
Leuconostoc mesenteroides LVBH107 Antibacterial Activity against Porphyromonas gingivalis and Anti-Inflammatory Activity against P. gingivalis Lipopolysaccharide-Stimulated RAW 264.7 Cells

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Culture Conditions, and Sample Preparation
2.2. Cell Culture Conditions
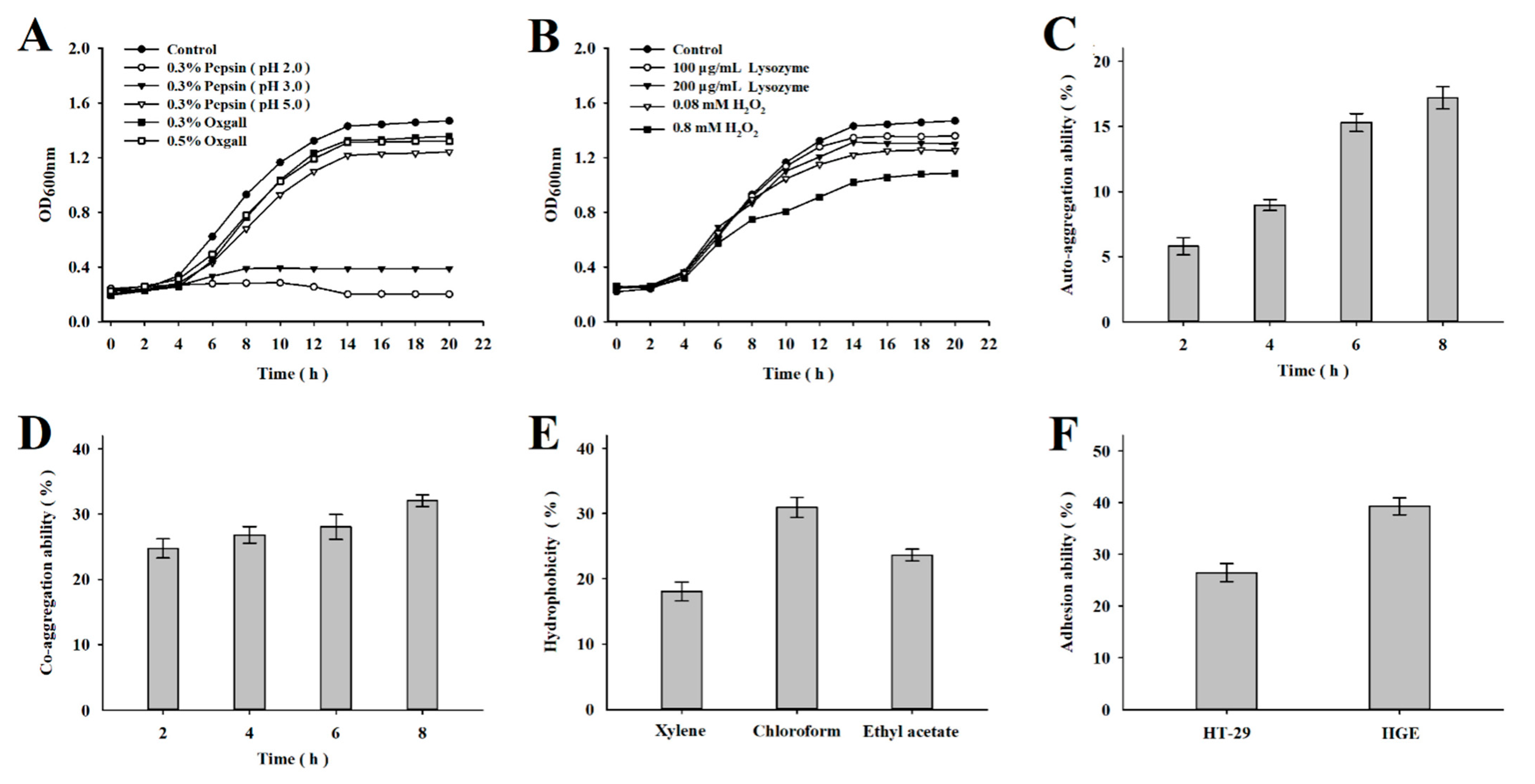
2.3. Probiotic Properties of L. mesenteroides LVBH107
2.3.1. Pepsin, Bile, Lysozyme, and Hydrogen Peroxide Tolerance
2.3.2. Auto-Aggregation Ability
2.3.3. Co-Aggregation Ability
2.3.4. Surface Hydrophobicity
2.3.5. Adhesion Ability
2.3.6. Safety Assessment
2.4. Antibacterial Activity of L. mesenteroides LVBH107 against P. gingivalis
2.4.1. Biofilm Formation Assay
2.4.2. Biofilm Activity Assay
2.5. Assessment of Functional Properties of L. mesenteroides LVBH107 Using the RAW 264.7 Cell Model
2.5.1. Cytotoxicity Assay
2.5.2. Cytokine and Inflammation Assays
2.5.3. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Assay
2.6. Statistical Analysis
3. Results
3.1. Probiotic Characterization of L. mesenteroides LVBH107
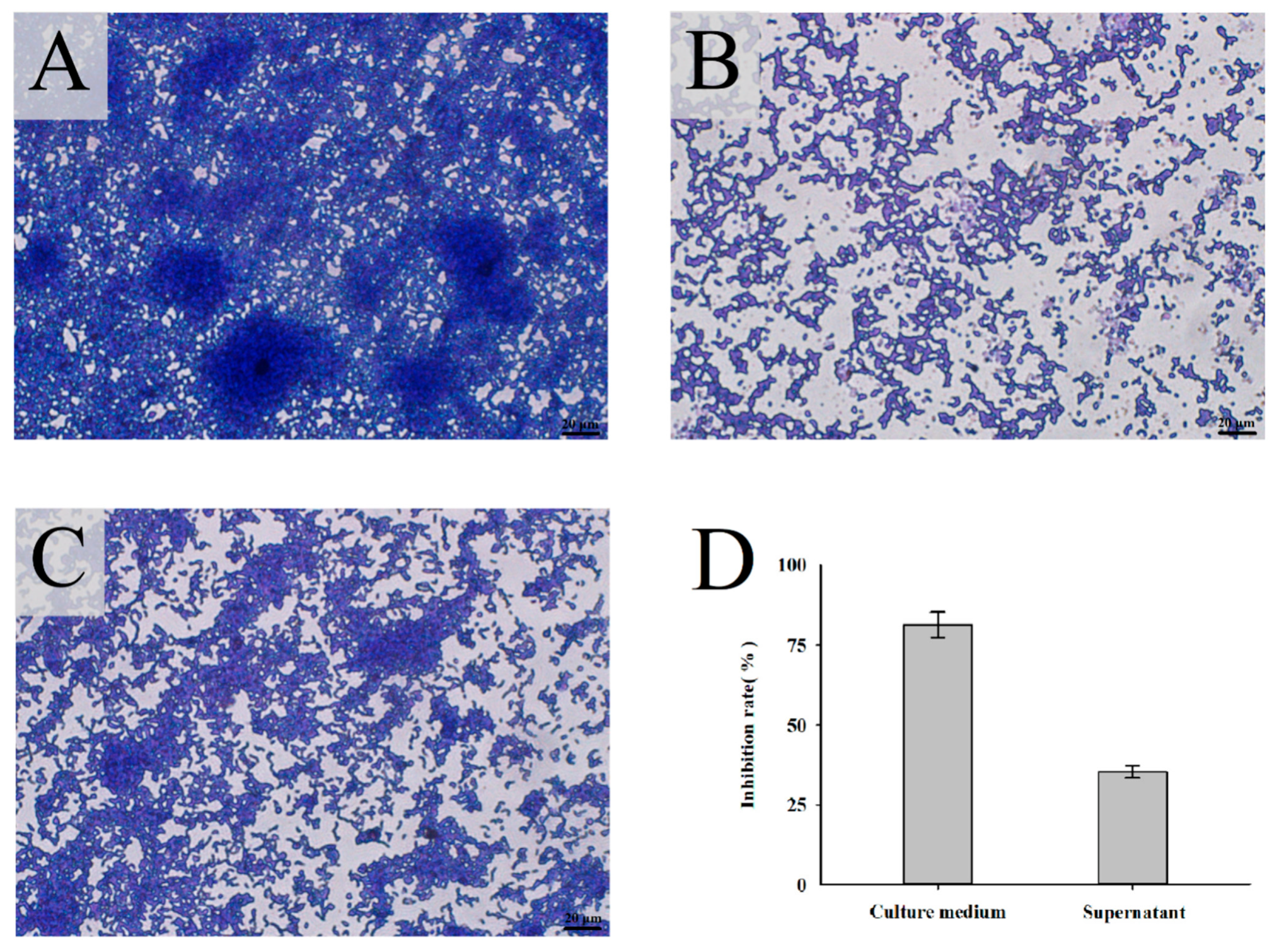
3.2. Inhibitory Effect of L. mesenteroides LVBH107 on P. gingivalis Biofilm Formation
3.2.1. Inhibitory Effect of L. mesenteroides LVBH107 on Biofilm Formation by P. gingivalis
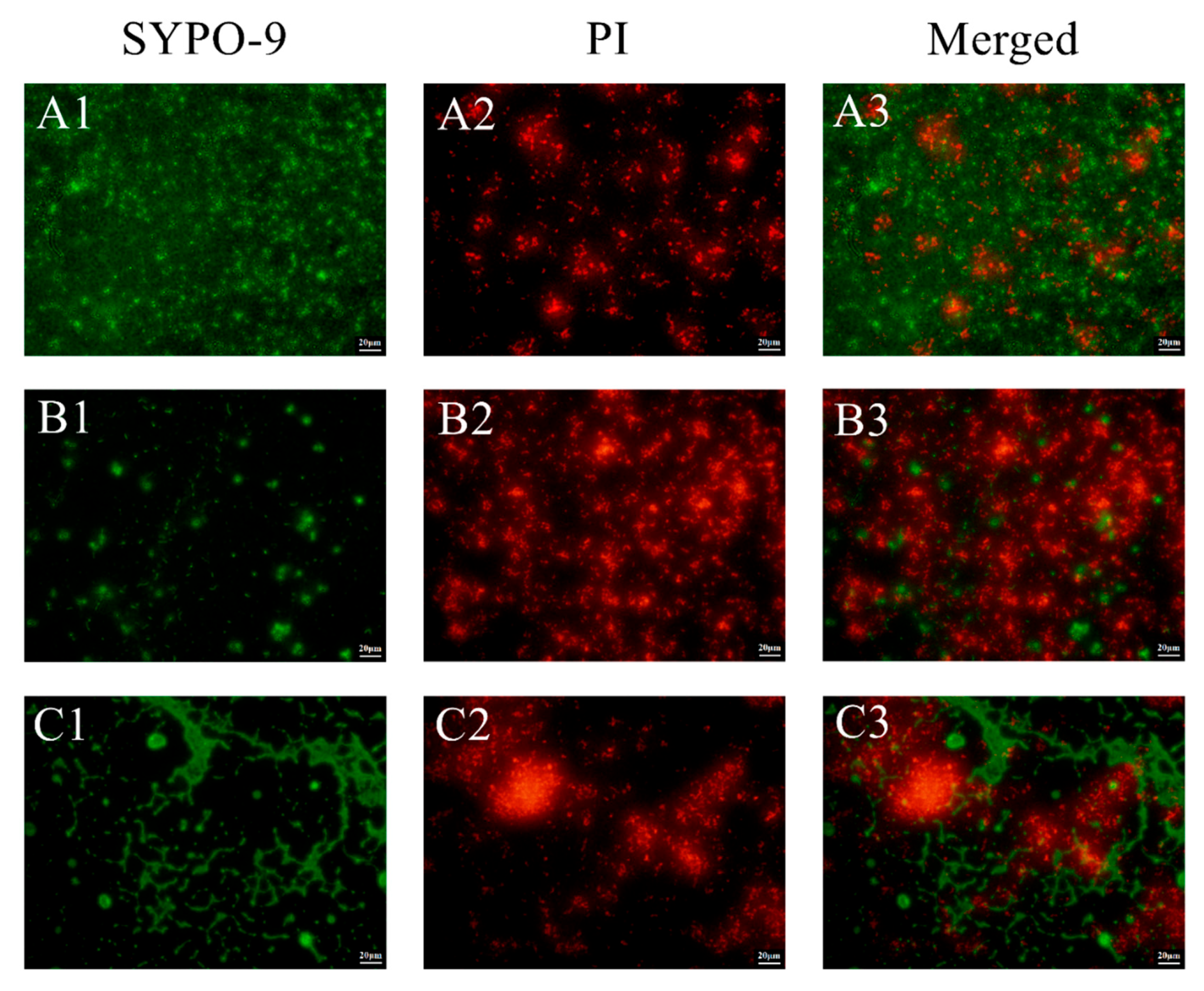
3.2.2. Inhibitory Effect of L. mesenteroides LVBH107 on Biofilm Structure
3.3. In Vitro Anti-Inflammatory Properties of L. mesenteroides LVBH107
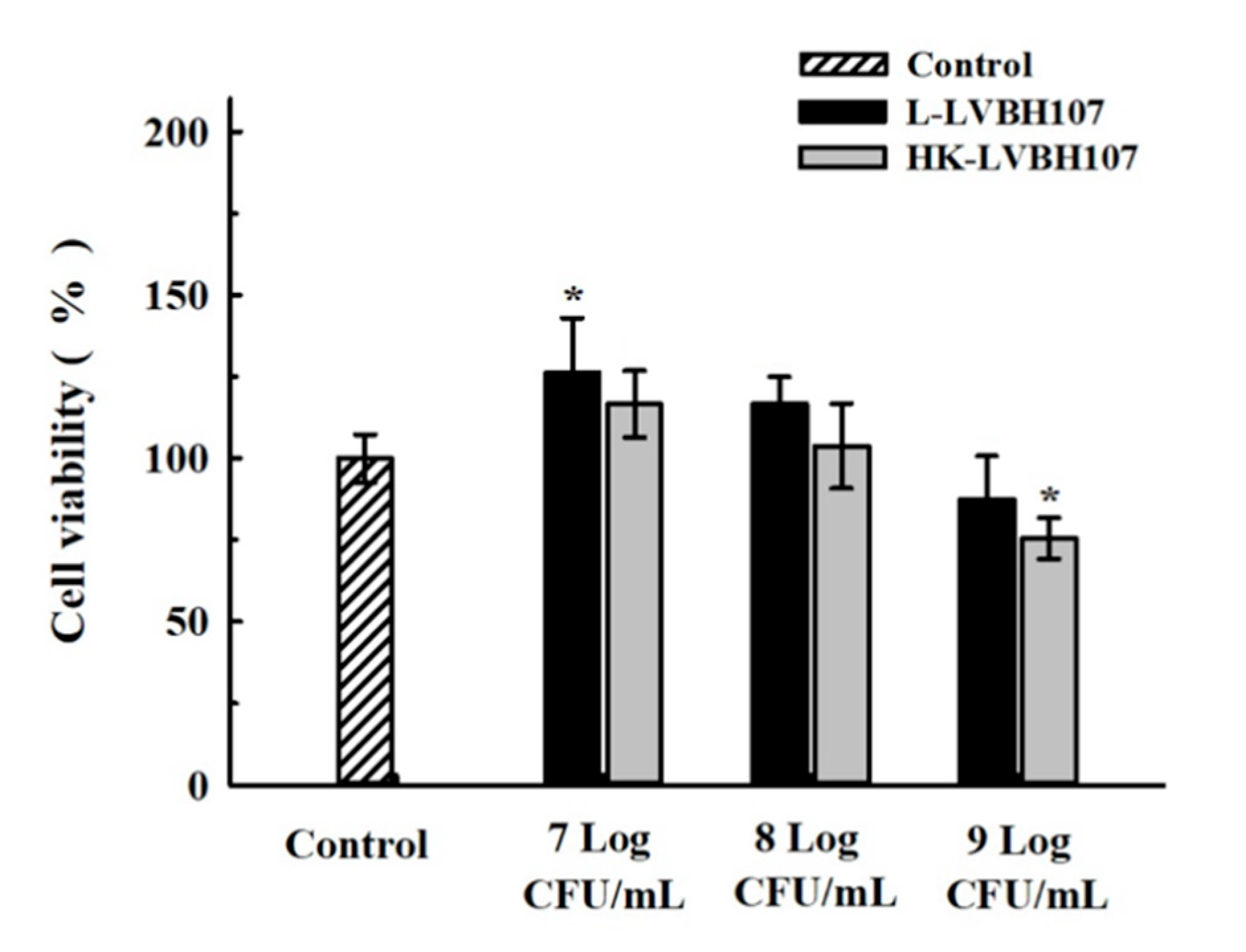
3.3.1. Effect of L. mesenteroides LVBH107 on RAW 264.7 Cells
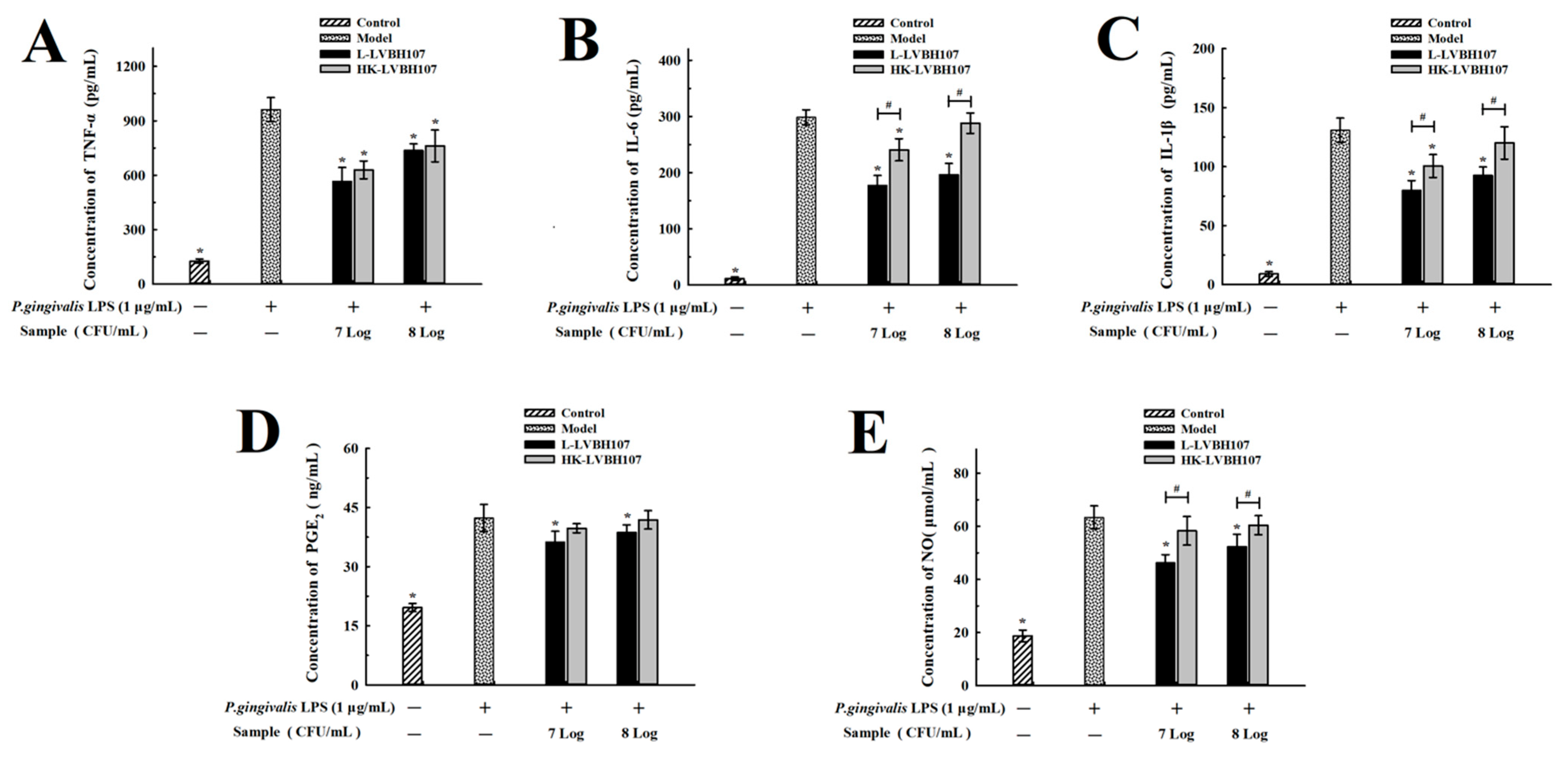
3.3.2. Anti-Inflammatory of L. mesenteroides LVBH107 on Proinflammatory Cytokines Expression in P. gingivalis LPS-Induced RAW 264.7 Cells
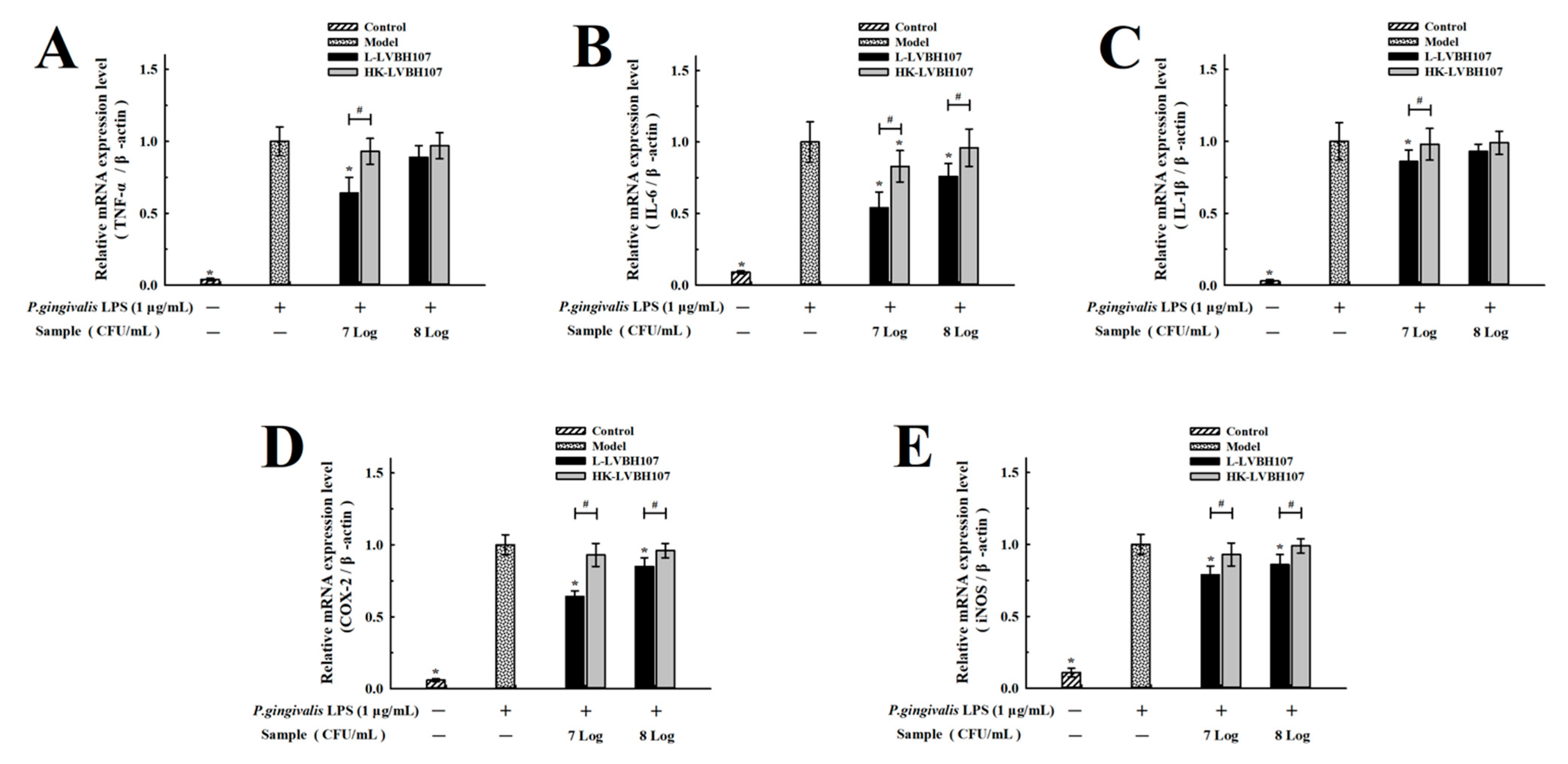
3.3.3. Effect of L. mesenteroides LVBH107 on mRNA Expression Levels in P. gingivalis LPS-Induced RAW 264.7 Cells
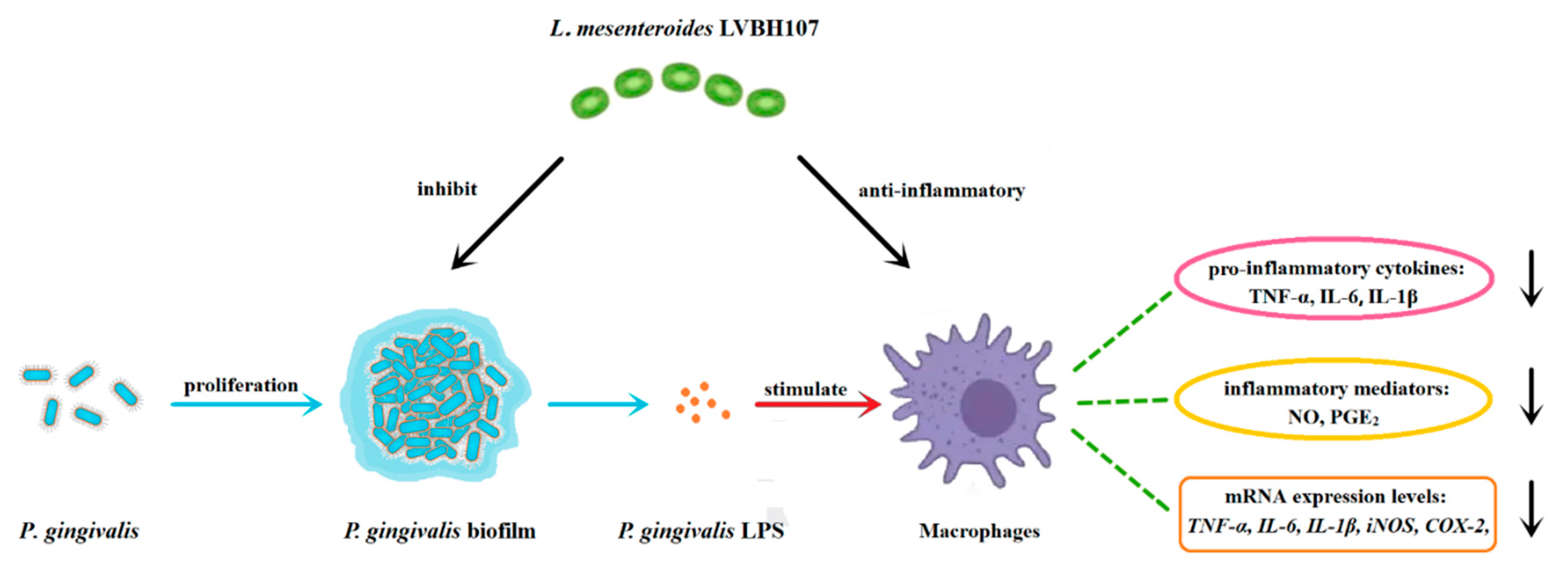
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Morelli, L.; Capurso, L. FAO/WHO guidelines on probiotics 10 years later foreword. Clin. J. Gastroenterol. 2012, 46, S1–S2. [Google Scholar] [CrossRef] [PubMed]
- Boirivant, M.; Strober, W. The mechanism of action of probiotics. Curr. Opin. Gastroenterol. 2007, 23, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Giorgetti, G.M.; Brandimarte, G.; Fabiocchi, F.; Ricci, S.; Flamini, P.; Sandri, G.; Trotta, M.C.; Elisei, W.; Penna, A.; Lecca, P.G.; et al. Interactions between innate immunity, microbiota, and probiotics. J. Immunol. Res. 2015, 2015, 501361. [Google Scholar] [CrossRef]
