


Early Prediction of Regional Red Needle Cast Outbreaks Using Climatic Data Trends and Satellite-Derived Observations

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
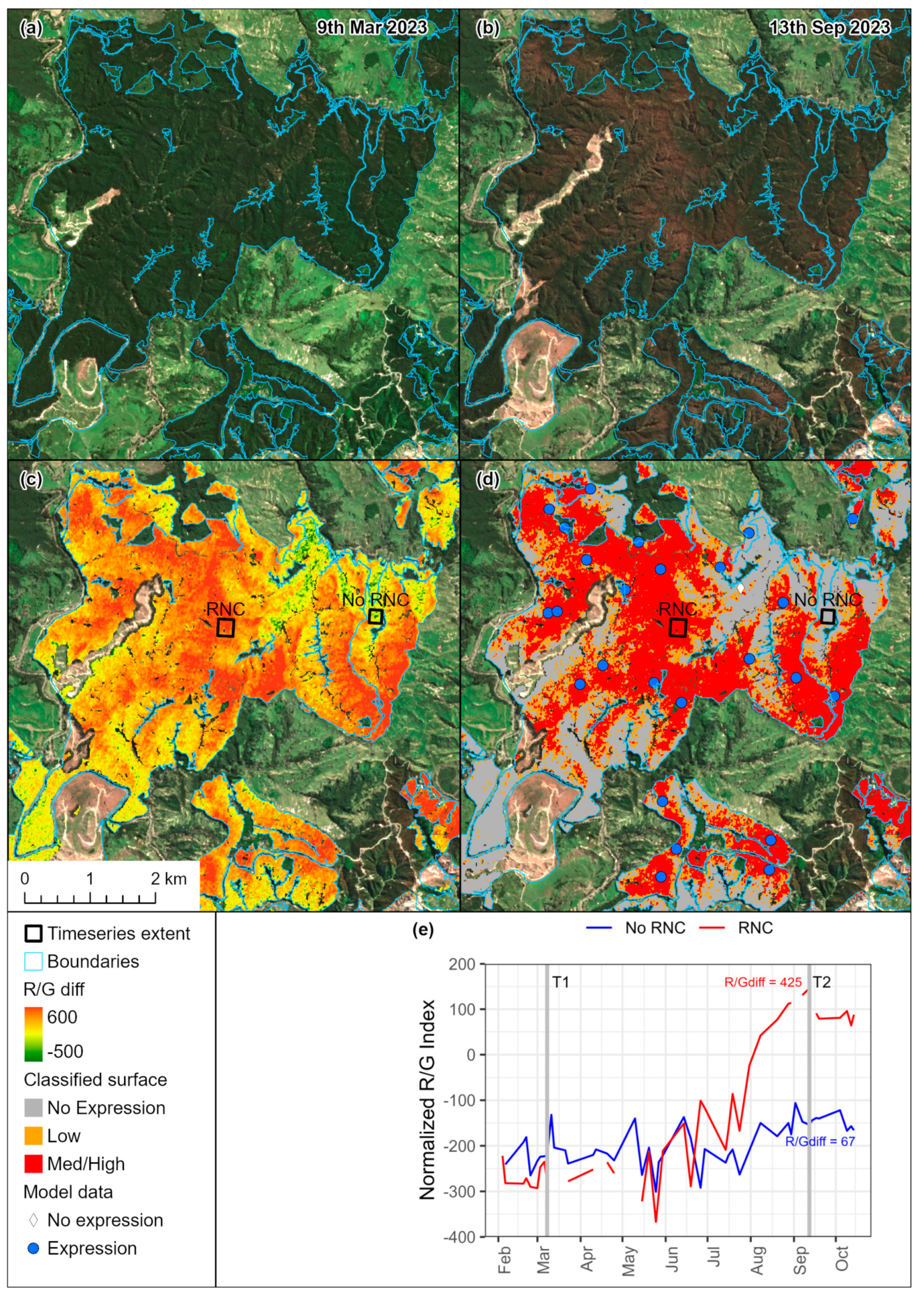
2.1. Spatial Predictions of RNC from Sentinel-2
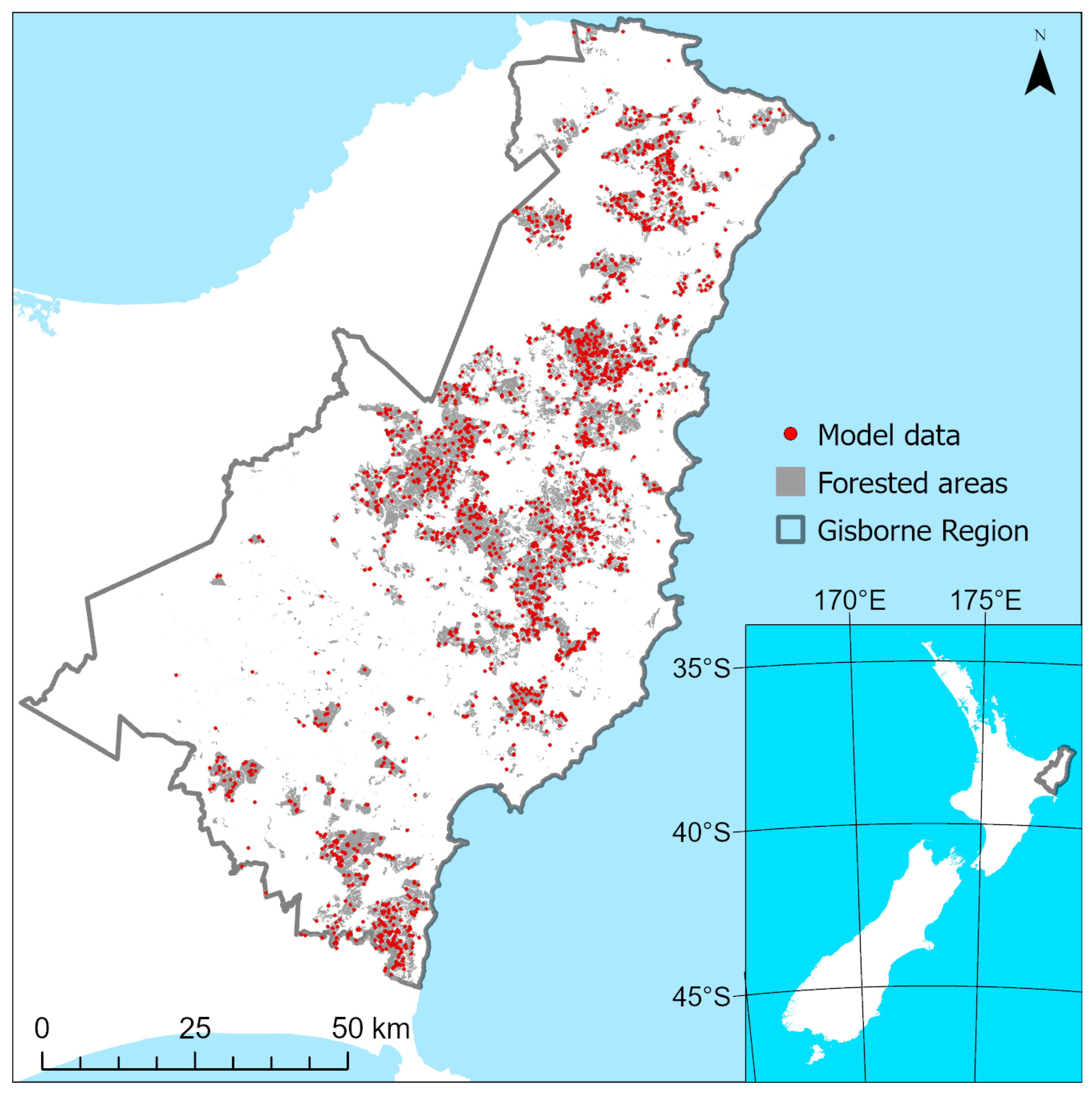
2.1.1. Study Area and Plantation Delineation
2.1.2. Data Collection
