
Estimating Surface Concentrations of Calanus finmarchicus Using Standardised Satellite-Derived Enhanced RGB Imagery

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
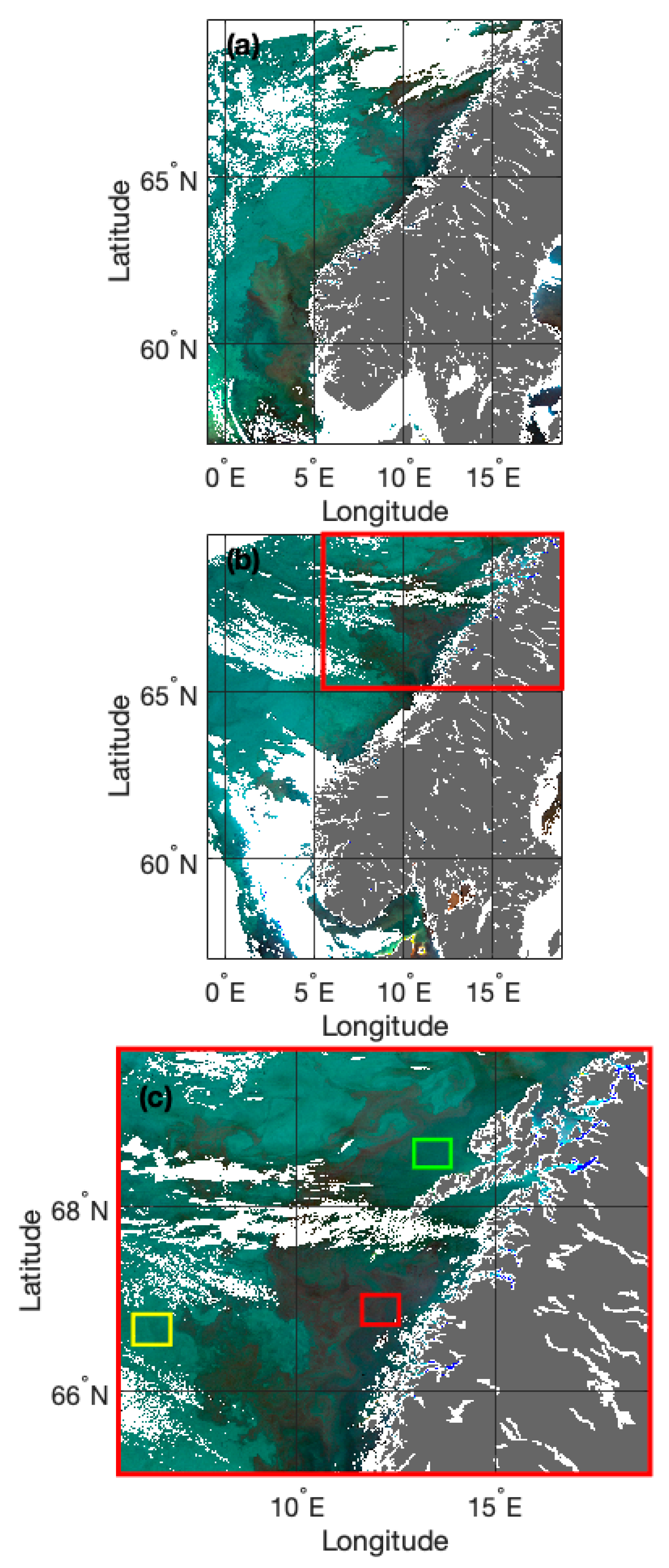
2.1. Study Site and Field Data
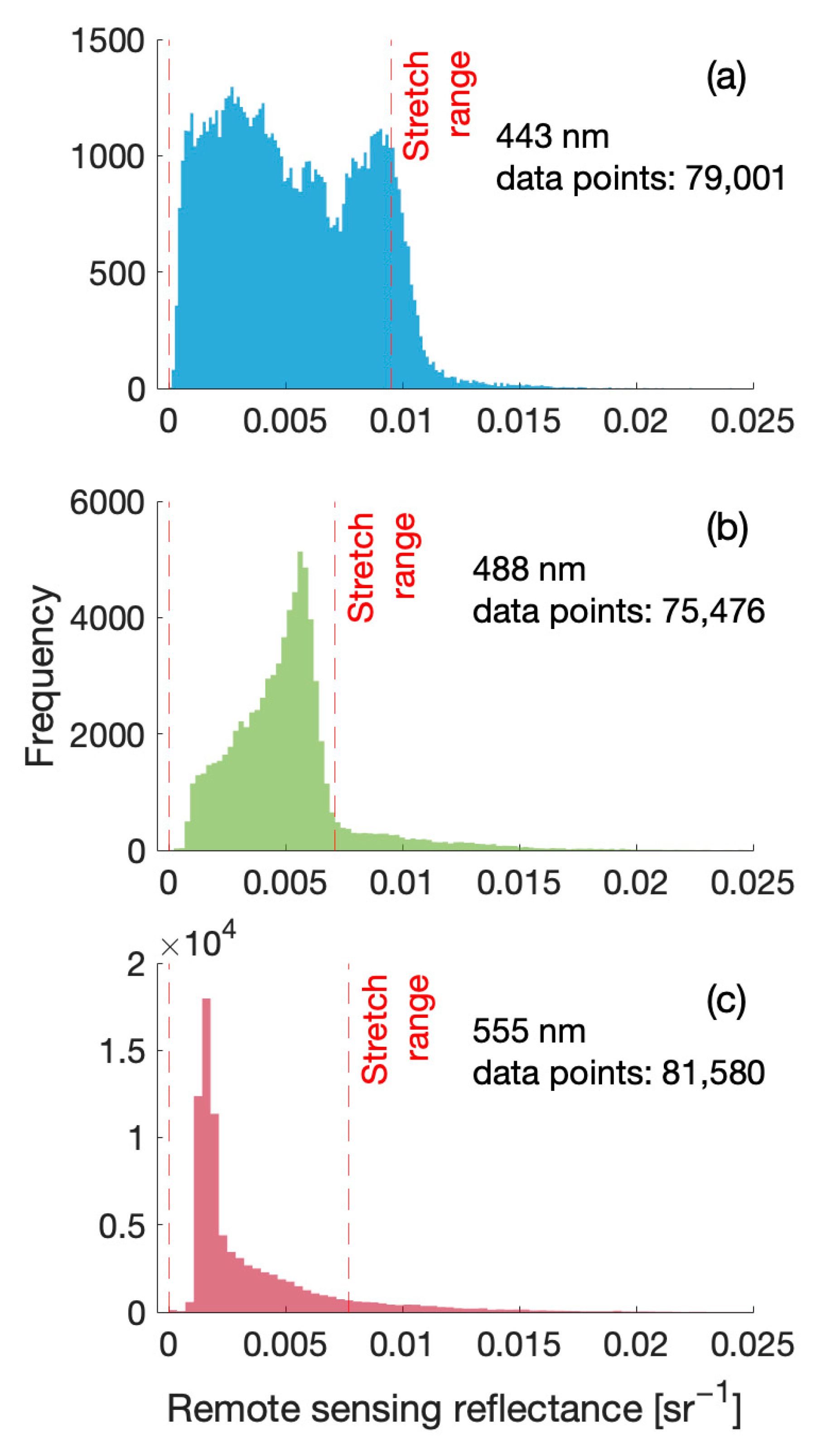
2.2. Satellite Data and eRGB Image Processing
2.3. Radiative Transfer Simulations
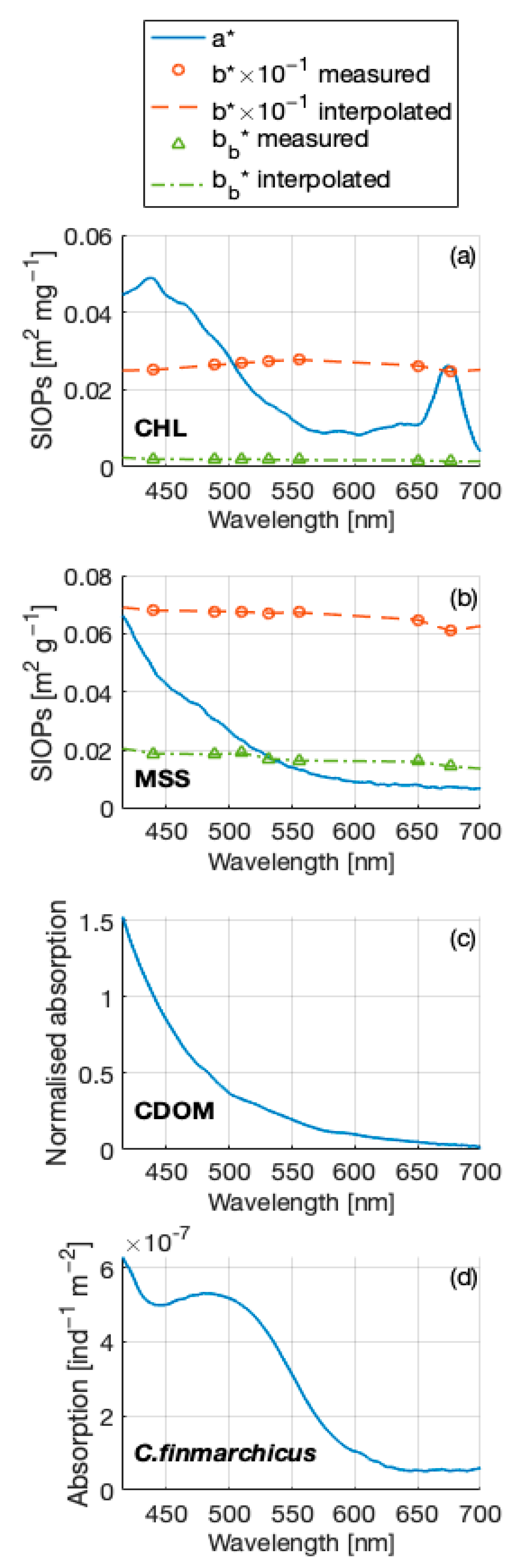
