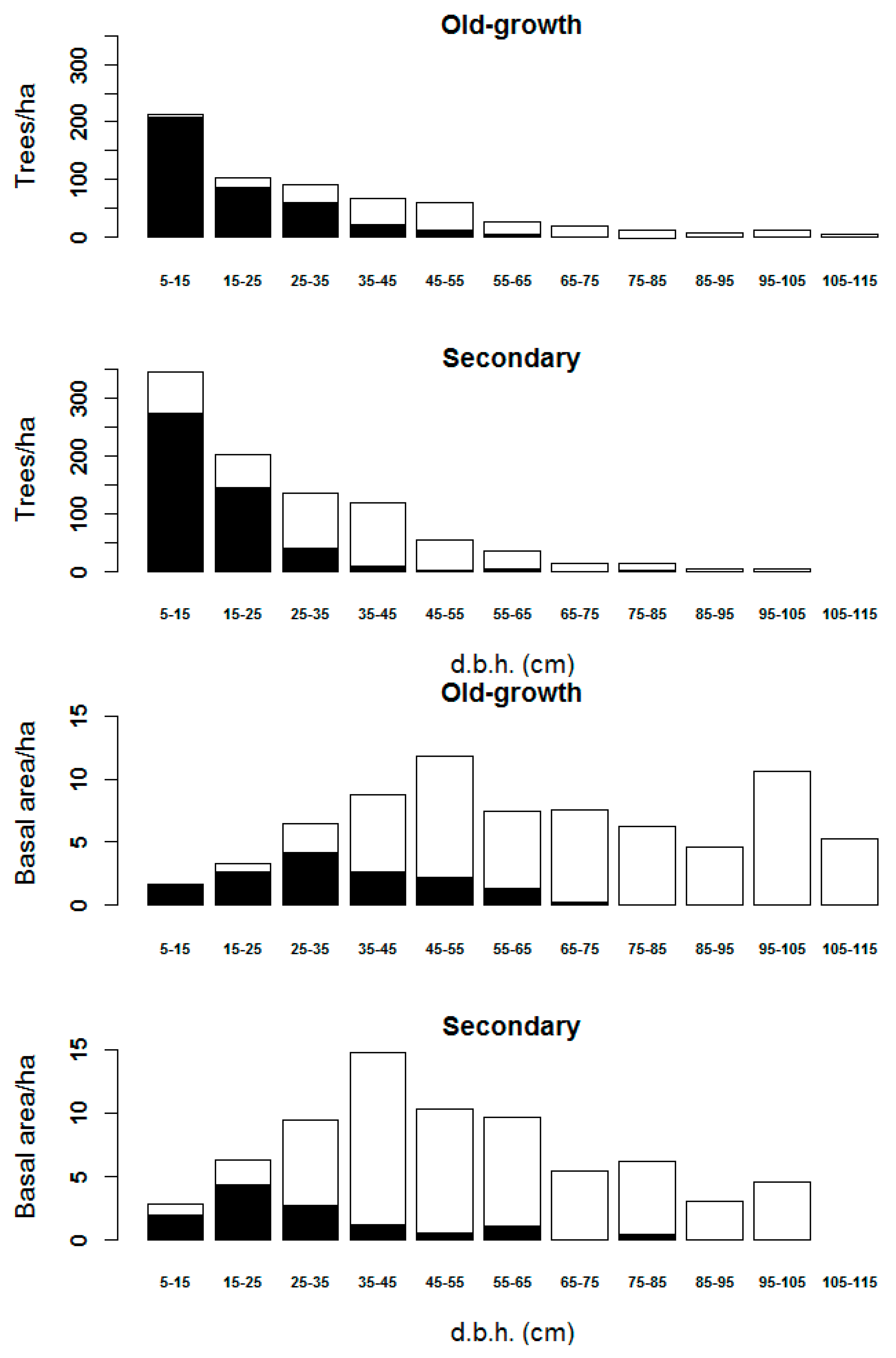
3.2. Diameter Classes Represented in the Remote Sensing Data
3.3. Stand Structure Variables
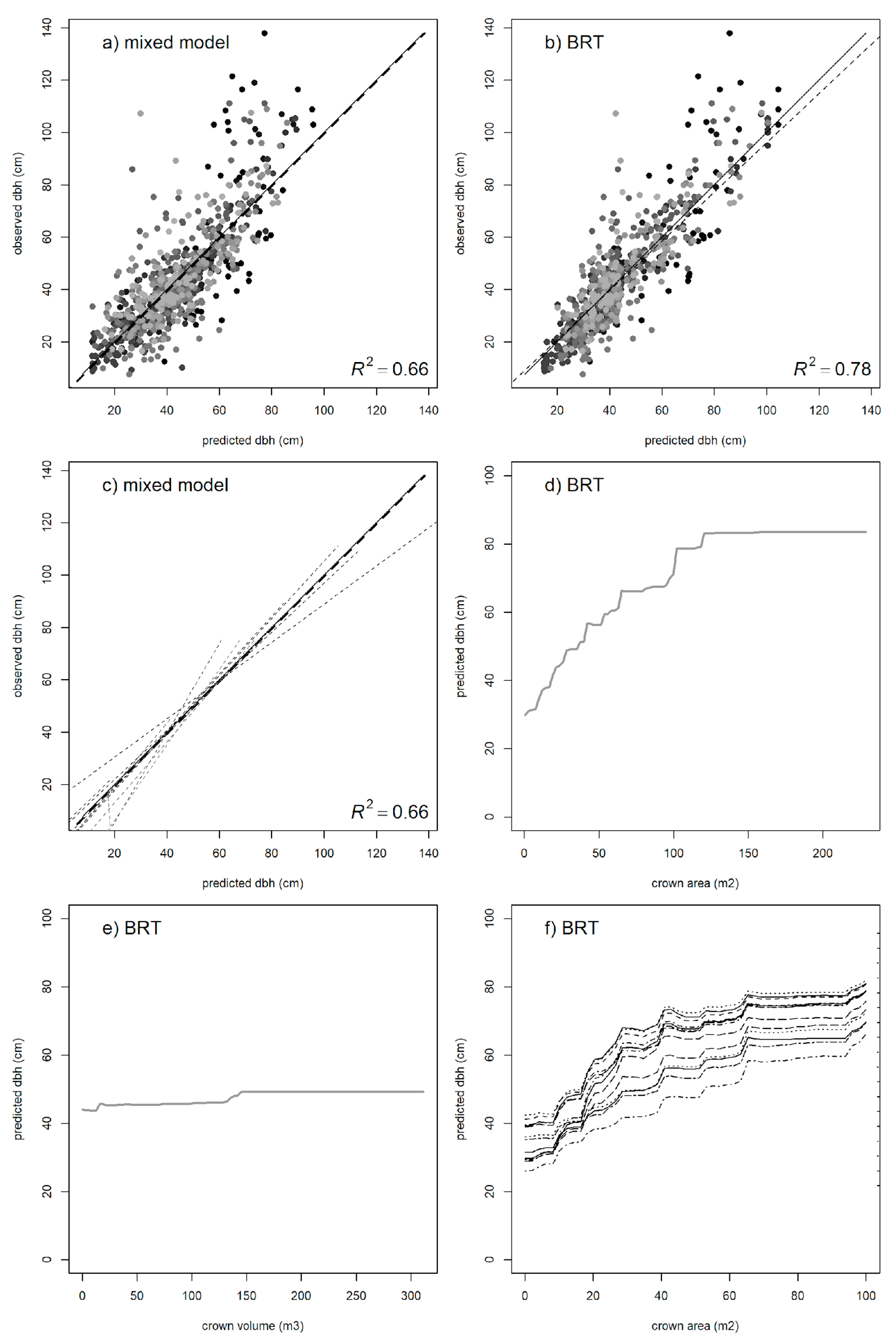
3.4. Individual-Tree DBH
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, X.-P.; Hansen, M.C.; Stehman, S.V.; Potapov, P.V.; Tyukavina, A.; Vermote, E.F.; Townshend, J.R. Global land change from 1982 to 2016. Nature 2018, 560, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Potapov, P.; Hansen, M.C.; Laestadius, L.; Turubanova, S.; Yaroshenko, A.; Thies, C.; Smith, W.; Zhuravleva, I.; Komarova, A.; Minnemeyer, S.; et al. The last frontiers of wilderness: Tracking loss of intact forest landscapes from 2000 to 2013. Sci. Adv. 2017, 3, e1600821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhl, M.; Lasco, R.; Cifuentes, M.; Jonsson, Ö.; Korhonen, K.T.; Mundhenk, P.; de Jesus Navar, J.; Stinson, G. Changes in forest production, biomass and carbon: Results from the 2015 UN FAO Global Forest Resource Assessment. For. Ecol. Manag. 2015, 352, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Whiteman, A.; Wickramasinghe, A.; Piña, L. Global trends in forest ownership, public income and expenditure on forestry and forestry employment. For. Ecol. Manag. 2015, 352, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Iglhaut, J.; Cabo, C.; Puliti, S.; Piermattei, L.; O’Connor, J.; Rosette, J. Structure from Motion Photogrammetry in Forestry: A Review. Curr. For. Rep. 2019, 5, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Anderson-Teixeira, K.J.; Davies, S.J.; Bennett, A.C.; Gonzalez-Akre, E.B.; Muller-Landau, H.C.; Joseph Wright, S.; Abu Salim, K.; Almeyda Zambrano, A.M.; Alonso, A.; Baltzer, J.L.; et al. CTFS-ForestGEO: A worldwide network monitoring forests in an era of global change. Glob. Chang. Biol. 2015, 21, 528–549. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.-D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, aaf8957. [Google Scholar] [CrossRef] [Green Version]
- Crowther, T.W.; Glick, H.B.; Covey, K.R.; Bettigole, C.; Maynard, D.S.; Thomas, S.M.; Smith, J.R.; Hintler, G.; Duguid, M.C.; Amatulli, G.; et al. Mapping tree density at a global scale. Nature 2015, 525, 201–205. [Google Scholar] [CrossRef]
