
Human Milk from Previously COVID-19-Infected Mothers: The Effect of Pasteurization on Specific Antibodies and Neutralization Capacity

 , , , , , ,
, , , , , ,  , ,
, ,  , , and
, , and  add
Show full author list
add
Show full author list
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Material Collection
2.3. Laboratory Analyses
2.3.1. Evaluation of Antibodies in the Serum and Human Milk
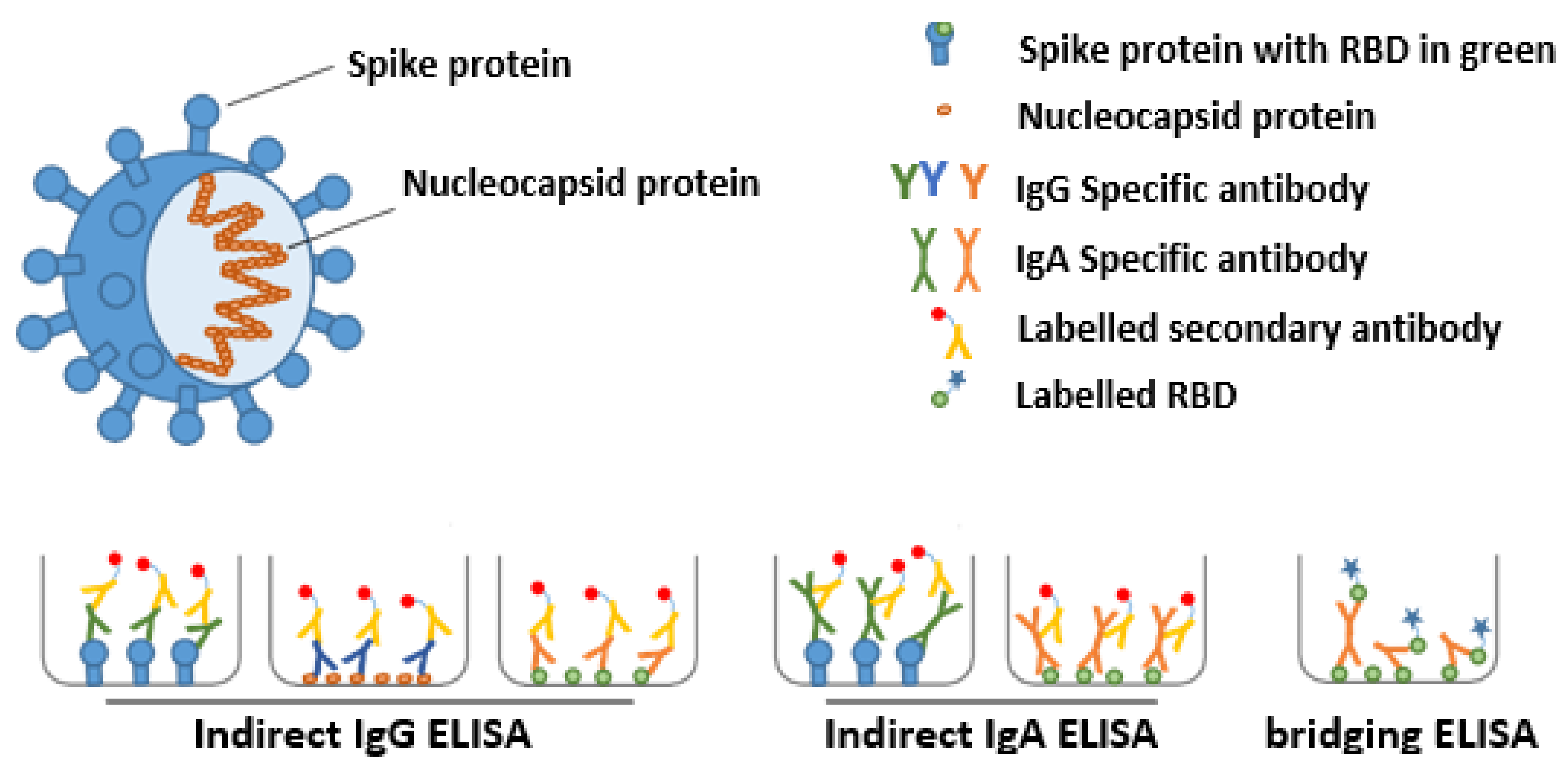
2.3.2. Detection of Anti-SARS-CoV-2 Ig in Serum and Human Milk with ELISA
2.3.3. Bridging ELISA with the SARS-CoV-2 RBD and Nucleocapsid Protein
2.3.4. Effect of Antibodies on Virus Replication
Pseudovirus Neutralization Assay
Replication Inhibition of a SARS-CoV-2 Clinical Isolate Assay
2.3.5. Evaluation of the Effect of Pasteurization of Human Milk on SARS-CoV-2 Antibodies
2.3.6. Monitoring IgA Clone Diversity in Human Milk by Mass Spectrometry
2.4. Statistical Analysis
3. Results
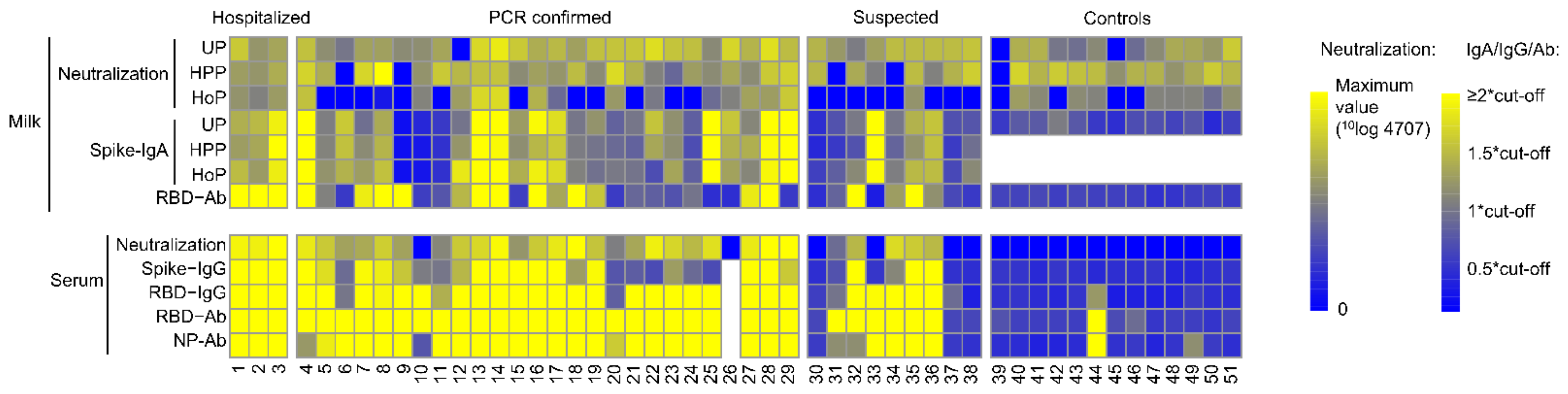
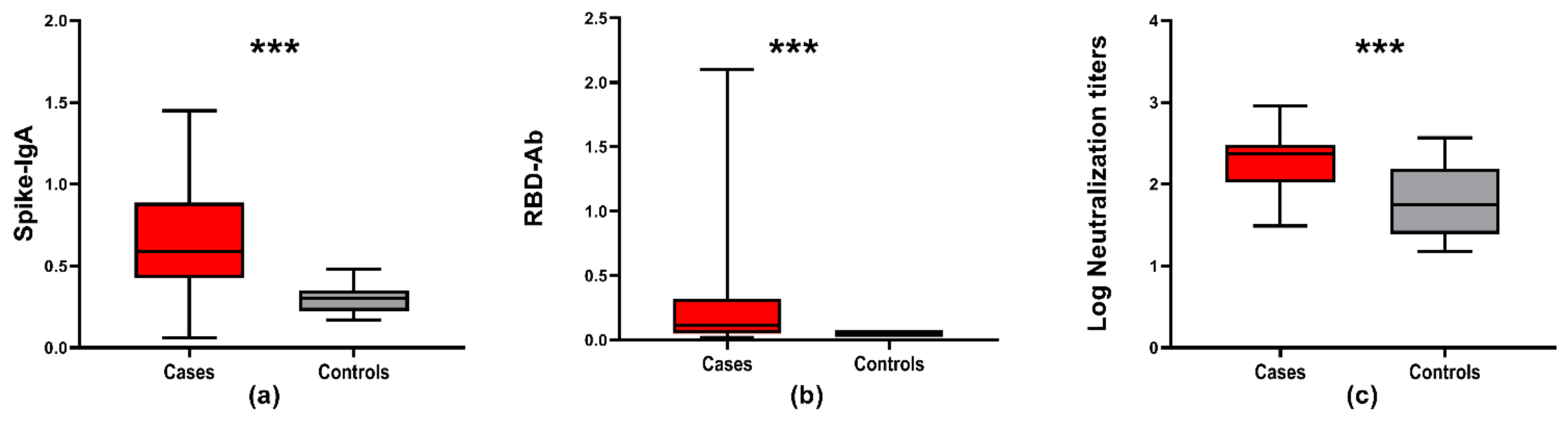
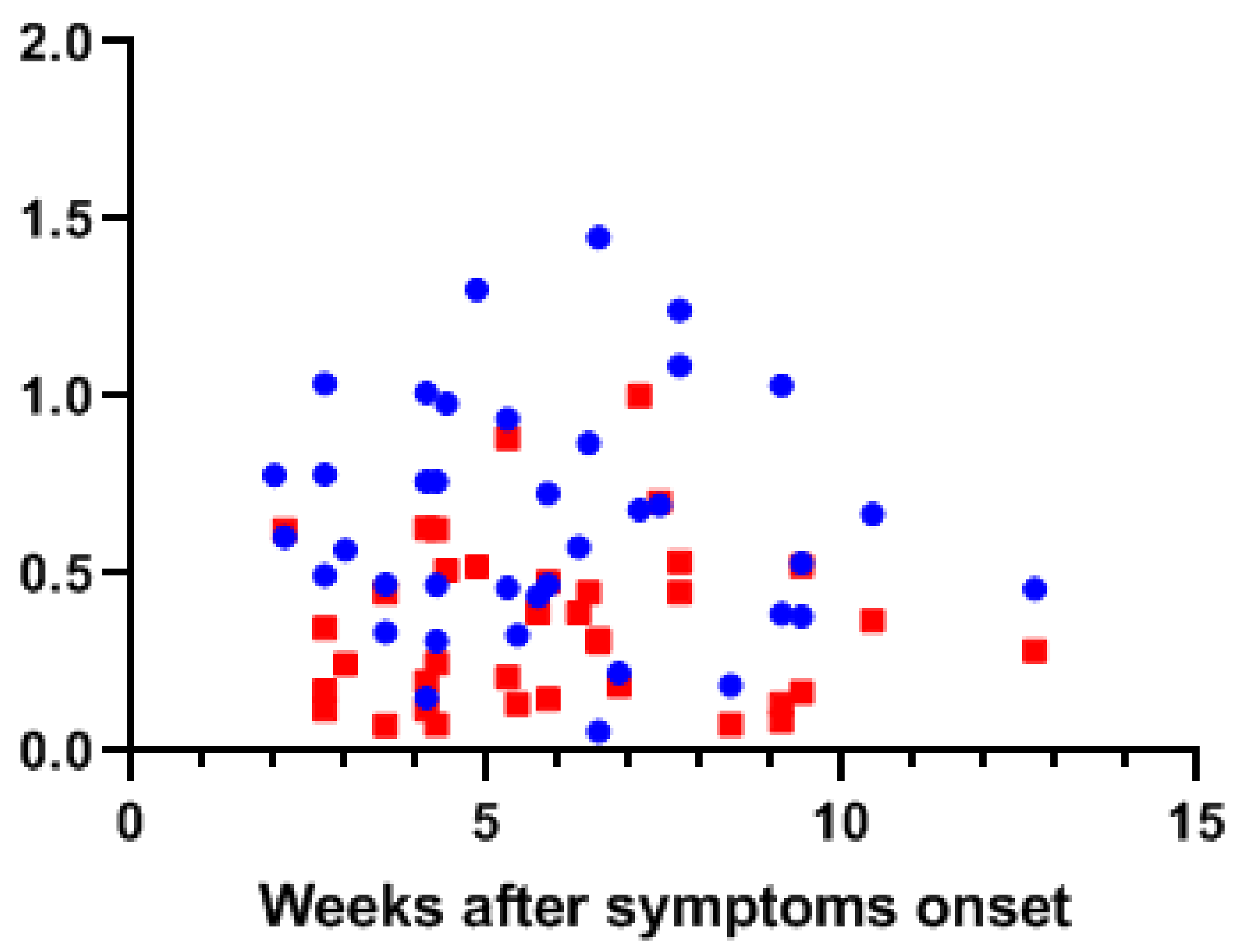
3.1. SARS-CoV-2 Antibodies in Human Milk and Serum
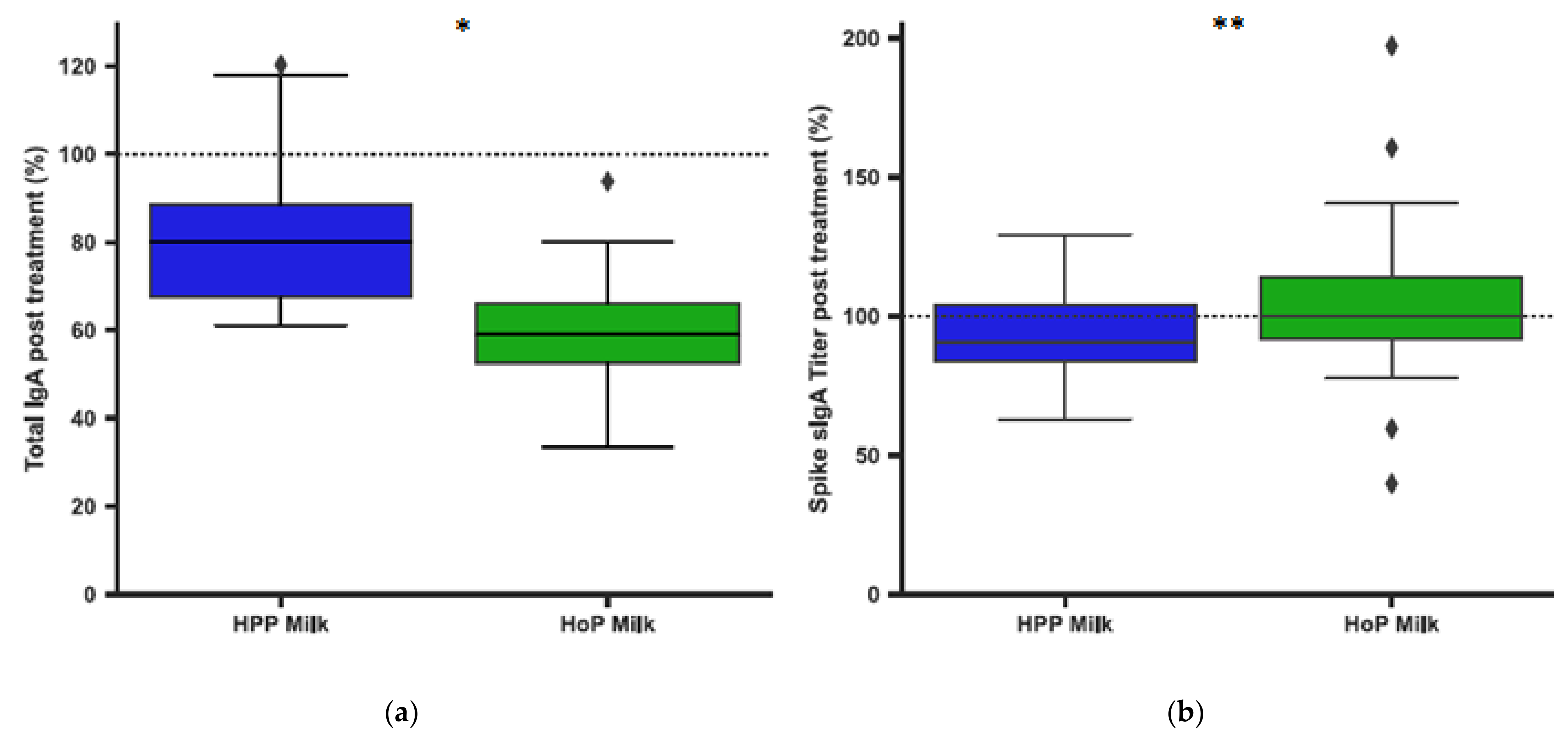
3.2. Pasteurization of Human Milk and IgA Antibodies
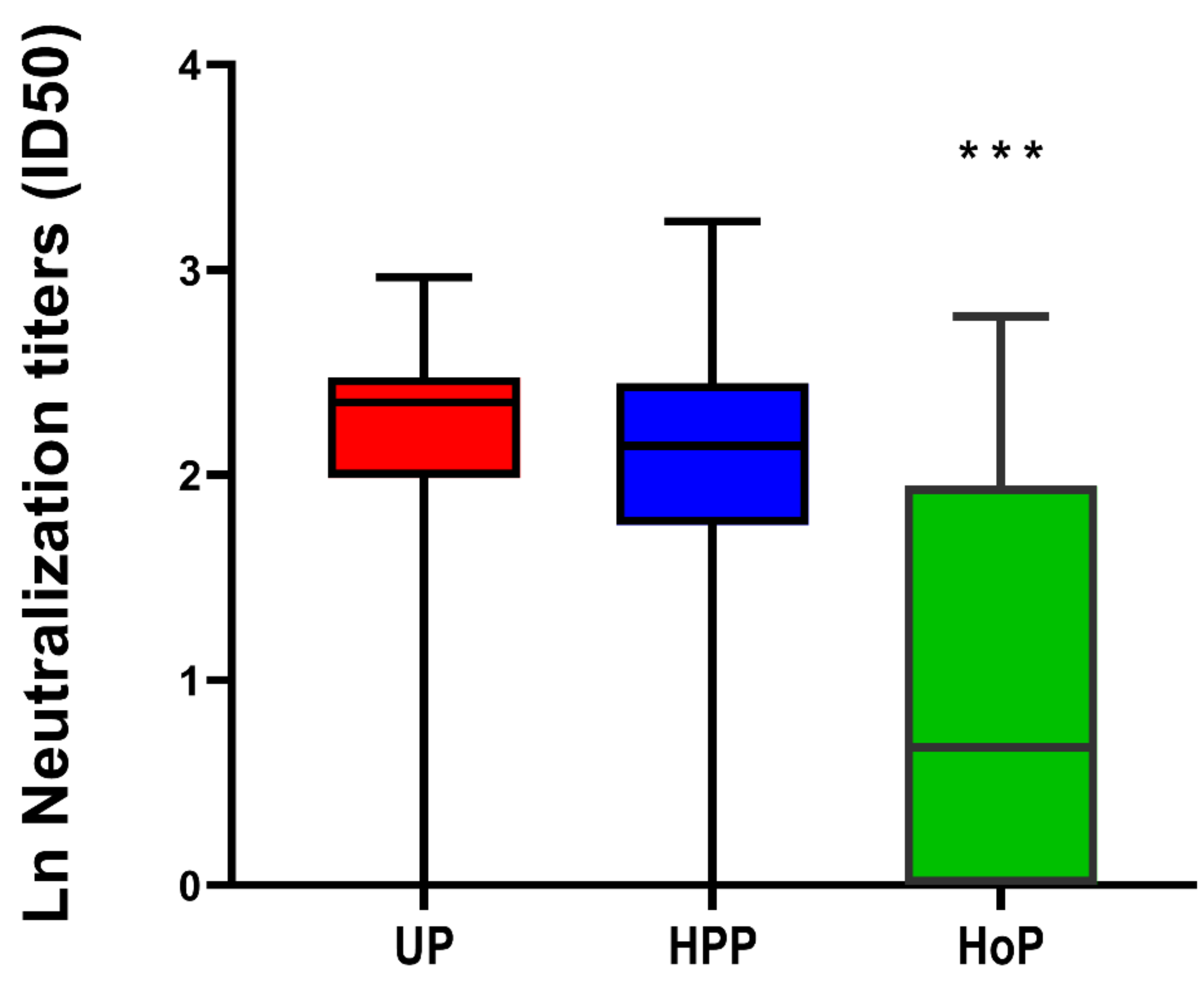
3.3. SARS-CoV-2 Virus Neutralization in Unpasteurized and Pasteurized Milk
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mullard, A. COVID-19 vaccine development pipeline gears up. Lancet 2020, 395, 1751–1752. [Google Scholar] [CrossRef]