2.1.3. Satellite Imagery Processing
2.1.4. Sampling
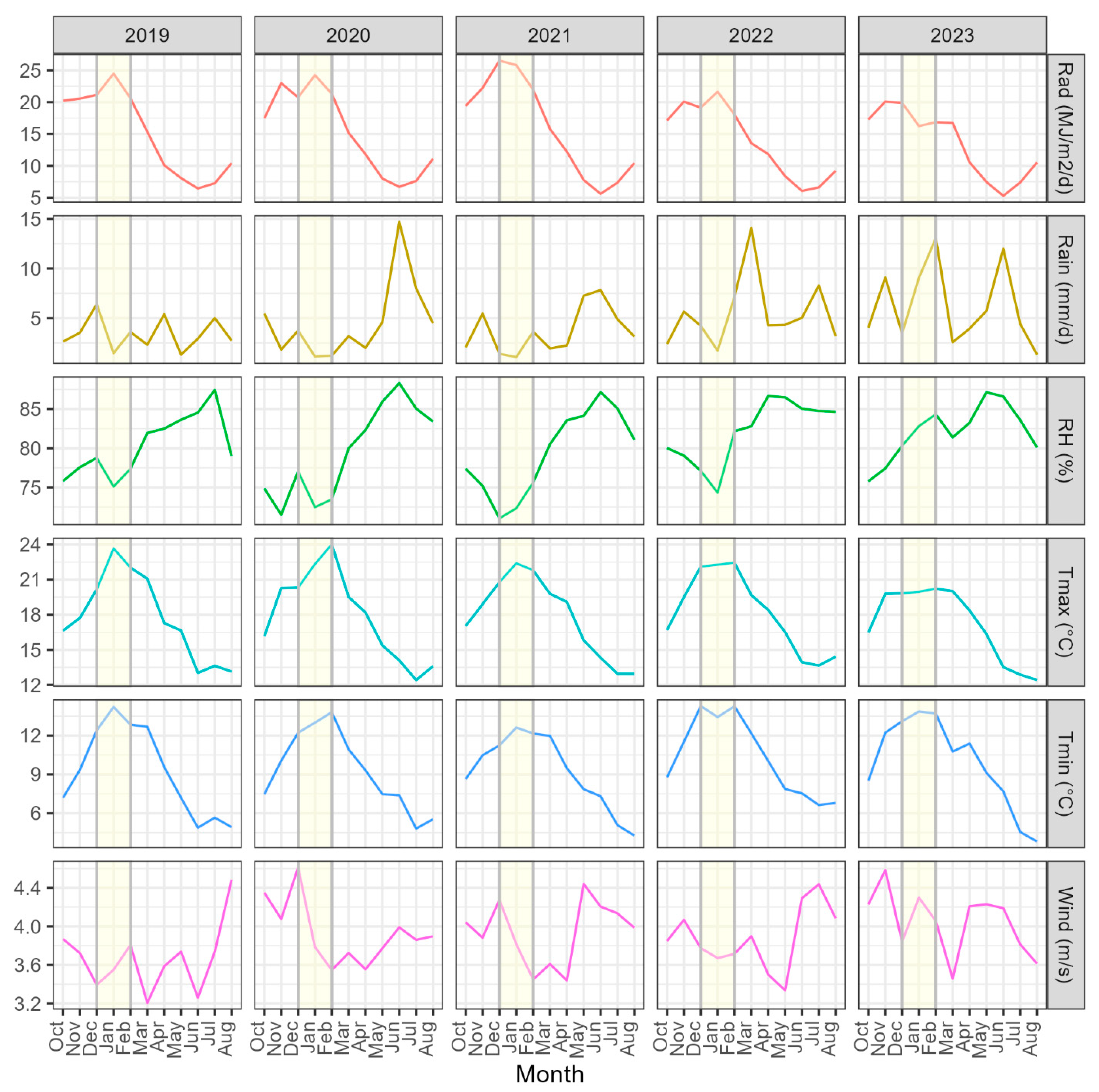
2.2. Weather Data
2.3. Data Analysis
2.3.1. Relationships between R/Gdiff and Weather Variables
2.3.2. Classification Model
3. Results
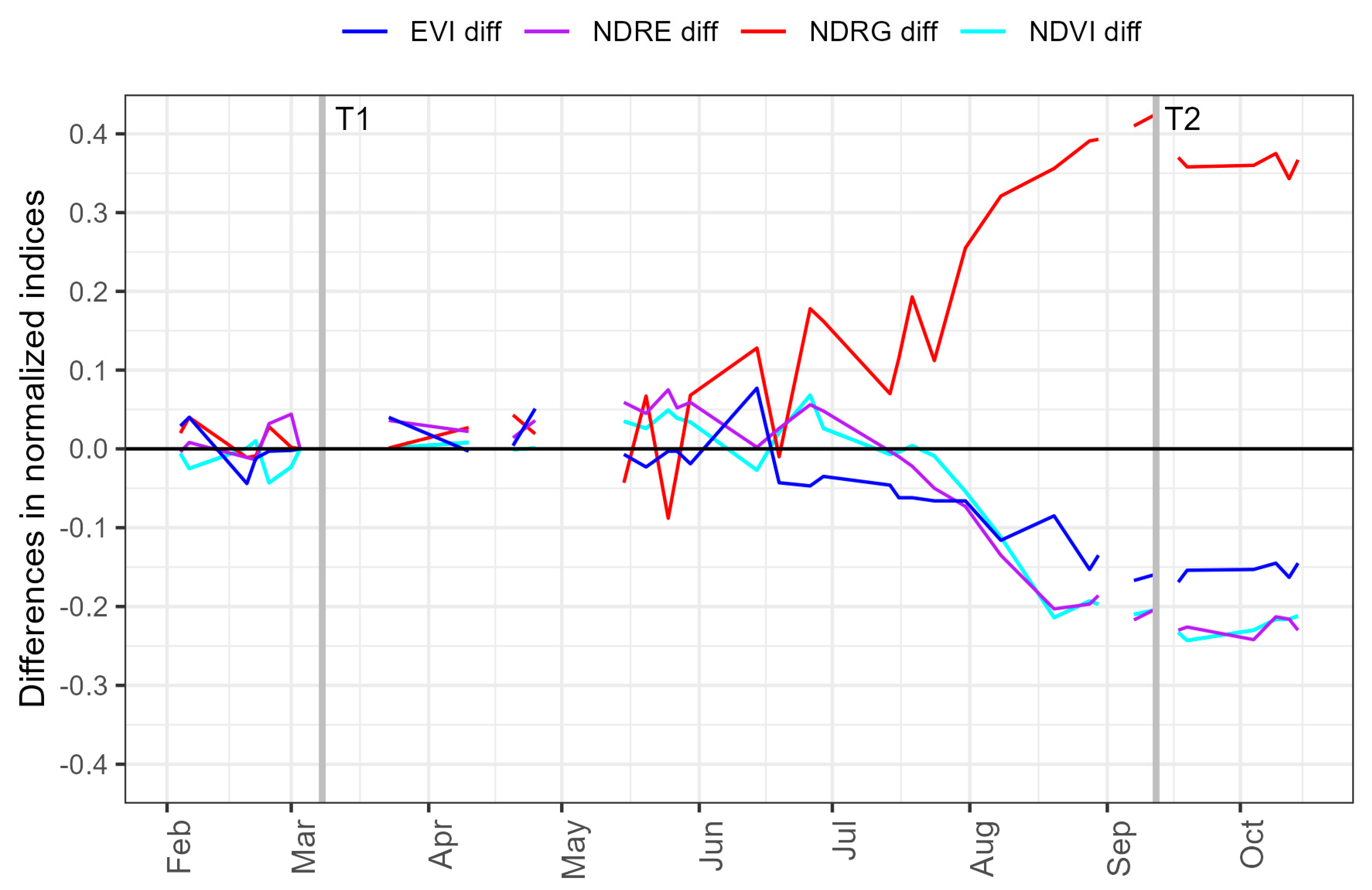
3.1. Variation in RNC Severity
3.2. Variation in Environmental Conditions
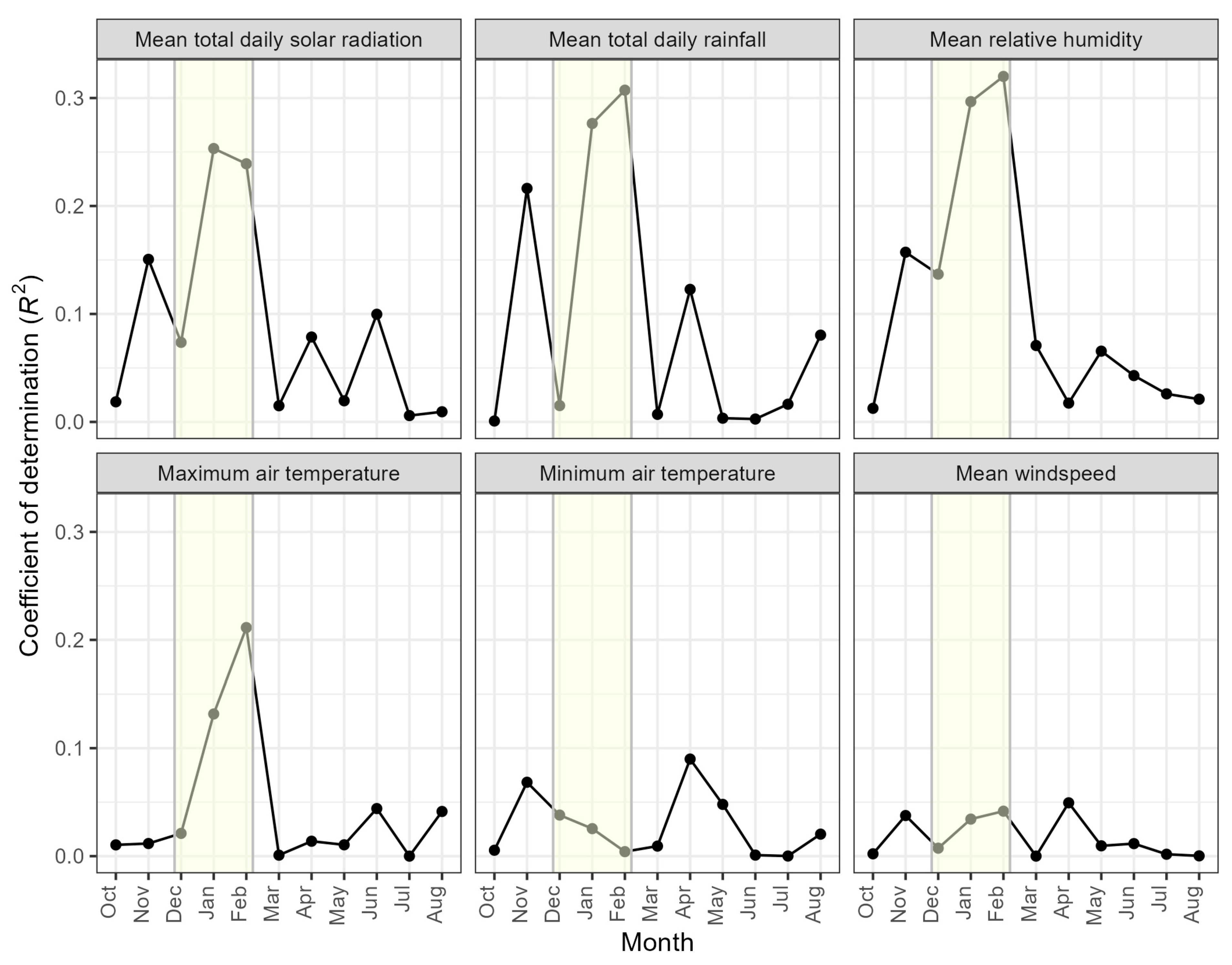
