
Antioxidant Compounds from Edible Mushrooms as Potential Candidates for Treating Age-Related Neurodegenerative Diseases
 , ,
, ,
Abstract
:1. Introduction
2. Metodology
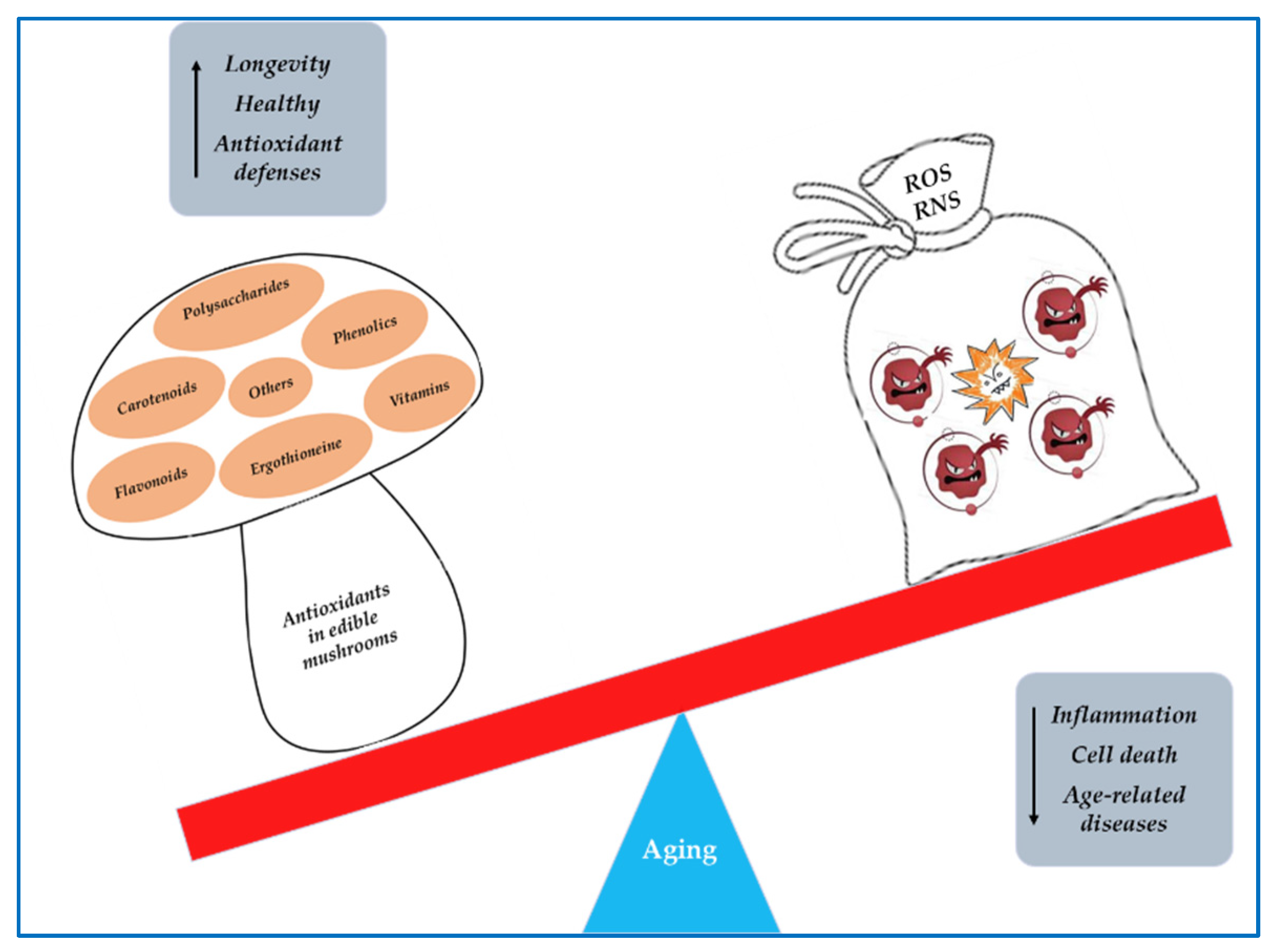
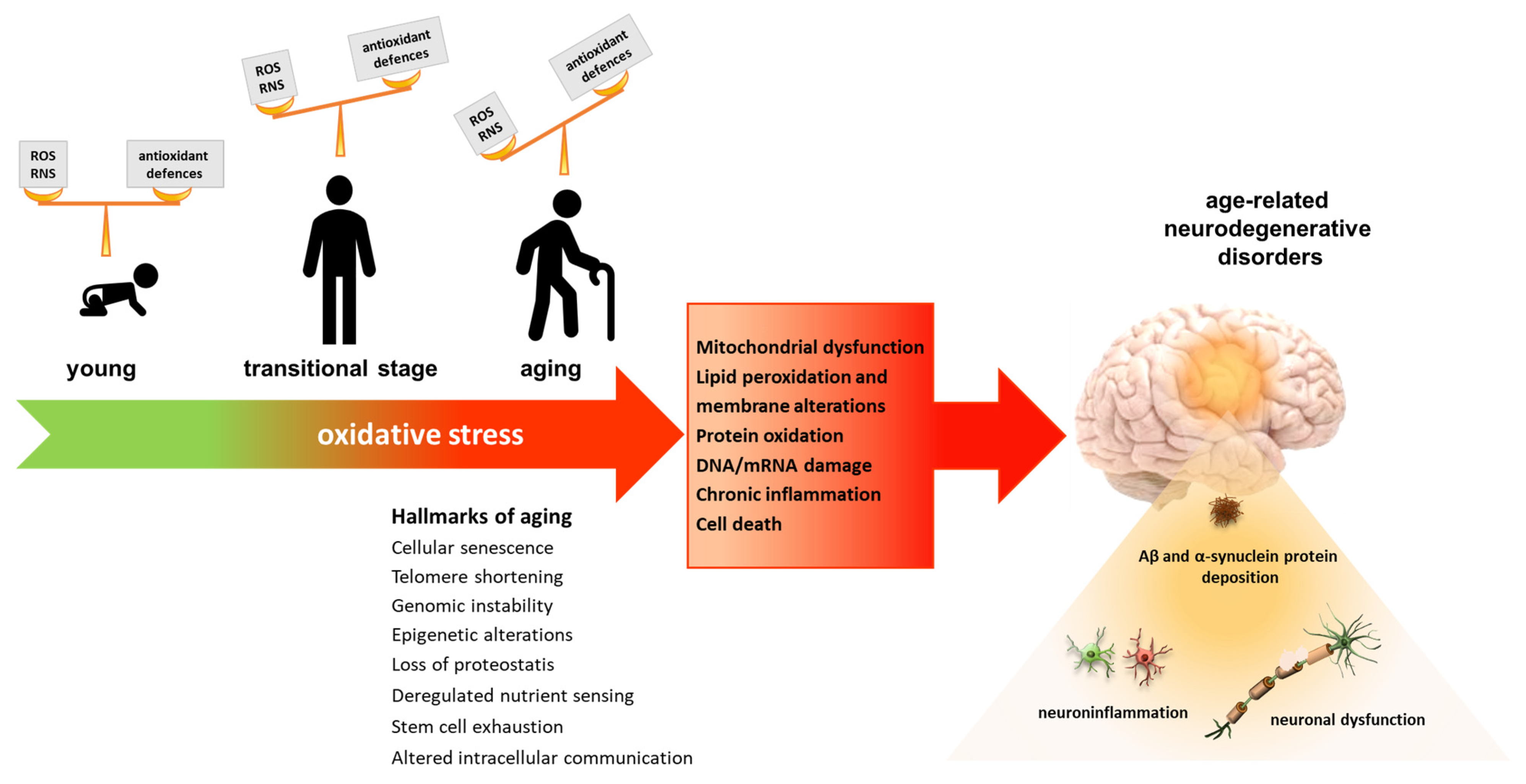
3. Oxidative Stress
4. Role of Oxidative Stress in Aging and Age-Related Neurodegenerative Diseases
4.1. Alzheimer’s Disease
4.2. Parkinson’s Disease
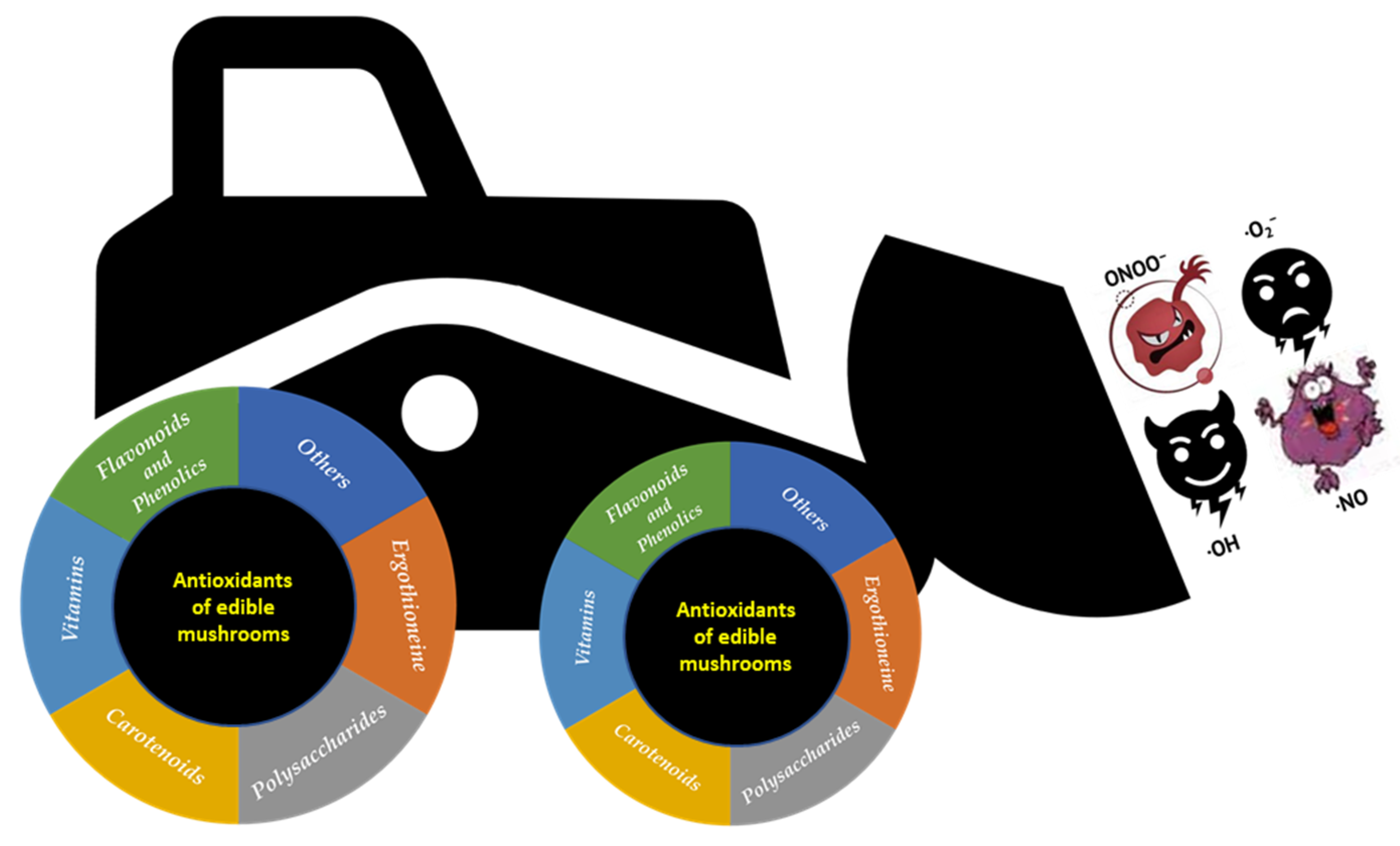
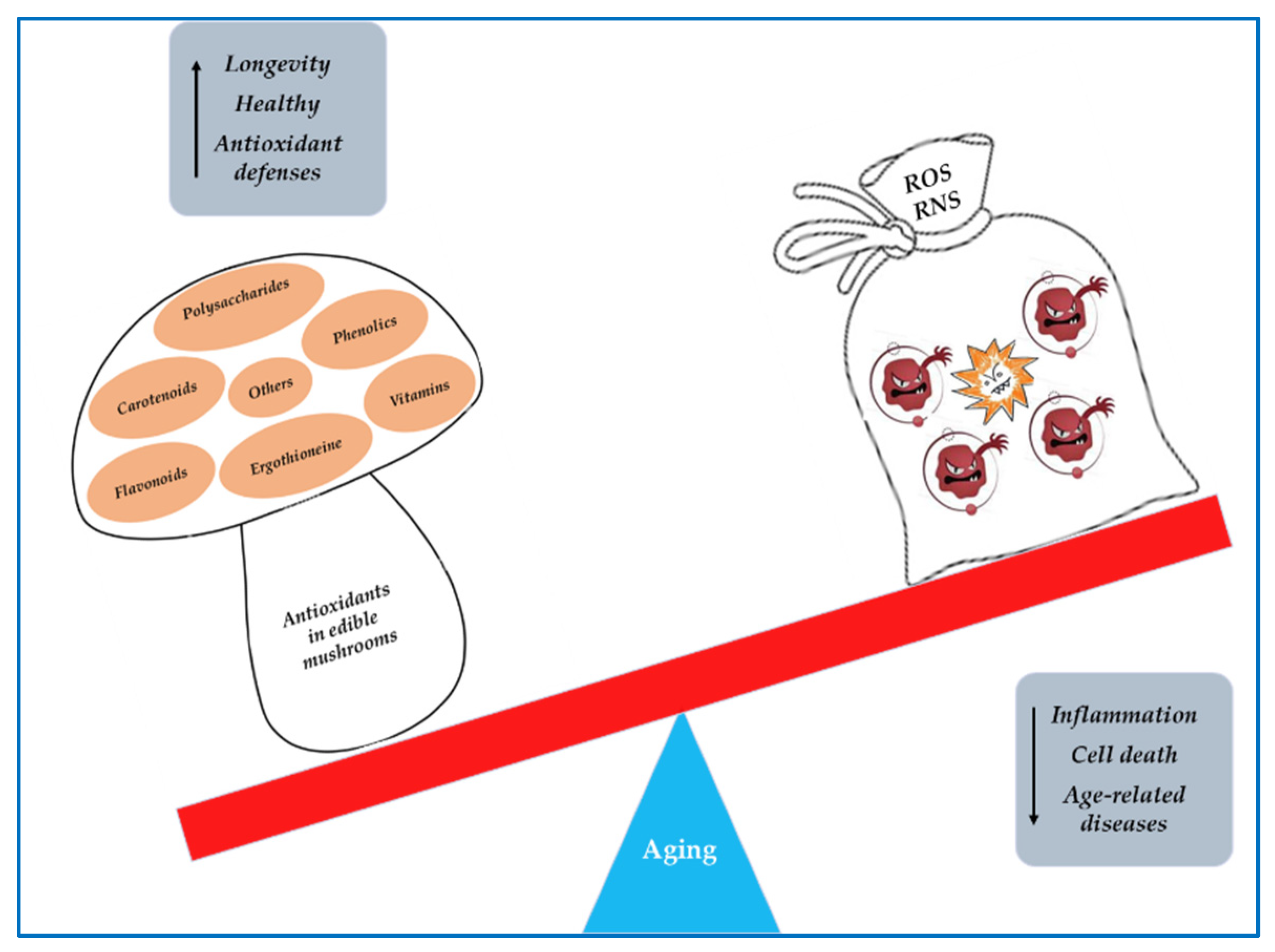
5. Mushrooms as Sources of Antioxidant Compounds
5.1. Polyphenols
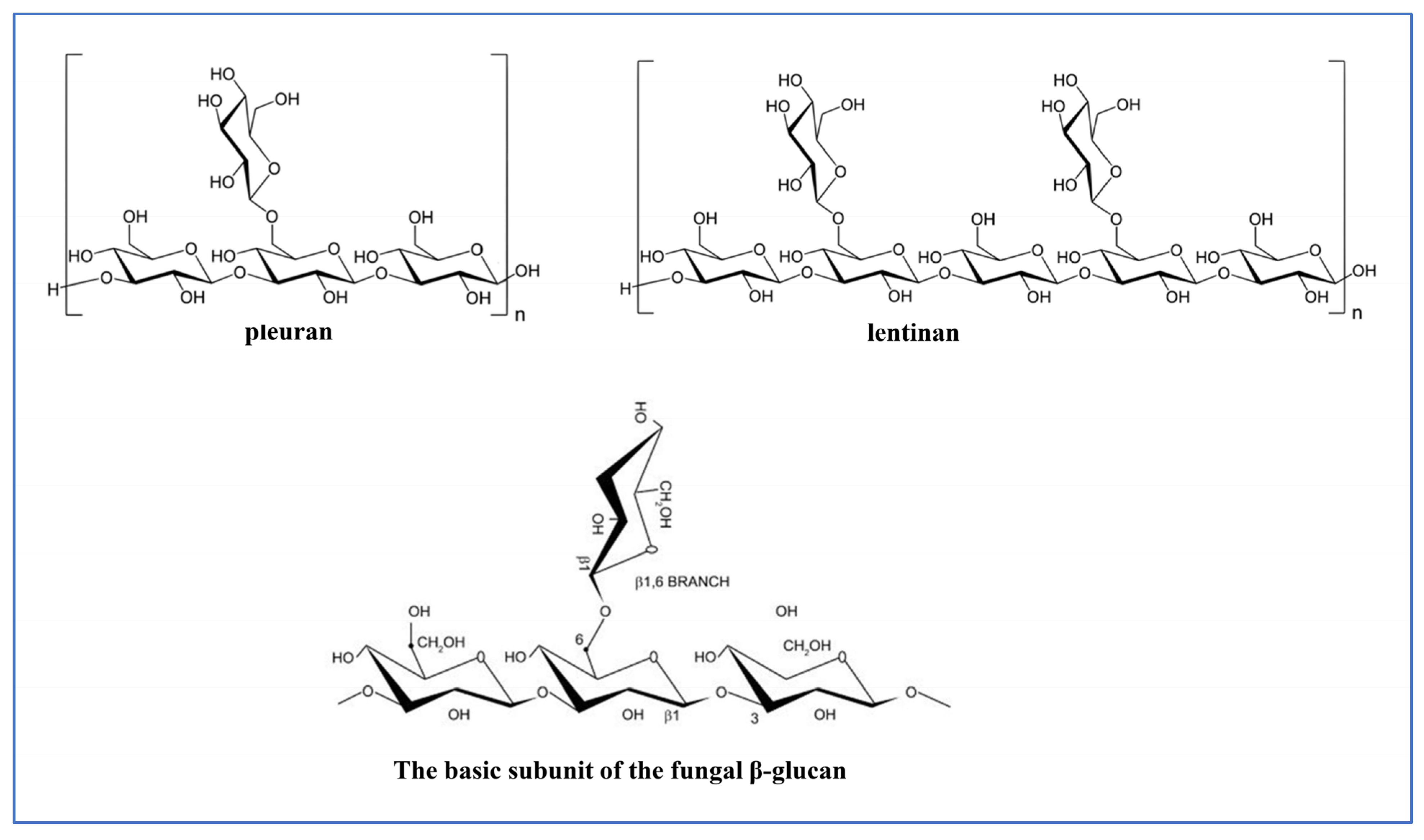
5.2. Polysaccharides
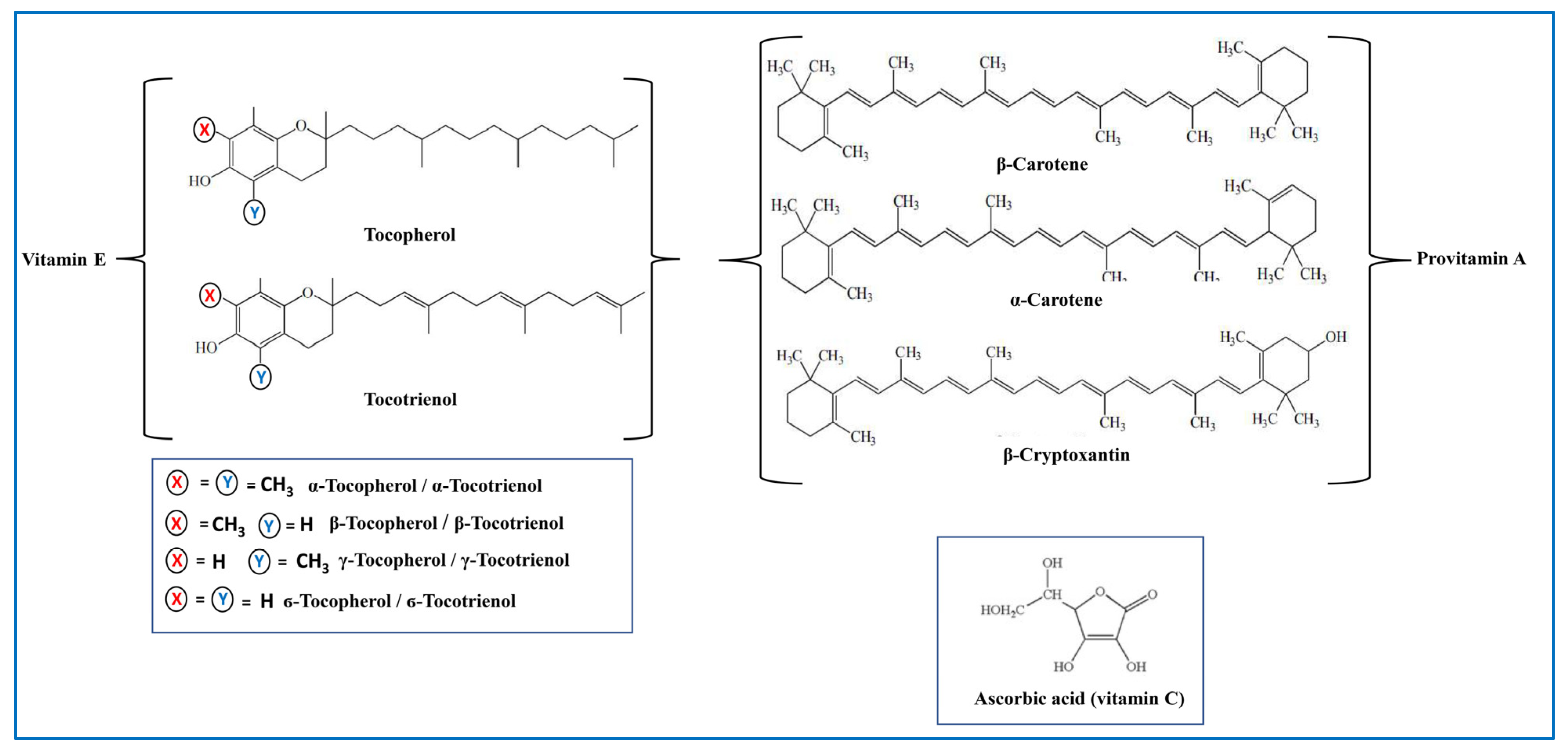
5.3. Antioxidant Vitamins
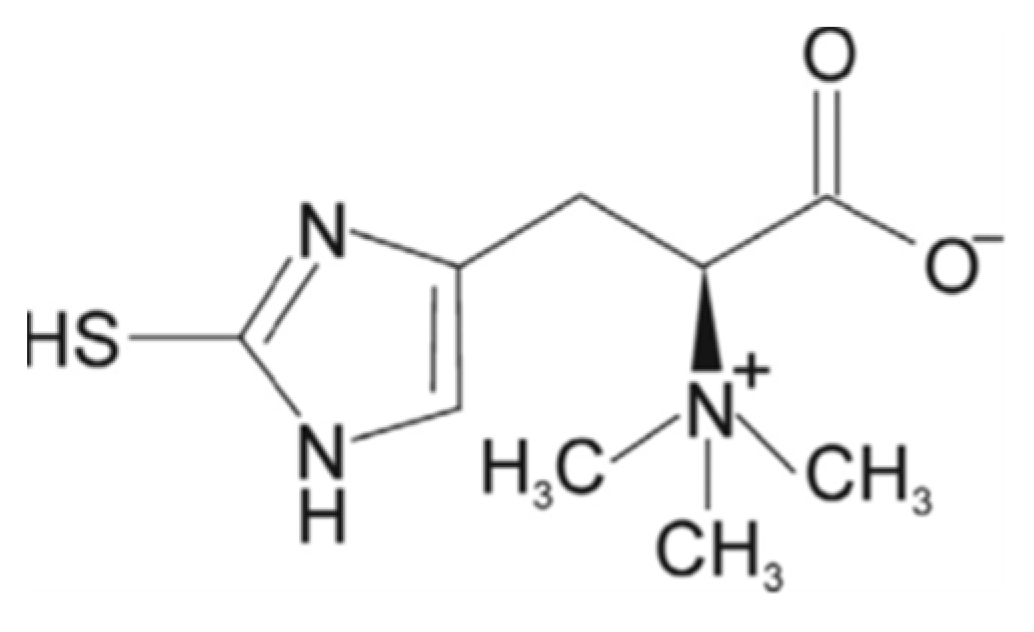
5.4. Ergothioneine
5.5. Other Antioxidants
5.5.1. Minerals
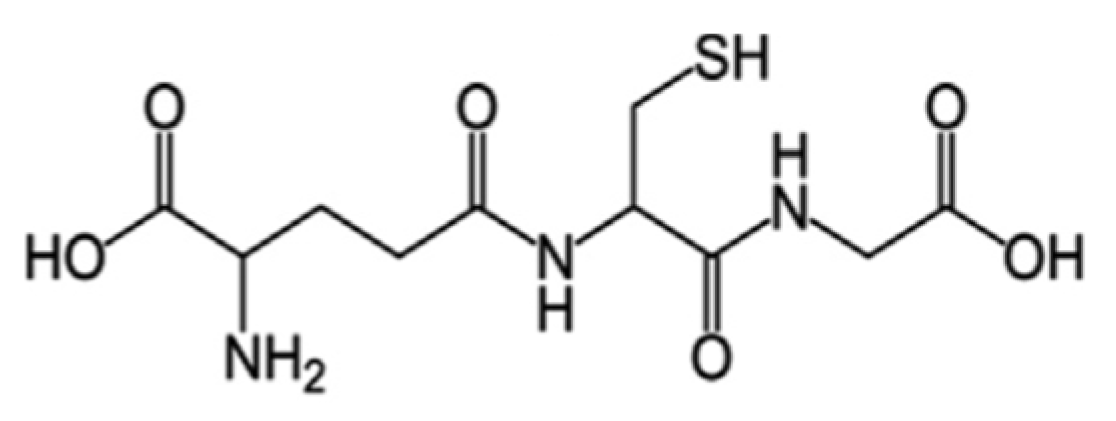
5.5.2. Glutathione (GSH)
6. Antioxidant Compounds of Mushrooms as Neuroprotective Agents
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Z.; Zhang, Z.; Ren, Y.; Wang, Y.; Fang, J.; Yue, H.; Ma, S.; Guan, F. Aging and age-related diseases: From mechanisms to therapeutic strategies. Biogerontology 2021, 22, 165–187. [Google Scholar] [CrossRef] [PubMed]
- de Magalhales, J.P.; Stevens, M.; Thornton, D. The business of anti-aging. Sci. Trends Biotechnol. 2017, 35, 1062–1073. [Google Scholar] [CrossRef] [Green Version]
- Abramov, A.Y.; Potapova, E.V.; Dremin, V.V.; Dunaev, A.V. Interaction of Oxidative Stress and Misfolded Proteins in the Mechanism of Neurodegeneration. Life 2020, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Angelova, P.R.; Abramov, A.Y. Alpha-synuclein and beta-amyloid—Different targets, same players: Calcium, free radicals and mitochondria in the mechanism of neurodegeneration. Biochem. Biophys. Res. Commun. 2017, 483, 1110–1115. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The role of oxidative stress in neurodegenerative diseases. Exp. Neurobiol. 2015, 24, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korovesis, D.; Rubio-Tomás, T.; Tavernarakis, N. Oxidative stress in age-related neurodegenerative diseases: An overview of recent tools and findings. Antioxidants 2023, 12, 131. [Google Scholar] [CrossRef]
- Cobley, J.N.; Fiorello, M.L.; Bailey, D.M. 13 reasons why the brain is susceptible to oxidative stress. Redox Biol. 2018, 15, 490–503. [Google Scholar] [CrossRef]
- Fekete, M.; Szarvas, Z.; Fazekas-Pongor, V.; Feher, A.; Csipo, T.; Forrai, J.; Dosa, N.; Peterfi, A.; Lehoczki, A.; Tarantini, S.; et al. Nutrition strategies promoting healthy aging: From improvement of cardiovascular and brain health to prevention of age-associated diseases. Nutrients 2022, 15, 47. [Google Scholar] [CrossRef]
- Patel, Y.; Naraian, R.; Singh, V.K. Medicinal properties of Pleurotus Species (Oyster Mushroom): A review. World J. Fungal Plant Biol. 2012, 3, 1–12. [Google Scholar]
- Giavasis, I. Bioactive fungal polysaccharides as potential functional ingredients in food and nutraceuticals. Curr. Opin. Biotechnol. 2014, 26, 162–173. [Google Scholar] [CrossRef]
- Kozarski, M.; Klaus, A.; Jakovljevic, D.; Todorovic, N.; Vunduk, J.; Petrović, P.; Niksic, M.; Vrvic, M.M.; van Griensven, L. Antioxidants of edible mushrooms. Molecules 2015, 20, 19489–19525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nohl, H.; Gille, L.; Staniek, K. Intracellular generation of reactive oxygen species by mitochondria. Biochem. Pharmacol. 2005, 69, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Magnani, F.; Mattevi, A. Structure and mechanisms of ROS generation by NADPH oxidases. Curr. Opin. Struct. Biol. 2019, 59, 91–97. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxid. Med. Cell Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The chemistry of reactive oxygen species (ROS) revisited: Outlining their role in biological macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals and antioxidants: Updating a personal view. Nutr. Rev. 2012, 70, 257–265. [Google Scholar] [CrossRef]