- Glick, H.B.; Bettigole, C.; Maynard, D.S.; Covey, K.R.; Smith, J.R.; Crowther, T.W. Spatially-explicit models of global tree density. Sci. Data 2016, 3. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Rosenqvist, A.; Mora, B. Current remote sensing approaches to monitoring forest degradation in support of countries measurement, reporting and verification (MRV) systems for REDD+. Carbon Balance Manag. 2017, 12, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, J.L.; Gergel, S.E.; Coops, N.C. Aerial Photography: A Rapidly Evolving Tool for Ecological Management. BioScience 2010, 60, 47–59. [Google Scholar] [CrossRef]
- Bagaram, M.B.; Giuliarelli, D.; Chirici, G.; Giannetti, F.; Barbati, A. UAV Remote Sensing for Biodiversity Monitoring: Are Forest Canopy Gaps Good Covariates? Remote Sens. 2018, 10, 1397. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, K.; Schöning, I. Assessing biodiversity in forests using very high-resolution images and unmanned aerial vehicles: Assessing biodiversity in forests. Methods Ecol. Evol. 2012, 3, 397–404. [Google Scholar] [CrossRef]
- Tomaštík, J.; Mokroš, M.; Surový, P.; Grznárová, A.; Merganič, J. UAV RTK/PPK Method—An Optimal Solution for Mapping Inaccessible Forested Areas? Remote Sens. 2019, 11, 721. [Google Scholar] [CrossRef] [Green Version]
- Brovkina, O.; Cienciala, E.; Surový, P.; Janata, P. Unmanned aerial vehicles (UAV) for assessment of qualitative classification of Norway spruce in temperate forest stands. Geo-Spat. Inf. Sci. 2018, 21, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Romijn, E.; Lantican, C.B.; Herold, M.; Lindquist, E.; Ochieng, R.; Wijaya, A.; Murdiyarso, D.; Verchot, L. Assessing change in national forest monitoring capacities of 99 tropical countries. For. Ecol. Manag. 2015, 352, 109–123. [Google Scholar] [CrossRef] [Green Version]
- White, J.C.; Coops, N.C.; Wulder, M.A.; Vastaranta, M.; Hilker, T.; Tompalski, P. Remote Sensing Technologies for Enhancing Forest Inventories: A Review. Can. J. Remote Sens. 2016, 42, 619–641. [Google Scholar] [CrossRef] [Green Version]
- Goodbody, T.R.H.; Coops, N.C.; White, J.C. Digital Aerial Photogrammetry for Updating Area-Based Forest Inventories: A Review of Opportunities, Challenges, and Future Directions. Curr. For. Rep. 2019, 5, 55–75. [Google Scholar] [CrossRef] [Green Version]
- Puliti, S.; Solberg, S.; Granhus, A. Use of UAV Photogrammetric Data for Estimation of Biophysical Properties in Forest Stands Under Regeneration. Remote Sens. 2019, 11, 233. [Google Scholar] [CrossRef] [Green Version]
