
Spatial Assessment of the Bioclimatic and Environmental Factors Driving Mangrove Tree Species’ Distribution along the Brazilian Coastline

 , ,
, ,
Abstract
:
1. Introduction
2. Material and Methods
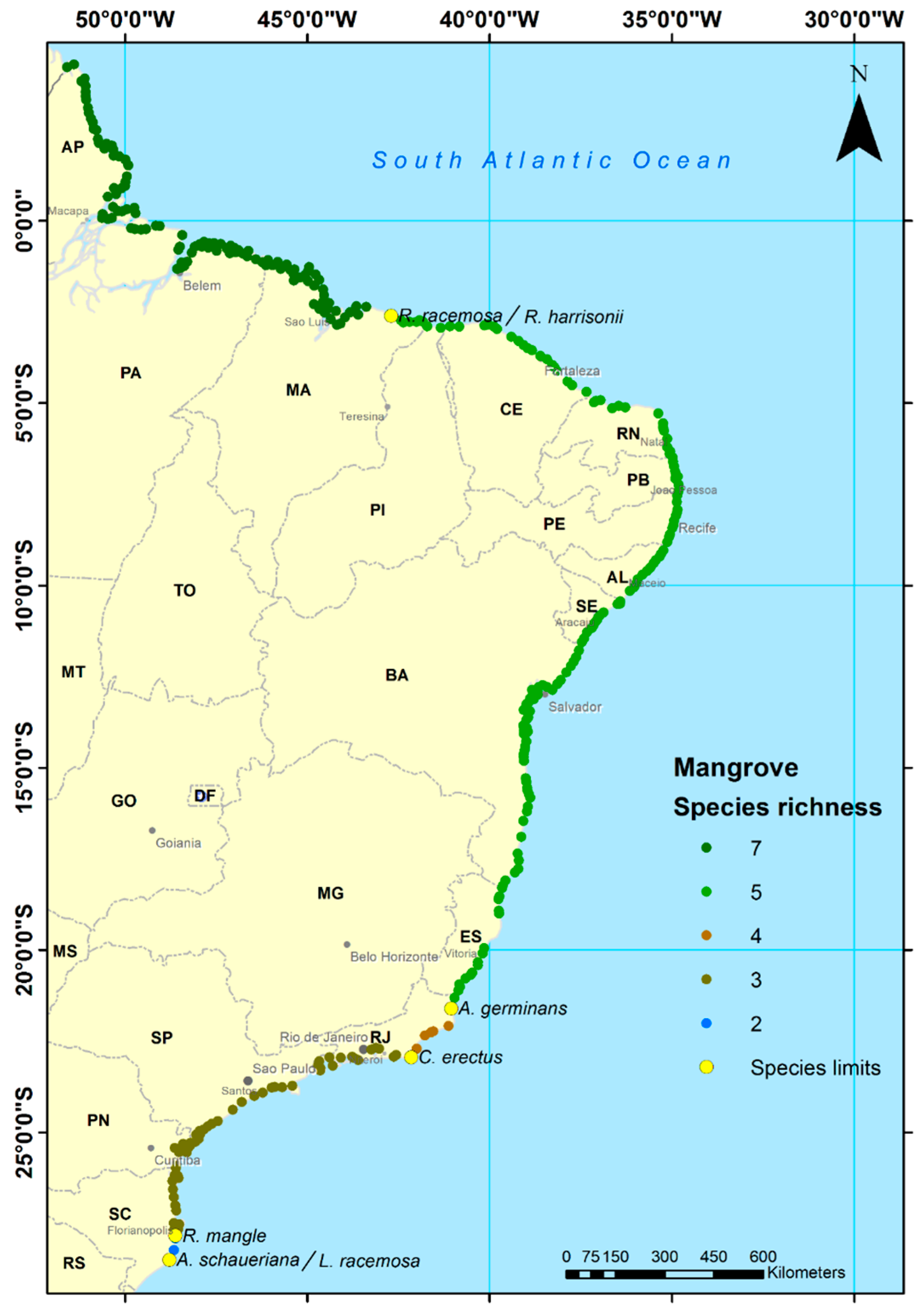
2.1. Study Area
2.2. Environmental Geodatabase
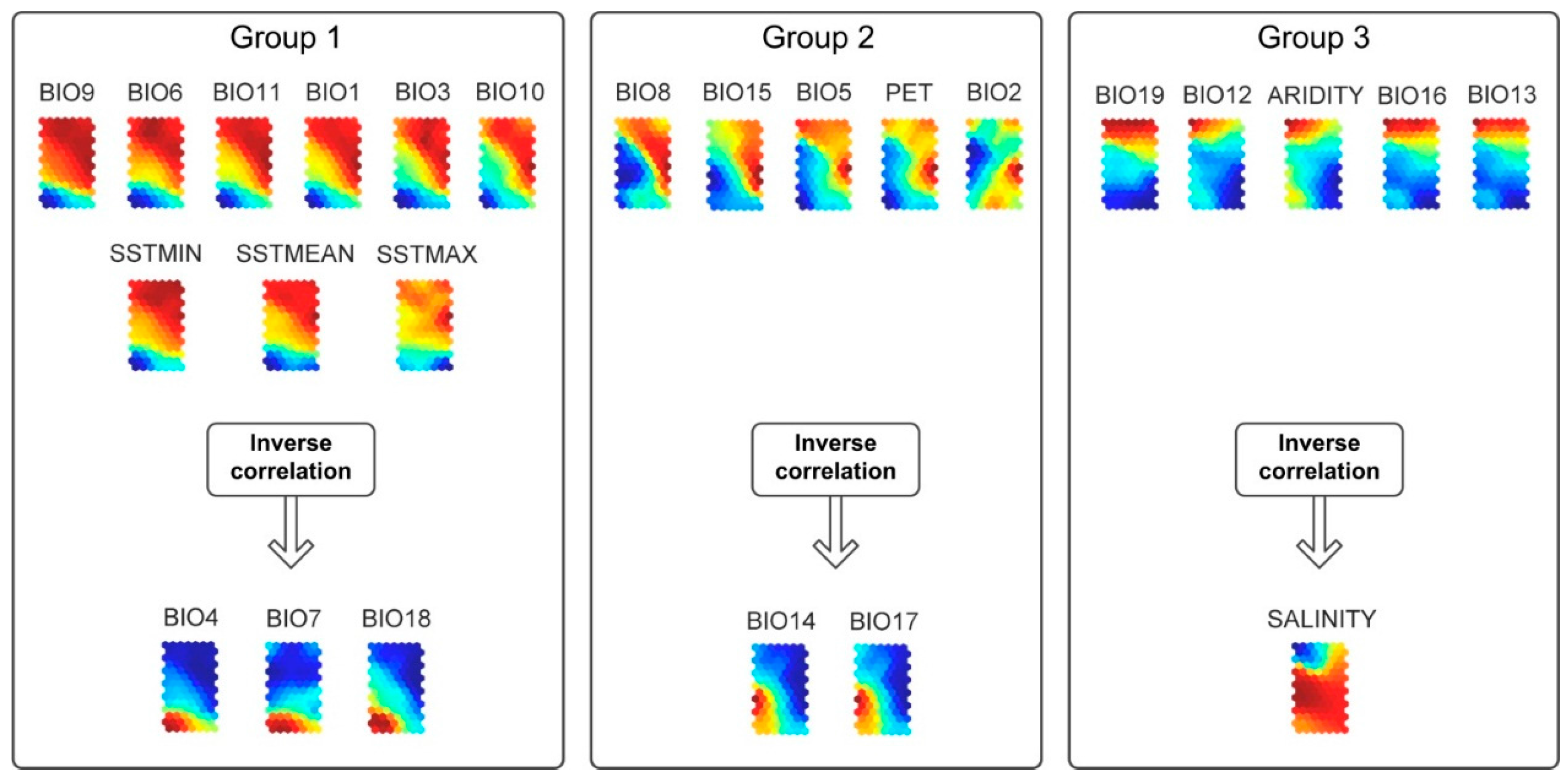
2.3. Self-Organizing Maps (Data-Driven Approach)
2.4. Descriptive Analysis
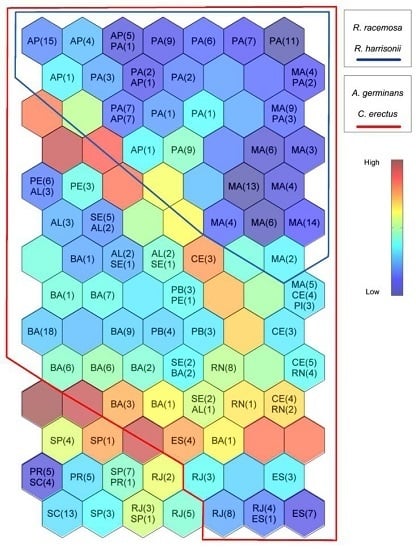
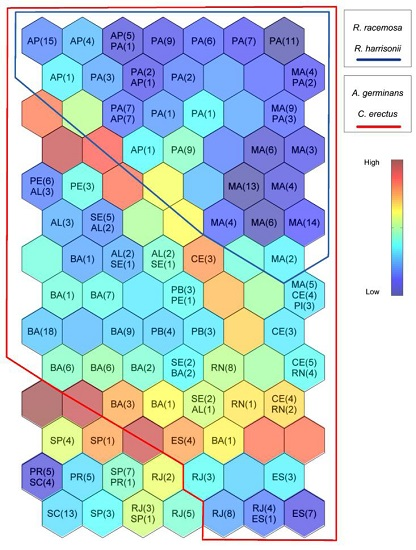
3. Results
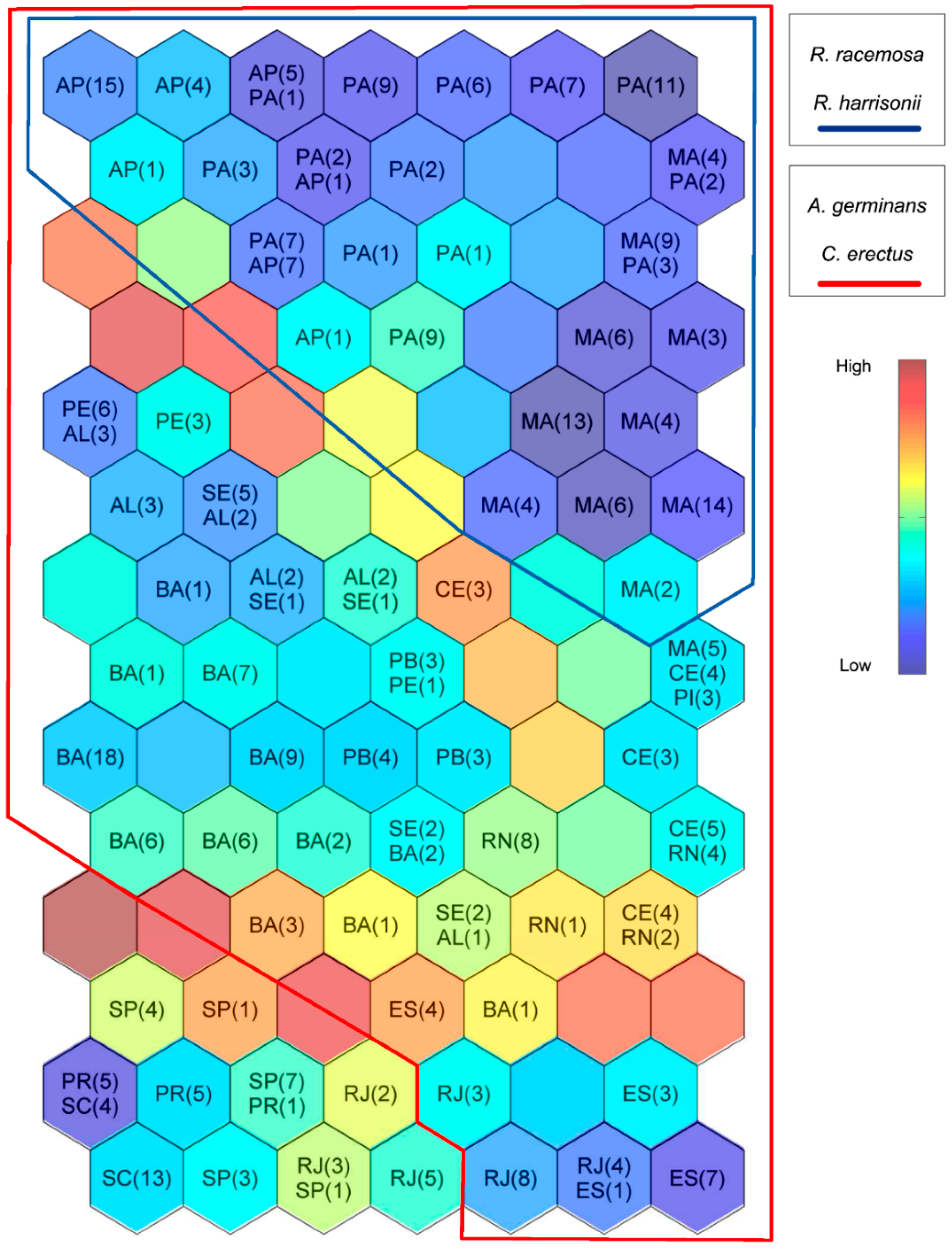
3.1. Self-Organizing Maps
3.2. Environmental Description
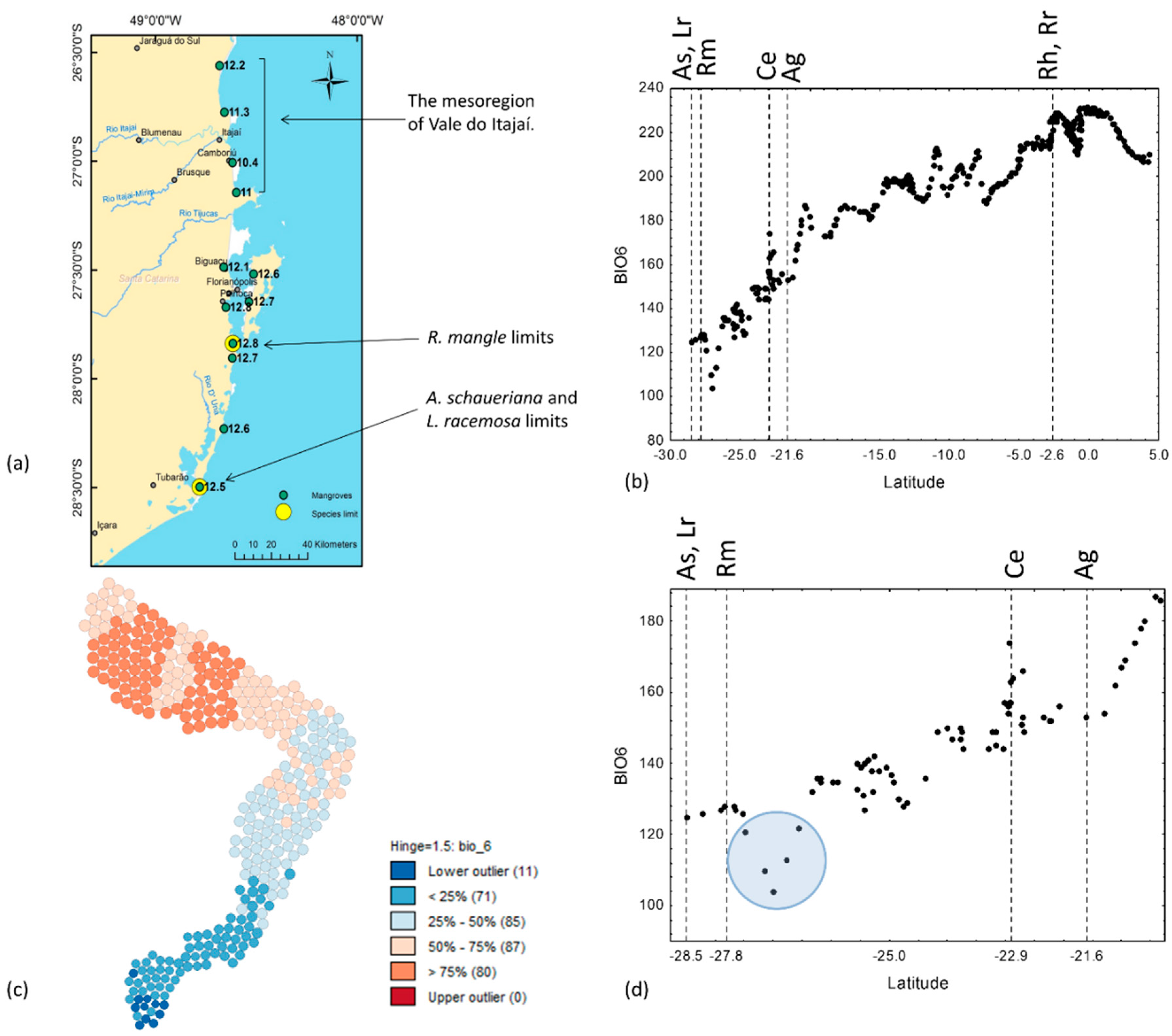
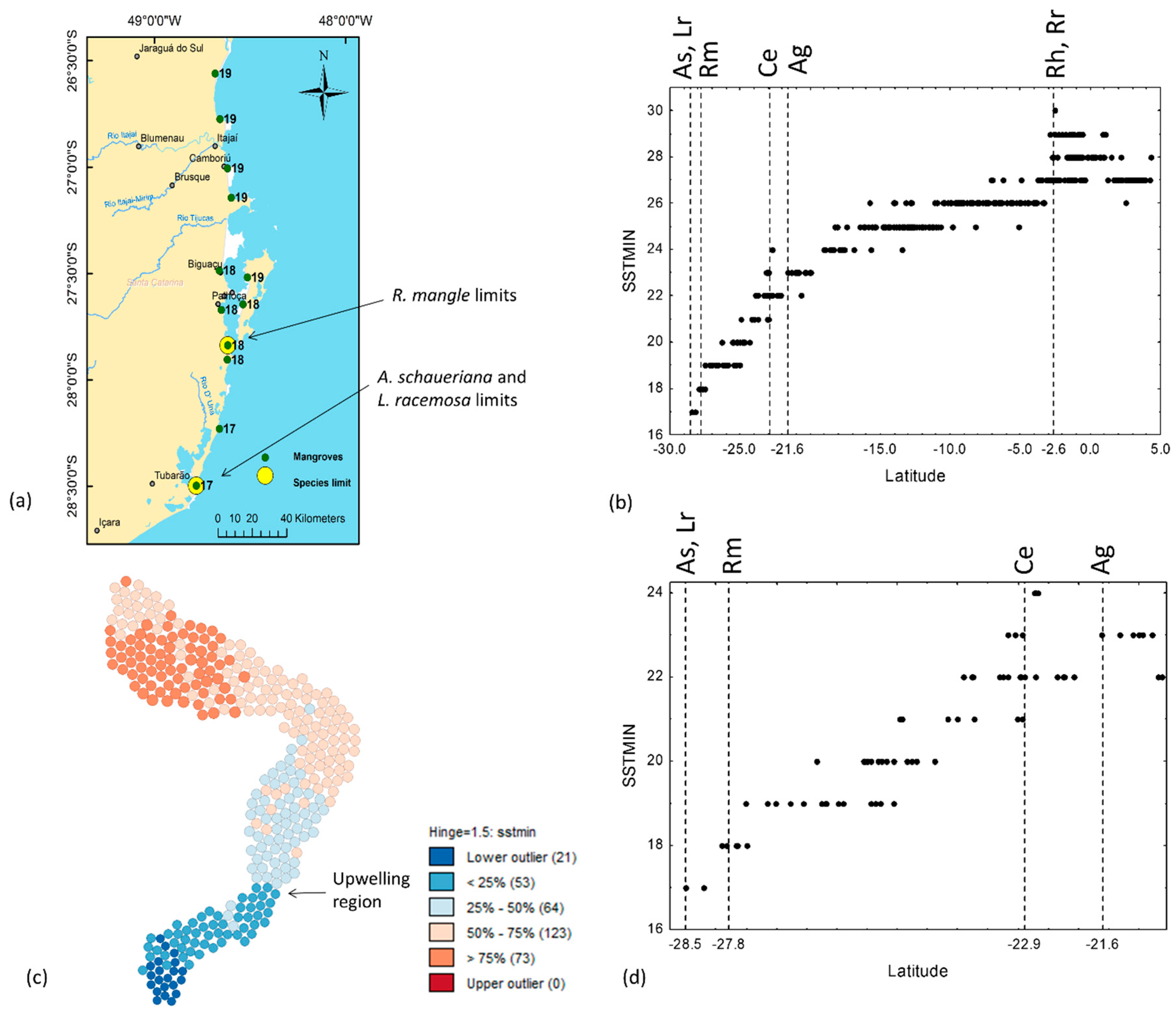
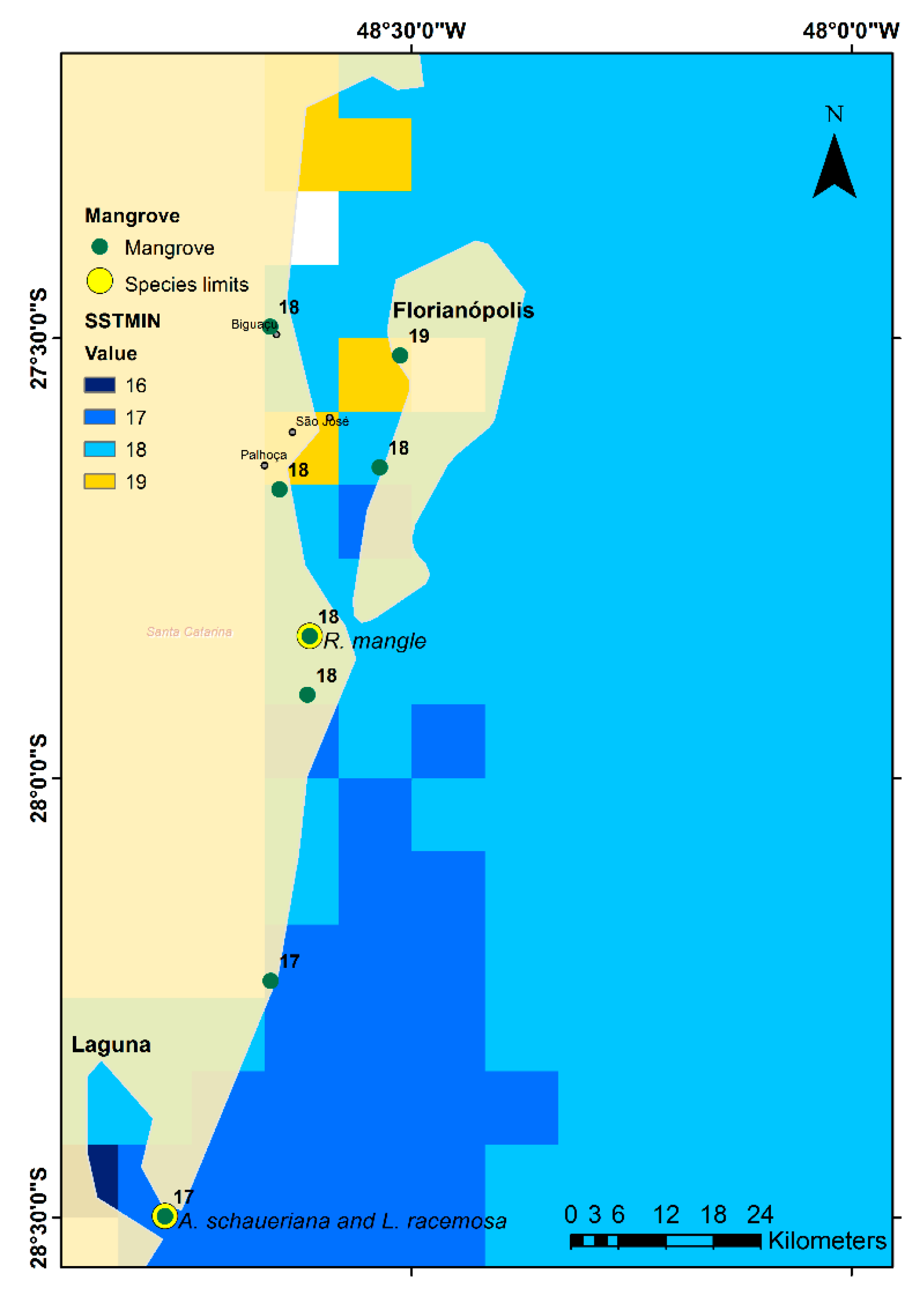
3.3. Air Temperature and Sea Surface Temperature
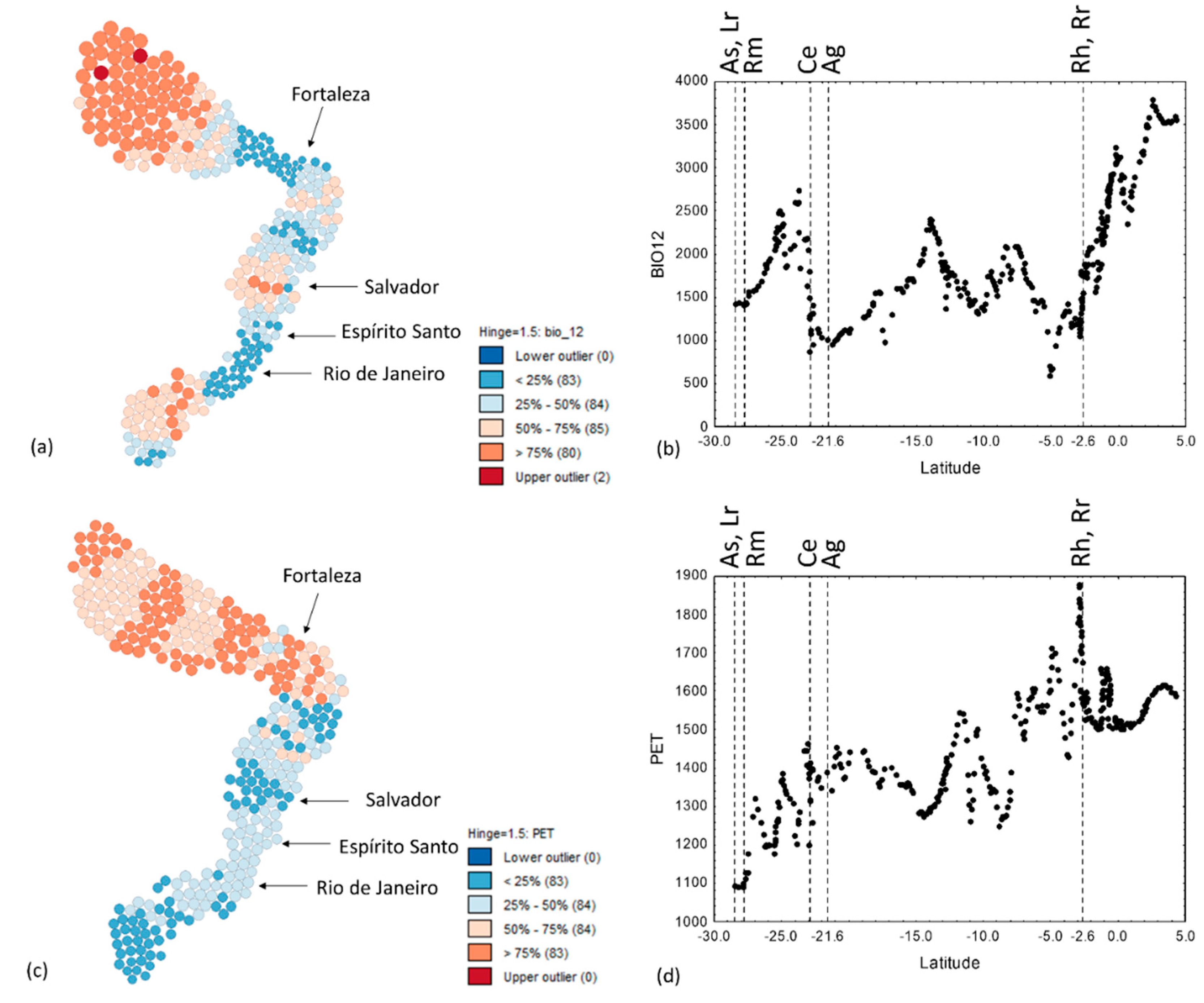
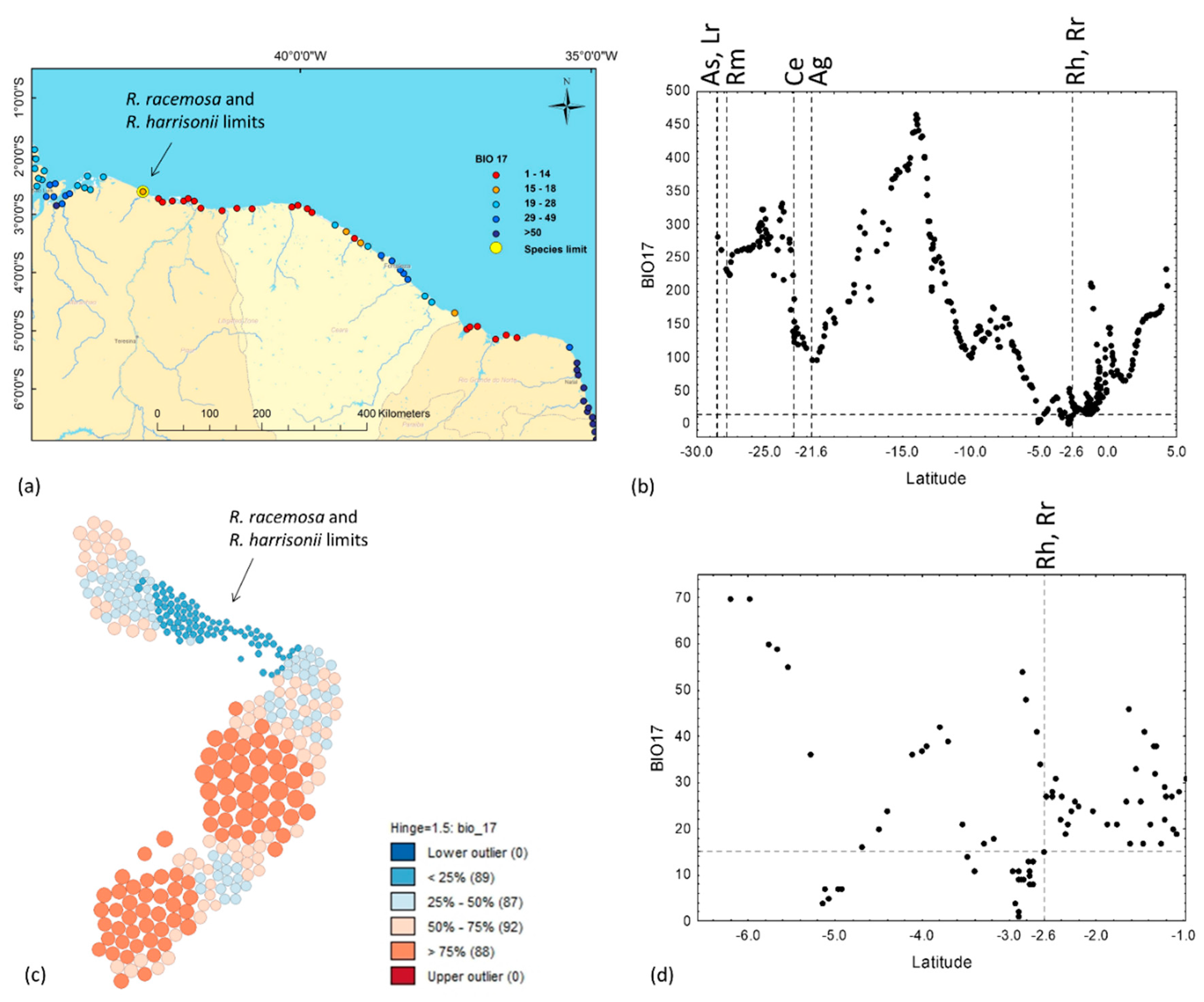
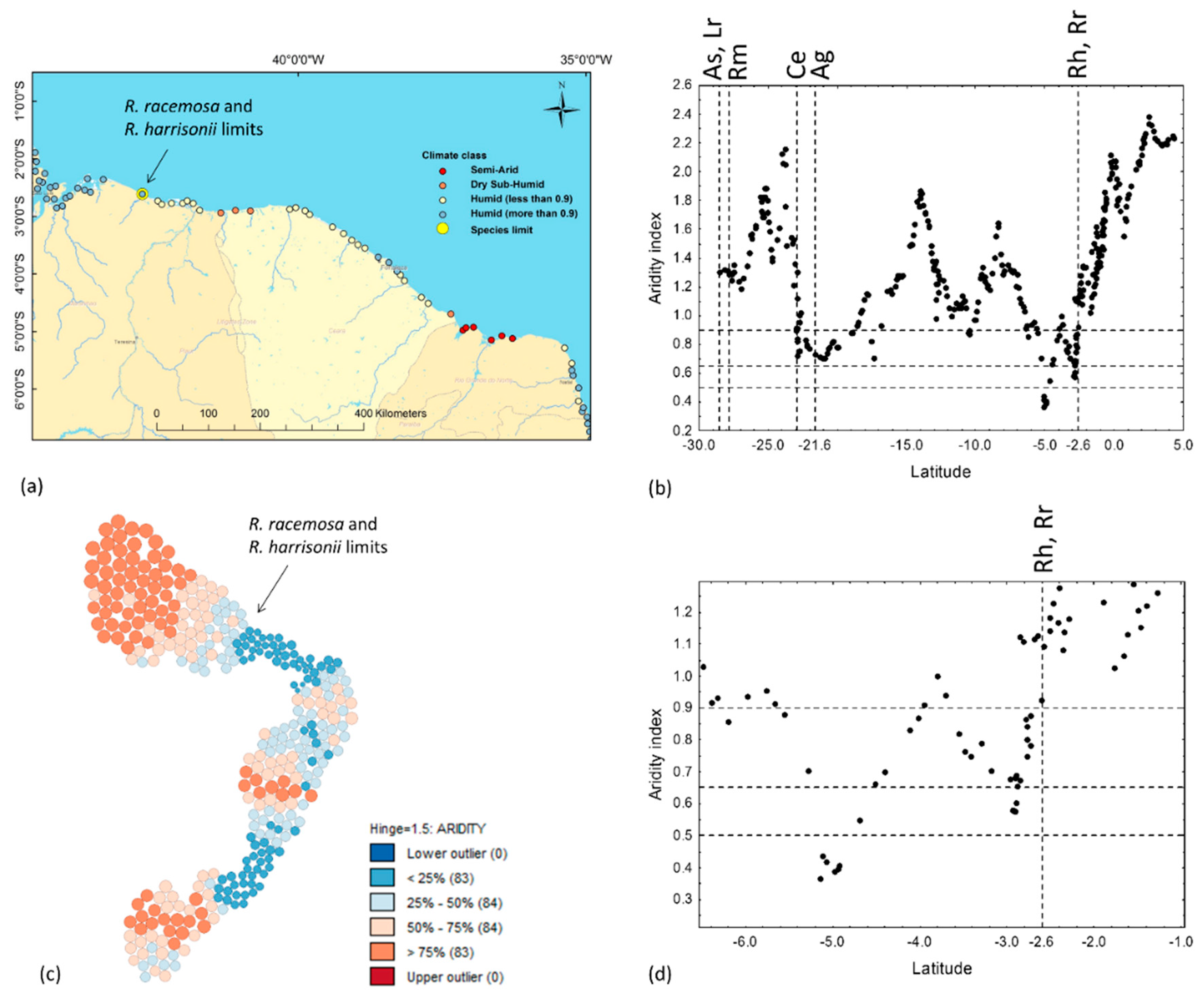
3.4. Annual Precipitation, PET and the Precipitation of the Driest Quarter
4. Discussion
4.1. Explaining the Species Limits
4.2. Comparison Climate Databases
4.3. Mangrove Mappings
4.4. Limitation of This Work
