


Recent Cereal Phenological Variations under Mediterranean Conditions

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Source
2.2.1. Detection of Cereal Zones
2.2.2. Remote Sensing Data and Processing
2.2.3. Hydroclimatic Data
2.3. Data Analyses
2.3.1. Phenology Parameter Extraction
2.3.2. Trend Analysis
2.3.3. Correlation Analysis
3. Results
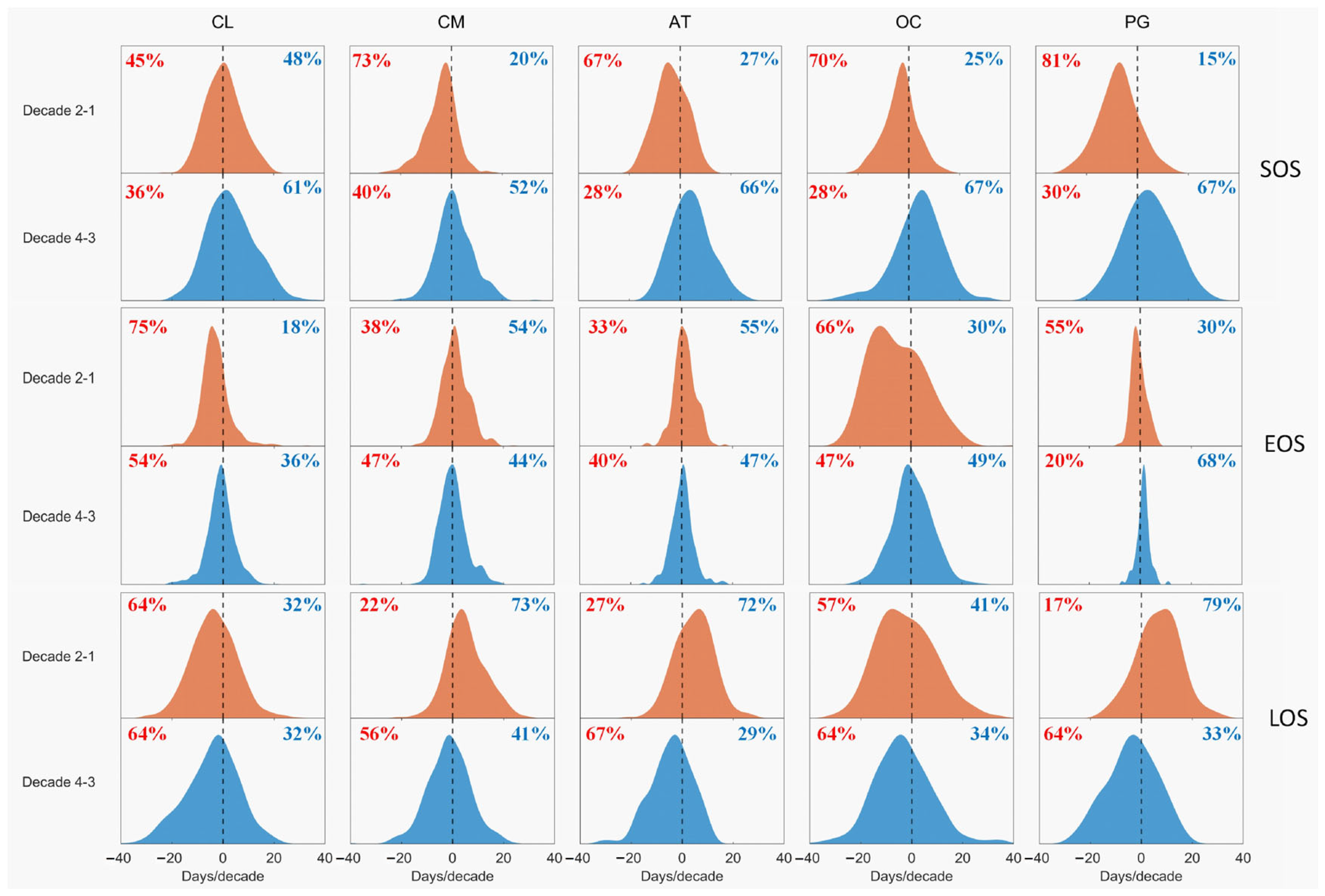
3.1. Phenological Dynamics over Decades
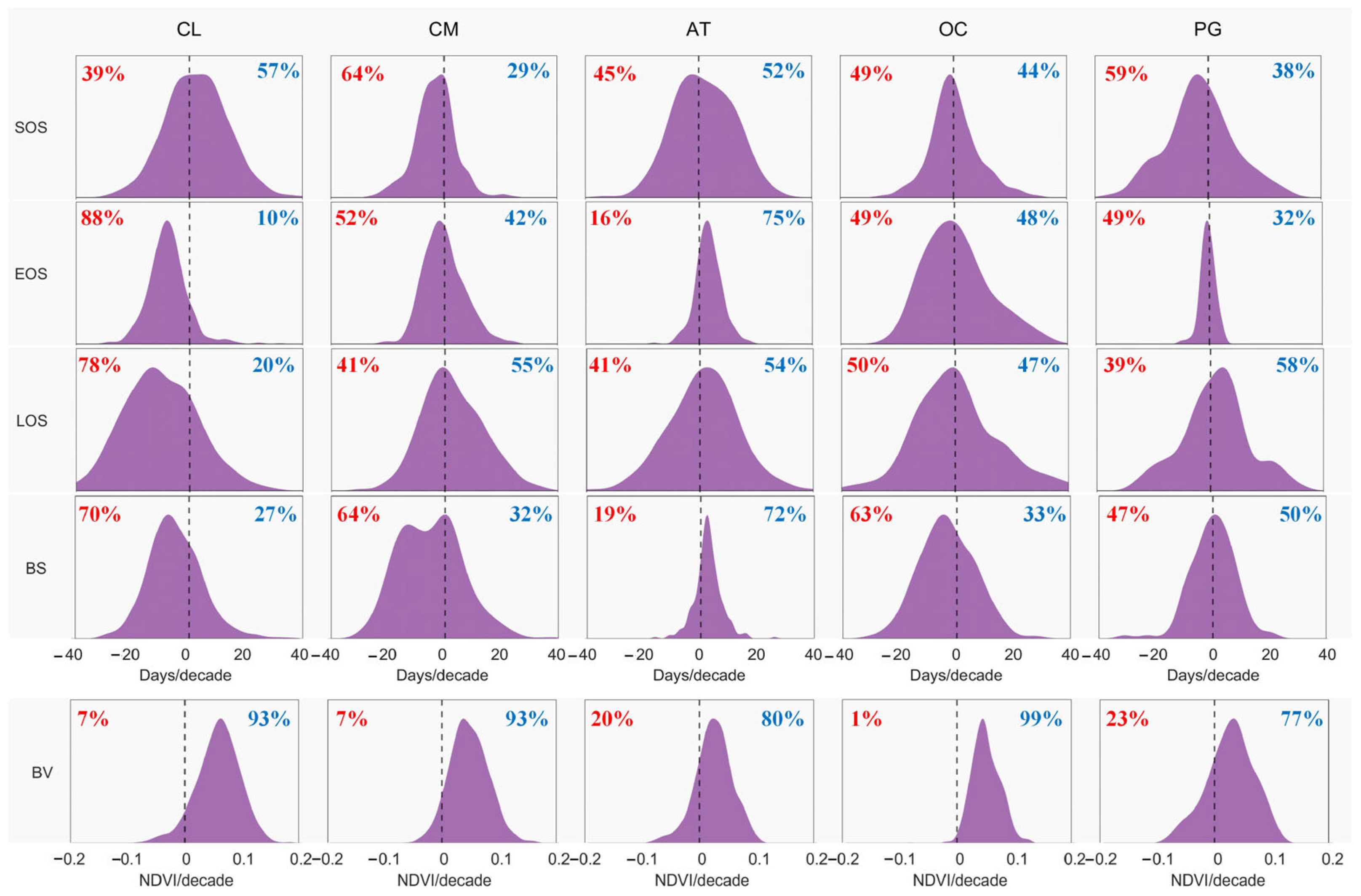
3.2. Temporal Patterns of Phenological Trends
3.3. Relationships between the Phenological Parameters of Vegetation
3.4. Influence of Hydroclimatic Variables on Phenological Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, H.; Zhang, Z.; Guo, X.; Li, X.; Chen, C. Spatial and temporal changes in vegetation phenology at middle and high latitudes of the Northern Hemisphere over the past three decades. Remote Sens. 2015, 7, 10973–10995. [Google Scholar] [CrossRef]
- Gu, L.; Chen, J.; Yin, J.; Sullivan, S.C.; Wang, H.-M.; Guo, S.; Zhang, L.; Kim, J.-S. Projected increases in magnitude and socioeconomic exposure of global droughts in 1.5 and 2 °C warmer climates. Hydrol. Earth Syst. Sci. 2020, 24, 451–472. [Google Scholar] [CrossRef]
- Tramblay, Y.; Koutroulis, A.; Samaniego, L.; Vicente-Serrano, S.M.; Volaire, F.; Boone, A.; Le Page, M.; Llasat, M.C.; Albergel, C.; Burak, S.; et al. Challenges for drought assessment in the Mediterranean region under future climate scenarios. Earth-Sci. Rev. 2020, 210, 103348. [Google Scholar] [CrossRef]
- Benito-Verdugo, P.; Martínez-Fernández, J.; González-Zamora, Á.; Almendra-Martín, L.; Gaona, J.; Herrero-Jiménez, C.M. Impact of Agricultural Drought on Barley and Wheat Yield: A Comparative Case Study of Spain and Germany. Agriculture 2023, 13, 2111. [Google Scholar] [CrossRef]
- Jacobsen, S.-E.; Jensen, C.R.; Liu, F. Improving crop production in the arid Mediterranean climate. Field Crop. Res. 2012, 128, 34–47. [Google Scholar] [CrossRef]
- Jiao, F.; Liu, H.; Xu, X.; Gong, H.; Lin, Z. Trend evolution of vegetation phenology in China during the period of 1981–2016. Remote Sens. 2020, 12, 572. [Google Scholar] [CrossRef]
- Zhan, W.; Luo, F.; Luo, H.; Li, J.; Wu, Y.; Yin, Z.; Wu, Y.; Wu, P. Time-Series-Based Spatiotemporal Fusion Network for Improving Crop Type Mapping. Remote Sens. 2024, 16, 235. [Google Scholar] [CrossRef]
- Meng, L.; Zhou, Y.; Gu, L.; Richardson, A.D.; Peñuelas, J.; Fu, Y.; Wang, Y.; Asrar, G.R.; De Boeck, H.J.; Mao, J.; et al. Photoperiod decelerates the advance of spring phenology of six deciduous tree species under climate warming. Glob. Change Biol. 2021, 27, 2914–2927. [Google Scholar] [CrossRef]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Liao, C.; Wang, J.; Shan, B.; Shang, J.; Dong, T.; He, Y. Near real-time detection and forecasting of within-field phenology of winter wheat and corn using Sentinel-2 time-series data. ISPRS J. Photogramm. Remote Sens. 2023, 196, 105–119. [Google Scholar] [CrossRef]
- Kibret, K.S.; Marohn, C.; Cadisch, G. Use of MODIS EVI to map crop phenology, identify cropping systems, detect land use change and drought risk in Ethiopia—An application of Google Earth Engine. Eur. J. Remote Sens. 2020, 53, 176–191. [Google Scholar] [CrossRef]
- Gerard, F.F.; George, C.T.; Hayman, G.; Chavana-Bryant, C.; Weedon, G.P. Leaf phenology amplitude derived from MODIS NDVI and EVI: Maps of leaf phenology synchrony for Meso-and South America. Geosci. Data J. 2020, 7, 13–26. [Google Scholar] [CrossRef]