- Agostoni, C.; Braegger, C.; Decsi, T.; Kolacek, S.; Koletzko, B.; Michaelsen, K.F.; Mihatsch, W.; Moreno, L.A.; Puntis, J.; Shamir, R.; et al. Breast-feeding: A commentary by the ESPGHAN Committee on Nutrition. J. Pediatr. Gastr. Nutr. 2009, 49, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Chantry, C.J.; Howard, C.R.; Auinger, P. Full breastfeeding duration and associated decrease in respiratory tract infection in US children. Pediatrics 2006, 117, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Hanson, L.A. Breastfeeding provides passive and likely long-lasting active immunity. Ann. Allergy Asthma. Immunol. 1998, 81, 523–537. [Google Scholar] [CrossRef]
- Mantis, N.J.; Rol, N.; Corthesy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Van de Perre, P. Transfer of antibody via mother’s milk. Vaccine 2003, 21, 3374–3376. [Google Scholar] [CrossRef]
- Hutchings, A.B.; Helander, A.; Silvey, K.J.; Chandran, K.; Lucas, W.T.; Nibert, M.L.; Neutra, M.R. Secretory immunoglobulin A antibodies against the sigma1 outer capsid protein of reovirus type 1 Lang prevent infection of mouse Peyer’s patches. J. Virol. 2004, 78, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Yang, M.; Hong, Z.; Zhang, L.; Huang, Z.; Chen, X.; He, S.; Zhou, Z.; Zhou, Z.; Chen, Q.; et al. Crystal structure of SARS-CoV-2 nucleocapsid protein RNA binding domain reveals potential unique drug targeting sites. Acta Pharm. Sin. B 2020, 10, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Schlaudecker, E.P.; Steinhoff, M.C.; Omer, S.B.; McNeal, M.M.; Roy, E.; Arifeen, S.E.; Dodd, C.N.; Raqib, R.; Breiman, R.F.; Zaman, K. IgA