- Albuquerque-Souza, E.; Balzarini, D.; Ando-Suguimoto, E.S.; Ishikawa, K.H.; Simionato, M.R.L.; Holzhausen, M.; Mayer, M.P.A. Probiotics alter the immune response of gingival epithelial cells challenged by Porphyromonas gingivalis. J. Periodontal Res. 2019, 54, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Nie, S.P.; Zhu, K.X.; Xiong, T.; Li, C.; Gong, J.; Xie, M.Y. Effect of Lactobacillus plantarum NCU116 on loperamide-induced constipation in mice. Int. J. Food Sci. Nutr. 2015, 66, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.; Xiaohong, Y.; Yi, L.; Yun, G.; Lin, G.; Fangfang, P.; Xinying, M.; Wenjing, C.; Francesco, M.; Fang, H. Bifidobacterium bifidum TMC3115 can characteristically influence glucose and lipid profile and intestinal microbiota in the middle-aged and elderly. Probiotics Antimicrob. Proteins 2019, 11, 1182–1194. [Google Scholar]
- Wang, C.; Nagata, S.; Chiba, Y.Y. The effects of the Lactobacillus casei strain on obesity in children: A pilot study. Benef. Microbe 2017, 8, 535–543. [Google Scholar]
- Shaaban, S.Y.; El Gendy, Y.G.; Mehanna, N.S.; El-Senousy, W.M.; El-Feki, H.S.A.; Saad, K.; El-Asheer, O.M. The role of probiotics in children with autism spectrum disorder: A prospective, open-label study. Nutr. Neurosci. 2018, 21, 676–681. [Google Scholar] [CrossRef]
- Gatej, S.; Gully, N.; Gibson, R.; Bartold, P.M. Probiotics and periodontitis—A literature review. J. Int. Acad. Periodontol. 2017, 19, 42–50. [Google Scholar]
- Morales, A.; Carvajal, P.; Silva, N.; Hernandez, M.; Godoy, C.; Rodriguez, G.; Cabello, R.; Garcia-Sesnich, J.; Hoare, A.; Diaz, P.I.; et al. Clinical effects of Lactobacillus rhamnosus in non-surgical treatment of chronic periodontitis: A randomized placebo-controlled trial with 1-year follow-up. J. Periodontol. 2016, 87, 944–952. [Google Scholar] [CrossRef] [Green Version]
- Slots, J. Periodontitis: Facts, fallacies and the future. Periodontology 2017, 75, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Wang, Q.; Chen, Q. The cytokine network involved in the host immune response to periodontitis. Int. J. Oral Sci. 2019, 11, 30–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.; Zhao, L.; Huang, X.; Ma, C.; Wang, Y.; Zhang, J.; Xuan, D. Enhanced activity of the macrophage M1/M2 phenotypes and phenotypic switch to M1 in periodontal infection. J. Periodontol. 2016, 87, 1092–1102. [Google Scholar] [CrossRef] [PubMed]