- Torresan, C.; Berton, A.; Carotenuto, F.; Di Gennaro, S.F.; Gioli, B.; Matese, A.; Miglietta, F.; Vagnoli, C.; Zaldei, A.; Wallace, L. Forestry applications of UAVs in Europe: A review. Int. J. Remote Sens. 2017, 38, 2427–2447. [Google Scholar] [CrossRef]
- Brach, M.; Chan, J.; Szymanski, P. Accuracy assessment of different photogrammetric software for processing data from low-cost UAV platforms in forest conditions. iForest Biogeosci. For. 2019, 12, 435–441. [Google Scholar] [CrossRef]
- Wallace, L.; Bellman, C.; Hally, B.; Hernandez, J.; Jones, S.; Hillman, S. Assessing the Ability of Image Based Point Clouds Captured from a UAV to Measure the Terrain in the Presence of Canopy Cover. Forests 2019, 10, 284. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Lee, X. Four-band Thermal Mosaicking: A New Method to Process Infrared Thermal Imagery of Urban Landscapes from UAV Flights. Remote Sens. 2019, 11, 1365. [Google Scholar] [CrossRef] [Green Version]
- Dandois, J.P.; Ellis, E.C. Remote Sensing of Vegetation Structure Using Computer Vision. Remote Sens. 2010, 2, 1157–1176. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Hu, J.; Lian, J.; Fan, Z.; Ouyang, X.; Ye, W. Seeing the forest from drones: Testing the potential of lightweight drones as a tool for long-term forest monitoring. Biol. Conserv. 2016, 198, 60–69. [Google Scholar] [CrossRef]
- Getzin, S.; Nuske, R.; Wiegand, K. Using Unmanned Aerial Vehicles (UAV) to Quantify Spatial Gap Patterns in Forests. Remote Sens. 2014, 6, 6988–7004. [Google Scholar] [CrossRef] [Green Version]
- Zielewska-Büttner, K.; Adler, P.; Ehmann, M.; Braunisch, V. Automated Detection of Forest Gaps in Spruce Dominated Stands Using Canopy Height Models Derived from Stereo Aerial Imagery. Remote Sens. 2016, 8, 175. [Google Scholar] [CrossRef] [Green Version]
- Mohan, M.; Silva, C.; Klauberg, C.; Jat, P.; Catts, G.; Cardil, A.; Hudak, A.; Dia, M. Individual Tree Detection from Unmanned Aerial Vehicle (UAV) Derived Canopy Height Model in an Open Canopy Mixed Conifer Forest. Forests 2017, 8, 340. [Google Scholar] [CrossRef] [Green Version]
- Reis, B.P.; Martins, S.V.; Fernandes Filho, E.I.; Sarcinelli, T.S.; Gleriani, J.M.; Marcatti, G.E.; Leite, H.G.; Halassy, M. Management Recommendation Generation for Areas Under Forest Restoration Process through Images Obtained by UAV and LiDAR. Remote Sens. 2019, 11, 1508. [Google Scholar] [CrossRef] [Green Version]