and neutralizing antibodies to influenza a virus in human milk: A randomized trial of antenatal influenza immunization. PLoS ONE 2013, 8, e70867. [Google Scholar] [CrossRef]
- Demers-Mathieu, V.; Huston, R.K.; Markell, A.M.; McCulley, E.A.; Martin, R.L.; Dallas, D.C. Antenatal Influenza A-Specific IgA, IgM, and IgG Antibodies in Mother’s Own Breast Milk and Donor Breast Milk, and Gastric Contents and Stools from Preterm Infants. Nutrients 2019, 11, 1567. [Google Scholar] [CrossRef]
- Fox, A.; Marino, J.; Amanat, F.; Krammer, F.; Hahn-Holbrook, J.; Zolla-Pazner, S.; Powell, R.L. Robust and Specific Secretory IgA Against SARS-CoV-2 Detected in Human Milk. iScience 2020, 23, 101735. [Google Scholar] [CrossRef]
- Italian Association of Human Milk Banks Associazione Italiana Banche del Latte Umano Donato; Arslanoglu, S.; Bertino, E.; Tonetto, P.; de Nisi, G.; Ambruzzi, A.M.; Biasini, A.; Profeti, C.; Spreghini, M.R.; Moro, G.E. Guidelines for the establishment and operation of a donor human milk bank. J. Matern. Fetal Neonatal Med. 2010, 23, 1–20. [Google Scholar] [CrossRef]
- Ford, J.E.; Law, B.A.; Marshall, V.M.; Reiter, B. Influence of the heat treatment of human milk on some of its protective constituents. J. Pediatr 1977, 90, 29–35. [Google Scholar] [CrossRef]