- You, X.; Meng, J.; Zhang, M.; Dong, T. Remote sensing based detection of crop phenology for agricultural zones in China using a new threshold method. Remote Sens. 2013, 5, 3190–3211. [Google Scholar] [CrossRef]
- Tian, R.; Li, J.; Zheng, J.; Liu, L.; Liu, Y.; Han, W.; Wang, X. The spatial-temporal patterns of spring phenology in the temperate grasslands of China and their response mechanisms to climatic factors. J. Spat. Sci. 2024, 1–19. [Google Scholar] [CrossRef]
- Meroni, M.; Verstraete, M.M.; Rembold, F.; Urbano, F.; Kayitakire, F. A phenology-based method to derive biomass production anomalies for food security monitoring in the Horn of Africa. Int. J. Remote Sens. 2014, 35, 2472–2492. [Google Scholar] [CrossRef]
- Fu, Y.; He, H.S.; Zhao, J.; Larsen, D.R.; Zhang, H.; Sunde, M.G.; Duan, S. Climate and spring phenology effects on autumn phenology in the Greater Khingan Mountains, Northeastern China. Remote Sens. 2018, 10, 449. [Google Scholar] [CrossRef]
- Gordo, O.; Sanz, J.J. Impact of climate change on plant phenology in Mediterranean ecosystems. Glob. Change Biol. 2010, 16, 1082–1106. [Google Scholar] [CrossRef]
- Menzel, A.; Yuan, Y.; Matiu, M.; Sparks, T.; Scheifinger, H.; Gehrig, R.; Estrella, N. Climate change fingerprints in recent European plant phenology. Glob. Change Biol. 2020, 26, 2599–2612. [Google Scholar] [CrossRef]
- Jin, H.; Jönsson, A.M.; Olsson, C.; Lindström, J.; Jönsson, P.; Eklundh, L. New satellite-based estimates show significant trends in spring phenology and complex sensitivities to temperature and precipitation at Northern European latitudes. Int. J. Biometeorol. 2019, 63, 763–775. [Google Scholar] [CrossRef]
- Yuan, M.; Wang, L.; Lin, A.; Liu, Z.; Qu, S. Variations in land surface phenology and their response to climate change in Yangtze River basin during 1982–2015. Theor. Appl. Climatol. 2019, 137, 1659–1674. [Google Scholar] [CrossRef]
- Stöckli, R.; Vidale, P.L. European plant phenology and climate as seen in a 20-Year AVHRR land-surface parameter dataset. Int. J. Remote Sens. 2004, 25, 3303–3330. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Zhou, L.; Ciais, P.; Zhu, B. Variations in satellite-derived phenology in China’s temperate vegetation. Glob. Chang. Biol. 2006, 12, 672–685. [Google Scholar] [CrossRef]
- Jeong, S.-J.; HO, C.-H.; GIM, H.-J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Chang. Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Cong, N.; Wang, T.; Nan, H.; Ma, Y.; Wang, X.; Myneni, R.B.; Piao, S. Changes in satellite-derived spring vegetation green-up date and its linkage to climate in China from 1982 to 2010: A multimethod analysis. Glob. Chang. Biol. 2013, 19, 881–891. [Google Scholar] [CrossRef]
- Fu, Y.H.; Piao, S.; Op de Beeck, M.; Cong, N.; Zhao, H.; Zhang, Y.; Menzel, A.; Janssens, I.A. Recent spring phenology shifts in western Central Europe based on multiscale observations. Glob. Ecol. Biogeogr. 2014, 23, 1255–1263. [Google Scholar] [CrossRef]
- Touhami, I.; Moutahir, H.; Assoul, D.; Bergaoui, K.; Aouinti, H.; Bellot, J.; Andreu, J.M. Multi-year monitoring land surface phenology in relation to climatic variables using MODIS-NDVI time-series in Mediterranean forest, Northeast Tunisia. Acta Oecol. 2022, 114, 103804. [Google Scholar] [CrossRef]