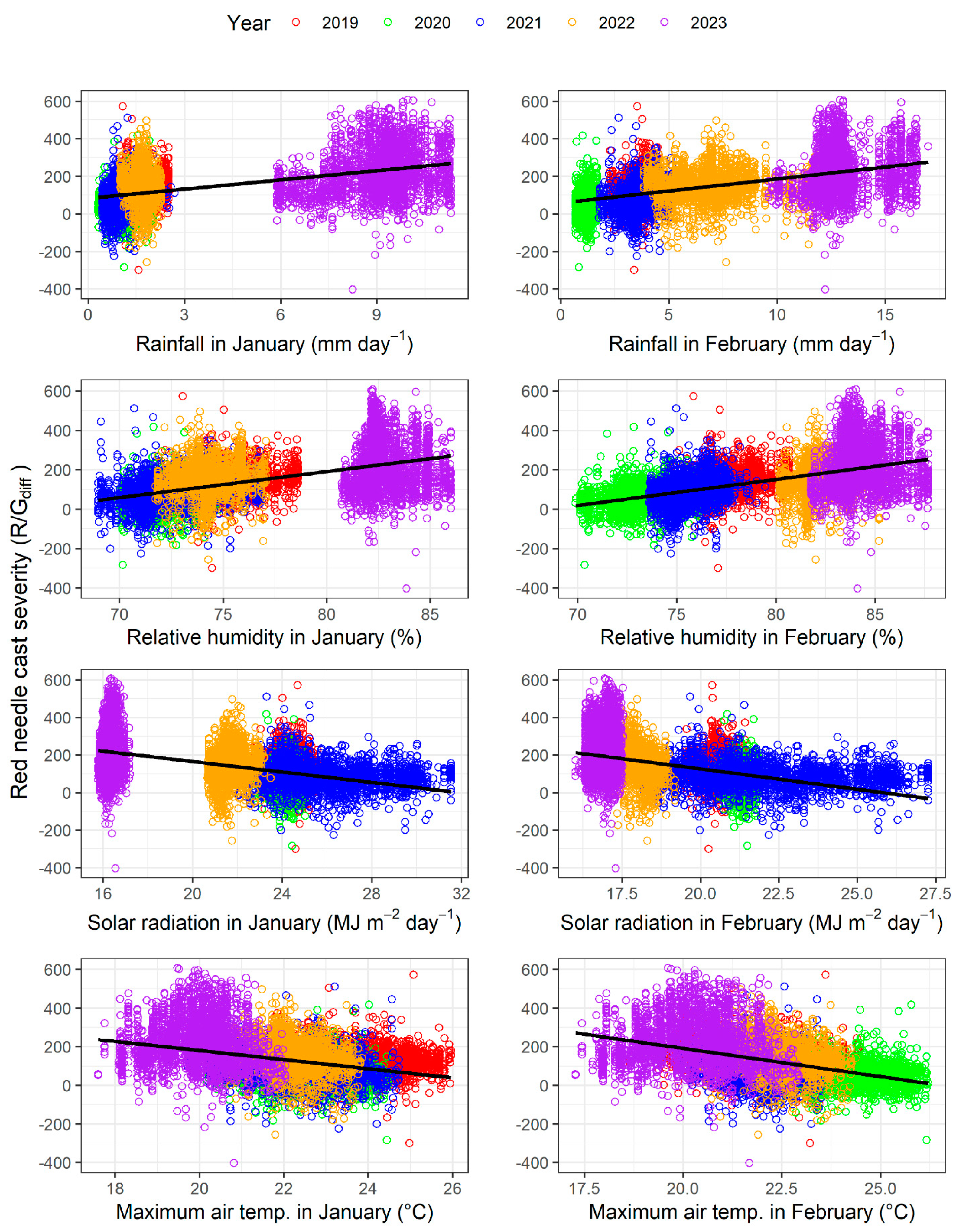
3.3. Relationships with Climatic Variables and Seasonal Patterns
3.4. Classification Model
3.4.1. Variable Selection and Model Performance
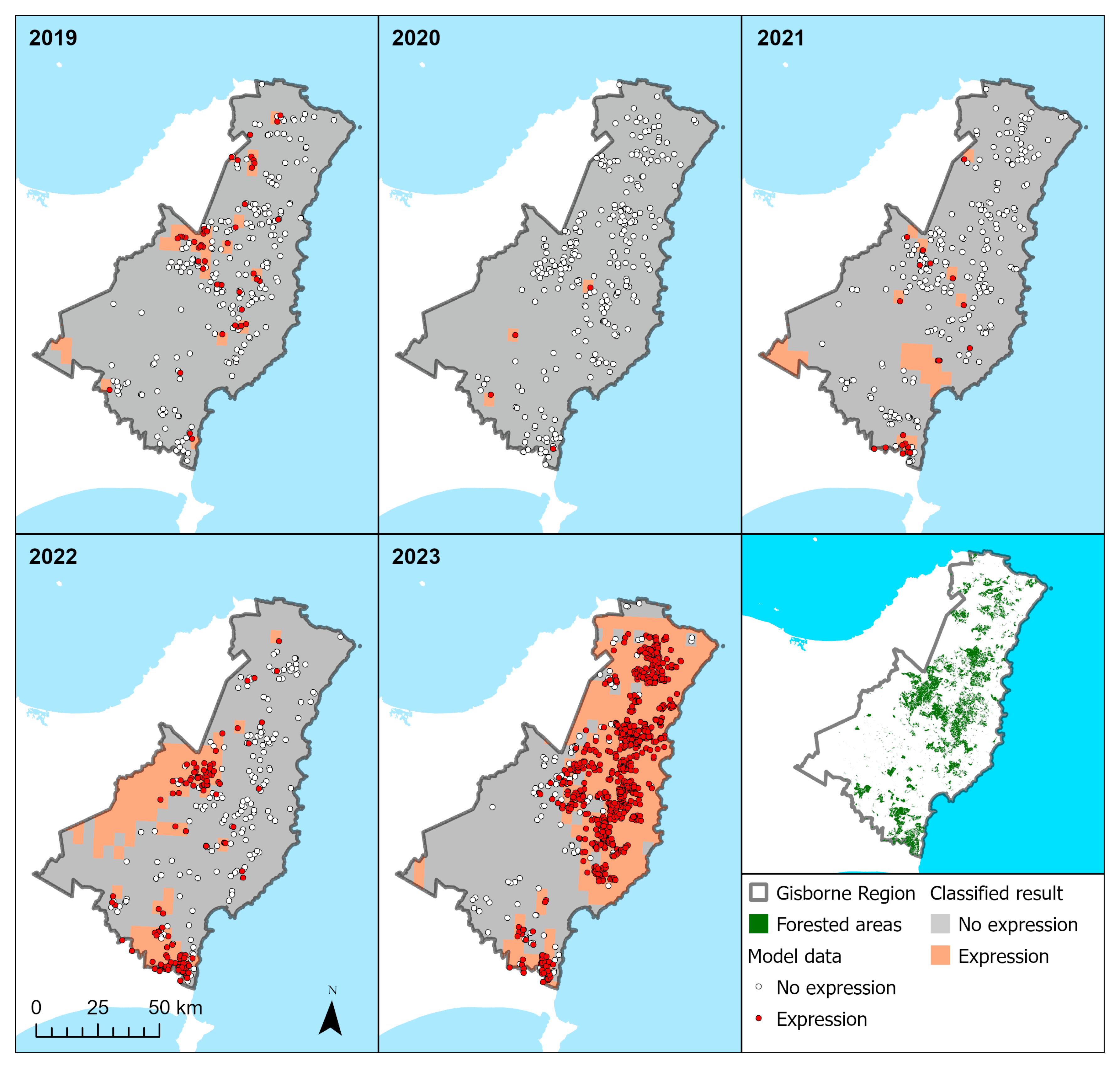
3.4.2. Model Predictions
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Month | Intercept | Slope | R2 |
|---|---|---|---|---|
| Rainfall (mm day−1) | January | 82.6 | 16.4 | 0.28 |
| Relative humidity (%) | January | −866 | 13.2 | 0.30 |
| Solar radiation (MJ m−2 day−1) | January | 443 | −13.9 | 0.25 |
| Maximum air temperature (°C) | January | 653 | −23.6 | 0.13 |
| Rainfall (mm day−1) | February | 58.1 | 12.8 | 0.31 |
| Relative humidity (%) | February | −910 | 13.3 | 0.32 |
| Solar radiation (MJ m−2 day−1) | February | 561 | −21.7 | 0.24 |
| Maximum air temperature (°C) | February | 777 | −29.3 | 0.21 |
References
- NZFOA. New Zealand Forestry Industry, Facts and Figures 2022/2023. New Zealand Plantation Forest Industry. New Zealand Forest Owners Association, Wellington, 2023. Available online: https://www.nzfoa.org.nz/images/Facts_and_Figures_2022-2023_-_WEB.pdf (accessed on 11 March 2024).