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pugnaire, F.I.; Valladares, F. Functional Plant Ecology; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2007. [Google Scholar]
- Schaeffer-Novelli, Y.; Cintrón-Molero, G.; Adaime, R.R.; Camargo, T.M. Variability of mangrove ecosystems along the Brazilian coast. Estuaries 1990, 13, 204–218. [Google Scholar] [CrossRef]
- de Larcila, L.N.; Morellato, L.P.C.; Machado, I.C. Reproductive phenology of a northeast Brazilian mangrove community: Environmental and biotic constraints. Flora Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 682–692. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A.J. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Boyer, T.P.; Antonov, J.I.; Baranova, O.K.; Garcia, H.E.; Johnson, D.R.; Locarnini, R.A.; Mishonov, A.V.; O’Brien, T.D.; Seidov, D.; Smolyar, I.V; et al. World Ocean Database 2009; Levitus, S., Ed.; NOAA Atlas.; U.S. Gov. Printing Office: Washington, DC, USA, 2009.
- Feldman, G.C.; McClain, C.R. Ocean Color Web. Available online: http://oceancolor.gsfc.nasa.gov/ (accessed on 12 Februray 2015).
- Spalding, M.; Blasco, F.; Field, C. World Mangrove Atlas; International Society for Mangrove Ecosystems: Okinawa, Japan, 1997. [Google Scholar]
- Spalding, M.; Kainuma, M.; Collins, L. World Atlas of Mangroves; Earthscan: London, UK, 2010. [Google Scholar]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Newman, H.B.; Ellisman, M.H.; Orcutt, J.A. Data-intensive e-science frontier research. Commun. ACM 2003, 46, 68–77. [Google Scholar] [CrossRef]
- Kelling, S.; Hochachka, W.M.; Fink, D.; Riedewald, M.; Caruana, R.; Ballard, G.; Hooker, G. Data-intensive science: A new paradigm for biodiversity studies. Bioscience 2009, 59, 613–620. [Google Scholar] [CrossRef]
- Kumpumäki, T.; Ruusuvuori, P.; Kangasniemi, V.; Lipping, T. Data-driven approach to benthic Cover type classification using bathymetric LiDAR waveform analysis. Remote Sens. 2015, 7, 13390–13409. [Google Scholar] [CrossRef]
- Kohonen, T. Self-organized formation of topologically correct feature maps. Biol. Cybern. 1982, 43, 59–69. [Google Scholar] [CrossRef]
- Kohonen, T. Self-Organizing Maps, 3rd ed.; Springer-Verlag Berlin Heidelberg: Berlin/Heidelberg, Germany, 2001. [Google Scholar]
