
Missing the Forest and the Trees: Utility, Limits and Caveats for Drone Imaging of Coastal Marine Ecosystems


Abstract
:1. Introduction
2. Materials and Methods
2.1. RGB Imaging of Rocky Reef Ecosystems
2.2. Multispectral Imaging
2.3. Analysis and Validation
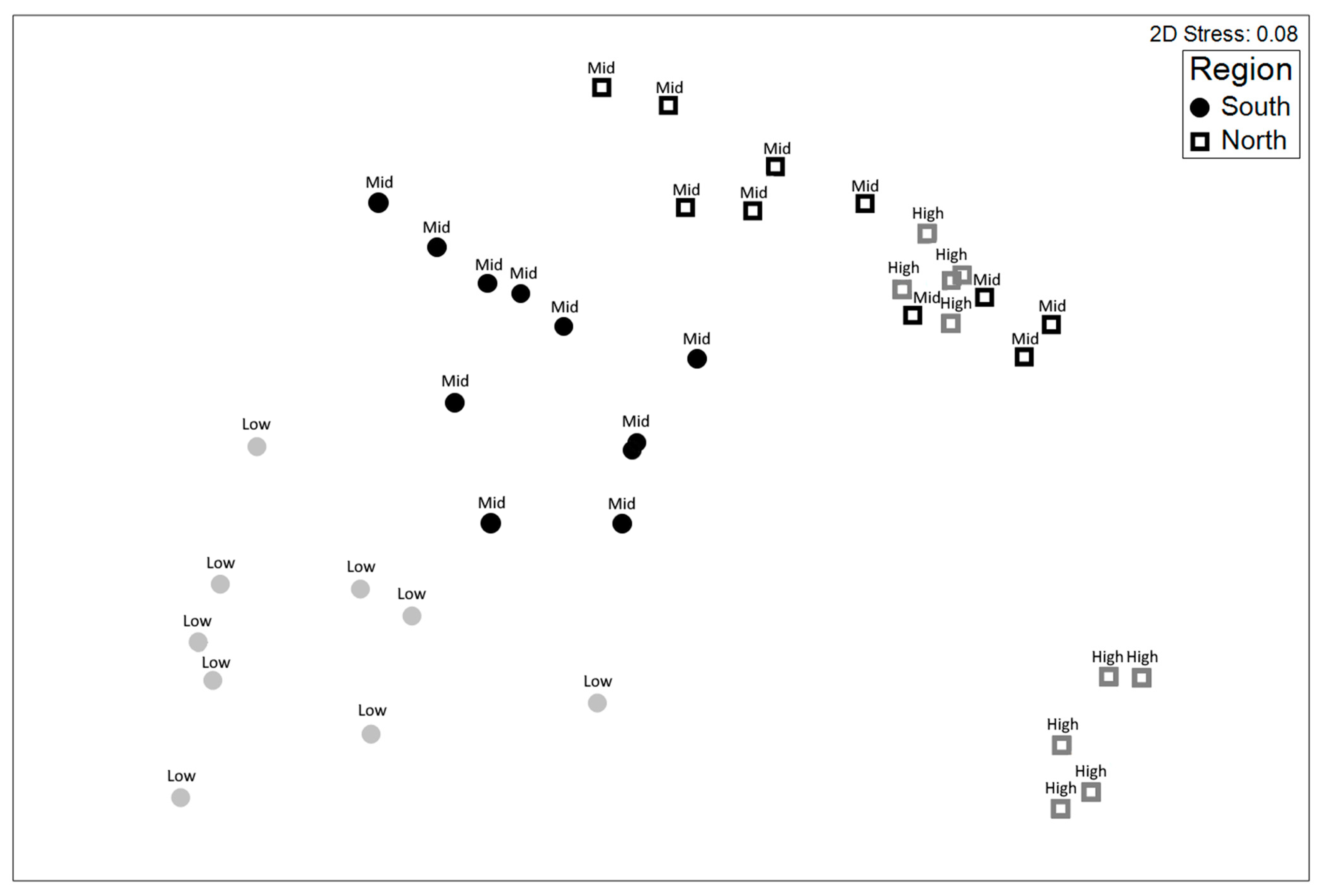
3. Results
3.1. RGB Imaging of Rocky Reef Ecosystems
3.2. Multispectral Imaging
4. Discussion
- Detection of unique spectral signatures is greatly impeded by overlying water, leading to further aggregation of unique habitat/species classes.