- Dick, M.A.; Williams, N.M.; Bader, M.K.-F.; Gardner, J.F.; Bulman, L.S. Pathogenicity of Phytophthora pluvialis to Pinus radiata and its relation with red needle cast disease in New Zealand. New Zealand J. For. Sci. 2014, 44, 6. [Google Scholar] [CrossRef]
- Graham, N.J.; Suontama, M.; Pleasants, T.; Li, Y.; Bader, M.K.F.; Klápště, J.; Dungey, H.S.; Williams, N.M. Assessing the genetic variation of tolerance to red needle cast in a Pinus radiata breeding population. Tree Genet. Genom. 2018, 14, 55. [Google Scholar] [CrossRef]
- Gomez-Gallego, M.; Gommers, R.; Bader, M.K.-F.; Williams, N.M. Modelling the key drivers of an aerial Phytophthora foliar disease epidemic, from the needles to the whole plant. PLoS ONE 2019, 14, e0216161. [Google Scholar] [CrossRef] [PubMed]
- Ganley, R.J.; Williams, N.M.; Rolando, C.A.; Hood, I.A.; Dungey, H.S.; Beets, P.N.; Bulman, L.S. Management of red needle cast caused by Phytophthora pluvialis a new disease of radiata pine in New Zealand. New Zealand Plant Prot. 2014, 67, 48–53. [Google Scholar] [CrossRef]
- Reeser, P.; Sutton, W.; Hansen, E.M. Phytophthora pluvialis, a new species from mixed tanoak-Douglas-fir forests of western Oregon, USA. North Am. Fungi 2013, 8, 1–8. [Google Scholar] [CrossRef]
- Shelley, B.A.; Luster, D.G.; Garrett, W.M.; McMahon, M.B.; Widmer, T.L. Effects of temperature on germination of sporangia, infection and protein secretion by Phytophthora kernoviae. Plant Pathol. 2018, 67, 719–728. [Google Scholar] [CrossRef]
- Fraser, S.; Gomez-Gallego, M.; Gardner, J.; Bulman, L.S.; Denman, S.; Williams, N.M. Impact of weather variables and season on sporulation of Phytophthora pluvialis and Phytophthora kernoviae. For. Pathol. 2020, 50, e12588. [Google Scholar] [CrossRef]
- Hood, I.A.; Husheer, S.; Gardner, J.F.; Evanson, T.W.; Tieman, G.; Banham, C.; Wright, L.A.H.; Fraser, S. Infection periods of Phytophthora pluvialis and Phytophthora kernoviae in relation to weather variables and season in Pinus radiata forests in New Zealand. New Zealand J. For. Sci. 2022, 52, 17. [Google Scholar] [CrossRef]
- Bulman, L.S.; Bradshaw, R.E.; Fraser, S.; Martín-García, J.; Barnes, I.; Musolin, D.L.; La Porta, N.; Woods, A.J.; Diez, J.J.; Koltay, A.; et al. A worldwide perspective on the management and control of Dothistroma needle blight. For. Pathol. 2016, 46, 472–488. [Google Scholar] [CrossRef]
- Fraser, S.; Baker, M.; Pearse, G.; Todoroki, C.L.; Estarija, H.J.; Hood, I.A.; Bulman, L.S.; Somchit, C.; Rolando, C.A. Efficacy and optimal timing of low-volume aerial applications of copper fungicides for the control of red needle cast of pine. New Zealand J. For. Sci. 2022, 52, 18. [Google Scholar] [CrossRef]
- Bárta, V.; Lukeš, P.; Homolová, L. Early detection of bark beetle infestation in Norway spruce forests of Central Europe using Sentinel-2. Int. J. Appl. Earth Obs. Geoinf. 2021, 100, 102335. [Google Scholar] [CrossRef]
- Bozzini, A.; Francini, S.; Chirici, G.; Battisti, A.; Faccoli, M. Spruce bark beetle outbreak prediction through automatic classification of Sentinel-2 imagery. Forests 2023, 14, 1116. [Google Scholar] [CrossRef]
- Dalponte, M.; Cetto, R.; Marinelli, D.; Andreatta, D.; Salvadori, C.; Pirotti, F.; Frizzera, L.; Gianelle, D. Spectral separability of bark beetle infestation stages: A single-tree time-series analysis using Planet imagery. Ecol. Indic. 2023, 153, 110349. [Google Scholar] [CrossRef]
- Senf, C.; Pflugmacher, D.; Wulder, M.A.; Hostert, P. Characterizing spectral–temporal patterns of defoliator and bark beetle disturbances using Landsat time series. Remote Sens. Environ. 2015, 170, 166–177. [Google Scholar] [CrossRef]
- Panzavolta, T.; Bracalini, M.; Benigno, A.; Moricca, S. Alien invasive pathogens and pests harming trees, forests, and plantations: Pathways, global consequences and management. Forests 2021, 12, 1364. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Slippers, B.; Wingfield, B.D. Novel Associations between Pathogens, Insects and Tree Species Threaten World Forests; University of Pretoria: Pretoria, South Africa, 2010. [Google Scholar]
- Poona, N.K.; Ismail, R. Discriminating the occurrence of pitch canker fungus in Pinus radiata trees using QuickBird imagery and artificial neural networks. South. For. J. For. Sci. 2013, 75, 29–40. [Google Scholar] [CrossRef]
- Camarretta, N.; Pearse, G.D.; Steer, B.S.C.; McLay, E.; Fraser, S.; Watt, M.S. Automatic Detection of Phytophthora pluvialis Outbreaks in Radiata Pine Plantations Using Multi-Scene, Multi-Temporal Satellite Imagery. Remote Sens. 2024, 16, 338. [Google Scholar] [CrossRef]
- Mantas, V.; Fonseca, L.; Baltazar, E.; Canhoto, J.; Abrantes, I. Detection of tree decline (Pinus pinaster Aiton) in European forests using Sentinel-2 data. Remote Sens. 2022, 14, 2028. [Google Scholar] [CrossRef]
- Migas-Mazur, R.; Kycko, M.; Zwijacz-Kozica, T.; Zagajewski, B. Assessment of Sentinel-2 images, support vector machines and change detection algorithms for bark beetle outbreaks mapping in the Tatra mountains. Remote Sens. 2021, 13, 3314. [Google Scholar] [CrossRef]
- Venter, Z.S.; Barton, D.N.; Chakraborty, T.; Simensen, T.; Singh, G. Global 10 m land use land cover datasets: A comparison of dynamic world, world cover and esri land cover. Remote Sens. 2022, 14, 4101. [Google Scholar] [CrossRef]