- Zhang, R.; Zhou, Y.; Hu, T.; Sun, W.; Zhang, S.; Wu, J.; Wang, H. Detecting the Spatiotemporal Variation of Vegetation Phenology in Northeastern China Based on MODIS NDVI and Solar-Induced Chlorophyll Fluorescence Dataset. Sustainability 2023, 15, 6012. [Google Scholar] [CrossRef]
- Zhu, W.; Tian, H.; Xu, X.; Pan, Y.; Chen, G.; Lin, W. Extension of the growing season due to delayed autumn over mid and high latitudes in North America during 1982–2006. Glob. Ecol. Biogeogr. 2012, 21, 260–271. [Google Scholar] [CrossRef]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Liu, L.; Lian, X.; Shen, M.; Zhu, X. Plant phenology and global climate change: Current progresses and challenges. Glob. Chang. Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef]
- Guo, J.; Hu, Y. Spatiotemporal Variations in Satellite-Derived Vegetation Phenological Parameters in Northeast China. Remote Sens. 2022, 14, 705. [Google Scholar] [CrossRef]
- Ren, S.; An, S. Temporal Pattern Analysis of Cropland Phenology in Shandong Province of China Based on Two Long-Sequence Remote Sensing Data. Remote Sens. 2021, 13, 4071. [Google Scholar] [CrossRef]
- Measho, S.; Li, F.; Chen, G.; Hirwa, H. Characterizing Cropland Patterns Across North-East Africa Using Time Series Vegetation Indices. J. Geophys. Res. Biogeosci. 2023, 128, e2022JG007075. [Google Scholar] [CrossRef]
- Savin, R.; Cossani, C.M.; Dahan, R.; Ayad, J.Y.; Albrizio, R.; Todorovic, M.; Karrou, M.; Slafer, G.A. Intensifying cereal management in dryland Mediterranean agriculture: Rainfed wheat and barley responses to nitrogen fertilisation. Eur. J. Agron. 2022, 137, 126518. [Google Scholar] [CrossRef]
- Mefleh, M. Cereals of the Mediterranean region: Their origin, breeding history and grain Quality Traits. In Cereal-Based Foodstuffs: The Backbone of Mediterranean Cuisine; Boukid, F., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 1–18. ISBN 978-3-030-69228-5. [Google Scholar]
- MAPA. Anuario de Estadística; Ministerio de Agricultura Pesca y Alimentación (MAPA): Madrid, Spain, 2023. [Google Scholar]
- Stoate, C.; Borralho, R.; Araújo, M. Factors affecting corn bunting Miliaria calandra abundance in a Portuguese agricultural landscape. Agric. Ecosyst Environ. 2000, 77, 219–226. [Google Scholar] [CrossRef]
- García-León, D.; López-Lozano, R.; Toreti, A.; Zampieri, M. Local-scale cereal yield forecasting in Italy: Lessons from different statistical models and spatial aggregations. Agronomy 2020, 10, 809. [Google Scholar] [CrossRef]
- Statistique Agricole Annuelle 2022 et Nouvelles Séries 2010–2022. DRAAF Occitanie. Available online: https://draaf.occitanie.agriculture.gouv.fr/statistique-agricole-annuelle-2022-et-nouvelles-series-2010-2022-a7672.html (accessed on 12 April 2024).
- Di Gregorio, A. Land Cover Classification System: Classification Concepts and User Manual: Software Version 2; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2005; ISBN 92-5-105327-8. [Google Scholar]
- Defourny, P.; Kirches, G.; Brockmann, C.; Boettcher, M.; Peters, M.; Bontemps, S.; Lamarche, C.; Schlerf, M.; Santoro, M. Land Cover CCI: Product User Guide Version 2; European Space Agency (ESA): Louvain-la-Neuve, Belgium, 2012. [Google Scholar]
- Siebert, S.; Henrich, V.; Frenken, K.; Burke, J. Update of the Digital Global Map of Irrigation Areas to Version 5; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013. [Google Scholar]
- Pinzon, J.E.; Pak, E.W.; Tucker, C.J.; Bhatt, U.S.; Frost, G.V.; Macander, M.J. Glob. Vegetation Greenness (NDVI) from AVHRR GIMMS-3G+, 1981–2022; ORNL DAAC: Oak Ridge, TN, USA, 2023. [Google Scholar] [CrossRef]