- Demers-Mathieu, V.; Huston, R.K.; Markell, A.M.; McCulley, E.A.; Martin, R.L.; Spooner, M.; Dallas, D.C. Differences in Maternal Immunoglobulins within Mother’s Own Breast Milk and Donor Breast Milk and across Digestion in Preterm Infants. Nutrients 2019, 11, 920. [Google Scholar] [CrossRef]
- Viazis, S.; Farkas, B.E.; Allen, J.C. Effects of high-pressure processing on immunoglobulin A and lysozyme activity in human milk. J. Hum. Lact. 2007, 23, 253–261. [Google Scholar] [CrossRef]
- Wesolowska, A.; Sinkiewicz-Darol, E.; Barbarska, O.; Bernatowicz-Lojko, U.; Borszewska-Kornacka, M.K.; van Goudoever, J.B. Innovative Techniques of Processing Human Milk to Preserve Key Components. Nutrients 2019, 11, 1169. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM Structure of the 2019-nCoV Spike in the Prefusion Conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef]
- Vogelzang, E.H.; Loeff, F.C.; Derksen, N.I.; Kruithof, S.; Ooijevaar-de Heer, P.; van Mierlo, G.; Linty, F.; Mok, J.Y.; van Esch, W.; de Bruin, S.; et al. Development of a SARS-CoV-2 Total Antibody Assay and the Dynamics of Antibody Response over Time in Hospitalized and Nonhospitalized Patients with COVID-19. J. Immunol. 2020, 205, 3491–3499. [Google Scholar] [CrossRef]
- Schmidt, F.; Schmidt, F.; Weisblum, Y.; Muecksch, F.; Hoffmann, H.H.; Michailidis, E.; Lorenzi, J.C.; Mendoza, P.; Rutkowska, M.; Bednarski, E.; et al. Measuring SARS-CoV-2 neutralizing antibody activity using pseudotyped and chimeric viruses. J. Exp. Med. 2020, 217, e20201181. [Google Scholar] [CrossRef]