- Zahawi, R.A.; Dandois, J.P.; Holl, K.D.; Nadwodny, D.; Reid, J.L.; Ellis, E.C. Using lightweight unmanned aerial vehicles to monitor tropical forest recovery. Biol. Conserv. 2015, 186, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Swinfield, T.; Lindsell, J.A.; Williams, J.V.; Harrison, R.D.; Agustiono; Habibi; Gemita, E.; Schönlieb, C.B.; Coomes, D.A. Accurate Measurement of Tropical Forest Canopy Heights and Aboveground Carbon Using Structure From Motion. Remote Sens. 2019, 11, 928. [Google Scholar] [CrossRef] [Green Version]
- Alonzo, M.; Andersen, H.-E.; Morton, D.; Cook, B. Quantifying Boreal Forest Structure and Composition Using UAV Structure from Motion. Forests 2018, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Goodbody, T.R.H.; Coops, N.C.; Marshall, P.L.; Tompalski, P.; Crawford, P. Unmanned aerial systems for precision forest inventory purposes: A review and case study. For. Chron. 2017, 93, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Otero, V.; Van De Kerchove, R.; Satyanarayana, B.; Martínez-Espinosa, C.; Fisol, M.A.B.; Ibrahim, M.R.B.; Sulong, I.; Mohd-Lokman, H.; Lucas, R.; Dahdouh-Guebas, F. Managing mangrove forests from the sky: Forest inventory using field data and Unmanned Aerial Vehicle (UAV) imagery in the Matang Mangrove Forest Reserve, peninsular Malaysia. For. Ecol. Manag. 2018, 411, 35–45. [Google Scholar] [CrossRef]
- Puliti, S.; Ørka, H.; Gobakken, T.; Næsset, E. Inventory of Small Forest Areas Using an Unmanned Aerial System. Remote Sens. 2015, 7, 9632–9654. [Google Scholar] [CrossRef] [Green Version]
- Matese, A.; Toscano, P.; Di Gennaro, S.; Genesio, L.; Vaccari, F.; Primicerio, J.; Belli, C.; Zaldei, A.; Bianconi, R.; Gioli, B. Intercomparison of UAV, Aircraft and Satellite Remote Sensing Platforms for Precision Viticulture. Remote Sens. 2015, 7, 2971–2990. [Google Scholar] [CrossRef] [Green Version]
- Koh, L.P.; Wich, S.A. Dawn of drone ecology: Low-cost autonomous aerial vehicles for conservation. Trop. Conserv. Sci. 2012. [Google Scholar] [CrossRef] [Green Version]
- Friedman, J.H. Stochastic gradient boosting. Comput. Stat. Data Anal. 2002, 38, 367–378. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J.; Hijmans, M.R.J. Package ‘dismo’. Circles 2017, 9, 1–68. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. Linear and nonlinear mixed effects models. R Package Version 2007, 3, 1–89. [Google Scholar]