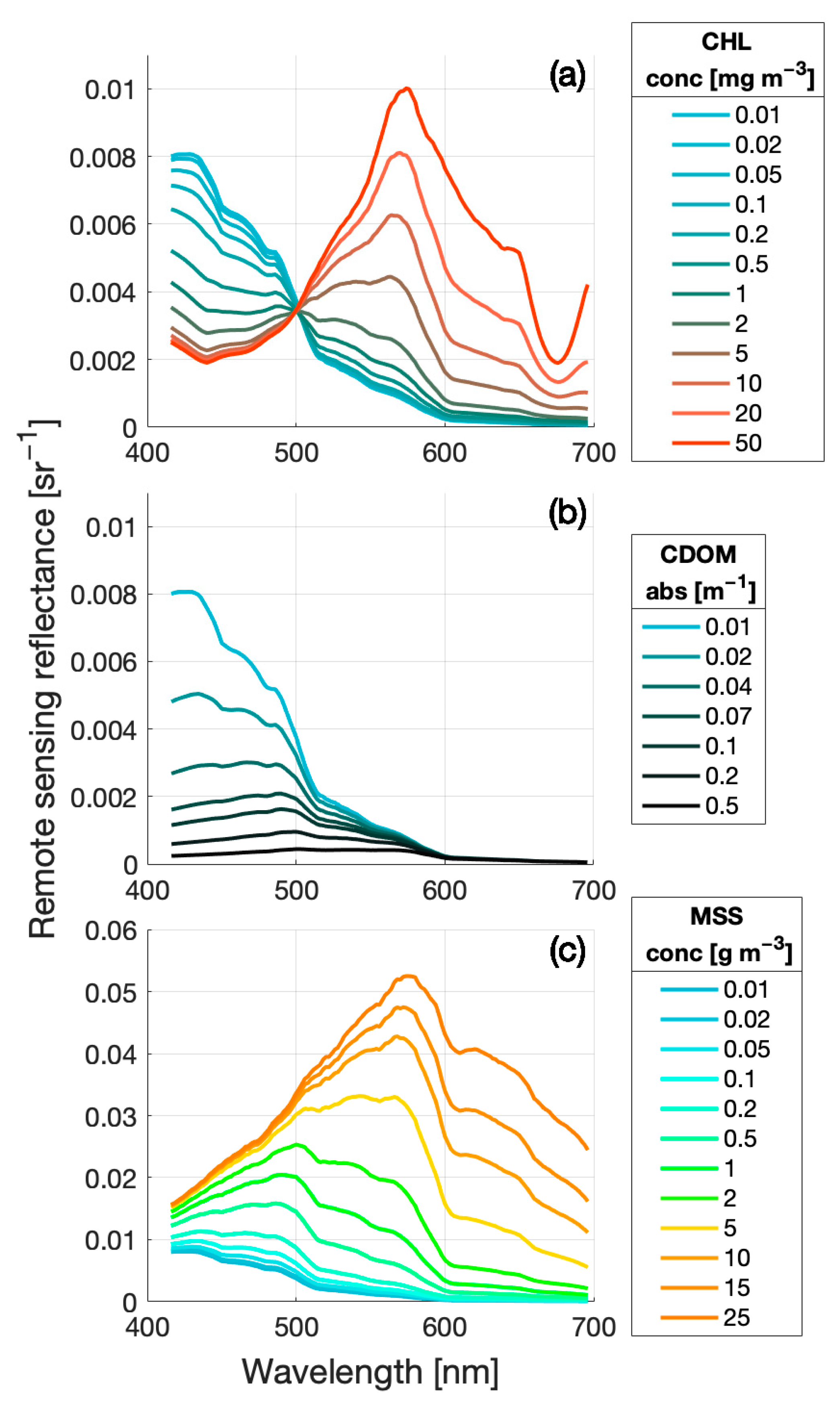
2.3.1. Bio-Optical Model Used
2.3.2. Ecolight Radiative Transfer Modelling
2.4. Quantification of Percieved Colour Difference
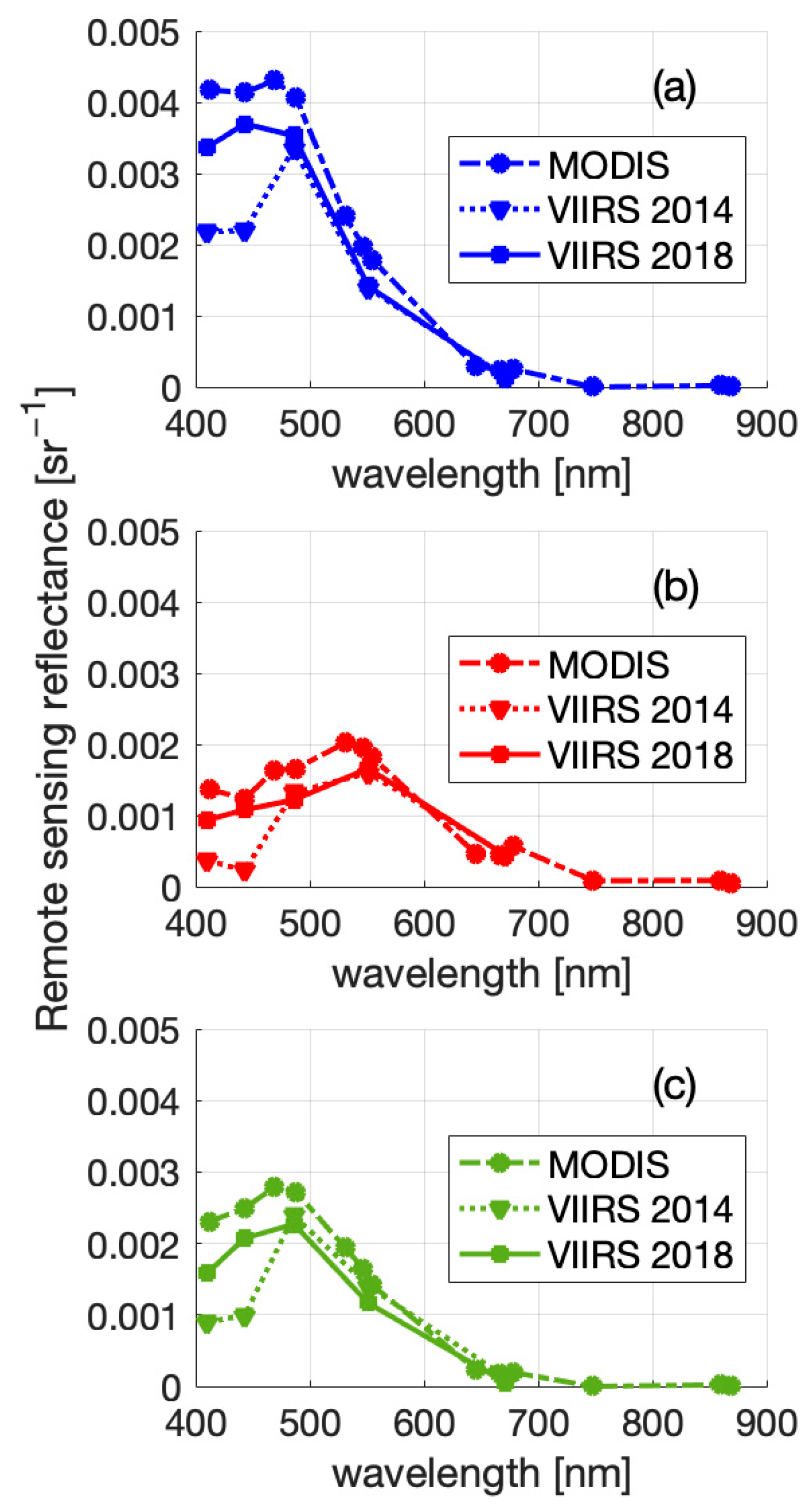
3. Results
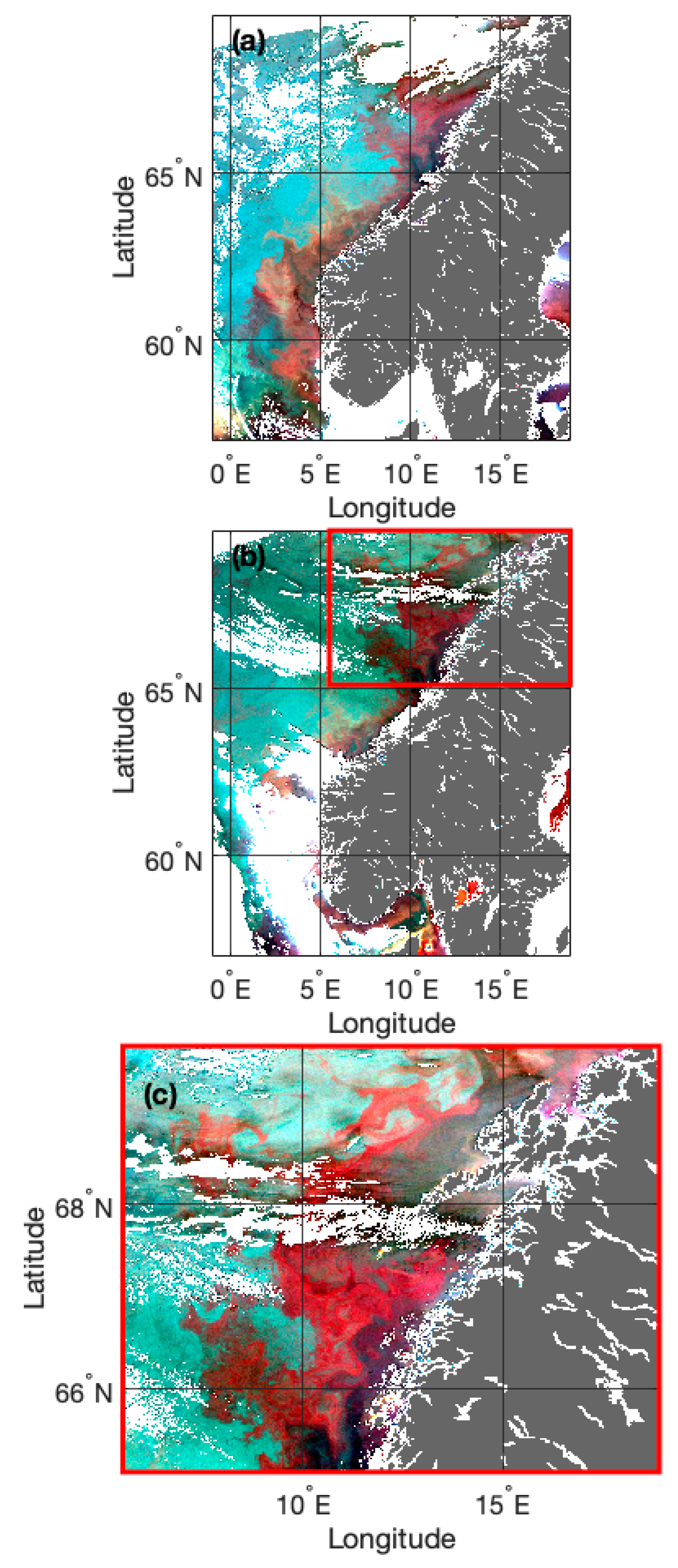
3.1. Standardised eRGB Imagery
3.2. eRGB Colour Simulation
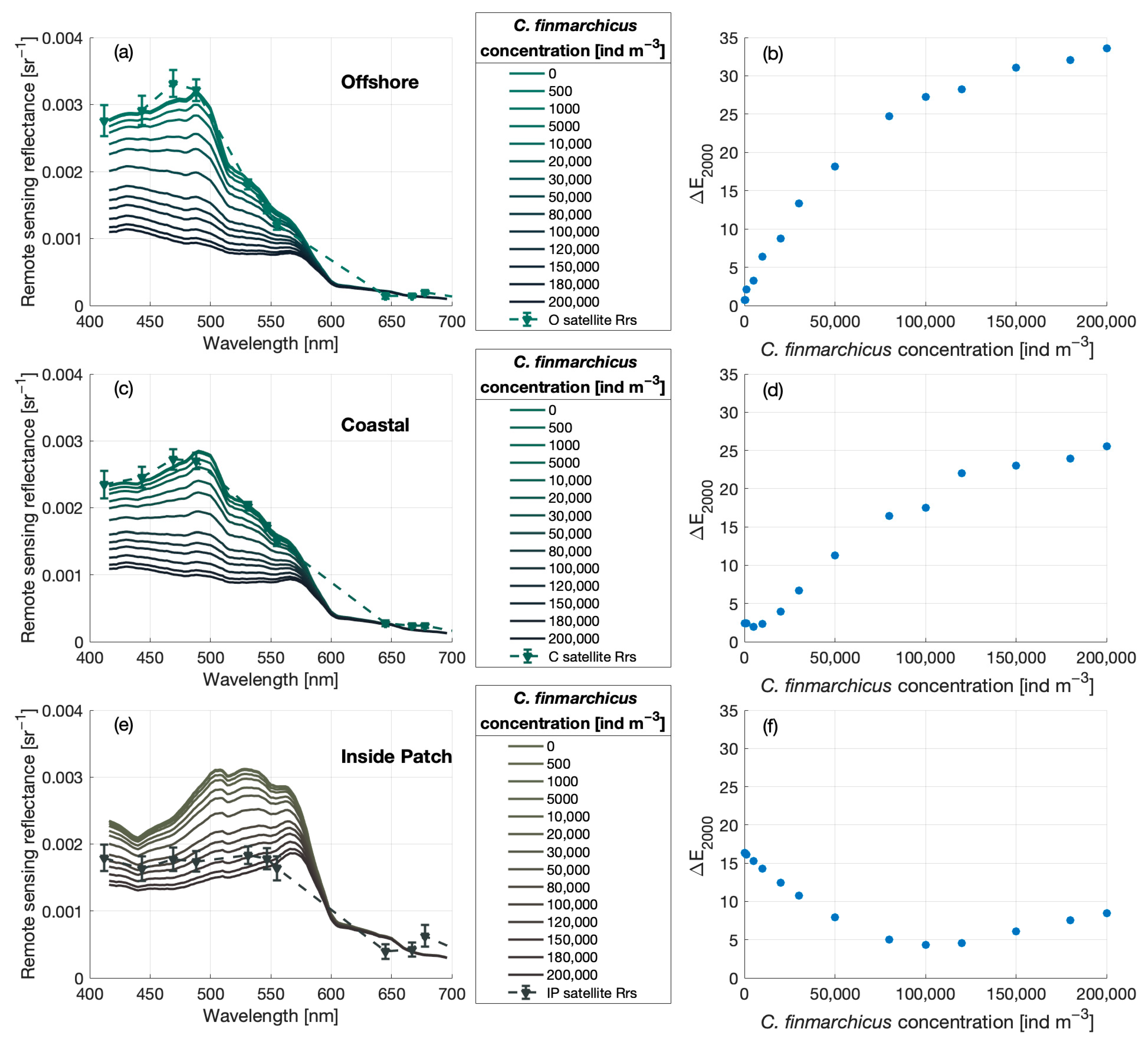
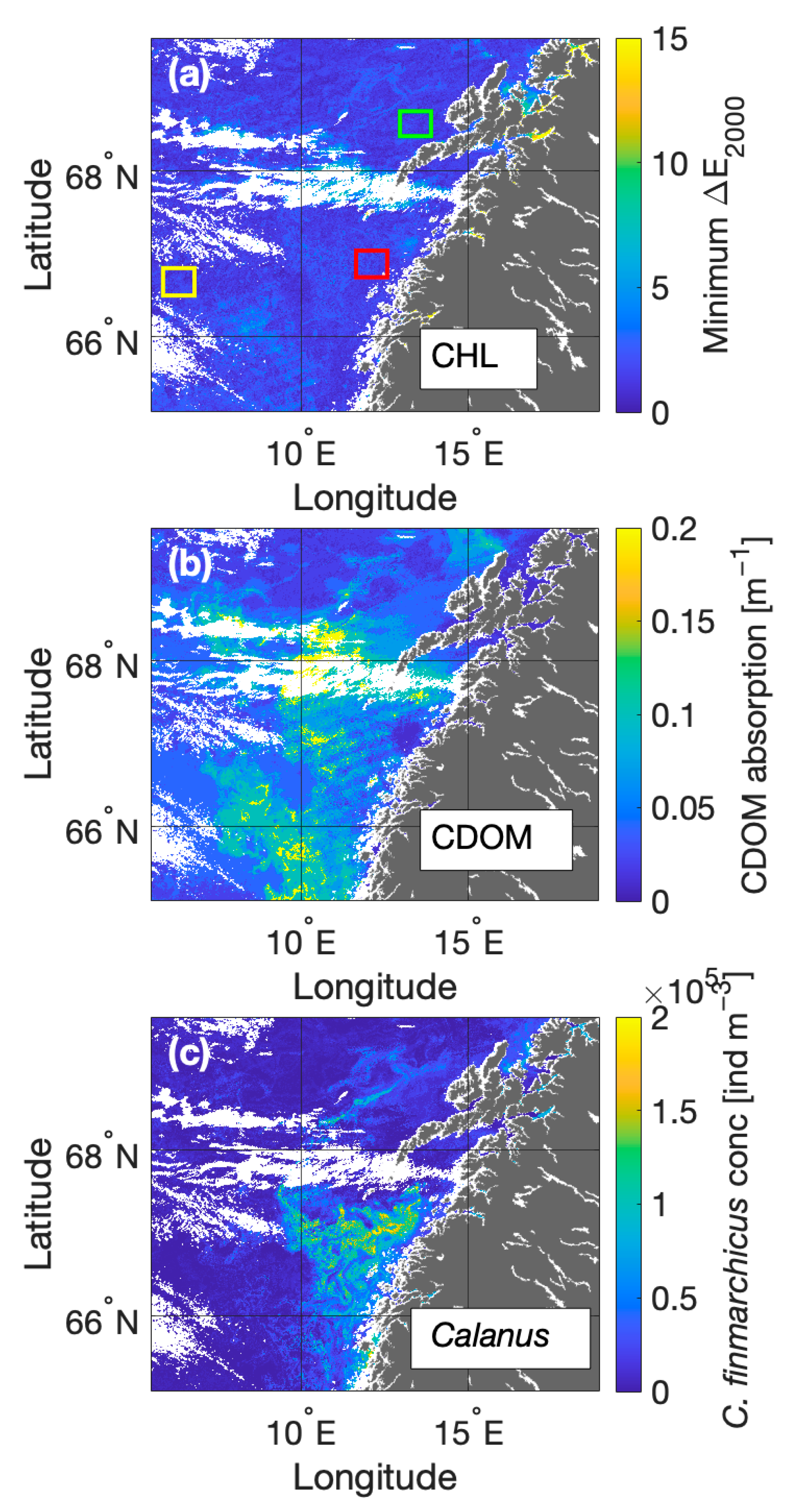
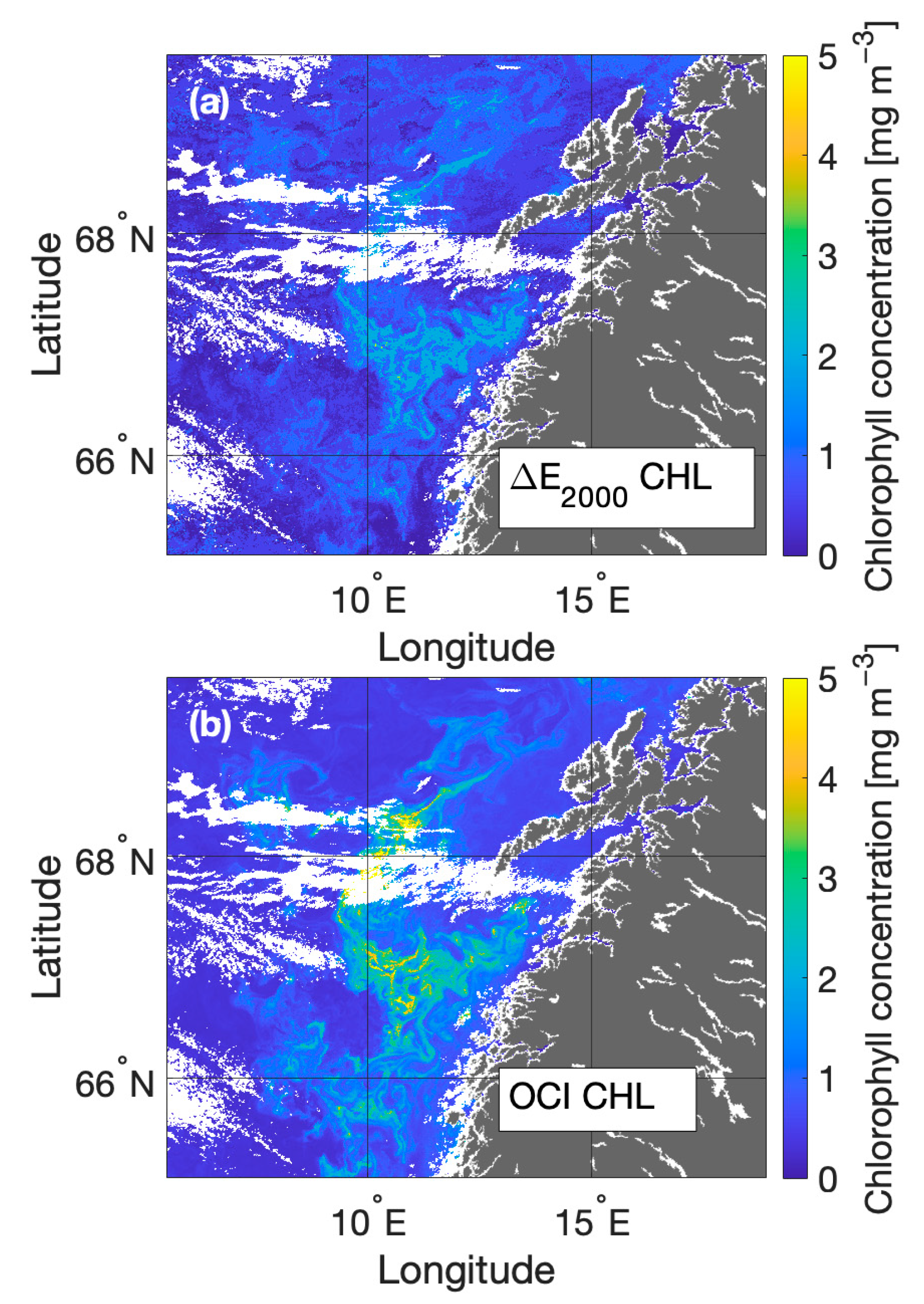