- Cornacchione, L.P.; Klein, B.A.; Duncan, M.J.; Hu, L.T. Interspecies inhibition of Porphyromonas gingivalis by yogurt-derived Lactobacillus delbrueckii requires active pyruvate oxidase. Appl. Environ. Microbiol. 2019, 85, e01271-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twetman, S.; Derawi, B.; Keller, M.; Ekstrand, K.; Yucel-Lindberg, T.; Stecksén-Blicks, C. Short-term effect of chewing gums containing probiotic Lactobacillus reuteri on the levels of inflammatory mediators in gingival crevicular fluid. Acta Odontol. Scand. 2008, 67, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Riccia, D.N.; Bizzini, F.; Perilli, M.G.; Polimeni, A.; Trinchieri, V.; Amicosante, G.; Cifone, M.G. Anti-inflammatory effects of Lactobacillus brevis (CD2) on periodontal disease. Oral Dis. 2017, 13, 376–385. [Google Scholar] [CrossRef]
- Vicario, M.; Santos, A.; Violant, D.; Nart, J.; Giner, L. Clinical changes in periodontal subjects with the probiotic Lactobacillus reuteri prodentis: A preliminary randomized clinical trial. Acta Odontol. Scand. 2013, 71, 813–819. [Google Scholar] [CrossRef]
- Lauritano, D.; Carinci, F.; Palmieri, A.; Cura, F.; Caruso, S.; Candotto, V. Reuterinos as adjuvant for peri-implant treatment: A pilot study. Int. J. Immunopathol. Pharmacol. 2019, 33, 2058738419827745. [Google Scholar] [CrossRef] [Green Version]
- Bostanci, N.; Belibasakis, G.N. Porphyromonas gingivalis: An invasive and evasive opportunistic oral pathogen. FEMS Microbiol. Lett. 2012, 333, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hajishengallis; George Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [CrossRef]
- Mysak, J.; Podzimek, S.; Sommerova, P.; Lyuya-Mi, Y.; Duskova, J. Porphyromonas gingivalis: Major periodontopathic pathogen overview. J. Immunol. Res. 2014, 3, 476068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Rodríguez, D.C.; Reyes-Castro, L.A.; Vega, C.C.; Rodríguez-González, G.L.; Yáñez-Fernández, J.; Zambrano, E. Leuconostoc mesenteroides subsp. mesenteroides SD23 prevents metabolic dysfunction associated with high-fat diet–induced obesity in male mice. Probiotics Antimicrob. Proteins 2020, 12, 505–516. [Google Scholar] [CrossRef]
- Le, B.; Yang, S.H. Effect of potential probiotic Leuconostoc mesenteroides FB111 in prevention of cholesterol absorption by modulating NPC1l1/PPARα/SREBP-2 pathways in epithelial Caco-2 cells. Int. Microbiol. 2019, 22, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Jjy, A.; Arr, B.; Ajy, B.; Thc, C.; Cmh, B. Production of electricity and reduction of high-fat diet-induced il-6 by glucose fermentation of Leuconostoc mesenteroides. Biochem. Biophys. Res. Commun. 2020, 533, 651–656. [Google Scholar]