- Gutiérrez, A.G.; Armesto, J.J.; Aravena, J.-C.; Carmona, M.; Carrasco, N.V.; Christie, D.A.; Peña, M.-P.; Pérez, C.; Huth, A. Structural and environmental characterization of old-growth temperate rainforests of northern Chiloé Island, Chile: Regional and global relevance. For. Ecol. Manag. 2009, 258, 376–388. [Google Scholar] [CrossRef]
- Ponce, D.B.; Donoso, P.J.; Salas-Eljatib, C. Índice de bosque adulto: Una herramienta para evaluar estados de desarrollo de bosques nativos de tierras bajas del centro-sur de Chile. Bosque (Valdivia) 2019, 40, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Donoso, P.J.; Lusk, C.H. Differential effects of emergent Nothofagus dombeyi on growth and basal area of canopy species in an old-growth temperate rainforest. J. Veg. Sci. 2007, 18, 675–684. [Google Scholar] [CrossRef]
- Donoso, P.; Promis, Á.; Soto, D. Silviculture in Native Forests Experiences in Silviculture and Restoration in Chile, Argentina and Western USA; The Chile Initiative & OSU College of Forestry: Valdivia, Chile, 2019. [Google Scholar]
- Donoso, P.J. Crown Index: A canopy balance indicator to assess growth and regeneration in uneven-aged forest stands of the Coastal Range of Chile. For. Int. J. For. Res. 2005, 78, 337–351. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H. Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. For. Ecol. Manag. 2014, 327, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Lausch, A.; Bannehr, L.; Beckmann, M.; Boehm, C.; Feilhauer, H.; Hacker, J.M.; Heurich, M.; Jung, A.; Klenke, R.; Neumann, C.; et al. Linking Earth Observation and taxonomic, structural and functional biodiversity: Local to ecosystem perspectives. Ecol. Indic. 2016, 70, 317–339. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E. Airborne spectranomics: Mapping canopy chemical and taxonomic diversity in tropical forests. Front. Ecol. Environ. 2009, 7, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Ganivet, E.; Bloomberg, M. Towards rapid assessments of tree species diversity and structure in fragmented tropical forests: A review of perspectives offered by remotely-sensed and field-based data. For. Ecol. Manag. 2019, 432, 40–53. [Google Scholar] [CrossRef]
- Franklin, J.F.; Van Pelt, R. Spatial Aspects of Structural Complexity in Old-Growth Forests. J. For. 2004, 102, 22–28. [Google Scholar] [CrossRef]