- Koua, E.L.; Maceachren, A.; Kraak, M.-J. Evaluating the usability of visualization methods in an exploratory geovisualization environment. Int. J. Geogr. Inf. Sci. 2006, 20, 425–448. [Google Scholar] [CrossRef]
- Ximenes, A.C. Mapas Auto-Organizáveis para a Identificação de Ecorregiões no Interflúvio Madeira-Purus: Uma Abordagem da Biogeografia Ecológica; Instituto Nacional de Pesquisas Espaciais: São José dos Campos, Brazil, 2008. [Google Scholar]
- Ximenes, A.C.; Amaral, S.; Monteiro, A.M.V.; Valeriano, D.M. A Self-organized map neural network approach for ecoregions mapping at Brazilian Amazon Forest. In First Latin American Landscape Ecology Conference; International Association for Landscape Ecology (IALE): Campos do Jordão, Brazil, 2009. [Google Scholar]
- Ximenes, A.C.; Amaral, S.; Valeriano, D.M. Os impactos dos cenários gerais de mudanças climáticas sobre as ecorregiões da Amazônia Legal; INPE: São José dos Campos, Brazil, 2012. [Google Scholar]
- Fendereski, F.; Vogt, M.; Payne, M.R.; Lachkar, Z.; Gruber, N.; Salmanmahiny, A.; Hosseini, S.A. Biogeographic classification of the Caspian Sea. Biogeosciences 2014, 11, 6451–6470. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Hoyos, A.; Martínez, B.; García-Haro, F.; Moreno, Á.; Gilabert, M. Identification of ecosystem functional types from coarse resolution imagery using a self-organizing map approach: A case study for Spain. Remote Sens. 2014, 6, 11391–11419. [Google Scholar] [CrossRef]
- Arcoverde, G.F.B.; Almeida, C.M.; Ximenes, A.C.; Maeda, E.E.; Araújo, L.S. de Identificação de áreas prioritárias para recuperação florestal com o uso de rede neural de mapas auto-organizáveis. Bol. Ciências Geodésicas 2011, 17, 379–400. [Google Scholar] [CrossRef]
- Ximenes, A.C.; Amaral, S.; Arcoverde, G.F.B.; Monteiro, A.M.V. Redes neurais para a seleção de variáveis ambientais no processo de modelagem de distribuição de espécies na região Norte do Brasil. In XIV Simpósio Brasileiro de Sensoriamento Remoto; INPE: Natal, Brazil, 2009; pp. 5531–5538. [Google Scholar]
- Vesanto, J.; Himberg, J.; Alhoniemi, E.; Parhankangas, J. SOM toolbox for Matlab. Available online: http://www.cis.hut.fi/somtoolbox/ (accessed on 26 May 2016).