- Features of interest (FOIs) smaller than c. 5–20 cm2 require very low elevation flights to define individual FOIs by >1–4 pixels. This greatly limits:
- ▪
- The use of imagery for collection of “training samples”.
- ▪
- The size of the area that can be covered under the limitations of UAV batteries, and tidal cycles.
- Full taxonomic inventory is not readily achievable (even for macroalgae) from aerial imagery alone.
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomsen, M.S.; Mondardini, L.; Alestra, T.; Gerrity, S.; Tait, L.; South, P.M.; Lilley, S.A.; Schiel, D.R. Local extinction of bull kelp (Durvillaea spp.) due to a marine heatwave. Front. Mar. Sci. 2019, 6, 84. [Google Scholar] [CrossRef] [Green Version]
- Wernberg, T.; Bennett, S.; Babcock, R.C.; De Bettignies, T.; Cure, K.; Depczynski, M.; Dufois, F.; Fromont, J.; Fulton, C.J.; Hovey, R.K. Climate-driven regime shift of a temperate marine ecosystem. Science 2016, 353, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Wernberg, T.; Smale, D.A.; Tuya, F.; Thomsen, M.S.; Langlois, T.J.; de Bettignies, T.; Bennett, S.; Rousseaux, C.S. An extreme climatic event alters marine ecosystem structure in a global biodiversity hotspot. Nat. Clim. Chang. 2013, 3, 78–82. [Google Scholar] [CrossRef]
- Schiel, D.R.; Lilley, S.A.; South, P.M.; Coggins, J.H. Decadal changes in sea surface temperature, wave forces and intertidal structure in New Zealand. Mar. Ecol. Prog. Ser. 2016, 548, 77–95. [Google Scholar] [CrossRef] [Green Version]
- Krumhansl, K.A.; Okamoto, D.K.; Rassweiler, A.; Novak, M.; Bolton, J.J.; Cavanaugh, K.C.; Connell, S.D.; Johnson, C.R.; Konar, B.; Ling, S.D.; et al. Global patterns of kelp forest change over the past half-century. Proc. Natl. Acad. Sci. USA 2016, 113, 13785–13790. [Google Scholar] [CrossRef] [Green Version]
- Graham, M.H.; Vasquez, J.A.; Buschmann, A.H. Global ecology of the giant kelp Macrocystis: From ecotypes to ecosystems. Oceanogr. Mar. Biol. 2007, 45, 39. [Google Scholar]
- Wernberg, T.; Coleman, M.A.; Babcock, R.C.; Bell, S.Y.; Bolton, J.J.; Connell, S.D.; Hurd, C.L.; Johnson, C.R.; Marzinelli, E.M.; Shears, N.T.; et al. Biology and ecology of the globally significant kelp Ecklonia radiata. Oceanogr. Mar. Biol. 2019, 57, 265–324. [Google Scholar]
- Krause-Jensen, D.; Duarte, C.M. Substantial role of macroalgae in marine carbon sequestration. Nat. Geosci. 2016, 9, 737–742. [Google Scholar] [CrossRef]
- Pedersen, M.F.; Filbee-Dexter, K.; Frisk, N.L.; Sárossy, Z.; Wernberg, T. Carbon sequestration potential increased by incomplete anaerobic decomposition of kelp detritus. Mar. Ecol. Prog. Ser. 2021, 660, 53–67. [Google Scholar] [CrossRef]
- Velásquez, M.; Fraser, C.I.; Nelson, W.A.; Tala, F.; Macaya, E.C. Concise review of the genus Durvillaea Bory de Saint-Vincent, 1825. J. Appl. Phycol. 2020, 32, 3–21. [Google Scholar] [CrossRef]