- Morató, L.; Bertini, E.; Verrigni, D.; Ardissone, A.; Ruiz, M.; Ferrer, I.; Uziel, G.; Pujol, A. Mitochondrial dysfunction in central nervous system white matter disorders. Glia 2014, 62, 1878–1894. [Google Scholar] [CrossRef]
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M.P. Pathways towards and away from Alzheimer’s disease. Nature 2004, 430, 631–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leng, F.; Edison, P. Neuroinflammation and microglial activation in Alzheimer’s disease: Where do we go from here? Nat. Rev. Neurol. 2021, 17, 157–172. [Google Scholar] [CrossRef]
- Nunomura, A.; Castellani, R.J.; Zhu, X.; Moreira, P.I.; Perry, G.; Smith, M.A. Involvement of oxidative stress in Alzheimer disease. J. Neuropathol. Exp. Neurol. 2006, 65, 631–641. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.H.; Beal, M.F. Amyloid beta, mitochondrial dysfunction and synaptic damage: Implications for cognitive decline in aging and Alzheimer’s disease. Trends Mol. Med. 2008, 14, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obulesu, M.; Venu, R.; Somashekhar, R. Lipid peroxidation in Alzheimer’s disease: Emphasis on metal-mediated neurotoxicity. Acta Neurol. Scand. 2011, 124, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Mantyh, P.W.; Ghilardi, J.R.; Rogers, S.; DeMaster, E.; Allen, C.J.; Stimson, E.R.; Maggio, J.E. Aluminum, Iron, and Zinc Ions Promote Aggregation of Physiological Concentrations of Β-amyloid Peptide. J. Neurochem. 1993, 61, 1171–1174. [Google Scholar] [CrossRef]
- Tönnies, E.; Trushina, E. Oxidative stress, synaptic dysfunction, and Alzheimer’s disease. J. Alzheimer’s Dis. 2017, 57, 1105–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Kamat, P.K.; Kalani, A.; Rai, S.; Swarnkar, S.; Tota, S.; Nath, C.; Tyagi, N. Mechanism of oxidative stress and synapse dysfunction in the pathogenesis of Alzheimer’s disease: Understanding the therapeutics strategies. Mol. Neurobiol. 2016, 53, 648–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultana, R.; Poon, H.F.; Cai, J.; Pierce, W.M.; Merchant, M.; Klein, J.B.; Markesbery, W.R.; Butterfield, D.A. Identification of nitrated proteins in Alzheimer’s disease brain using a redox proteomics approach. Neurobiol. Dis. 2006, 22, 76–87. [Google Scholar] [CrossRef]
- Sultana, R.; Perluigi, M.; Newman, S.F.; Pierce, W.M.; Cini, C.; Coccia, R.; Butterfield, D.A. Redox proteomic analysis of carbonylated brain proteins in mild cognitive impairment and early Alzheimer’s disease. Antioxid. Redox Signal 2010, 12, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butterfield, D.A.; Bader Lange, M.L.; Sultana, R. Involvements of the lipid peroxidation product, HNE, in the pathogenesis and progression of Alzheimer’s disease. Biochim. Biophys. Acta 2010, 1801, 924–929. [Google Scholar] [CrossRef] [Green Version]
- Markesbery, W.R.; Kryscio, R.J.; Lovell, M.A.; Morrow, J.D. Lipid peroxidation is an early event in the brain in amnestic mild cognitive impairment. Ann. Neurol. 2005, 58, 730–735. [Google Scholar] [CrossRef]
- Montine, T.J.; Quinn, J.; Kaye, J.; Morrow, J.D. F(2)-isoprostanes as biomarkers of late-onset Alzheimer’s disease. J. Mol. Neurosci. 2007, 33, 114–119. [Google Scholar] [CrossRef]
- Padurariu, M.; Ciobica, A.; Hritcu, L.; Stoica, B.; Bild, W.; Stefanescu, C. Changes of some oxidative stress markers in the serum of patients with mild cognitive impairment and Alzheimer’s disease. Neurosci. Lett. 2010, 469, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, M.; Sato, T.; Takino, J.; Kobayashi, Y.; Furuno, S.; Kikuchi, S.; Yamagishi, S. Diagnostic utility of serum or cerebrospinal fluid levels of toxic advanced glycation end-products (TAGE) in early detection of Alzheimer’s disease. Med. Hypotheses. 2007, 69, 1358–1366. [Google Scholar] [CrossRef] [PubMed]