- Spies, T.A. Ecological Concepts and Diversity of Old-Growth Forests. J. For. 2004, 102, 14–20. [Google Scholar] [CrossRef]
- Laurance, W.F.; Nascimento, H.E.M.; Laurance, S.G.; Andrade, A.; Ribeiro, J.E.L.S.; Giraldo, J.P.; Lovejoy, T.E.; Condit, R.; Chave, J.; Harms, K.E.; et al. Rapid decay of tree-community composition in Amazonian forest fragments. Proc. Natl. Acad. Sci. USA 2006, 103, 19010–19014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutsch, W.L.; Wirth, C.; Kattge, J.; Nöllert, S.; Herbst, M.; Kappen, L. Ecophysiological Characteristics of Mature Trees and Stands—Consequences for Old-Growth Forest Productivity. In Old-Growth Forests; Wirth, C., Gleixner, G., Heimann, M., Eds.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2009; Volume 207, pp. 57–79. ISBN 978-3-540-92705-1. [Google Scholar]
- Wirth, C. Old-Growth Forests: Function, Fate and Value—A Synthesis. In Old-Growth Forests; Wirth, C., Gleixner, G., Heimann, M., Eds.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2009; Volume 207, pp. 465–491. ISBN 978-3-540-92705-1. [Google Scholar]
- Vásquez-Grandón, A.; Donoso, P.; Gerding, V. Forest Degradation: When Is a Forest Degraded? Forests 2018, 9, 726. [Google Scholar] [CrossRef] [Green Version]
- Ghazoul, J.; Burivalova, Z.; Garcia-Ulloa, J.; King, L.A. Conceptualizing Forest Degradation. Trends Ecol. Evol. 2015, 30, 622–632. [Google Scholar] [CrossRef]
- Stanturf, J.A.; Palik, B.J.; Dumroese, R.K. Contemporary forest restoration: A review emphasizing function. For. Ecol. Manag. 2014, 331, 292–323. [Google Scholar] [CrossRef]
- Xiang, W.; Hassegawa, M.; Franceschini, T.; Leitch, M.; Achim, A. Characterizing wood density–climate relationships along the stem in black spruce (Picea mariana (Mill.) B.S.P.) using a combination of boosted regression trees and mixed-effects models. For. Int. J. For. Res. 2019, 92, 357–374. [Google Scholar] [CrossRef]
- Cardil, A.; Mola-Yudego, B.; Blázquez-Casado, Á.; González-Olabarria, J.R. Fire and burn severity assessment: Calibration of Relative Differenced Normalized Burn Ratio (RdNBR) with field data. J. Environ. Manag. 2019, 235, 342–349. [Google Scholar] [CrossRef]
- Reder, S.; Waßermann, L.; Mund, J.-P. UAV-based Tree Height Estimation in Dense Tropical Rainforest Areas in Ecuador and Brazil. Gi_Forum 2019, 1, 47–59. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