- Wolberg, G.; Alfy, I. Monotonic cubic spline interpolation. In Proceedings of the Computer Graphics International, Canmore, AB, Canada, 7–11 June 1999; ISBN 0-7695-0185-0. [Google Scholar]
- Talebi, H.; Samadianfard, S.; Valizadeh Kamran, K. Estimation of daily reference evapotranspiration implementing satellite image data and strategy of ensemble optimization algorithm of stochastic gradient descent with multilayer perceptron. Environ. Dev. Sustain. 2023. [Google Scholar] [CrossRef]
- Bandhauer, M.; Isotta, F.; Lakatos, M.; Lussana, C.; Båserud, L.; Izsák, B.; Szentes, O.; Tveito, O.E.; Frei, C. Evaluation of daily precipitation analyses in E-OBS (V19.0e) and ERA5 by comparison to regional high-resolution datasets in European regions. Int. J. Climatol. 2022, 42, 727–747. [Google Scholar] [CrossRef]
- Yoder, R.E.; Odhiambo, L.O.; Wright, W.C. Effects of vapor-pressure deficit and net-irradiance calculation methods on accuracy of standardized Penman-Monteith equation in a humid climate. J. Irrig. Drain. Eng. 2005, 131, 228–237. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Requirements; FAO Irrigation and Drainage Paper, 56; FAO: Rome, Italy, 1998. [Google Scholar]
- Muñoz-Sabater, J.; Dutra, E.; Agustí-Panareda, A.; Albergel, C.; Arduini, G.; Balsamo, G.; Boussetta, S.; Choulga, M.; Harrigan, S.; Hersbach, H.; et al. ERA5-Land: A state-of-the-art global reanalysis dataset for land applications. Earth Syst. Sci. Data 2021, 13, 4349–4383. [Google Scholar] [CrossRef]
- González-Zamora, Á.; Almendra-Martín, L.; de Luis, M.; Gaona, J.; Martínez-Fernández, J. How Are Pine Species Responding to Soil Drought and Climate Change in the Iberian Peninsula? Forests 2023, 14, 1530. [Google Scholar] [CrossRef]
- Almendra-Martín, L.; Martínez-Fernández, J.; Piles, M.; González-Zamora, Á.; Benito-Verdugo, P.; Gaona, J. Influence of atmospheric patterns on soil moisture dynamics in Europe. Sci. Total Environ. 2022, 846, 157537. [Google Scholar] [CrossRef]
- Gaona, J.; Benito-Verdugo, P.; Martínez-Fernández, J.; González-Zamora, Á.; Almendra-Martín, L.; Herrero-Jiménez, C.M. Soil Moisture Outweighs Climatic Factors in Critical Periods for Rainfed Cereal Yields: An Analysis in Spain. Agriculture 2022, 12, 533. [Google Scholar] [CrossRef]
- Huang, X.; Liu, J.; Zhu, W.; Atzberger, C.; Liu, Q. The Optimal Threshold and Vegetation Index Time Series for Retrieving Crop Phenology Based on a Modified Dynamic Threshold Method. Remote Sens. 2019, 11, 2725. [Google Scholar] [CrossRef]
- White, M.A.; Thornton, P.E.; Running, S.W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Glob. Biogeochem. Cycles 1997, 11, 217–234. [Google Scholar] [CrossRef]
- Jonsson, P.; Eklundh, L. Seasonality extraction by function fitting to time-series of satellite sensor data. IEEE Trans. Geosci. Remote Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- MAPA. Calendario de Siembra, Recolección y Comercialización; Ministerio de Agricultura Pesca y Alimentación (MAPA): Madrid, Spain, 2023. [Google Scholar]
- Morais, T.G.; Silva, C.; Jebari, A.; Álvaro-Fuentes, J.; Domingos, T.; Teixeira, R.F. A proposal for using process-based soil models for land use Life cycle impact assessment: Application to Alentejo, Portugal. J. Clean. Prod. 2018, 192, 864–876. [Google Scholar] [CrossRef]