- Marcus, D.L.; Thomas, C.; Rodriguez, C.; Simberkoff, K.; Tsai, J.S.; Strafaci, J.A.; Freedman, M.L. Increased peroxidation and reduced antioxidant enzyme activity in Alzheimer’s disease. Exp. Neurol. 1998, 150, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Forno, L.S. Neuropathology of Parkinson’s disease. J. Neuropathol. Exp Neurol. 1996, 55, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Spillantini, M.G.; Goedert, M. The alpha-synucleinopathies: Parkinson’s disease, dementia with Lewy bodies, and multiple system atrophy. Ann. N. Y. Acad. Sci. 2000, 920, 16–27. [Google Scholar] [CrossRef]
- Danielson, S.R.; Andersen, J.K. Oxidative and nitrative protein modifications in Parkinson’s disease. Free Radic. Biol. Med. 2008, 44, 1787–1794. [Google Scholar] [CrossRef] [Green Version]
- Selley, M.L. (E)-4-hydroxy-2-nonenal may be involved in the pathogenesis of Parkinson’s disease. Free Radic. Biol. Med. 1998, 25, 169–174. [Google Scholar] [CrossRef]
- Abe, T.; Isobe, C.; Murata, T.; Sato, C.; Tohgi, H. Alteration of 8-hydroxyguanosine concentrations in the cerebrospinal fluid and serum from patients with Parkinson’s disease. Neurosci. Lett. 2003, 336, 105–108. [Google Scholar] [CrossRef]
- Zhang, J.; Perry, G.; Smith, M.A.; Robertson, D.; Olson, S.J.; Graham, D.G.; Montine, T.J. Parkinson’s disease is associated with oxidative damage to cytoplasmic DNA and RNA in substantia nigra neurons. Am. J. Pathol. 1999, 154, 1423–1429. [Google Scholar] [CrossRef] [PubMed]
- Burai, R.; Ait-Bouziad, N.; Chiki, A.; Lashuel, H.A. Elucidating the role of site-specific nitration of α-synuclein in the pathogenesis of Parkinson’s disease via protein semisynthesis and mutagenesis. J. Am. Chem. Soc. 2015, 137, 5041–5052. [Google Scholar] [CrossRef]
- Trist, B.G.; Hare, D.J.; Double, K.L. Oxidative stress in the aging substantia nigra and the etiology of Parkinson’s disease. Aging Cell. 2019, 18, e13031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative sress: A key modulator in neurodegenerative diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [Green Version]
- Dalfó, E.; Ferrer, I. Early alpha-synuclein lipoxidation in neocortex in Lewy body diseases. Neurobiol. Aging 2008, 29, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Castellani, R.; Smith, M.A.; Richey, P.L.; Perry, G. Glycoxidation and oxidative stress in Parkinson disease and diffuse Lewy body disease. Brain Res. 1996, 737, 195–200. [Google Scholar] [CrossRef]
- Kunikowska, G.; Jenner, P. The distribution of copper, zinc- and manganese-superoxide dismutase, and glutathione peroxidase messenger ribonucleic acid in rat basal ganglia. Biochem. Pharmacol. 2002, 63, 1159–1164. [Google Scholar] [CrossRef]
- Schulz, J.B.; Lindenau, J.; Seyfried, J.; Dichgans, J. Glutathione, oxidative stress and neurodegeneration. Eur. J. Biochem. 2000, 267, 4904–4911. [Google Scholar] [CrossRef] [Green Version]
- Weinreb, O.; Mandel, S.; Youdim, M.B.H.; Amit, T. Targeting dysregulation of brain iron homeostasis in Parkinson’s disease by iron chelators. Free Radic. Biol. Med. 2013, 62, 52–64. [Google Scholar] [CrossRef]
- Kumar, K.; Mehra, R.; Guiné, R.P.F.; Lima, M.J.; Kumar, N.; Kaushik, R.; Ahmed, N.; Yadav, A.N.; Kumar, H. Edible Mushrooms: A comprehensive review on bioactive compounds with health benefits and processing aspects. Foods 2021, 10, 2996. [Google Scholar] [CrossRef]
- Das, A.K.; Nanda, P.K.; Dandapat, P.; Bandyopadhyay, S.; Gullón, P.; Sivaraman, G.K.; McClements, D.J.; Gullón, B.; Lorenzo, J.M. Edible mushrooms as functional ingredients for development of healthier and more sustainable muscle foods: A flexitarian approach. Molecules 2021, 26, 2463. [Google Scholar] [CrossRef]