- Giraudel, J.L.; Lek, S. A comparison of self-organizing map algorithm and some conventional statistical methods for ecological community ordination. Ecol. Modell. 2001, 146, 329–339. [Google Scholar] [CrossRef]
- Crisci, J.V.; Sala, O.E.; Katinas, L.; Posadas, P. Bridging historical and ecological approaches in biogeography. Aust. Syst. Bot. 2006, 19, 1–10. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Concluding Remarks. Cold Spring Harb. Symp. Quant. Biol. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- FAO. The World’s Mangroves 1980–2005; FAO Forest.; Food and Agricultural Organization: Rome, Italy, 2007. [Google Scholar]
- Saenger, P.; Hegerl, E.J.J.; Davie, J.D.S. Global status of mangrove ecosystems. Environmentalist 1983, 3, 80–88. [Google Scholar]
- Lacerda, L.; Conde, J.E.; Alarcon, C.; Alvarez-León, R.; Bacon, P.; D’Croz, L.; Kjerfve, B.; Polanía, J.; Vannucci, M. Mangrove Ecosystems of Latin America and the Caribbean: A Summary; Lacerda, L.D., Ed.; Mangrove Ecosystems Technical Reports; International Society for Mangrove Ecosystems: Okinawa, Japan, 1993. [Google Scholar]
- Kuenzer, C.; Bluemel, A.; Gebhardt, S.; Quoc, T.V.; Dech, S. Remote sensing of mangrove ecosystems: A review. Remote Sens. 2011, 3, 878–928. [Google Scholar] [CrossRef]
- Heumann, B.W. Satellite remote sensing of mangrove forests: Recent advances and future opportunities. Prog. Phys. Geogr. 2011, 35, 87–108. [Google Scholar] [CrossRef]
- Kovacs, J.M.; King, J.M.L.; Flores de Santiago, F.; Flores-Verdugo, F. Evaluating the condition of a mangrove forest of the Mexican Pacific based on an estimated leaf area index mapping approach. Environ. Monit. Assess. 2009, 157, 137–149. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.J.; Merton, E.J.; Muller-Karger, F.E. Improved coastal wetland mapping using very-high 2-meter spatial resolution imagery. Int. J. Appl. Earth Obs. Geoinf. 2015, 40, 11–18. [Google Scholar] [CrossRef]
- Ximenes, A.C. Global mangrove mapping: A critical tool for conservation. GLP News 2015, 12, 65–69. [Google Scholar]
- Zomer, R.J.; Trabucco, A.; Van Straaten, O.; Bossio, D.A. Carbon, Land and Water: A Global Analysis of the Hydrologic Dimensions of Climate Change Mitigation through Afforestation/Reforestation; IWMI Resea; International Water Management Institute: Colombo, Sri Lanka, 2006; Volume 101. [Google Scholar]
- Zomer, R.J.; Trabucco, A.; Bossio, D.A.; van Straaten, O.; Verchot, L.V. Climate change mitigation: A spatial analysis of global land suitability for clean development mechanism afforestation and reforestation. Agric. Ecosyst. Environ. 2008, 126, 67–80. [Google Scholar] [CrossRef]
- United Nations Environment Programme (UNEP). World Atlas of Desertification, 2nd ed.; UNEP: London, UK, 1997. [Google Scholar]
- Tyberghein, L.; Verbruggen, H.; Pauly, K.; Troupin, C.; Mineur, F.; De Clerck, O. Bio-ORACLE: A global environmental dataset for marine species distribution modelling. Glob. Ecol. Biogeogr. 2012, 21, 272–281. [Google Scholar] [CrossRef]
- Brown, O.B.; Minnett, P.J. MODIS Infrared Sea Surface Temperature Algorithm (ATBD 25, v2); University of Miami: Miami, FL, USA, 1999. [Google Scholar]
- Vesanto, J. Data Exploration Process Based on the Self-Organizing Map; Helsinki University of Technology: Tieteiden, (Helsinki) Teknillisten, 2002. [Google Scholar]
- Astel, A.; Tsakovski, S.; Barbieri, P.; Simeonov, V. Comparison of self-organizing maps classification approach with cluster and principal components analysis for large environmental data sets. Water Res. 2007, 41, 4566–4578. [Google Scholar] [CrossRef] [PubMed]
- Ultsch, A. Knowledge extraction from self-organizing neural networks. In Information and Classification; Opitz, O., Lausen, B., Klar, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1993; pp. 301–306. [Google Scholar]
- Poncelet, P.; Masseglia, F.; Teisseire, M. Successes and New Directions in Data Mining; Information Science Reference—Imprint of IGI Publishing: Hershey, PA, USA, 2007. [Google Scholar]
- Brereton, R.G. Self organising maps for visualising and modelling. Chem. Cent. J. 2012, 6, S1. [Google Scholar] [CrossRef] [PubMed]
- MATLAB Matlab Version 7.10.0 (R2010a) Natick, Massachusetts; The MathWorks Inc.: Natick, MA, USA, 2010.
- Ellison, A.M. Macroecology of mangroves: Large-scale patterns and processes in tropical coastal forests. Trees 2002, 16, 181–194. [Google Scholar] [CrossRef]
- Ricklefs, R.E.; Latham, R.E.; Ricklefs, R.E.; Latham, R.E. Global patterns of diversity in mangrove floras. In Species Diversity in Ecological Communities: Historical and Geographical Perspectives; Ricklefs, R.E., Schluter, D., Eds.; University of Chicago Press: Chicago, IL, USA, 1993; pp. 215–229. [Google Scholar]
- Wade, T.; Sommer, S. A to Z GIS: An Illustrated Dictionary of Geographic Information Systems, 2nd ed.; Esri Press: Redlands, CA, USA, 2006. [Google Scholar]
- Anselin, L.; Syabri, I.; Kho, Y. GeoDa: An introduction to spatial data analysis. Geogr. Anal. 2006, 38, 5–22. [Google Scholar] [CrossRef]
- Environmental Systems Research Institute. ESRI ArcGIS Desktop Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- Saenger, P. Mangrove Ecology, Silviculture and Conservation; Springer: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Saenger, P.; Moverley, J. Vegetative phenology of mangroves along the Queensland coastline. Proc. Ecol. Soc. Aust. 1985, 13, 257–265. [Google Scholar]
- Smith, T.J.; Duke, N.C. Physical determinants of inter-estuary variation in mangrove species richness around the tropical coastline of Australia. J. Biogeogr. 1987, 14, 9–19. [Google Scholar] [CrossRef]
- Jiménez, J.A.; Sauter, K. Structure and dynamics of mangrove forests along a flooding gradient. Estuaries 1991, 14, 49–56. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F.; Centre, P.C.; Wetterdienst, D. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Kampel, M.; Lorenzetti, J.A.; Silva, C.L., Jr. Observação por satélite de ressurgências na costa S-SE brasileira. In Congresso Latino-AMERICANO sobre Ciências do Mar (Colacmar); Colacmar: Santos-SP, Brazil, 1997; pp. 38–40. [Google Scholar]
- Oliveira Filho, E.C. Algas Marinhas Bentônicas do Brasil; Universidade de São Paulo: São Paulo, Brazil, 1977. [Google Scholar]
- Yoneshigue, Y. Taxonomie et ecologie des algues marines dans la région de Cabo Frio (Rio de Janeiro, Brésil); Université d'Aix Marseille: Marseille, France, 1985. [Google Scholar]