- Aslim, B.; Onal, D.; Beyatli, Y. Factors influencing autoaggregation and aggregation of Lactobacillus delbrueckii subsp. bulgaricus isolated from handmade yogurt. J. Food Prot. 2007, 70, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Vlková, E.; Rada, V.; Šmehilová, M.; Killer, J. Auto-aggregation and co-aggregation ability in Bifidobacteria and Clostridia. Folia Microbiol. 2008, 53, 263–269. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Arroyo-López, F.N.; Rantsiou, K.; Jiménez-Díaz, R.; Garrido-Fernández, A.; Cocolin, L. Screening of lactic acid bacteria isolated from fermented table olives with probiotic potential. Food Res. Int. 2013, 50, 135–142. [Google Scholar] [CrossRef]
- Chen, S.; Chen, L.; Chen, L.; Ren, X.; Ge, H.; Li, B.; Ma, G.; Ke, X.; Zhu, J.; Li, L.; et al. Potential probiotic characterization of Lactobacillus reuteri from traditional Chinese highland barley wine and application for room-temperature-storage drinkable yogurt. J. Dairy Sci. 2018, 101, 5780–5788. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.J.; Yu, H.H.; Kim, Y.J.; Lee, N.K.; Paik, H.D. Anti-biofilm activity of grapefruit seed extract against Staphylococcus aureus and Escherichia coli. J. Microbiol. Biotechnol. 2019, 29, 1177–1183. [Google Scholar] [CrossRef]
- Zaidi, S.; Singh, S.L.; Khan, A.U. Exploring antibiofilm potential of bacitracin against Streptococcus mutans. Microb. Pathog. 2020, 149, 104279. [Google Scholar] [CrossRef]
- Yu, H.S.; Lee, N.K.; Choi, A.J.; Choe, J.S.; Bae, C.H.; Paik, H.D. Anti-inflammatory potential of probiotic strain Weissella cibaria JW15 isolated from kimchi through regulation of NF-κB and MAPKs pathways in LPS-induced RAW 264.7 cells. J. Microbiol. Biotechnol. 2019, 29, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Luan, C.; Jiang, N.; Zhou, X.; Zhang, C.; Zhao, Y.; Li, Z.; Li, C. Antibacterial and anti-biofilm activities of probiotic Lactobacillus curvatus BSF206 and Pediococcus pentosaceus AC1-2 against Streptococcus mutans. Microb. Pathog. 2022, 164, 105446. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W. Regression analysis for continuous independent variables in medical research: Statistical standard and guideline of life cycle committee. Life Cycle 2022, 2, 2799–8894.e3. [Google Scholar] [CrossRef]
- Angmo, K.; Kumari, A.; Bhalla, T.C. Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh. LWT Food. Sci. Technol. 2016, 66, 428–435. [Google Scholar] [CrossRef]
- Lee, C.S.; Kim, S.H. Anti-inflammatory and anti-osteoporotic potential of Lactobacillus plantarum A41 and L. fermentum SRK414 as probiotics. Probiotics Antimicrob. Proteins 2019, 12, 623–634. [Google Scholar] [CrossRef]
- Maldonado, N.C.; de Ruiz, C.S.; Otero, M.C.; Sesma, F.; Nader-Macías, M.E. Lactic acid bacteria isolated from young calves—Characterization and potential as probiotics. Res. Vet. Sci. 2012, 92, 342–349. [Google Scholar] [CrossRef]