- Carrasco, J.; Zied, D.C.; Pardo, J.E.; Preston, G.M.; Pardo-Gimenez, A. Supplementation in mushroom crops and its impact on yield and quality. AMB Express 2018, 8, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Tian, Y.; Menolli, N.; Ye, L.; Karunarathna, S.C.; Perez-Moreno, J.; Rahman, M.M.; Rashid, M.H.; Phengsintham, P.; Rizal, L.; et al. Reviewing the world’s edible mushroom species: A new evidence-based classification system. Comprehensive Reviews. Food Sci. Food Saf. 2021, 20, 1982–2014. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.; Negi, P.S. Bioactive components of mushrooms: Processing effects and health benefits. Food Res. Int. 2021, 148, 110599. [Google Scholar] [CrossRef]
- Rathore, H.; Prasad, S.; Sharma, S. Mushroom nutraceuticals for improved nutrition and better human health: A review. Pharma Nutr. 2017, 5, 35–46. [Google Scholar] [CrossRef]
- Gaoxing, M.; Wenjian, Y.; Liyan, Z.; Fei, P.; Donglu, F.; Qiuhui, H. A critical review on the health promoting effects of mushrooms nutraceuticals. Food Sci. Hum. Wellness. 2018, 7, 125–133. [Google Scholar]
- You, S.W.; Hoskin, R.T.; Komarnytsky, S.; Moncada, M. Mushrooms as functional and nutritious food ingredients for multiple applications. ACS Food Sci. Technol. 2022, 2, 1184–1195. [Google Scholar] [CrossRef]
- Reis, F.S.; Martins, A.; Vasconcelos, M.H.; Morales, P.; Ferreira, I.C.F.R. Functional foods based on extracts or compounds derived from mushrooms. Trend Food Sci. Technol. 2017, 66, 48–62. [Google Scholar] [CrossRef]
- Chang, S.T.; Wasser, S.P. The role of culinary-medicinal mushrooms on human welfare with a pyramid model for human health. Int. J. Med. Mushrooms 2012, 14, 95–134. [Google Scholar] [CrossRef] [PubMed]
- Lindequist, U.; Niedermayer, T.H.J.; Julich, W.D. The pharmacological potential of mushrooms. Evid. Based Complement. Alternat. Med. 2005, 2, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [PubMed]
- Feeney, M.J.; Dwyer, J.; Hasler-Lewis, C.M.; Milner, J.A.; Noakes, M.; Rowe, S.; Wach, M.; Beelman, R.B.; Caldwell, J.; Cantorna, M.T. Mushrooms and health summit proceedings. J. Nutr. 2014, 144, 1128S–1136S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Du, B.; Bian, Z.; Xu, B. Beta-glucans from edible and medicinal mushrooms: Characteristics, physicochemical and biological activities. J. Food Compos. Anal. 2015, 41, 165–173. [Google Scholar] [CrossRef]
- Islam, T.; Ganesan, K.; Xu, B. New insight into mycochemical profiles and antioxidant potential of edible and medicinal mushrooms: A Review. Int. J. Med. Mushrooms 2019, 21, 237–251. [Google Scholar] [CrossRef]
- Petraglia, T.; Latronico, T.; Liuzzi, G.M.; Fanigliulo, A.; Crescenzi, A.; Rossano, R. Edible mushrooms as source of fibrin (ogen) olytic enzymes: Comparison between four cultivated species. Molecules 2022, 27, 8145. [Google Scholar] [CrossRef]
- Slusarczyk, J.; Adamska, E.; Czerwik-Marcinkowska, J. Fungi and algae as sources of medicinal and other biologically active compounds: A Review. Nutrients 2021, 13, 3178. [Google Scholar] [CrossRef]
- Dubost, N.J.; Ou, B.; Beelman, R.B. Quantification of polyphenols and ergothioneine in cultivated mushrooms and correlation to total antioxidant capacity. Food Chem. 2007, 105, 727–735. [Google Scholar] [CrossRef]
- Sanchez, C. Reactive oxygen species and antioxidant properties from mushrooms. Synth. Syst. Biotechnol. 2017, 2, 13–22. [Google Scholar] [CrossRef]
- Sganzerla, W.G.; Todorov, S.D.; da Silva, A.P.G. Research trends in the study of edible mushrooms: Nutritional properties and health benefits. Int. J. Med. Mushrooms 2022, 24, 1–18. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.P.P.; Sulaiman Rahman, H. Antioxidant and oxidative stress: A mutual interplay in age-related diseases. Front. Pharmacol. 2018, 9, 1162. [Google Scholar] [CrossRef] [Green Version]