- Martínez-Monteagudo, S.I.; Balasubramaniam, V.M. Fundamentals and Applications of High-Pressure Processing. In High Pressure Processing of Food; Balasubramaniam, V., Barbosa-Cánovas, G., Lelieveld, H., Eds.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Timmermans, R.; Hayrapetyan, H.; Vollebregt, M.; Dijksterhuis, J. Comparing thermal inactivation to a combined process of moderate heat and high pressure: Effect on ascospores in strawberry puree. Int. J. Food Microbiol. 2020, 325, 108629. [Google Scholar] [CrossRef]
- Brouwer, P.J.M.; Caniels, T.G.; van der Straten, K.; Snitselaar, J.L.; Aldon, Y.; Bangaru, S.; Torres, J.L.; Okba, N.M.; Claireaux, M.; Kerster, G.; et al. Potent neutralizing antibodies from COVID-19 patients define multiple targets of vulnerability. Science 2020, 369, 643–650. [Google Scholar] [CrossRef]
- Fan, H.; Hong, B.; Luo, Y.; Peng, Q.; Wang, L.; Jin, X.; Chen, Y.; Hu, Y.; Shi, Y.; Li, T.; et al. The effect of whey protein on viral infection and replication of SARS-CoV-2 and pangolin coronavirus in vitro. Signal Transduct. Target. Ther. 2020, 5, 1–3. [Google Scholar] [CrossRef]
- Okba, N.M.A.; Müller, M.A.; Li, W.; Wang, C.; GeurtsvanKessel, C.H.; Corman, V.M.; Lamers, M.M.; Sikkema, R.S.; de Bruin, E.; Chandler, F.D.; et al. SARS-CoV-2 specific antibody responses in COVID-19 patients. MedRxiv 2020. [Google Scholar] [CrossRef]
- Sterlin, D.; Mathian, A.; Miyara, M.; Mohr, A.; Anna, F.; Claër, L.; Quentric, P.; Fadlallah, J.; Devilliers, H.; Ghillani, P.; et al. IgA dominates the early neutralizing antibody response to SARS-CoV-2. Sci. Transl. Med. 2021, 13, eabd2223. [Google Scholar] [CrossRef]