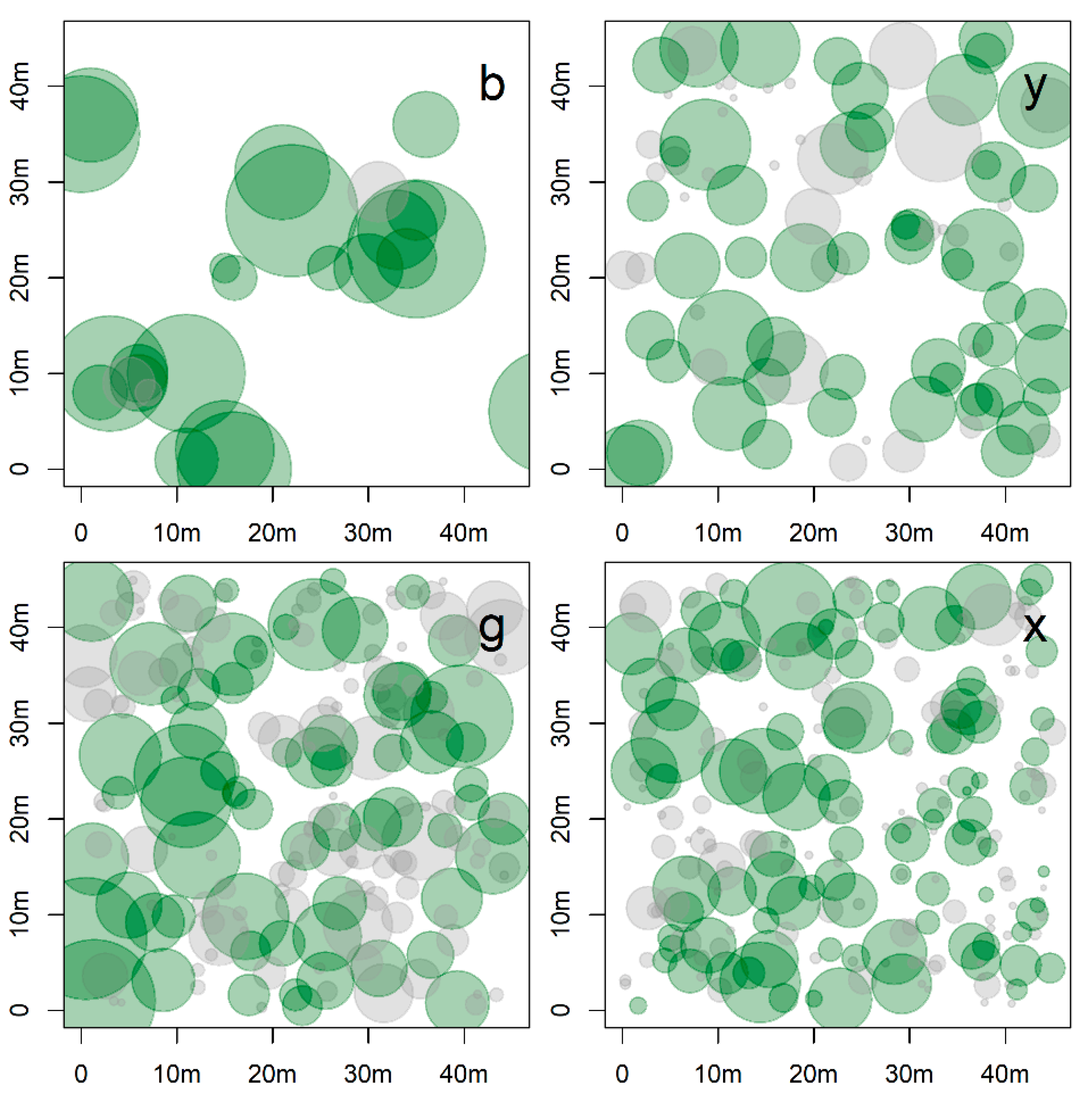
| Plot | Density (Trees/ha) | BA (m2) | QMD (cm) | Trees Detected (%) | BA Detected (%) | Cover Area (%) | Forest Canopy | Mean Crown Area (m2) | Mean Crown Volume (m3) | Dominant Species |
|---|---|---|---|---|---|---|---|---|---|---|
| Height (m) | ||||||||||
| Old-growth forest | ||||||||||
| a | 572.8 | 67.9 | 38.9 | 31.9 | 74.3 | 77 | 12.7 (cv = 28) | 41.9 (cv = 109) | 35.8 (cv = 194) | Aa |
| b | 123.5 | 53.4 | 74.2 | 88.0 | 96.0 | 31 | 14 (cv = 25) | 28.6 (cv = 42) | 17.4 (cv = 87) | Aa |
| c | 439.5 | 68.9 | 44.7 | 49.4 | 87.2 | 79 | 21.3 (cv = 12) | 36.4 (cv = 112) | 21.3 (cv = 149) | Aa-Nd |
| d | 849.4 | 67.6 | 31.8 | 26.2 | 71.3 | 98 | 20.2 (cv = 21) | 44.3 (cv = 131) | 36.8 (cv = 155) | Nd-Pl |
| e | 355.6 | 87.0 | 55.8 | 63.9 | 86.8 | 79 | 21.2(cv = 12) | 34.6 (cv = 114) | 22.3 (cv = 144) | Ap-Lp |
| f | 864.2 | 78.3 | 34.0 | 28.0 | 75.8 | 100 | 21.7 (cv = 20) | 41.3 (cv = 108) | 31.3 (cv = 136) | Nd-Ap |
| g | 928.4 | 97.3 | 36.5 | 34.6 | 70.6 | 100 | 20.7 (cv = 17) | 32.2 (cv = 108) | 19.6 (cv = 153) | Ap-Nd |
| Mean | 590.5 | 74.3 | 45.1 | 46.0 | 80.3 | 81 | 18.8 (cv = 19) | 37 (cv = 104) | 26. 4 (cv = 145) | |
| Secondary forest | ||||||||||
| t | 1037.0 | 48.9 | 24.5 | 35.7 | 75.1 | 84 | 21.1 (cv = 23) | 22.8 (cv = 72) | 14.5 (cv = 119) | No-Pl |
| u | 1101.2 | 73.5 | 29.1 | 43.0 | 83.0 | 100 | 18 (cv = 31) | 21.9 (cv = 157) | 13.5 (cv = 189) | Pl-Nd |
| v | 1165.4 | 60.6 | 25.7 | 30.9 | 75.8 | 94 | 21.7 (cv = 15) | 25.9 (cv = 69) | 19.6 (cv = 108) | Pl-No |
| w | 661.7 | 71.3 | 37.0 | 61.2 | 92.2 | 82 | 19.7 (cv = 16) | 20.4 (cv = 92) | 12.5 (cv = 145) | No-Ec |
| x | 1170.4 | 69.5 | 27.5 | 40.9 | 80.1 | 88 | 16.2 (cv = 19) | 18.4 (cv = 100) | 7.8 (cv = 177) | Ap-Ec |
| y | 444.4 | 61.9 | 42.1 | 57.8 | 78.9 | 88 | 22.5 (cv = 12) | 34.4 (cv = 86) | 20.9 (cv = 113) | Nd-No |
| z | 834.6 | 70.7 | 32.8 | 44.4 | 76.7 | 84 | 22.8 (cv = 20) | 22.6 (cv = 81) | 13.2 (cv = 106) | No-Nd |
| Mean | 916.4 | 65.2 | 31.3 | 44.9 | 80.3 | 89 | 20.2 (cv = 19) | 23.7 (cv = 94) | 14.6 (cv = 137) | |
| Variable | Estimate | Std. Error | p-Value |
|---|---|---|---|
| (a) Relative density | |||
| Intercept (β0) | −339.06 | 246.6 | 0.19 |
| Forest canopy area (β1) | 0.83 | 0.15 | <0.001 |
| SD trees crown area (β2) | −11.33 | 4.03 | 0.02 |
| (b) Quadratic mean diameter | |||
| Intercept (β0) | 88.32 | 9.33 | <0.001 |