- Nile, S.H.; Ko, E.Y.; Kim, D.H.; Keum, Y.S. Screening of ferulic acid related compounds as inhibitors of xanthine oxidase and cyclooxygenase-2 with anti-inflammatory activity. Rev. Bras. Farmacogn. 2016, 26, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Rashmi, H.B.; Negi, P.S. Phenolic acids from vegetables: A review on processing stability and health benefits. Food Res. Int. 2020, 136, 109298. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Luo, T.; Li, S.; Zhou, Y.; Shen, X.Y.; He, F.; Xu, J.; Wang, H.O. Quercetin protects against Okadaic acid-induced injury via MAPK and PI3K/Akt/GSK3β signaling pathways in HT22 hippocampal neurons. PLoS ONE 2016, 11, 0152371. [Google Scholar] [CrossRef] [PubMed]
- Grassi, D.; Ferri, C.; Desideri, G. Brain protection and cognitive function: Cocoa flavonoids as nutraceuticals. Curr. Pharm. Des. 2016, 22, 145–151. [Google Scholar] [CrossRef]
- Braidy, N.; Behzad, S.; Habtemariam, S.; Ahmed, T.; Daglia, M.; Nabavi, S.M.; Sobarzo-Sanchez, E.; Nabavi, S.F. Neuroprotective effects of citrus fruit-derived flavonoids, nobiletin and tangeretin in Alzheimer’s and Parkinson’s disease. CNS Neurol. Disord. Drug Targets 2017, 16, 387–397. [Google Scholar] [CrossRef]
- Muszyńska, B.; Grzywacz-Kisielewska, A.; Kała, K.; Argasińska, J.G. Anti-inflammatory properties of edible mushrooms: A review. Food Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef]
- Çayan, F.; Deveci, E.; Tel-Çayan, G.; Duru, M.E. Identification and quantification of phenolic acid compounds of twenty-six mushrooms by HPLC–DAD. J. Food Meas. Charact. 2020, 14, 1690–1698. [Google Scholar] [CrossRef]
- Reis, F.S.; Barros, L.; Martins, A.; Ferreira, I.C.F.R. Chemical composition and nutritional value of the most widely appreciated cultivated mushrooms: An inter-species comparative study. Food Chem. Toxicol. 2012, 50, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Palacios, I.; Lozano, M.; Moro, C.; Arrigo, M.; Rostagno, M.A.; Martinez, J.A.; Garcia-Lafuente, A.; Guillamon, E.; Villares, A. Antioxidant properties of phenolic compound occurring in edible mushrooms. Food Chem. 2011, 128, 674–678. [Google Scholar] [CrossRef]
- Barros, L.; Duenas, M.; Ferreira, I.C.F.R.; Baptista, P.; Santos-Buelga, C. Phenolic acids determination by HPLC-DAD-ESI/MS in sixteen different Portuguese wild mushrooms species. Food Chem. Toxicol. 2019, 47, 1076–1079. [Google Scholar] [CrossRef]
- Butkhup, L.; Samappito, W.; Jorjong, S. Evaluation of bioactivities and phenolic contents of wild edible mushrooms from northeastern Thailand. Food Sci. Biotechnol. 2017, 27, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Cerletti, C.; Esposito, S.; Iacoviello, L. Edible mushrooms and Beta-Glucans: Impact on human health. Nutrients 2021, 13, 2195. [Google Scholar] [CrossRef] [PubMed]
- Kozarski, M.; Klaus, A.; Niksic, M.; Jakovljevic, D.; Helsper, J.P.F.G. Antioxidative and immunomodulating activities of polysaccharide extracts of the medicinal mushroom Agaricus bisporus, Agaricus brasiliensis, Ganoderma lucidum and Phelinus linteus. Food Chem. 2011, 129, 1667–1675. [Google Scholar] [CrossRef]
- Kozarski, M.S.; Klaus, A.S.; Niksic, M.P.; van Griensven, L.J.L.D.; Vrvic, M.M.; Jakovljevic, D.M. Polysaccharides of higher fungi: Biological role, structure and antioxidative activity. Chem. Ind. 2014, 68, 305–320. [Google Scholar] [CrossRef]
- Ping, C.X.; Yan, C.; Bing, L.S.; Guo, C.Y.; Yun, L.J.; Ping, L.L. Free radical scavenging of Ganoderma lucidum polysaccharides and its effect on antioxidant enzymes and immunity activities in cervical carcinoma rats. Carbohydr. Polym. 2009, 77, 389–393. [Google Scholar]
- Yan, J.; Zhu, L.; Qu, Y.; Qu, X.; Mu, M.; Zhang, M.; Muneer, G.; Zhou, Y.; Sun, L. Analyses of active antioxidant polysaccharides from four edible mushrooms. Int. J. Biol. Macromol. 2019, 123, 945–956. [Google Scholar] [CrossRef]