- Hernández-Clemente, R.; Hornero, A.; Mottus, M.; Penuelas, J.; González-Dugo, V.; Jiménez, J.C.; Suárez, L.; Alonso, L.; Zarco-Tejada, P.J. Early Diagnosis of Vegetation Health From High-Resolution Hyperspectral and Thermal Imagery: Lessons Learned from Empirical Relationships and Radiative Transfer Modelling. Curr. For. Rep. 2019, 5, 169–183. [Google Scholar] [CrossRef]
- Hornero, A.; Zarco-Tejada, P.J.; Quero, J.L.; North, P.R.J.; Ruiz-Gómez, F.J.; Sánchez-Cuesta, R.; Hernandez-Clemente, R. Modelling hyperspectral-and thermal-based plant traits for the early detection of Phytophthora-induced symptoms in oak decline. Remote Sens. Environ. 2021, 263, 112570. [Google Scholar] [CrossRef]
- Mahlein, A.K.; Rumpf, T.; Welke, P.; Dehne, H.W.; Plümer, L.; Steiner, U.; Oerke, E.C. Development of spectral indices for detecting and identifying plant diseases. Remote Sens. Environ. 2013, 128, 21–30. [Google Scholar] [CrossRef]
- Poblete, T.; Camino, C.; Beck, P.S.A.; Hornero, A.; Kattenborn, T.; Saponari, M.; Boscia, D.; Navas-Cortes, J.A.; Zarco-Tejada, P.J. Detection of Xylella fastidiosa infection symptoms with airborne multispectral and thermal imagery: Assessing bandset reduction performance from hyperspectral analysis. ISPRS J. Photogramm. Remote Sens. 2020, 162, 27–40. [Google Scholar] [CrossRef]
- Tian, L.; Xue, B.; Wang, Z.; Li, D.; Yao, X.; Cao, Q.; Zhu, Y.; Cao, W.; Cheng, T. Spectroscopic detection of rice leaf blast infection from asymptomatic to mild stages with integrated machine learning and feature selection. Remote Sens. Environ. 2021, 257, 112350. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Camino, C.; Beck, P.S.A.; Calderon, R.; Hornero, A.; Hernández-Clemente, R.; Kattenborn, T.; Montes-Borrego, M.; Susca, L.; Morelli, M.; et al. Previsual symptoms of Xylella fastidiosa infection revealed in spectral plant-trait alterations. Nat. Plants 2018, 4, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Calderón, R.; Navas-Cortés, J.A.; Zarco-Tejada, P.J. Early detection and quantification of Verticillium wilt in olive using hyperspectral and thermal imagery over large areas. Remote Sens. 2015, 7, 5584–5610. [Google Scholar] [CrossRef]
- Lee, E.H.; Beedlow, P.A.; Waschmann, R.S.; Burdick, C.A.; Shaw, D.C. Tree-ring analysis of the fungal disease Swiss needle cast in western Oregon coastal forests. Can. J. For. Res. 2013, 43, 677–690. [Google Scholar] [CrossRef]
- Lech, P.; Mychayliv, O.; Hildebrand, R.; Orman, O. Weather Conditions Drive the Damage Area Caused by Armillaria Root Disease in Coniferous Forests across Poland. Plant Pathol. J. 2023, 39, 548–565. [Google Scholar] [CrossRef]
- Ades, P.K.; Simpson, J.A.; Eldridge, K.G.; Eldridge, R.H. Genetic variation in susceptibility to Dothistroma needle blight among provenances and families of Pinus muricata. Can. J. For. Res. 1992, 22, 1111–1117. [Google Scholar] [CrossRef]
- Woods, A.; Coates, K.; Hamann, A. Is an unprecedented Dothistroma needle blight epidemic related to climate change? BioScience 2005, 55, 761–769. [Google Scholar] [CrossRef]
- Bulman, L.S.; Dick, M.A.; Ganley, R.J.; McDougal, R.L.; Schwelm, A.; Bradshaw, R.E.; Gonthier, P.; Nicolotti, G. Dothistroma needle blight. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CPI Group (UK) Ltd.: Croydon, UK, 2013; pp. 436–457. [Google Scholar]
- McDougal, R.L.; Cunningham, L.; Hunter, S.; Caird, A.; Flint, H.; Lewis, A.; Ganley, R.J. Molecular detection of Phytophthora pluvialis, the causal agent of red needle cast in Pinus radiata. J. Microbiol. Methods 2021, 189, 106299. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.S.; Tan, A.Y.S.; Fraser, S.; Bulman, L.S. Use of advanced modelling methods to predict dothistroma needle blight on Pinus radiata at a fine resolution within New Zealand. For. Ecol. Manage. 2021, 492, 119226. [Google Scholar] [CrossRef]
- Chen, L.-C.; Zhu, Y.; Papandreou, G.; Schroff, F.; Adam, H. Encoder-decoder with atrous separable convolution for semantic image segmentation. In Proceedings of the European Conference on Computer Vision (ECCV), Online, 6 October 2018; pp. 801–818. [Google Scholar]
- He, K.; Zhang, X.; Ren, S.; Sun, J. Deep residual learning for image recognition. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Vegas, NV, USA, 27–30 June 2016; pp. 770–778. [Google Scholar]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Drusch, M.; Del Bello, U.; Carlier, S.; Colin, O.; Fernandez, V.; Gascon, F.; Hoersch, B.; Isola, C.; Laberinti, P.; Martimort, P. Sentinel-2: ESA’s optical high-resolution mission for GMES operational services. Remote Sens. Environ. 2012, 120, 25–36. [Google Scholar] [CrossRef]
- Main-Knorn, M.; Pflug, B.; Louis, J.; Debaecker, V.; Müller-Wilm, U.; Gascon, F. Sen2Cor for sentinel-2. In Proceedings of the Image and Signal Processing for Remote Sensing XXIII, SPIE, Warsaw, Poland, 4 October 2017; Volume 10427, pp. 37–48. [Google Scholar]
- Pasquarella, V.J.; Brown, C.F.; Czerwinski, W.; Rucklidge, W.J. Comprehensive Quality Assessment of Optical Satellite Imagery Using Weakly Supervised Video Learning. In Proceedings of the IEEE/CVF Conference on Computer Vision and Pattern Recognition, Vancouver, BC, Canada, 17–24 June 2023; pp. 2124–2134. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 15 January 2024).