| Forest canopy area (β1) | −0.03 | 0.01 | <0.00 |
| SD trees crown area (β2) | 0.32 | 0.15 | 0.06 |
| (c) Basal area | |||
| Intercept (β0) | 41.51 | 15.11 | 0.02 |
| Forest canopy area (β1) | 0.01 | 0.01 | 0.24 |
| SD trees crown area (β2) | 0.26 | 0.25 | 0.31 |
| Parameter | Variable | Value | SE | p-Value |
|---|---|---|---|---|
| β0 | −7.99 | 3.08 | 0.010 | |
| β1 | log (crown area) | 17.89 | 0.67 | <0.001 |
| β2 | crown area−1 | 33.56 | 4.09 | <0.002 |
| σsp | species | 6.81 | ||
| σe | residual | 12.06 |
| Species | β0 (1) | BRT (2) |
|---|---|---|
| Nothofagus obliqua | −9.71 | 37.87 |
| Persea lingue | −4.17 | 42.90 |
| Eucryphia cordifolia | −0.51 | 44.44 |
| Gevuina avellana | −3.57 | 45.73 |
| N. dombeyi | −0.01 | 46.18 |
| Peumus boldus | −4.95 | 46.52 |
| Lomatia dentata | −5.90 | 46.73 |
| Aextoxicon punctatum | 4.51 | 52.05 |
| Laureliopsis philippiana | 5.20 | 53.19 |
| Araucaria araucana | 9.81 | 53.62 |
| Laurelia sempervirens | 8.49 | 54.02 |
| Lomatia hirsuta | 0.81 | 54.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miranda, A.; Catalán, G.; Altamirano, A.; Zamorano-Elgueta, C.; Cavieres, M.; Guerra, J.; Mola-Yudego, B. How Much Can We See from a UAV-Mounted Regular Camera? Remote Sensing-Based Estimation of Forest Attributes in South American Native Forests. Remote Sens. 2021, 13, 2151. https://doi.org/10.3390/rs13112151
Miranda A, Catalán G, Altamirano A, Zamorano-Elgueta C, Cavieres M, Guerra J, Mola-Yudego B. How Much Can We See from a UAV-Mounted Regular Camera? Remote Sensing-Based Estimation of Forest Attributes in South American Native Forests. Remote Sensing. 2021; 13(11):2151. https://doi.org/10.3390/rs13112151
Chicago/Turabian StyleMiranda, Alejandro, Germán Catalán, Adison Altamirano, Carlos Zamorano-Elgueta, Manuel Cavieres, Javier Guerra, and Blas Mola-Yudego. 2021. "How Much Can We See from a UAV-Mounted Regular Camera? Remote Sensing-Based Estimation of Forest Attributes in South American Native Forests" Remote Sensing 13, no. 11: 2151. https://doi.org/10.3390/rs13112151
APA StyleMiranda, A., Catalán, G., Altamirano, A., Zamorano-Elgueta, C., Cavieres, M., Guerra, J., & Mola-Yudego, B. (2021). How Much Can We See from a UAV-Mounted Regular Camera? Remote Sensing-Based Estimation of Forest Attributes in South American Native Forests. Remote Sensing, 13(11), 2151. https://doi.org/10.3390/rs13112151