- Schiel, D.R. Experimental analyses of diversity partitioning in southern hemisphere algal communities. Oecologia 2019, 190, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Tait, L.W.; Schiel, D.R. Legacy Effects of Canopy Disturbance on Ecosystem Functioning in Macroalgal Assemblages. PLoS ONE 2011, 6, e26986. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.J.; Nissen, E.K.; Howarth, J.D.; Hamling, I.J.; Mountjoy, J.J.; Ries, W.F.; Jones, K.; Goldstien, S.; Cochran, U.A.; Villamor, P.; et al. Highly variable coastal deformation in the 2016 MW7.8 Kaikōura earthquake reflects rupture complexity along a transpressional plate boundary. Earth Planet. Sci. Lett. 2017, 474, 334–344. [Google Scholar] [CrossRef]
- Hamling, I.J.; Hreinsdóttir, S.; Clark, K.; Elliott, J.; Liang, C.; Fielding, E.; Litchfield, N.; Villamor, P.; Wallace, L.; Wright, T.J.; et al. Complex multifault rupture during the 2016 Mw 7.8 Kaikōura earthquake, New Zealand. Science 2017, 7194. [Google Scholar] [CrossRef] [Green Version]
- Quigley, M.C.; Hughes, M.W.; Bradley, B.A.; van Ballegooy, S.; Reid, C.; Morgenroth, J.; Horton, T.; Duffy, B.; Pettinga, J.R. The 2010–2011 Canterbury Earthquake Sequence: Environmental effects, seismic triggering thresholds and geologic legacy. Tectonophysics 2016, 672–673, 228–274. [Google Scholar] [CrossRef] [Green Version]
- Schiel, D.R.; Alestra, T.; Gerrity, S.; Orchard, S.; Dunmore, R.; Pirker, J.; Lilley, S.; Tait, L.; Hickford, M.; Thomsen, M. The Kaikōura earthquake in southern New Zealand: Loss of connectivity of marine communities and the necessity of a cross-ecosystem perspective. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 1520–1534. [Google Scholar] [CrossRef]
- Layton, C.; Coleman, M.A.; Marzinelli, E.M.; Steinberg, P.D.; Swearer, S.E.; Vergés, A.; Wernberg, T.; Johnson, C.R. Kelp forest restoration in Australia. Front. Mar. Sci. 2020, 7, 74. [Google Scholar] [CrossRef] [Green Version]
- Bell, T.W.; Allen, J.G.; Cavanaugh, K.C.; Siegel, D.A. Three decades of variability in California’s giant kelp forests from the Landsat satellites. Remote Sens. Environ. 2020, 238, 110811. [Google Scholar] [CrossRef]
- Bell, T.W.; Cavanaugh, K.C.; Reed, D.C.; Siegel, D.A. Geographical variability in the controls of giant kelp biomass dynamics. J. Biogeogr. 2015, 42, 2010–2021. [Google Scholar] [CrossRef]
- Butler, C.L.; Lucieer, V.L.; Wotherspoon, S.J.; Johnson, C.R. Multi-decadal decline in cover of giant kelp Macrocystis pyrifera at the southern limit of its Australian range. Mar. Ecol. Prog. Ser. 2020, 653, 1–18. [Google Scholar] [CrossRef]
- Cavanaugh, K.C.; Siegel, D.A.; Kinlan, B.P.; Reed, D.C. Scaling giant kelp field measurements to regional scales using satellite observations. Mar. Ecol. Prog. Ser. 2010, 403, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Mora-Soto, A.; Palacios, M.; Macaya, E.C.; Gómez, I.; Huovinen, P.; Pérez-Matus, A.; Young, M.; Golding, N.; Toro, M.; Yaqub, M.; et al. A High-Resolution Global Map of Giant Kelp (Macrocystis pyrifera) Forests and Intertidal Green Algae (Ulvophyceae) with Sentinel-2 Imagery. Remote Sens. 2020, 12, 694. [Google Scholar] [CrossRef] [Green Version]
- Tait, L.; Bind, J.; Charan-Dixon, H.; Hawes, I.; Pirker, J.; Schiel, D. Unmanned Aerial Vehicles (UAVs) for monitoring macroalgal biodiversity: Comparison of RGB and multispectral imaging sensors for biodiversity assessments. Remote Sens. 2019, 11, 2332. [Google Scholar] [CrossRef] [Green Version]