- Zhang, A.; Li, X.; Xing, C.; Yang, J.; Sun, P. Antioxidant activity of polysaccharide extracted from Pleurotus eryngii using response surface methodology. Int. J. Biol. Macromol. 2014, 65, 28–32. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, X.; Jia, W.; Gong, G.; Zhao, Y.; Li, G.; Zhou, J.; Li, X.; Zhao, Y.; Ma, W. Extraction, isolation, characterisation, antioxidant and anti-fatigue activities of Pleurotus eryngii polysaccharides. Int. J. Food Sci. Technol. 2020, 55, 2492–2508. [Google Scholar] [CrossRef]
- Li, F.; Zhang, Y.; Zhang, Z. Antihyper glycemic effect of Ganoderma lucidum polysaccharides on streptozocin-induced diabetic mice. Int. J. Mol. Sci. 2011, 12, 6135–6145. [Google Scholar] [CrossRef] [Green Version]
- Seweryn, E.; Ziała, A.; Gamian, A. Health-promoting of polysaccharides extracted from Ganoderma lucidum. Nutrients 2021, 13, 2725. [Google Scholar] [CrossRef]
- Petraglia, T.; Latronico, T.; Fanigliulo, A.; Crescenzi, A.; Liuzzi, G.M.; Rossano, R. Antioxidant activity of polysaccharides from the edible mushroom Pleurotus eryngii. Molecules 2023, 28, 2176. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Jiang, A.; Tian, M. Extraction optimization of antioxidant polysaccharides from Auricularia auricula fruiting bodies. Food Sci. Technol. 2015, 35, 428–433. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on mechanisms of in vitro antioxidant activity of polysaccharides. Oxid. Med. Cell. Long. 2016, 2016, 5692852. [Google Scholar]
- Kishk, Y.F.M.; Al-Sayed, H.M.A. Free-radical scavenging and antioxidative activities of some polysaccharides in emulsions. Food Sci. Technol. 2007, 40, 270–277. [Google Scholar] [CrossRef]
- Jung, H.Y.; Bae, I.Y.; Lee, S.; Lee, H.G. Effect of the degree of sulfation on the physicochemical and biological properties of Pleurotus eryngii polysaccharides. Food Hydrocoll. 2011, 25, 1291–1295. [Google Scholar] [CrossRef]
- Liu, W.; Wang, H.; Yao, W.; Gao, X.; Yu, L. Effects of sulfation on the physicochemical and functional properties of a water-insoluble polysaccharide preparation from Ganoderma lucidum. J. Agric. Food Chem. 2010, 58, 3336–3341. [Google Scholar] [CrossRef]
- Lo, T.C.T.; Chang, C.A.; Chiu, K.H.; Tsay, P.K.; Jen, J.F. Correlation evaluation of antioxidant properties on the monosaccharide components and glycosyl linkages of polysaccharide with different measuring methods. Carbohydr. Polym. 2011, 86, 320–327. [Google Scholar] [CrossRef]
- Berman, J.; Zorrilla-López, U.; Farré, G.; Zhu, C.; Sandmann, G.; Twyman, R.; Capell, T.; Christou, P. Nutritionally important carotenoids as consumer products. Phytochem. Rev. 2015, 14, 727–743. [Google Scholar] [CrossRef]
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- Sandmann, G. Carotenoids of biotechnological importance. Adv. Biochem. Eng. Biotechnol. 2015, 148, 449–467. [Google Scholar]
- Nishino, A.; Yasui, H.; Maoka, T. Reaction of paprika carotenoids, capsanthin and capsorubin, with reactive oxygen species. J. Agric. Food Chem. 2016, 64, 4786–4792. [Google Scholar] [CrossRef] [PubMed]
- Mohammadzadeh Honarvar, N.; Saedisomeolia, A.; Abdolahi, M.; Shayeganrad, A.; Taheri Sangsari, G.; Hassanzadeh Rad, B.; Muench, G. Molecular antiinflammatory mechanisms of retinoids and carotenoids in Alzheimer’s disease: A review of current evidence. J. Mol. Neurosci. 2017, 61, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, L.; Wang, Z.; Cui, Y.; Tan, X.; Yuan, T.; Liu, Q.; Liu, Z.; Liu, X. Supplementation of lycopene attenuates lipopolysaccharide-induced amyloidogenesis and cognitive impairments via mediating neuroinflammation and oxidative stress. J. Nutr. Biochem. 2018, 56, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Zhang, Y.; Li, Y.; Lu, K.; Shen, Y.; Guo, Y.; Qi, O.; Wang, M.; Zhang, S. NrF2/ARE and NF-kB pathway regulation may be the mechanism for lutein inhibition of human breast cancer cell. Future Oncol. 2018, 14, 719–726. [Google Scholar] [CrossRef]