- Manfron, G.; Delmotte, S.; Busetto, L.; Hossard, L.; Ranghetti, L.; Brivio, P.A.; Boschetti, M. Estimating inter-annual variability in winter wheat sowing dates from satellite time series in Camargue, France. Int. J. Appl. Earth Obs. Geoinf. 2017, 57, 190–201. [Google Scholar] [CrossRef]
- Yang, C.; Fraga, H.; van Ieperen, W.; Santos, J.A. Assessing the impacts of recent-past climatic constraints on potential wheat yield and adaptation options under Mediterranean climate in southern Portugal. Agric. Syst. 2020, 182, 102844. [Google Scholar] [CrossRef]
- Ventrella, D.; Stellacci, A.M.; Castrignano, A.; Charfeddine, M.; Castellini, M. Effects of crop residue management on winter durum wheat productivity in a long term experiment in Southern Italy. Eur. J. Agron. 2016, 77, 188–198. [Google Scholar] [CrossRef]
- Meyer, N.; Bergez, J.-E.; Constantin, J.; Belleville, P.; Justes, E. Cover crops reduce drainage but not always soil water content due to interactions between rainfall distribution and management. Agric. Water Manag. 2020, 231, 105998. [Google Scholar] [CrossRef]
- Ersi, C.; Bayaer, T.; Bao, Y.; Bao, Y.; Yong, M.; Lai, Q.; Zhang, X.; Zhang, Y. Comparison of Phenological Parameters Extracted from SIF, NDVI and NIRv Data on the Mongolian Plateau. Remote Sens. 2023, 15, 187. [Google Scholar] [CrossRef]
- Kern, A.; Marjanović, H.; Barcza, Z. Spring vegetation green-up dynamics in Central Europe based on 20-Year Long MODIS NDVI data. Agric. For. Meteorol. 2020, 287, 107969. [Google Scholar] [CrossRef]
- Pan, Z.; Huang, J.; Zhou, Q.; Wang, L.; Cheng, Y.; Zhang, H.; Blackburn, G.A.; Yan, J.; Liu, J. Mapping crop phenology using NDVI time-series derived from HJ-1 A/B data. Int. J. Appl. Earth Obs. Geoinf. 2015, 34, 188–197. [Google Scholar] [CrossRef]
- Benedetti, R.; Rossini, P. On the use of NDVI profiles as a tool for agricultural statistics: The case study of wheat yield estimate and forecast in Emilia Romagna. Remote Sens. Environ. 1993, 45, 311–326. [Google Scholar] [CrossRef]
- Mann, H.B. Nonparametric Tests against Trend. Econometrica 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods; Griffin: London, UK, 1948. [Google Scholar]
- Karkauskaite, P.; Tagesson, T.; Fensholt, R. Evaluation of the Plant Phenology Index (PPI), NDVI and EVI for Start-of-Season Trend Analysis of the Northern Hemisphere Boreal Zone. Remote Sens. 2017, 9, 485. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Comas, P. Changed plant and animal life cycles from 1952 to 2000 in the Mediterranean region. Glob. Chang. Biol. 2002, 8, 531–544. [Google Scholar] [CrossRef]
- Fan, J.; Min, J.; Yang, Q.; Na, J.; Wang, X. Spatial-Temporal Relationship Analysis of Vegetation Phenology and Meteorological Parameters in an Agro-Pasture Ecotone in China. Remote Sens. 2022, 14, 5417. [Google Scholar] [CrossRef]
- Liu, Y.; Shen, X.; Zhang, J.; Wang, Y.; Wu, L.; Ma, R.; Lu, X.; Jiang, M. Variation in Vegetation Phenology and Its Response to Climate Change in Marshes of Inner Mongolian. Plants 2023, 12, 2072. [Google Scholar] [CrossRef] [PubMed]
- Zhu, E.; Fang, D.; Chen, L.; Qu, Y.; Liu, T. The Impact of Urbanization on Spatial–Temporal Variation in Vegetation Phenology: A Case Study of the Yangtze River Delta, China. Remote Sens. 2024, 16, 914. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, J.; Wang, Y.; Zhang, H.; Zhang, Z.; Guo, X. Comparison of land surface phenology in the Northern Hemisphere based on AVHRR GIMMS3g and MODIS datasets. ISPRS J. Photogramm. Remote Sens. 2020, 169, 1–16. [Google Scholar] [CrossRef]