- Menge, B.A.; Farrell, T.M.; Oison, A.M.; van Tamelen, P.; Turner, T. Algal recruitment and the maintenance of a plant mosaic in the low intertidal region on the Oregon coast. J. Exp. Mar. Biol. Ecol. 1993, 170, 91–116. [Google Scholar] [CrossRef]
- Tait, L.W.; Schiel, D.R. Ecophysiology of Layered Macroalgal Assemblages: Importance of Subcanopy Species Biodiversity in Buffering Primary Production. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef]
- Murfitt, S.L.; Allan, B.M.; Bellgrove, A.; Rattray, A.; Young, M.A.; Ierodiaconou, D. Applications of unmanned aerial vehicles in intertidal reef monitoring. Sci. Rep. 2017, 7, 10259. [Google Scholar] [CrossRef] [Green Version]
- Rossiter, T.; Furey, T.; McCarthy, T.; Stengel, D.B. Application of multiplatform, multispectral remote sensors for mapping intertidal macroalgae: A comparative approach. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 1595–1612. [Google Scholar] [CrossRef]
- Ventura, D.; Bonifazi, A.; Gravina, M.F.; Belluscio, A.; Ardizzone, G. Mapping and Classification of Ecologically Sensitive Marine Habitats Using Unmanned Aerial Vehicle (UAV) Imagery and Object-Based Image Analysis (OBIA). Remote Sens. 2018, 10, 1331. [Google Scholar] [CrossRef] [Green Version]
- Rossiter, T.; Furey, T.; McCarthy, T.; Stengel, D.B. UAV-mounted hyperspectral mapping of intertidal macroalgae. Estuar. Coast. Shelf Sci. 2020, 242, 106789. [Google Scholar] [CrossRef]
- Olmedo-Masat, O.M.; Raffo, M.P.; Rodríguez-Pérez, D.; Arijón, M.; Sánchez-Carnero, N. How Far Can We Classify Macroalgae Remotely? An Example Using a New Spectral Library of Species from the South West Atlantic (Argentine Patagonia). Remote Sens. 2020, 12, 3870. [Google Scholar] [CrossRef]
- Parsons, M.; Bratanov, D.; Gaston, K.J.; Gonzalez, F. UAVs, hyperspectral remote sensing, and machine learning revolutionizing reef monitoring. Sensors 2018, 18, 2026. [Google Scholar] [CrossRef] [Green Version]
- Alestra, T.; Gerrity, S.; Dunmore, R.A.; Crossett, D.; Orchard, S.; Schiel, D.R. Rocky reef impacts of the 2016 Kaikōura earthquake: Extended monitoring of nearshore habitats and communities to 3.5 years. In New Zealand Aquatic Environment and Biodiversity Report No. 253; Ministry for Primary Industries: Wellington, New Zealand, 2021. [Google Scholar]
- Thomsen, M.S.; Metcalfe, I.; Siciliano, A.; South, P.M.; Gerrity, S.; Alestra, T.; Schiel, D.R. Earthquake-driven destruction of an intertidal habitat cascade. Aquat. Bot. 2020, 164, 103217. [Google Scholar] [CrossRef]
- Schiel, D.R. Rivets or bolts? When single species count in the function of temperate rocky reef communities. J. Exp. Mar. Biol. Ecol. 2006, 338, 233–252. [Google Scholar] [CrossRef]
- Lilley, S.A.; Schiel, D.R. Community effects following the deletion of a habitat-forming alga from rocky marine shores. Oecologia 2006, 148, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Schiel, D.R.; Lilley, S.A. Gradients of disturbance to an algal canopy and the modification of an intertidal community. Mar. Ecol. Prog. Ser. 2007, 339, 1–11. [Google Scholar] [CrossRef]