- Carini, F.; David, S.; Tomasello, G.; Mazzola, M.; Damiani, P.; Rappa, F.; Battaglia, L.; Gerges Geagea, A.; Jurjus, R.; Leone, A. Colorectal cancer: An update on the effects of lycopene on tumor progression and cell proliferation. J. Biol. Regul. Homeost. Agents 2017, 31, 769–774. [Google Scholar] [PubMed]
- Aizawa, K.; Liu, C.; Tang, S.; Veeramachaneni, S.; Hu, K.Q.; Smith, D.E.; Wang, X.D. Tobacco carcinogen induces both lung cancer and nonalcoholic steatohepatitis and hepatocellular carcinomas in ferrets which can be attenuated by lycopene supplementation. Int. J. Cancer 2016, 139, 1171–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feart, C.; Letenneur, L.; Helmer, C.; Samieri, C.; Schalch, W.; Etheve, S.; Delcourt, C.; Dartigues, J.F.; Barberger-Gateau, P. Plasma carotenoids Aareinversely associated with dementia risk in an elderly French cohort. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 71, 683–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworska, G.; Pogon, K.; Skrzypczak, A.; Berna´s, E. Composition and antioxidant properties of wild mushrooms Boletus edulis and Xerocomus badius prepared for consumption. J. Food Sci. Technol. 2015, 52, 7944–7953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Li, S.L.; Chen, H.Y.; Zou, Y.; Zheng, Q.W.; Guo, L.Q.; Wu, G.H.; Lu, J.; Lin, J.F.; Ye, Z.W. Enhancement of carotenoid production and its regulation in edible mushroom Cordyceps militaris by abiotic stresses. Enzyme Microb. Technol. 2021, 148, 109808. [Google Scholar] [CrossRef]
- Elkanah, F.A.; Oke, M.A.; Adebayo, E.A. Substrate composition effect on the nutritional quality of Pleurotus ostreatus (MK751847) fruiting body. Heliyon 2022, 8, 1184. [Google Scholar] [CrossRef]
- Davey, M.W.; Van Montagu, M.; Inze, D.; Sanmartin, M.; Kanellis, A.; Smirnoff, N.; Benzie, I.J.J.; Strain, J.J.; Favell, D.; Fletcher, J. Plant L-ascorbic acid: Chemistry, function, metabolism, bioavailability and effects of processing. J. Sci. Food Agric. 2000, 80, 825–860. [Google Scholar] [CrossRef]
- May, J.M.; Harrison, F.E. Role of vitamin C in the function of the vascular endothelium. Antioxid. Redox Signal 2013, 19, 2068–2083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spector, R.; Johanson, C.E. The nexus of vitamin homeostasis and DNA synthesis and modification in mammalian brain. Mol. Brain 2014, 7, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sies, H.; Stahl, W.; Sundquist, A.R. Antioxidant functions of vitamins—Vitamins E and C, β-Carotene, and other carotenoids. Ann. N.Y. Acad. Sci. 1992, 669, 7–20. [Google Scholar] [CrossRef]
- Dixit, S.; Bernardo, A.; Walker, J.M.; Kennard, J.A.; Kim, G.Y.; Kessler, E.S.; Harrison, F.E. Vitamin C deficiency in the brain impairs cognition, increases amyloid accumulation and deposition, and oxidative stress in APP/PSEN1 and normally-aging mice. ACS Chem. Neurosci. 2015, 6, 570–581. [Google Scholar] [CrossRef] [Green Version]
- Mishra, K.K.; Pal, R.S.; Arunkumar, R.; Chandrashekara, C.; Jain, S.K.; Bhatt, J.C. Antioxidant properties of different edible mushroom species and increased bioconversion efficiency of Pleurotus eryngii using locally available casing materials. Food Chem. 2013, 138, 1557–1563. [Google Scholar] [CrossRef]
- Miyazawa, T.; Burdeos, G.C.; Itaya, M.; Nakagawa, K.; Miyazawa, T. Vitamin E: Regulatory redox interactions. IUBMB Life. 2019, 71, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Hensley, K.; Barnes, L.L.; Christov, A.; Tangney, C.; Honer, W.G.; Schneider, J.A.; Bennett, D.A.; Morris, M.C. Analysis of postmortem ventricular cerebrospinal fluid from patients with and without dementia indicates association of vitamin E with neuritic plaques and specific measures of cognitive performance. J. Alzheimer Dis. 2011, 24, 767–774. [Google Scholar] [CrossRef]