3.3. Anomaly Detection
3.4. Impact of C. finmarchicus Absorption on Reflectance Signals
Estimating Surface C. finmarchicus Concentrations from eRGB Imagery
3.5. Impact of C. finmarchicus Absorption on OC3M Algorithm Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilstone, G.H.; Pardo, S.; Dall’Olmo, G.; Brewin, R.J.W.; Nencioli, F.; Dessailly, D.; Kwiatkowska, E.; Casal, T.; Donlon, C. Performance of Ocean Colour Chlorophyll a algorithms for Sentinel-3 OLCI, MODIS-Aqua and Suomi-VIIRS in open-ocean waters of the Atlantic. Remote Sens. Environ. 2021, 260, 112444. [Google Scholar] [CrossRef]
- Blondeau-Patissier, D.; Gower, J.F.R.; Dekker, A.G.; Phinn, S.R.; Brando, V.E. A review of ocean color remote sensing methods and statistical techniques for the detection, mapping and analysis of phytoplankton blooms in coastal and open oceans. Prog. Oceanogr. 2014, 123, 123–144. [Google Scholar] [CrossRef] [Green Version]
- Falk-Petersen, S.; Mayzaud, P.; Kattner, G.; Sargent, J.R. Lipids and life strategy of Arctic Calanus. Mar. Biol. Res. 2009, 5, 18–39. [Google Scholar] [CrossRef] [Green Version]
- Melle, W.; Runge, J.; Head, E.; Plourde, S.; Castellani, C.; Licandro, P.; Pierson, J.; Jonasdottir, S.; Johnson, C.; Broms, C.; et al. The North Atlantic Ocean as habitat for Calanus finmarchicus: Environmental factors and life history traits. Prog. Oceanogr. 2014, 129, 244–284. [Google Scholar] [CrossRef] [Green Version]
- Wiborg, K.F. Fishery and commercial exploitation of Calanus finmarchicus in Norway. ICES J. Mar. Sci. 1976, 36, 251–258. Available online: https://academic.oup.com/icesjms/article/36/3/251/639243 (accessed on 8 March 2023).