- Gao, W.; Zheng, C.; Liu, X.; Lu, Y.; Chen, Y.; Wei, Y.; Ma, Y. NDVI-based vegetation dynamics and their responses to climate change and human activities from 1982 to 2020: A case study in the Mu Us Sandy Land, China. Ecol. Indic. 2022, 137, 108745. [Google Scholar] [CrossRef]
- De Jong, R.; Verbesselt, J.; Zeileis, A.; Schaepman, M.E. Shifts in Global Vegetation Activity Trends. Remote Sens. 2013, 5, 1117–1133. [Google Scholar] [CrossRef]
- Mishra, N.B.; Mainali, K.P. Greening and browning of the Himalaya: Spatial patterns and the role of climatic change and human drivers. Sci. Total Environ. 2017, 587–588, 326–339. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Nath, A.J.; Nath, A.; Sahu, N.; Pandey, R. Landsat-based multi-decadal spatio-temporal assessment of the vegetation greening and browning trend in the Eastern Indian Himalayan Region. Remote Sens. Appl. Soc. Environ. 2022, 25, 100695. [Google Scholar] [CrossRef]
- Piao, S.; Yin, G.; Tan, J.; Cheng, L.; Huang, M.; Li, Y.; Liu, R.; Mao, J.; Myneni, R.B.; Peng, S.; et al. Detection and attribution of vegetation greening trend in China over the last 30 years. Glob. Chang. Biol. 2015, 21, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Calvin, K.; Dasgupta, D.; Krinner, G.; Mukherji, A.; Thorne, P.; Trisos, C.; Romero, J.; Aldunce, P.; Barrett, K. Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2023; pp. 35–115. [Google Scholar] [CrossRef]
- Julien, Y.; Sobrino, J. Global land surface phenology trends from GIMMS database. Int. J. Remote Sens. 2009, 30, 3495–3513. [Google Scholar] [CrossRef]
- Wu, C.; Chen, J.; Gonsamo, A.; Price, D.; Black, T.; Kurz, W. Interannual variability of carbon sequestration is determined by the lag between ends of net uptake and photosynthesis: Evidence from long records of two contrasting forest stands. Agric. For. Meteorol. 2012, 164, 29–38. [Google Scholar] [CrossRef]
- Wu, C.; Hou, X.; Peng, D.; Gonsamo, A.; Xu, S. Land surface phenology of China’s temperate ecosystems over 1999–2013: Spatial–temporal patterns, interaction effects, covariation with climate and implications for productivity. Agric. For. Meteorol. 2016, 216, 177–187. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrêne, E.; Pontailler, J.-Y.; Delpierre, N.; Aubinet, M.; Caquet, B.; de Grandcourt, A.; Burban, B.; Flechard, C.; Granier, A.; et al. Evaluation of the potential of MODIS satellite data to predict vegetation phenology in different biomes: An investigation using ground-based NDVI measurements. Remote Sens. Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- Yuan, Z.; Bao, G.; Dorjsuren, A.; Oyont, A.; Chen, J.; Li, F.; Dong, G.; Guo, E.; Shao, C.; Du, L. Climatic Constraints of Spring Phenology and Its Variability on the Mongolian Plateau From 1982 to 2021. J. Geophys. Res. Biogeosci. 2024, 129, e2023JG007689. [Google Scholar] [CrossRef]
- Yuan, W.; Zheng, Y.; Piao, S.; Ciais, P.; Lombardozzi, D.; Wang, Y.; Ryu, Y.; Chen, G.; Dong, W.; Hu, Z.; et al. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2019, 5, eaax1396. [Google Scholar] [CrossRef] [PubMed]
- Braganza, K.; Karoly, D.J.; Arblaster, J.M. Diurnal temperature range as an index of global climate change during the twentieth century. Geophys. Res. Lett. 2004, 31, L13217. [Google Scholar] [CrossRef]
- Almendra-Martín, L.; Martínez-Fernández, J.; Piles, M.; González-Zamora, Á.; Benito-Verdugo, P.; Gaona, J. Analysis of soil moisture trends in Europe using rank-based and empirical decomposition approaches. Glob. Planet. Chang. 2022, 215, 103868. [Google Scholar] [CrossRef]