- Sousa, S.G.; Delgadillo, I.; Saraiva, J.A. Effect of thermal pasteurisation and high-pressure processing on immunoglobulin content and lysozyme and lactoperoxidase activity in human colostrum. Food Chem. 2014, 151, 79–85. [Google Scholar] [CrossRef]
- Viazis, S.; Farkas, B.E.; Jaykus, L.A. Inactivation of bacterial pathogens in human milk by high-pressure processing. J. Food Prot. 2008, 71, 109–118. [Google Scholar] [CrossRef]
- Peila, C.; Moro, G.E.; Bertino, E.; Cavallarin, L.; Giribaldi, M.; Giuliani, F.; Cresi, F.; Coscia, A. The Effect of Holder Pasteurization on Nutrients and Biologically-Active Components in Donor Human Milk: A Review. Nutrients 2016, 8, 477. [Google Scholar] [CrossRef]
- Pace, R.M.; Williams, J.E.; Järvinen, K.M.; Belfort, M.B.; Pace, C.D.; Lackey, K.A.; Gogel, A.C.; Nguyen-Contant, P.; Kanagaiah, P.; Fitzgerald, T.; et al. COVID-19 and human milk: SARS-CoV-2, antibodies, and neutralizing capacity. MedRxiv 2020. preprint. [Google Scholar]
- Furlow, B. US NICUs and donor milk banks brace for COVID-19. Lancet Child Adolesc. Health 2020, 4, 355. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Confirmed COVID-19 N = 29 | Suspected COVID-19 N = 9 | Controls N = 13 | ||
|---|---|---|---|---|---|
| Gestational age—weeks median (IQR) | 39.7 (38.5, 40.7) | 38.8 (36.8, 40.4) | 40.7 (39.6, 41.1) | ||
| Age of child—weeks median (IQR) | 28.9 (12.1, 39.5) | 12.6 (8.4, 40.7) | 6.1 (4.3, 7.4) | ||