- Guimaraens, M.A.; Coutinho, R. Spatial and temporal variation of benthic marine algae at the Cabo Frio upwelling region, Rio de Janeiro, Brazil. Aquat. Bot. 1996, 52, 283–299. [Google Scholar] [CrossRef]
- Martin, L.; Flexor, J.M.; Valentin, J.L. The influence of the “El Niño” phenomenon on the enhancement or annihilation of Cabo Frio upwelling on the Brazilian coast of the State of Rio de Janeiro. In International Symposium on Global Changes in South America during the Quaternary. Special Publ. No. 1.; Associação Brasileira de Estudos do Quaternário: São Paulo, Brésil, 1989; pp. 225–227. [Google Scholar]
- Valentin, J. Analyses des paramètres hidrobiologiques darts la remontée de Cabo Frio (Brésil). Mar. Biol. 1984, 82, 259–276. [Google Scholar] [CrossRef]
- Calado, L.; da Silveira, I.C. A.; Gangopadhyay, A.; de Castro, B.M. Eddy-induced upwelling off Cape São Tomé (22°S, Brazil). Cont. Shelf Res. 2010, 30, 1181–1188. [Google Scholar] [CrossRef]
- McMillan, C. Environmental factors affecting seedling establishment of the black mangrove-D on the central Texas coast. Ecology 1971, 52, 927–930. [Google Scholar] [CrossRef]
- Tognella, M.M.P.; Oliveira, R.G. Manguezal do Rio Camboriú: Dados preliminares. Enciclopédia Biosf. 2012, 8, 1828–1844. [Google Scholar]
- Massó i Alemán, S.; Bourgeois, C.; Appeltans, W.; Vanhoorne, B.; De Hauwere, N.; Stoffelen, P.; Heughebaert, A.; Dahdouh-Guebas, F. The “Mangrove Reference Database and Herbarium”. Plant Ecol. Evol. 2010, 143, 225–232. [Google Scholar] [CrossRef]
- Stuart, S.A.; Choat, B.; Martin, K.C.; Holbrook, N.M.; Ball, M.C. The role of freezing in setting the latitudinal limits of mangrove forests. New Phytol. 2007, 173, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Quisthoudt, K.; Schmitz, N.; Randin, C.F.; Dahdouh-Guebas, F.; Robert, E.M.R.; Koedam, N. Temperature variation among mangrove latitudinal range limits worldwide. Trees 2012, 26, 1919–1931. [Google Scholar] [CrossRef]
- Soares, M.L.G.; Estrada, G.C.D.; Fernandez, V.; Tognella, M.M.P. Southern limit of the Western South Atlantic mangroves: Assessment of the potential effects of global warming from a biogeographical perspective. Estuar. Coast. Shelf Sci. 2012, 101, 44–53. [Google Scholar] [CrossRef]
- Quisthoudt, K.; Adams, J.; Rajkaran, A.; Dahdouh-Guebas, F.; Koedam, N.; Randin, C. Disentangling the effects of global climate and regional land-use change on the current and future distribution of mangroves in South Africa. Biodivers. Conserv. 2013, 22, 1369–1390. [Google Scholar] [CrossRef]
- Record, S.; Charney, N.D.; Zakaria, R.M.; Ellison, A.M. Projecting global mangrove species and community distributions under climate change. Ecosphere 2013, 4. art34. [Google Scholar] [CrossRef]
- Menezes, M.P.M.; Berger, U.; Mehlig, U. Mangrove vegetation in Amazonia: A review of studies from the coast of Pará and Maranhão States, North Brazil. Acta Amaz. 2008, 38, 403–420. [Google Scholar] [CrossRef]
- Twilley, R.R.; Rivera-Monroy, V.H.; Chen, R.; Botero, L. Adapting an ecological mangrove Model to simulate trajectories in restoration ecology. Mar. Pollut. Bull. 1998, 37, 404–419. [Google Scholar] [CrossRef]
- Lugo, A.E.; Snedaker, S.C. The ecology of mangroves. Annu. Rev. Ecol. Syst. 1974, 5, 39–64. [Google Scholar] [CrossRef]
- Blasco, F. Climatic factors and the biology of mangrove plants. In The Mangrove Ecosystem: Research Methods; Snedaker, S.C., Snedaker, J.G., Eds.; UNESCO: Bungay, UK, 1983; pp. 18–35. [Google Scholar]
- Droogers, P.; Allen, R.G. Estimating reference evapotranspiration under inaccurate data conditions. Irrig. Drain. Syst. 2002, 16, 33–45. [Google Scholar] [CrossRef]
- Higazi, L. Dispersion and Establishment of Mangrove Propagules in the Micro-Tidal Area of Pambala-Chilaw Lagoon (Sri Lanka): An Experimental Approach; Vrije Universiteit Brussel: Ixelles, Belgium, 2008. [Google Scholar]
- Lara, R.J.; Cohen, M.C.L. Sediment porewater salinity, inundation frequency and mangrove vegetation height in Bragança, North Brazil: An ecohydrology-based empirical model. Wetl. Ecol. Manag. 2006, 14, 349–358. [Google Scholar] [CrossRef]
- Santos, M.V.; Zieman, J.C.; Cohen, R.H. Interpreting the upper mid-littoral zonation patterns of mangroves in Maranhão (Brazil) in response to microtopography and hydrology. In Mangrove Ecosystem Studies in Latin America and Africa; Kjerfve, B., Lacerda, L.D., Diop, E.H., Eds.; UNESCO: Paris, France, 1997; p. 149. [Google Scholar]
- Reiser, A. Estimates of Biomass and Productivity in Fringe Mangroves of North-Brazil; University of Bremen: Bremen, Germany, 2003. [Google Scholar]
- Matthijs, S.; Tack, J.; van Speybroeck, D.; Koedam, N. Mangrove species zonation and soil redox state, sulphide concentration and salinity in Gazi Bay (Kenya), a preliminary study. Mangroves Salt Marshes 1999, 3, 243–249. [Google Scholar] [CrossRef]
- Dahdouh-Guebas, F.; Koedam, N. Are the northernmost mangroves of West Africa viable?—A case study in Banc d’Arguin National Park, Mauritania. Hydrobiologia 2001, 458, 241–253. [Google Scholar] [CrossRef]
- Dahdouh-Guebas, F.; De Bondt, R.; Abeysinghe, P.D.; Kairo, J.G.; Cannicci, S.; Triest, L.; Koedam, N. Comparative study of the disjunct zonation pattern of the grey mangrove Avicennia marina (Forsk.) Vierh. in Gazi Bay (Kenya). Bull. Mar. Sci. 2004, 74, 237–252. [Google Scholar]
- Dana, J.D. On an isothermal oceanic chart, illustrating the geographical distribution of marine animals. Am. J. Sci. 1853, 16, 314–327. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Species Abbreviation | Family | Location | Latitude | STATE | |
|---|---|---|---|---|---|---|
| Degrees | Decimal | |||||
| Rhizophora harrisonii Leechman | R. harrisonii | Rhizophoraceae | Preguiças River | 2°40'S | 2.6°S | Maranhão, MA |
| Rhizophora racemosa G.F.W. Meyer | R. racemosa | Rhizophoraceae | Preguiças River | 2°40'S | 2.6°S | Maranhão, MA |
| Avicennia germinans L. | A. germinans | Acanthaceae | Atafona | 21°37'S | 21.6°S | Rio de Janeiro, RJ |
| Conocarpus erectus L. | C. erectus | Combretaceae | Araruama | 22°55'S | 22.9°S | Rio de Janeiro, RJ |
| Rhizophora mangle L. | R. mangle | Rhizophoraceae | Praia do Sonho | 27°53'S | 27.8°S | Santa Catarina, SC |
| Laguncularia racemosa Gaertn. | L. racemosa | Combretaceae | Laguna | 28°30'S | 28.5°S | Santa Catarina, SC |