- del Río, S.; Cano-Ortiz, A.; Herrero, L.; Penas, A. Recent trends in mean maximum and minimum air temperatures over Spain (1961–2006). Theor. Appl. Climatol. 2012, 109, 605–626. [Google Scholar] [CrossRef]
- Fyfe, J.C.; Gillett, N.P.; Zwiers, F.W. Overestimated global warming over the past 20 years. Nat. Clim. Chang. 2013, 3, 767–769. [Google Scholar] [CrossRef]
- Medhaug, I.; Stolpe, M.B.; Fischer, E.M.; Knutti, R. Reconciling controversies about the Global Warming Hiatus. Nature 2017, 545, 41–47. [Google Scholar] [CrossRef]
- Zahradníček, P.; Brázdil, R.; Štěpánek, P.; Trnka, M. Reflections of global warming in trends of temperature characteristics in the Czech Republic, 1961–2019. Int. J. Climatol. 2021, 41, 1211–1229. [Google Scholar] [CrossRef]
- Huang, X.; Ma, L.; Liu, T.; Sun, B.; Chen, Y.; Qiao, Z.; Liang, L. Response relationship between the abrupt temperature change-climate warming hiatus and changes in influencing factors in China. Int. J. Climatol. 2021, 41, 5178–5200. [Google Scholar] [CrossRef]
- Kosaka, Y.; Xie, S.-P. Recent global-warming hiatus tied to equatorial Pacific surface cooling. Nature 2013, 501, 403–407. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regions | 1st Period (%) | 2nd Period (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| P | N | S | SP | SN | P | N | S | SP | SN | |
| SOS | ||||||||||
| CL | 38 | 62 | 62 | 22 | 40 | 68 | 32 | 64 | 45 | 19 |
| CM | 26 | 73 | 67 | 13 | 54 | 66 | 33 | 65 | 44 | 21 |
| AT | 18 | 81 | 75 | 7 | 68 | 89 | 11 | 77 | 73 | 4 |
| OC | 21 | 79 | 70 | 13 | 57 | 75 | 25 | 81 | 64 | 17 |
| PG | 14 | 86 | 75 | 7 | 68 | 55 | 38 | 60 | 40 | 20 |
| Average | 23 | 76 | 69 | 12 | 57 | 71 | 28 | 69 | 53 | 16 |
| EOS | ||||||||||
| CL | 15 | 85 | 70 | 8 | 62 | 39 | 61 | 59 | 21 | 38 |
| CM | 45 | 54 | 54 | 25 | 30 | 53 | 46 | 62 | 34 | 28 |
| AT | 48 | 51 | 59 | 27 | 32 | 73 | 27 | 67 | 51 | 16 |
| OC | 32 | 68 | 80 | 22 | 58 | 58 | 41 | 63 | 40 | 22 |
| PG | 35 | 65 | 70 | 25 | 45 | 75 | 17 | 72 | 59 | 13 |
| Average | 35 | 65 | 66 | 21 | 45 | 60 | 38 | 65 | 41 | 23 |
| LOS | ||||||||||
| CL | 38 | 61 | 61 | 20 | 41 | 30 | 70 | 68 | 18 | 50 |
| CM | 66 | 34 | 55 | 41 | 14 | 39 | 60 | 62 | 23 | 39 |
| AT | 78 | 22 | 68 | 59 | 10 | 21 | 78 | 66 | 8 | 58 |
| OC | 44 | 55 | 64 | 27 | 37 | 32 | 68 | 62 | 15 | 47 |
| PG | 82 | 18 | 71 | 60 | 11 | 40 | 52 | 56 | 26 | 29 |
| Average | 62 | 38 | 64 | 41 | 23 | 32 | 66 | 63 | 18 | 45 |
| BS | ||||||||||
| CL | 7 | 93 | 80 | 4 | 77 | 67 | 33 | 73 | 51 | 22 |
| CM | 21 | 78 | 65 | 10 | 56 | 38 | 61 | 82 | 29 | 54 |
| AT | 27 | 73 | 63 | 8 | 55 | 84 | 15 | 73 | 67 | 6 |
| OC | 16 | 84 | 74 | 7 | 67 | 49 | 51 | 71 | 34 | 37 |
| PG | 49 | 49 | 56 | 22 | 34 | 57 | 35 | 61 | 39 | 22 |
| Average | 24 | 75 | 68 | 10 | 58 | 59 | 39 | 72 | 44 | 28 |
| BV | ||||||||||
| CL | 70 | 30 | 62 | 49 | 13 | 79 | 21 | 73 | 62 | 11 |
| CM | 58 | 42 | 62 | 41 | 21 | 84 | 16 | 83 | 73 | 11 |
| AT | 58 | 42 | 48 | 28 | 20 | 90 | 10 | 84 | 80 | 4 |
| OC | 98 | 2 | 96 | 94 | 1 | 92 | 8 | 93 | 87 | 6 |
| PG | 68 | 32 | 74 | 50 | 24 | 66 | 27 | 74 | 52 | 22 |
| Average | 70 | 30 | 68 | 52 | 16 | 82 | 17 | 81 | 71 | 11 |
| Periods | Regions | SOS | EOS | LOS | BS | BV | |
|---|---|---|---|---|---|---|---|
| Days/Period | NDVI/Period | Δ% 2P-1P | |||||
| 1 | CL | −3.2 | −6.6 | −3.3 | −13.0 | 0.009 | |