| Age of mother—years mean (SD) | 31.1 (3.1) | 30.3 (4.1) | 33.2 (3.3) | ||
| Time between start of clinical symptoms and collection of human milk—weeks mean (SD) | 5.9 (2.6) | 5.7 (2.1) | NA | ||
| Symptoms and duration in days | N. (%) | Median (IQR) | N. (%) | Median (IQR) | |
| Fever > 37.5 °C | 21 (72%) | 3 (1, 5) | 6 (67%) | 1 (0, 4) | NA |
| Cold | 24 (83%) | 12 (5, 20) | 4 (44%) | 7 (4, 54) | NA |
| Cough | 21 (72%) | 14 (5, 28) | 5 (56%) | 6 (3, 12) | NA |
| Sore throat | 21 (72%) | 6 (4, 14) | 5 (56%) | 6 (3, 12) | NA |
| Tachypnea | 5 (17%) | 11 (4, 14) | 2 (22%) | NA | NA |
| Dyspnea | 14 (38%) | 7 (3, 28) | 1 (11%) | NA | NA |
| Stomachache | 5 (17%) | 2 (1, 9) | 2 (22%) | NA | NA |
| Nausea | 5 (17%) | 3 (2.5, 14) | 1 (11%) | NA | NA |
| Vomiting | 2 (7%) | NA | 0 | NA | NA |
| Diarrhea | 5 (17%) | 2 (1, 25) | 3 (33%) | NA | NA |
| Headache | 24 (83%) | 5 (2, 12) | 8 (89%) | 7 (4, 14) | NA |
| Photophobia | 2 (7%) | NA | 0 | NA | NA |
| Anosmia | 18 (62%) | 20 (13) | 6 (67%) | 9 (4, 22) | NA |
| Ageusia | 17 (59%) | 19 (13) | 4 (44%) | 14 (11, 25) | NA |
| Fatigue | 24 (83%) | 20 (16) | 8 (89%) | 10 (4, 36) | NA |
| Anorexia | 10 (34%) | 12 (8, 21) | 3 (33%) | NA | NA |
| Hospital admission | 3 (10%) | NA | 0 | NA | NA |
| Non-invasive respiratory support (O2) during admission | 2 (7%) | NA | 0 | NA | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Keulen, B.J.; Romijn, M.; Bondt, A.; Dingess, K.A.; Kontopodi, E.; van der Straten, K.; den