- Roos, S.; Jonsson, H. A high-molecular-mass cell-surface protein from Lactobacillus reuteri 1063 adheres to mucus components. Microbiology 2002, 148, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Dlamini, Z.C.; Langa, R.L.S.; Aiyegoro, O.A.; Okoh, A.I. Safety evaluation and colonisation abilities of four lactic acid bacteria as future probiotics. Probiotics Antimicrob. Proteins 2019, 11, 397–402. [Google Scholar] [CrossRef]
- Juntunen, M.; Kirjavainen, P.V.; Ouwehand, A.C.; Salminen, S.J.; Isolauri, E. Adherence of probiotic bacteria to human intestinal mucus in healthy infants and during rotavirus infection. Clin. Diagn. Lab. Immunol. 2001, 8, 293–296. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.M.; Lee, N.K.; Paik, H.D. Antibacterial and anticavity activity of probiotic Lactobacillus plantarum 200661 isolated from fermented foods against Streptococcus mutans. LWT 2020, 118, 108840. [Google Scholar] [CrossRef]
- Kakisu, E.; Bolla, P.; Abraham, A.G.; Urraza, P.D.; Antoni, G. Lactobacillus plantarum isolated from Kefir: Protection of cultured Hep-2 cells against shigella invasion. Int. Dairy J. 2013, 33, 22–26. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, Y.; Tian, F.; Jin, X.; Chen, H.; Liu, X.; Zhang, Q.; Zhao, J.; Chen, Y.; Zhang, H.; et al. Screening of adhesive Lactobacilli with antagonistic activity against Campylobacter jejuni. Food Control 2014, 44, 49–57. [Google Scholar] [CrossRef]
- Vasiljevic, T.; Shah, N.P. Probiotics—From Metchnikoff to bioactives. Int. Dairy J. 2008, 18, 714–728. [Google Scholar] [CrossRef]
- Gon Alves, D.; Ferrari, Í.D.S.; Souza, J.D.; Silva, C.; Costa, M.M.D.; Dias, F.S. Characterization and evaluation of lactic acid bacteria isolated from goat milk. Food Control 2015, 53, 96–103. [Google Scholar]
- Essid, I.; Medini, M.; Hassouna, M. Technological and safety properties of Lactobacillus plantarum strains isolated from a Tunisian traditional salted meat. Meat Sci. 2009, 81, 203–208. [Google Scholar] [CrossRef]
- Tulumoglu, S.; Yuksekdag, Z.N.; Beyatli, Y.; Simsek, O.; Cinar, B.; Ya?Ar, E. Probiotic properties of Lactobacilli species isolated from children’s feces. Anaerobe 2013, 24, 36–42. [Google Scholar] [CrossRef]
- Mathara, J.M.; Schillinger, U.; Kutima, P.M.; Mbugua, S.K.; Guigas, C.; Franz, C.; Holzapfel, W.H. Functional properties of Lactobacillus plantarum strains isolated from maasai traditional fermented milk products in Kenya. Curr. Microbiol. 2008, 56, 315–321. [Google Scholar] [CrossRef]