| CM | −6.1 | 0.0 | 5.6 | −6.9 | 0.005 | ||
| AT | −10.6 | −1.1 | 9.5 | −3.7 | 0.004 | ||
| OC | −5.9 | −7.3 | −1.1 | −10.6 | 0.021 | ||
| PG | −11.4 | −0.6 | 10.5 | −1.0 | 0.005 | ||
| Average | −7.5 | −3.1 | 4.3 | −7.0 | 0.009 | ||
| 2 | CL | 6.5 | −2.0 | −8.7 | 4.7 | 0.015 | 40 |
| CM | 4.2 | 1.4 | −3.3 | −5.0 | 0.018 | 69 | |
| AT | 11.8 | 3.3 | −8.7 | 6.6 | 0.020 | 81 | |
| OC | 9.4 | 2.9 | −6.5 | −0.6 | 0.020 | −8 | |
| PG | 5.5 | 2.8 | −2.6 | 2.4 | 0.012 | 54 | |
| Average | 7.5 | 1.7 | −6.0 | 1.6 | 0.017 | 47 | |
| Regions | 1st Period | 2nd Period | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P | N | S | SP | SN | R | P | N | S | SP | SN | R | |
| SOS—EOS | ||||||||||||
| CL | 57 | 41 | 52 | 32 | 20 | 0.09 | 55 | 45 | 50 | 28 | 23 | 0.05 |
| CM | 47 | 52 | 52 | 26 | 27 | −0.02 | 55 | 44 | 56 | 33 | 23 | 0.08 |
| AT | 41 | 57 | 52 | 20 | 32 | −0.09 | 68 | 32 | 53 | 43 | 10 | 0.24 |
| OC | 62 | 37 | 61 | 41 | 21 | 0.16 | 57 | 43 | 56 | 34 | 23 | 0.09 |
| PG | 46 | 52 | 52 | 24 | 28 | −0.01 | 51 | 49 | 59 | 33 | 25 | 0.02 |
| Average | 51 | 48 | 54 | 28 | 26 | 0.03 | 57 | 43 | 55 | 34 | 21 | 0.09 |
| SOS—LOS | ||||||||||||
| CL | 5 | 95 | 81 | 0 | 81 | −0.67 | 2 | 98 | 92 | 0 | 91 | −0.82 |
| CM | 7 | 92 | 82 | 3 | 79 | −0.65 | 8 | 92 | 86 | 4 | 82 | −0.70 |
| AT | 0 | 100 | 95 | 0 | 95 | −0.87 | 2 | 98 | 96 | 0 | 95 | −0.84 |
| OC | 22 | 77 | 65 | 7 | 57 | −0.41 | 6 | 93 | 84 | 1 | 83 | −0.70 |
| PG | 0 | 100 | 100 | 0 | 100 | −0.96 | 1 | 99 | 99 | 0 | 99 | −0.94 |
| Average | 7 | 93 | 85 | 2 | 83 | −0.71 | 4 | 96 | 91 | 1 | 90 | −0.80 |
| EOS—LOS | ||||||||||||
| CL | 88 | 12 | 74 | 72 | 2 | 0.56 | 79 | 21 | 60 | 54 | 6 | 0.40 |
| CM | 90 | 9 | 78 | 77 | 1 | 0.64 | 83 | 17 | 67 | 62 | 5 | 0.49 |
| AT | 83 | 17 | 68 | 62 | 6 | 0.46 | 68 | 32 | 54 | 38 | 16 | 0.19 |
| OC | 93 | 7 | 86 | 84 | 2 | 0.71 | 83 | 17 | 67 | 62 | 5 | 0.50 |
| PG | 66 | 34 | 61 | 42 | 19 | 0.22 | 61 | 39 | 62 | 45 | 17 | 0.20 |
| Average | 84 | 16 | 73 | 67 | 6 | 0.52 | 75 | 25 | 62 | 52 | 10 | 0.36 |
| Variables | SOS | EOS | ||||||
|---|---|---|---|---|---|---|---|---|
| Autumn | Last Summer | Summer | Spring | |||||
| 1P | 2P | 1P | 2P | 1P | 2P | 1P | 2P | |
| P | −0.10 | −0.17 | 0.04 | 0.11 | −0.16 | 0.08 | 0.09 | 0.06 |
| Tmax | 0.27 | 0.53 * | −0.52 * | 0.30 | −0.37 | 0.45 * | −0.71 * | 0.26 |
| Tmin | 0.08 | 0.03 | −0.71 * | −0.14 | −0.61 * | 0.04 | −0.58 * | −0.04 |
| VPD | 0.36 | 0.40 | −0.45 * | 0.28 | −0.26 | 0.41 | −0.54 * | 0.19 |
| SM | −0.17 | −0.36 | 0.40 | 0.07 | 0.29 | −0.04 | 0.50 * | −0.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benito-Verdugo, P.; González-Zamora, Á.; Martínez-Fernández, J. Recent Cereal Phenological Variations under Mediterranean Conditions. Remote Sens. 2024, 16, 1879. https://doi.org/10.3390/rs16111879
Benito-Verdugo P, González-Zamora Á, Martínez-Fernández J. Recent Cereal Phenological Variations under Mediterranean Conditions. Remote Sensing. 2024; 16(11):1879. https://doi.org/10.3390/rs16111879
Chicago/Turabian StyleBenito-Verdugo, Pilar, Ángel González-Zamora, and José Martínez-Fernández. 2024. "Recent Cereal Phenological Variations under Mediterranean Conditions" Remote Sensing 16, no. 11: 1879. https://doi.org/10.3390/rs16111879
APA StyleBenito-Verdugo, P., González-Zamora, Á., & Martínez-Fernández, J. (2024). Recent Cereal Phenological Variations under Mediterranean Conditions. Remote Sensing, 16(11), 1879. https://doi.org/10.3390/rs16111879