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V. Scikit-learn: Machine Learning in Python. J. Mach. Learn.Res. 2011, 12, 2825–2830. [Google Scholar]
- Yuan, X.; Chen, S.; Sun, C.; Yuwen, L. A novel early diagnostic framework for chronic diseases with class imbalance. Sci. Rep. 2022, 12, 8614. [Google Scholar] [CrossRef] [PubMed]
- Coops, N.; Stanford, M.; Old, K.; Dudzinski, M.; Culvenor, D.; Stone, C. Assessment of dothistroma needle blight of Pinus radiata using airborne hyperspectral imagery. Phytopathology 2003, 93, 1524–1532. [Google Scholar] [CrossRef] [PubMed]
- Leckie, D.; Jay, C.; Gougeon, F.; Sturrock, R.; Paradine, D. Detection and assessment of trees with Phellinus weirii (laminated root rot) using high resolution multi-spectral imagery. Int. J. Remote Sens. 2004, 25, 793–818. [Google Scholar] [CrossRef]
- Rolando, C.A.; Dick, M.A.; Gardner, J.; Bader, M.K.F.; Williams, N.M. Chemical control of two Phytophthora species infecting the canopy of Monterey pine (Pinus radiata). For. Pathol. 2017, 47, e12327. [Google Scholar] [CrossRef]
- Rolando, C.; Somchit, C.; Bader, M.K.F.; Fraser, S.; Williams, N. Can copper be used to treat foliar Phytophthora infections in Pinus radiata? Plant Dis. 2019, 103, 1828–1834. [Google Scholar] [CrossRef] [PubMed]
- Van der Plank, J.E. Plant Diseases: Epidemics and Control; Academic Press: New York, NY, USA, 1963; p. 349. [Google Scholar]
- Sabatia, C.O.; Burkhart, H.E. Predicting site index of plantation loblolly pine from biophysical variables. For. Ecol. Manag. 2014, 326, 142–156. [Google Scholar] [CrossRef]
- González-Rodríguez, M.Á.; Diéguez-Aranda, U. Rule-based vs parametric approaches for developing climate-sensitive site index models: A case study for Scots pine stands in northwestern Spain. Ann. For. Sci. 2021, 78, 23. [Google Scholar] [CrossRef]
- Watt, M.; Moore, J.R. Modelling spatial variation in radiata pine slenderness (height/diameter ratio) and vulnerability to wind damage under current and future climate in New Zealand. Front. For. Glob. Chang. 2023, 6, 1188094. [Google Scholar] [CrossRef]
- Spoto, F.; Sy, O.; Laberinti, P.; Martimort, P.; Fernandez, V.; Colin, O.; Hoersch, B.; Meygret, A. Overview of Sentinel-2. In Proceedings of the 2012 IEEE International Geoscience and Remote Sensing Symposium, Munich, Germany, 22–27 July 2012; pp. 1707–1710. [Google Scholar]
- Schepaschenko, D.; See, L.; Lesiv, M.; Bastin, J.-F.; Mollicone, D.; Tsendbazar, N.-E.; Bastin, L.; McCallum, I.; Laso Bayas, J.C.; Baklanov, A. Recent advances in forest observation with visual interpretation of very high-resolution imagery. Surv. Geophys. 2019, 40, 839–862. [Google Scholar] [CrossRef]
- Gavilán-Acuña, G.; Olmedo, G.F.; Mena-Quijada, P.; Guevara, M.; Barría-Knopf, B.; Watt, M.S. Reducing the Uncertainty of Radiata Pine Site Index Maps Using an Spatial Ensemble of Machine Learning Models. Forests 2021, 12, 77. [Google Scholar] [CrossRef]









| 2019 | 2020 | 2021 | 2022 | 2023 | |
|---|---|---|---|---|---|
| R/Gdiff | 132 (1.1) | 60 (1.0) | 88 (1.3) | 150 (1.7) | 225 (2.5) |
| RNC severity by class (%) | |||||
| None | 89.9 | 99.0 | 94.8 | 77.1 | 47.3 |
| Low | 8.4 | 0.9 | 4.3 | 17.4 | 21.8 |
| Med | 1.6 | 0.0 | 0.7 | 5.0 | 18.0 |
| High | 0.1 | 0.1 | 0.2 | 0.5 | 12.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watt, M.S.; Holdaway, A.; Watt, P.; Pearse, G.D.; Palmer, M.E.; Steer, B.S.C.; Camarretta, N.; McLay, E.; Fraser, S. Early Prediction of Regional Red Needle Cast Outbreaks Using Climatic Data Trends and Satellite-Derived Observations. Remote Sens. 2024, 16, 1401. https://doi.org/10.3390/rs16081401
Watt MS, Holdaway A, Watt P, Pearse GD, Palmer ME, Steer BSC, Camarretta N, McLay E, Fraser S. Early Prediction of Regional Red Needle Cast Outbreaks Using Climatic Data Trends and Satellite-Derived Observations. Remote Sensing. 2024; 16(8):1401. https://doi.org/10.3390/rs16081401
Chicago/Turabian StyleWatt, Michael S., Andrew Holdaway, Pete Watt, Grant D. Pearse, Melanie E. Palmer, Benjamin S. C. Steer, Nicolò Camarretta, Emily McLay, and Stuart Fraser. 2024. "Early Prediction of Regional Red Needle Cast Outbreaks Using Climatic Data Trends and Satellite-Derived Observations" Remote Sensing 16, no. 8: 1401. https://doi.org/10.3390/rs16081401
APA StyleWatt, M. S., Holdaway, A., Watt, P., Pearse, G. D., Palmer, M. E., Steer, B. S. C., Camarretta, N., McLay, E., & Fraser, S. (2024). Early Prediction of Regional Red Needle Cast Outbreaks Using Climatic Data Trends and Satellite-Derived Observations. Remote Sensing, 16(8), 1401. https://doi.org/10.3390/rs16081401