- Eilertsen, K.-E.; Mæhre, H.K.; Jensen, I.J.; Devold, H.; Olsen, J.O.; Lie, R.K.; Brox, J.; Berg, V.; Elvevoll, E.O.; Østerud, B. A Wax Ester and Astaxanthin-Rich Extract from the Marine Copepod Calanus finmarchicus Attenuates Atherogenesis in Female Apolipoprotein EDeficient Mice. J. Nutr. 2012, 142, 508–512. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, A.M.; Vang, B.; Olsen, R.L. Oil from Calanus finmarchicus—Composition and Possible Use: A Review. J. Aquat. Food Prod. Technol. 2014, 23, 633–646. [Google Scholar] [CrossRef]
- Vilgrain, L.; Maps, F.; Trudnowska, E.; Basedow, S.L.; Madoui, A.; Niehoff, B.; Irisson, J.-O.; Ayata, S.-D. Copepods’ true colours: Pigmentation as an indicator of fitness. Ecosphere 2023. [Google Scholar] [CrossRef]
- Yiki, V. Biological functions and activities of animal carotenoids. Pure Appl. Chem. 1991, 63, 141–146. [Google Scholar] [CrossRef]
- Morel, A.; Prieur, L. Analysis of variations in ocean color. Limnol. Oceanogr. 1977, 22, 709–722. [Google Scholar] [CrossRef]
- Stramski, D.; Kiefer, D.A. Light scattering by microorganisms in the open ocean. Prog. Oceanogr. 1991, 28, 343–383. [Google Scholar] [CrossRef]
- Stramski, D.; Mobley, C.D. Effects of microbial particles on oceanic optics: A database of single-particle optical properties. Limnol. Oceanogr. 1997, 42, 538–549. [Google Scholar] [CrossRef] [Green Version]
- Mobley, C.D. Optical Properties of Water. In Light and Water: Radiative Transfer in Natural Waters; Mobley, C.D., Preisendorfer, R.W., Eds.; Academic Press: Cambridge, UK, 1994. [Google Scholar]
- Davies, E.J.; Basedow, S.L.; McKee, D. The hidden influence of large particles on ocean colour. Sci. Rep. 2021, 11, 3999. [Google Scholar] [CrossRef] [PubMed]
- Bullen, G. Makerel and Calanus. Nature 1913, 91, 531. [Google Scholar] [CrossRef]
- Sars, G.O. An Account of the Crustacea of Norway: Copepoda, Calanoida; University Museum of Bergen: Bergen, Norway, 1903; Volume 4. [Google Scholar]
- Basedow, S.L.; McKee, D.; Lefering, I.; Gislason, A.; Daase, M.; Trudnowska, E.; Egeland, E.S.; Choquet, M.; Falk-Petersen, S. Remote sensing of zooplankton swarms. Sci. Rep. 2019, 9, 686. [Google Scholar] [CrossRef] [Green Version]
- Weng, F.; Choi, T.; Cao, C.; Zhang, B. Reprocessing of SUOMI NPP VIIRS sensor data records and impacts on environmental applications. In Proceedings of the 2017 IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Fort Worth, TX, USA, 23–28 July 2017; pp. 293–296. [Google Scholar] [CrossRef]
- Zhu, Y.; Tande, K.S.; Zhou, M. Mesoscale physical processes and zooplankton transport-retention in the northern Norwegian shelf region. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 1922–1933. [Google Scholar] [CrossRef]
- Dong, H.; Zhou, M.; Hu, Z.; Zhang, Z.; Zhong, Y.; Basedow, S.L.; Smith, W.O., Jr. Transport Barriers and the Retention of Calanus finmarchicus on the Lofoten Shelf in Early Spring. J. Geophys. Res. Oceans 2021, 126, e2021JC017408. [Google Scholar] [CrossRef]
- Mobley, C.D.; Werdell, J.; Franz, B.; Ahmad, Z.; Bailey, S. Atmospheric Correction for Satellite Ocean Color Radiometry; National Aeronautics and Space Administration: Greenbelt, MD, USA, 2016.