- Foody, G.M. Status of land cover classification accuracy assessment. Remote Sens. Environ. 2002, 80, 185–201. [Google Scholar] [CrossRef]
- Kutser, T.; Miller, I.; Jupp, D.L. Mapping coral reef benthic substrates using hyperspectral space-borne images and spectral libraries. Estuar. Coast. Shelf Sci. 2006, 70, 449–460. [Google Scholar] [CrossRef]
- Rodríguez, Y.C.; Gómez, J.D.; Sánchez-Carnero, N.; Rodríguez-Pérez, D. A comparison of spectral macroalgae taxa separability methods using an extensive spectral library. Algal Res. 2017, 26, 463–473. [Google Scholar] [CrossRef]
- Vahtmäe, E.; Kutser, T.; Martin, G.; Kotta, J. Feasibility of hyperspectral remote sensing for mapping benthic macroalgal cover in turbid coastal waters—A Baltic Sea case study. Remote Sens. Environ. 2006, 101, 342–351. [Google Scholar] [CrossRef]
- Tait, L.W. Giant kelp forests at critical light thresholds show compromised ecological resilience to environmental and biological drivers. Estuar. Coast. Shelf Sci. 2019, 219, 231–241. [Google Scholar] [CrossRef]
- Schiel, D.R.; Howard-Williams, C. Controlling inputs from the land to sea: Limit-setting, cumulative impacts and ki uta ki tai. Mar. Freshw. Res. 2016, 67, 57–64. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | Eco-Region | Coordinates | Earthquake Vertical Deformation (m) |
|---|---|---|---|
| Cape Campbell | North | −41.724430, 174.2772 | 1.1 |
| Gate Reef | North | −41.812753, 174.216667 | 1.4 |
| Waipapa Reef | North | −42.209011, 173.879163 | 5.5 |
| Okiwi Reef | North | −42.218046, 173.87096 | 2.3 |
| Paia Point | South | −42.472953, 173.536960 | 1.1 |
| Omihi Reef | South | −42.49100, 173.523796 | 1.7 |
| Oaro North | South | −42.516454, 173.508488 | 0.0 |
| Oaro South | South | −42.521991, 173.506124 | −0.3 |
| Species/Habitat | Producer Accuracy | User Accuracy |
|---|---|---|
| Ulva | 0.94 | 0.63 |
| Brown algae | 0.97 | 0.92 |
| Red algae | 0.99 | 0.93 |
| Coralline algae | 0.998 | 0.91 |
| Durvillaea | 0.99 | 0.98 |
| Mussel beds | 0.994 | 0.89 |
| Rock | 0.994 | 0.86 |
| Water | 0.998 | 0.98 |
| Seafoam | 1 | 0.996 |
| Total | 0.986 | 0.90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tait, L.W.; Orchard, S.; Schiel, D.R. Missing the Forest and the Trees: Utility, Limits and Caveats for Drone Imaging of Coastal Marine Ecosystems. Remote Sens. 2021, 13, 3136. https://doi.org/10.3390/rs13163136
Tait LW, Orchard S, Schiel DR. Missing the Forest and the Trees: Utility, Limits and Caveats for Drone Imaging of Coastal Marine Ecosystems. Remote Sensing. 2021; 13(16):3136. https://doi.org/10.3390/rs13163136
Chicago/Turabian StyleTait, Leigh W., Shane Orchard, and David R. Schiel. 2021. "Missing the Forest and the Trees: Utility, Limits and Caveats for Drone Imaging of Coastal Marine Ecosystems" Remote Sensing 13, no. 16: 3136. https://doi.org/10.3390/rs13163136
APA StyleTait, L. W., Orchard, S., & Schiel, D. R. (2021). Missing the Forest and the Trees: Utility, Limits and Caveats for Drone Imaging of Coastal Marine Ecosystems. Remote Sensing, 13(16), 3136. https://doi.org/10.3390/rs13163136