Boer, M.A.; Burger, J.A.; Poniman, M.; Bosch, B.J.; et al. Human Milk from Previously COVID-19-Infected Mothers: The Effect of Pasteurization on Specific Antibodies and Neutralization Capacity. Nutrients 2021, 13, 1645. https://doi.org/10.3390/nu13051645
van Keulen BJ, Romijn M, Bondt A, Dingess KA, Kontopodi E, van der Straten K, den Boer MA, Burger JA, Poniman M, Bosch BJ, et al. Human Milk from Previously COVID-19-Infected Mothers: The Effect of Pasteurization on Specific Antibodies and Neutralization Capacity. Nutrients. 2021; 13(5):1645. https://doi.org/10.3390/nu13051645
Chicago/Turabian Stylevan Keulen, Britt J., Michelle Romijn, Albert Bondt, Kelly A. Dingess, Eva Kontopodi, Karlijn van der Straten, Maurits A. den Boer, Judith A. Burger, Meliawati Poniman, Berend J. Bosch, and et al. 2021. "Human Milk from Previously COVID-19-Infected Mothers: The Effect of Pasteurization on Specific Antibodies and Neutralization Capacity" Nutrients 13, no. 5: 1645. https://doi.org/10.3390/nu13051645
APA Stylevan Keulen, B. J., Romijn, M., Bondt, A., Dingess, K. A., Kontopodi, E., van der Straten, K., den Boer, M. A., Burger, J. A., Poniman, M., Bosch, B. J., Brouwer, P. J. M., de Groot, C. J. M., Hoek, M., Li, W., Pajkrt, D., Sanders, R. W., Schoonderwoerd, A., Tamara, S., Timmermans, R. A. H., ... van Goudoever, J. B. (2021). Human Milk from Previously COVID-19-Infected Mothers: The Effect of Pasteurization on Specific Antibodies and Neutralization Capacity. Nutrients, 13(5), 1645. https://doi.org/10.3390/nu13051645