- Hu, C.; Feng, L.; Lee, Z.; Franz, B.A.; Bailey, S.W.; Werdell, P.J.; Proctor, C.W. Improving Satellite Global Chlorophyll a Data Products through Algorithm Refinement and Data Recovery. J. Geophys. Res. Oceans 2019, 124, 1524–1543. [Google Scholar] [CrossRef]
- Hu, C.; Lee, Z.; Franz, B. Chlorophyll a algorithms for oligotrophic oceans: A novel approach based on three-band reflectance difference. J. Geophys. Res. Oceans 2012, 117, C01011. [Google Scholar] [CrossRef] [Green Version]
- Bengil, F.; McKee, D.; Beşiktepe, S.T.; Calzado, V.S.; Trees, C. A bio-optical model for integration into ecosystem models for the Ligurian Sea. Prog. Oceanogr. 2016, 149, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zaneveld, J.R.V.; Kitchen, J.C.; Moore, C.C. Scattering Error Correction of Reflecting-Tube Absorption Meters. In Proceedings of the SPIE 2258, Ocean Optics XII, Bergen, Norway, 26 October 1994; pp. 44–55. [Google Scholar] [CrossRef]
- Lefering, I.; Bengil, F.; Trees, C.; Röttgers, R.; Bowers, D.; Nimmo-Smith, A.; Schwarz, J.; McKee, D. Optical closure in marine waters from in situ inherent optical property measurements. Opt. Express 2016, 24, 14036–14052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, D.; Röttgers, R.; Neukermans, G.; Calzado, V.S.; Trees, C.; Ampolo-Rella, M.; Neil, C.; Cunningham, A. Impact of measurement uncertainties on determination of chlorophyll-specific absorption coefficient for marine phytoplankton. J. Geophys. Res. Oceans 2014, 119, 9013–9025. [Google Scholar] [CrossRef] [Green Version]
- Prejato, M.L.; McKee, D.; Mitchell, C. Inherent Optical Properties-Reflectance Relationships Revisited. J. Geophys. Res. Oceans 2020, 125, e2020JC016661. [Google Scholar] [CrossRef]
- Luo, M.R.; Cui, G.; Rigg, B. The development of the CIE 2000 colour-difference formula: CIEDE2000. Color Res. Appl. 2001, 26, 340–350. [Google Scholar] [CrossRef]
- Sharma, G.; Wu, W.; Dalal, E.N. The CIEDE2000 Color-Difference Formula: Implementation Notes, Supplementary Test Data, and Mathematical Observations. Color Res. Appl. 2005, 30, 21–30. [Google Scholar] [CrossRef]
- Liu, H.X.; Wu, B.; Liu, Y.; Huang, M.; Xu, Y.F. A Discussion on Printing Color Difference Tolerance by CIEDE2000 Color Difference Formula. Appl. Mech. Mater. 2013, 262, 96–99. [Google Scholar] [CrossRef]
- Valente, A.; Sathyendranath, S.; Brotas, V.; Groom, S.; Grant, M.; Jackson, T.; Chuprin, A.; Taberner, M.; Airs, R.; Antoine, D.; et al. A compilation of global bio-optical in situ data for ocean colour satellite applications—Version three. Earth Syst. Sci. Data 2022, 14, 5737–5770. [Google Scholar] [CrossRef]
- Madsen, S.; Nielsen, T.; Tervo, O.; Söderkvist, J. Importance of feeding for egg production in Calanus finmarchicus and C. glacialis during the Arctic spring. Mar. Ecol. Prog. Ser. 2008, 353, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Valente, A.; Sathyendranath, S.; Brotas, V.; Groom, S.; Grant, M.; Taberner, M.; Antoine, D.; Arnone, R.; Balch, W.M.; Barker, K.; et al. A compilation of global bio-optical in situ data for ocean-colour satellite applications. Earth Syst. Sci. Data 2016, 8, 235–252. [Google Scholar] [CrossRef] [Green Version]
- Niehoff, B.; Klenke, U.; Hirche, H.-J.; Irigoien, X.; Head, R.; Harris, R. A high frequency time series at weathership M, Norwegian Sea, during the 1997 spring bloom: Feeding of adult female Calanus finmarchicus. Mar. Ecol. Prog. Ser. 1999, 176, 81–92. [Google Scholar] [CrossRef]
- Berge, J.; Geoffroy, M.; Daase, M.; Cottier, F.; Priou, P.; Cohen, J.H.; Johnsen, G.; McKee, D.; Kostakis, I.; Renaud, P.E.; et al. Artificial light during the polar night disrupts Arctic fish and zooplankton behaviour down to 200 m depth. Commun. Biol. 2020, 3, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandara, K.; Basedow, S.L.; Pedersen, G.; Tverberg, V. Mid-summer vertical behavior of a high-latitude oceanic zooplankton community. J. Mar. Syst. 2022, 230, 103733. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCarry, C.L.; Basedow, S.L.; Davies, E.J.; McKee, D. Estimating Surface Concentrations of Calanus finmarchicus Using Standardised Satellite-Derived Enhanced RGB Imagery. Remote Sens. 2023, 15, 2987. https://doi.org/10.3390/rs15122987
McCarry CL, Basedow SL, Davies EJ, McKee D. Estimating Surface Concentrations of Calanus finmarchicus Using Standardised Satellite-Derived Enhanced RGB Imagery. Remote Sensing. 2023; 15(12):2987. https://doi.org/10.3390/rs15122987
Chicago/Turabian StyleMcCarry, Cait L., Sünnje L. Basedow, Emlyn J. Davies, and David McKee. 2023. "Estimating Surface Concentrations of Calanus finmarchicus Using Standardised Satellite-Derived Enhanced RGB Imagery" Remote Sensing 15, no. 12: 2987. https://doi.org/10.3390/rs15122987