- Nagano, K.; Hasegawa, Y.; Iijima, Y.; Kikuchi, T.; Mitani, A. Distribution of Porphyromonas gingivalis fimA and mfa1 fimbrial genotypes in subgingival plaques. PeerJ 2018, 6, e5581. [Google Scholar] [CrossRef] [Green Version]
- Terai, T.; Okumura, T.; Imai, S.; Nakao, M.; Yamaji, K.; Ito, M.; Nagata, T.; Kaneko, K.; Miyazaki, K.; Okada, A. Screening of probiotic candidates in human oral bacteria for the prevention of dental disease. PLoS ONE 2015, 10, e0128657. [Google Scholar] [CrossRef]
- Kang, M.S.; Oh, J.S.; Lee, H.C.; Lim, H.S.; Lee, S.W.; Yang, K.H.; Choi, N.K.; Kim, S.M. Inhibitory effect of Lactobacillus reuteri on periodontopathic and cariogenic bacteria. J. Microbiol. 2011, 49, 193–199. [Google Scholar] [CrossRef]
- Invernici, M.M.; Salvador, S.L.; Silva, P.H.F.; Soares, M.S.M.; Casarin, R.; Palioto, D.B.; Souza, S.L.S.; Taba, M.; Novaes, A.B.; Furlaneto, F.A.C.; et al. Effects of Bifidobacterium probiotic on the treatment of chronic periodontitis: A randomized clinical trial. J. Clin. Periodontol. 2018, 45, 1198–1210. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Wyant, T.; Anaya-Bergman, C.; Aduse-Opoku, J.; Brunner, J.; Laine, M.L.; Curtis, M.A.; Lewis, J.P. The capsule of Porphyromonas gingivalis leads to a reduction in the host inflammatory response, evasion of phagocytosis, and increase in virulence. Infect. Immun. 2011, 79, 4533–4542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zagozdzon, R.; Foroncewicz, B.; Paczek, L. The aging of the immune system. Przegląd Lek. 2003, 60, 156–160. [Google Scholar]
- Murray, P.J. Macrophage polarization. Annu. Rev. Physiol. 2016, 79, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, A.S.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Zhou, Y.Q.; Wang, M.H. Activation of the RON receptor tyrosine kinase protects murine macrophages from apoptotic death induced by bacterial lipopolysaccharide. J. Leukoc. Biol. 2002, 71, 359–366. [Google Scholar]
- Huang, H.; Al-Shabrawey, M.; Wang, M.H. Cyclooxygenase- and cytochrome P450-derived eicosanoids in stroke. Prostaglandins Other Lipid Mediat. 2016, 122, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Tanigawa, T.; Odkhuu, E.; Morikawa, A.; Hayashi, K.; Sato, T.; Shibata, R.; Goto, F.; Ueda, H.; Yokochi, T. Immunological role of prostaglandin E2 production in mouse auditory cells in response to LPS. Innate Immun. 2014, 20, 639–646. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.W.; Ong, W.K.; Su, Y.W.; Hsu, C.C.; Cheng, T.H.; Tsai, Y.C. Anti-inflammatory effects of Lactobacillus brevis K65 on RAW 264.7 cells and in mice with dextran sulphate sodium-induced ulcerative colitis. Benef. Microbes 2016, 7, 387–396. [Google Scholar] [CrossRef]