| Avicennia schaueriana Stapf. and Leech | A. schaueriana | Acanthaceae | Laguna | 28°30'S | 28.5°S | Santa Catarina, SC |
| Code | Unit | Resolution | Variables |
|---|---|---|---|
| BIO1 | °C | 2.5 arc-minute | Annual Mean Temperature |
| BIO2 | °C | Mean Diurnal Range (Mean of monthly (max temp–min temp)) | |
| BIO3 | °C | Isothermality (BIO2/BIO7) (×100) | |
| BIO4 | - | Temperature Seasonality (standard deviation × 100) | |
| BIO5 | °C | Max Temperature of Warmest Month | |
| BIO6 | °C | Min Temperature of Coldest Month | |
| BIO7 | °C | Temperature Annual Range (BIO5-BIO6) | |
| BIO8 | °C | Mean Temperature of Wettest Quarter | |
| BIO9 | °C | Mean Temperature of Driest Quarter | |
| BIO10 | °C | Mean Temperature of Warmest Quarter | |
| BIO11 | °C | Mean Temperature of Coldest Quarter | |
| BIO12 | mm | Annual Precipitation | |
| BIO13 | mm | Precipitation of Wettest Month | |
| BIO14 | mm | Precipitation of Driest Month | |
| BIO15 | - | Precipitation Seasonality (Coefficient of Variation) | |
| BIO16 | mm | Precipitation of Wettest Quarter | |
| BIO17 | mm | Precipitation of Driest Quarter | |
| BIO18 | mm | Precipitation of Warmest Quarter | |
| BIO19 | mm | Precipitation of Coldest Quarter | |
| ARIDITY | - | 30 arc-second | Aridity Index |
| PET | mm/day | Potential EvapoTranspiration | |
| SSTMIN | °C | 5 arc-minute | Min Sea Surface Temperature |
| SSTMEAN | °C | Mean Sea Surface Temperature | |
| SSTMAX | °C | Max Sea Surface Temperature | |
| SALINITY | PSU | Sea salinity |
| Species | Latitude | Richness | BIO1 | BIO6 | BIO11 | SSTMIN | SSTMEAN | SSTMAX |
|---|---|---|---|---|---|---|---|---|
| R. racemosa and R. harrisonii | 2.6°S | 7 | 27.3 | 22.1 | 26.7 | 27 | 29 | 29 |
| A. germinans | 21.6°S | 5 | 23.0 | 15.3 | 20.7 | 23 | 26 | 28 |
| C. erectus | 22.9°S | 4 | 23.0 | 17.4 | 20.9 | 22 | 25 | 27 |
| R. mangle | 27.8°S | 3 | 20.0 | 12.8 | 16.4 | 18 | 22 | 26 |
| A. schaueriana and L. racemosa | 28.5°S | 2 | 19.9 | 12.5 | 16.3 | 17 | 22 | 27 |
| Species | Latitude | Richness | BIO12 | BIO14 | BIO17 | BIO18 | Aridity |
|---|---|---|---|---|---|---|---|
| R. racemosa and R. harrisonii | 2.6°S | 7 | 1549 | 5 | 15 | 16 | 0.92 |
| A. germinans | 21.6°S | 5 | 1014 | 27 | 98 | 329 | 0.73 |
| C. erectus | 22.9°S | 4 | 870 | 39 | 125 | 242 | 0.72 |
| R. mangle | 27.8°S | 3 | 1420 | 72 | 230 | 535 | 1.29 |
| A. schaueriana and L. racemosa | 28.5°S | 2 | 1431 | 89 | 282 | 434 | 1.30 |
| Variables | Codes | This Study | S-N et al. (1990) | This Study | S-N et al. (1990) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Min | Location | Min | Location | Max | Location | Max | Location | |||
| Annual Mean Temperature | Group 1 | BIO1 | 19.8 | Florianópolis/Biguaçu, SC | <20 | Laguna, SC | 27.9 | Camocim, CE | ca. 26.8 | Recife, PE |
| Isothermality | BIO3 | 43 | Laguna, SC | 89 | near to Belém, PA | |||||
| Temperature Seasonality | BIO4 | 306 | near to Belém, PA | 3129 | Paranaguá Bay, PR | |||||
| Temperature Annual Range | BIO7 | 8.5 | Aracajú, SE | >1 | Belém, PA | 18.3 | Camboriú, SC | >8 | Laguna, SC | |
| Mean Temperature of Driest Quarter | BIO9 | 16.2 | Camboriú, SC | 28.2 | Acaraú, CE | |||||
| Mean Temperature of Warmest Quarter | BIO10 | 23.2 | Florianópolis, SC | 28.7 | Camocim, CE | |||||
| Min Temperature of Coldest Month | BIO6 | 10.4 | Camboriú, SC | 15.7 | Laguna, SC | 23.2 | Marajó Island, PA | ca. 25.5 | Belém, PA | |
| Mean Temperature of Coldest Quarter | BIO11 | 16.2 | Camboriú/Biguaçu, SC | 27.1 | Camocim, CE | |||||
| Min Sea surface temperature | SSTMIN | 17 | Laguna, SC | 30 | Icatu, MA | |||||
| Mean Sea surface temperature | SSTMEAN | 21.50 | Imbituba, SC | 31.21 | Bequimão, MA | |||||
| Max Sea surface temperature | SSTMAX | 25 | Imbituba, SC | 33 | *Four locations | |||||
| Precipitation of Warmest Quarter | BIO18 | 7 | Cajueiro da Praia, PI | 1029 | Cananéia, SP | |||||
| Mean Diurnal Range | Group 2 | BIO2 | 57 | Aracajú, SE | 110 | Camocim, CE | ||||
| Mean Temperature of Wettest Quarter | BIO8 | 22.5 | Belmonte, BA | 27.6 | Areia Branca, RN | |||||
| Max Temperature of Warmest Month | BIO5 | 27.6 | Laguna, SC | 35.1 | Camocim, CE | |||||
| Precipitation Seasonality | BIO15 | 9 | Una, BA | 118 | Camocim, CE | |||||
| Precipitation of Driest Month | BIO14 | 0 | Macau, RN | 143 | Camamu/Maraú, BA | |||||
| Precipitation of Driest Quarter | BIO17 | 1 | Camocim, CE | 466 | Camamu/Maraú, BA | |||||
| Potential Evapotranspiration | PET | 1092 | Madre River, SC | ca. 950 | Florianópolis, SC | 1877 | Camocim, CE | 1600 | Golfão-Belém, PA | |
| Annual Precipitation | Group 3 | BIO12 | 600 | Macau, RN | 1090 | Rio de Janeiro, RJ | 3791 | Nazaré, AP | 3250 | Maracá, AP |
| Precipitation of Wettest Month | BIO13 | 114 | Arraial do Cabo, RJ | 613 | Algodoal, PA | |||||
| Precipitation of Wettest Quarter | BIO16 | 300 | Arraial do Cabo, RJ | 1655 | Nazaré, AP | |||||
| Precipitation of Coldest Quarter | BIO19 | 61 | Areia Branca, RN | 1634 | Marajó Island, PA | |||||
| Aridity index | ARIDITY | 0.36 | Macau, RN | 2.38 | Nazaré, AP | |||||
| Salinity | SALINITY | 27.96 | Oiapoque River, AM | 37.17 | Belmonte, BA | |||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ximenes, A.C.; Maeda, E.E.; Arcoverde, G.F.B.; Dahdouh-Guebas, F. Spatial Assessment of the Bioclimatic and Environmental Factors Driving Mangrove Tree Species’ Distribution along the Brazilian Coastline. Remote Sens. 2016, 8, 451. https://doi.org/10.3390/rs8060451
Ximenes AC, Maeda EE, Arcoverde GFB, Dahdouh-Guebas F. Spatial Assessment of the Bioclimatic and Environmental Factors Driving Mangrove Tree Species’ Distribution along the Brazilian Coastline. Remote Sensing. 2016; 8(6):451. https://doi.org/10.3390/rs8060451
Chicago/Turabian StyleXimenes, Arimatéa C., Eduardo Eiji Maeda, Gustavo Felipe Balué Arcoverde, and Farid Dahdouh-Guebas. 2016. "Spatial Assessment of the Bioclimatic and Environmental Factors Driving Mangrove Tree Species’ Distribution along the Brazilian Coastline" Remote Sensing 8, no. 6: 451. https://doi.org/10.3390/rs8060451
APA StyleXimenes, A. C., Maeda, E. E., Arcoverde, G. F. B., & Dahdouh-Guebas, F. (2016). Spatial Assessment of the Bioclimatic and Environmental Factors Driving Mangrove Tree Species’ Distribution along the Brazilian Coastline. Remote Sensing, 8(6), 451. https://doi.org/10.3390/rs8060451