- Dinarello, C.A. Interleukin-1 and interleukin-1 antagonism. Blood 1991, 77, 1627–1652. [Google Scholar] [CrossRef] [Green Version]
- Heo, S.J.; Yoon, W.J.; Kim, K.N.; Ahn, G.N.; Kang, S.M.; Kang, D.H.; Ff An, A.A.; Oh, C.; Jung, W.K.; Jeon, Y.J. Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.L.; Pan, J.S.; Lu, Y.P.; Sun, P.; Han, J. Inflammatory signaling and cellular senescence. Cell. Signal. 2008, 21, 378–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghadimi, D.; Nielsen, A.; Hassan, M.F.Y.; Folster-Holst, R.; de Vrese, M.; Heller, K.J. Modulation of GSK-3β/β-catenin cascade by commensal Bifidobateria plays an important role for the inhibition of metaflammation-related biomarkers in response to LPS or non-physiological concentrations of fructose: An in vitro study—Sciencedirect. Pharma Nutr. Sci. Verse Sci. 2019, 8, 100145. [Google Scholar]
- Xiao, H.; Sun, K.-Y.; Yang, X.F.; Tang, J.; Plummer, S.X. Lactobacillus paracasei modulates LPS-induced inflammatory cytokine release by monocyte-macrophages via the up-regulation of negative regulators of NF-kappaB signaling in a TLR2-dependent manner. Cytokine 2017, 92, 1–11. [Google Scholar]
- Yamashita, M.; Ukibe, K.; Uenishi, H.; Hosoya, T.; Sakai, F. Lactobacillus helveticus SBT2171, a cheese starter, regulates proliferation and cytokine production of immune cells. J. Dairy Sci. 2014, 97, 4772–4779. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, C.; Hayakawa, A.; Matsumoto, K.; Katoh, T.; Yamamoto, K.; Hisa, K. Exopolysaccharides produced by Leuconostoc mesenteroides strain NTM048 as an immunostimulant to enhance the mucosal barrier and influence the systemic immune response. J. Agric. Food Chem. 2015, 63, 7009–7015. [Google Scholar] [CrossRef]
- Kook, S.Y.; Lee, Y.; Jeong, E.C.; Kim, S. Immunomodulatory effects of exopolysaccharides produced by Bacillus licheniformis and Leuconostoc mesenteroides isolated from Korean kimchi. J. Funct. Foods 2019, 54, 211–219. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) |
|---|---|
| β-actin | Forward: GTGGGCCGCCCTAGGCACCAG |
| Reverse: GGAGGAAGAGGATGCGGCAGT | |
| TNF-α | Forward: TTGACCTCAGCGCTGAGTTG |
| Reverse: CCTGTAGCCCACGTCTAGC | |
| IL-6 | Forward: GTACTCCAGAAGACCAGAGG |
| Reverse: TGCTGGTGACAACCACGGCC | |
| IL-1β | Forward: CAGGATGAGGACATGAGCACC |
| Reverse: CTCTGCAGACTCAAACTCCAC | |
| COX-2 | Forward: CACTACATCCTGACCCACTT |
| Reverse: ATGCTCCTGCTTGAGTATGT | |
| iNOS | Forward: CCCTTCCGAAGTTTCTGGCAGCAGC |
| Reverse: GGCTGTCAGAGCCTCGTGGCTTTGG |
| Antimicrobial Agent | Disk Content (μg) | Criteria of Inhibition Zone Diameters (mm) | Detection Result | ||||
|---|---|---|---|---|---|---|---|
| Group | Drug | R | I | S | Inhibition Zone (mm) | Sensibility | |
| macrolides | Erythromycin | 15 | ≤15 | 16–20 | ≥21 | 28.2 ± 2.3 | S |
| Tetracyclines | Doxycycline | 30 | ≤12 | 13–15 | ≥16 | 19.9 ± 1.4 | S |
| Tetracycline | 30 | ≤11 | 12–14 | ≥15 | 14.6 ± 0.6 | I | |
| Minocyline | 30 | ≤12 | 13–15 | ≥16 | 22.3 ± 2.4 | S | |
| β-Lactams Penicillins | Penicillin | 10 | ≤18 | - | ≥29 | 24.5 ± 1.7 | R |
| Piperacillin | 100 | ≤17 | 18–20 | ≥21 | 19.4 ± 0.9 | I | |
| Ampicillin | 10 | ≤11 | 12–14 | ≥15 | 15.4 ± 0.7 | S | |
| Aminoglycosides | Gentamicin | 10 | ≤12 | 13–14 | ≥15 | 0 | R |
| Streptomycin | 10 | ≤12 | 13–14 | ≥15 | 0 | R | |
| Lipopeptides | Polymyxin B | 300 | ≤8 | 9–11 | ≥12 | 6.2 ± 1.1 | R |
| Glycopeptides | Vancomycin | 30 | ≤14 | - | ≥15 | 0 | R |
| Chloramphenicol | Chloramphenicol | 30 | ≤13 | 14–17 | ≥18 | 10.8 ± 1.0 | R |
| ansamycins | Rifampicin | 5 | ≤16 | 17–19 | ≥20 | 13.7 ± 0.8 | R |
| Cephems | Cefazolin | 30 | ≤19 | 20–22 | ≥23 | 10.1 ± 0.6 | R |
| Cefuroxime | 30 | ≤14 | 15–22 | ≥23 | 21.6 ± 1.5 | S | |
| Cefoperazone | 75 | ≤15 | 16–20 | ≥21 | 19.5 | I | |
| Ceftazidime | 30 | ≤17 | 18–20 | ≥21 | 0 | R | |
| Ceftriaxone | 30 | ≤19 | 20–22 | ≥23 | 21.0 | I | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luan, C.; Yan, J.; Jiang, N.; Zhang, C.; Geng, X.; Li, Z.; Li, C. Leuconostoc mesenteroides LVBH107 Antibacterial Activity against Porphyromonas gingivalis and Anti-Inflammatory Activity against P. gingivalis Lipopolysaccharide-Stimulated RAW 264.7 Cells. Nutrients 2022, 14, 2584. https://doi.org/10.3390/nu14132584
Luan C, Yan J, Jiang N, Zhang C, Geng X, Li Z, Li C. Leuconostoc mesenteroides LVBH107 Antibacterial Activity against Porphyromonas gingivalis and Anti-Inflammatory Activity against P. gingivalis Lipopolysaccharide-Stimulated RAW 264.7 Cells. Nutrients. 2022; 14(13):2584. https://doi.org/10.3390/nu14132584
Chicago/Turabian StyleLuan, Chang, Jiaqing Yan, Ning Jiang, Chuang Zhang, Xu Geng, Zhengqiang Li, and Chen Li. 2022. "Leuconostoc mesenteroides LVBH107 Antibacterial Activity against Porphyromonas gingivalis and Anti-Inflammatory Activity against P. gingivalis Lipopolysaccharide-Stimulated RAW 264.7 Cells" Nutrients 14, no. 13: 2584. https://doi.org/10.3390/nu14132584
APA StyleLuan, C., Yan, J., Jiang, N., Zhang, C., Geng, X., Li, Z., & Li, C. (2022). Leuconostoc mesenteroides LVBH107 Antibacterial Activity against Porphyromonas gingivalis and Anti-Inflammatory Activity against P. gingivalis Lipopolysaccharide-Stimulated RAW 264.7 Cells. Nutrients, 14(13), 2584. https://doi.org/10.3390/nu14132584