- Xu, W.; He, P.; He, S.; Cui, P.; Mi, Y.; Yang, Y. Gamma tocotrienol stimulates the proliferation, differentiation, and mineralization in osteoblastic MC3T3-E1 cells. J. Chem. 2018, 2018, 3805932. [Google Scholar] [CrossRef] [Green Version]
- Obodai, M.; Mensah, D.L.; Fernandes, Â.; Kortei, N.K.; Dzomeku, M.; Teegarden, M.; Schwartz, S.J.; Barros, L.; Prempeh, J.; Takli, R.K.; et al. Chemical characterization and antioxidant potential of wild Ganoderma species from Ghana. Molecules 2017, 22, 196. [Google Scholar] [CrossRef] [Green Version]
- Vieira, V.; Barros, L.; Martins, A.; Ferreira, I.C. Nutritional and biochemical profiling of Leucopaxillus candidus (Bres.) singer wild mushroom. Molecules 2016, 21, 99. [Google Scholar] [CrossRef] [Green Version]
- Borodina, I.; Kenny, L.C.; McCarthy, C.M.; Paramasivan, K.; Pretorius, E.; Roberts, T.J.; van der Hoek, S.A.; Kell, D.B. The biology of ergothioneine, an antioxidant nutraceutical. Nutr. Res. Rev. 2020, 33, 190–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apparoo, Y.; Phan, C.W.; Kuppusamy, U.R.; Sabaratnam, V. Ergothioneine and its prospects as an anti-ageing compound. Exp. Gerontol. 2022, 170, 111982. [Google Scholar] [CrossRef] [PubMed]
- Song, T.Y.; Lin, H.C.; Chen, C.L.; Wu, J.H.; Liao, J.W.; Hu, M.L. Ergothioneine and melatonin attenuate oxidative stress and protect against learning and memory deficits in C57BL/6J mice treated with D-galactose. Free Radic. Res. 2014, 48, 1049–1060. [Google Scholar] [CrossRef]
- Tang, R.M.Y.; Cheah, I.K.M.; Yew, T.S.K.; Halliwell, B. Distribution and accumulation of dietary ergothioneine and its metabolites in mouse tissues. Sci. Rep. 2018, 8, 1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N.C.; Lin, H.C.; Wu, J.H.; Ou, H.C.; Chai, Y.C.; Tseng, C.Y.; Liao, J.W.; Song, T.Y. Ergothioneine protects against neuronal injury induced by β-amyloid in mice. Food Chem. Toxicol. 2012, 50, 3902–3911. [Google Scholar] [CrossRef] [PubMed]
- Cheah, I.K.; Feng, L.; Tang, R.M.Y.; Lim, K.H.C.; Halliwell, B. Ergothioneine levels in an elderly population decrease with age and incidence of cognitive decline; a risk factor for neurodegeneration? Biochem. Biophys. Res. Commun. 2016, 478, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Sotgia, S.; Zinellu, A.; Mangoni, A.A.; Pintus, G.; Attia, J.; Carru, C.; McEvoy, M. Clinical and biochemical correlates of serum L-ergothioneine concentrations in community-dwelling middle-aged and older adults. PLoS ONE 2014, 9, e84918. [Google Scholar] [CrossRef]
- Phan, C.W.; David, P.; Sabaratnam, V. Edible and medicinal mushrooms: Emerging brain food for the mitigation of neurodegenerative diseases. J. Med. Food. 2017, 20, 1–10. [Google Scholar] [CrossRef]
- Halliwell, B.; Cheah, I.K.; Tang, R.M.Y. Ergothioneine a diet derived antioxidant with therapeutic potential. FEBS Lett. 2018, 592, 3357–3366. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gapstur, S.M.; Carter, B.D.; Hartman, T.J.; Stevens, V.L.; Gaudet, M.M.; McCullough, M.L. Untargeted metabolomics identifies novel potential biomarkers of habitual food intake in a cross-sectional study of postmenopausal women. J. Nutr. 2018, 148, 932–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Huang, Y.; Wang, J.; Zhou, S.; Wang, Y.; Cai, M.; Yu, L.; Tang, Q. A study on the antioxidant properties and stability of ergothioneine from culinary-medicinal mushrooms. Int. J. Med. Mushrooms 2020, 22, 211–220. [Google Scholar] [CrossRef]
- Dubost, N.J.; Beelman, R.; Peterson, D.; Royse, D. Identification and quantification of ergothioneine in cultivated mushrooms by liquid chromatography–mass spectroscopy. Int. J. Med. Mushrooms 2005, 8, 215–222. [Google Scholar] [CrossRef]
- Kalaras, M.D.; Richie, J.P.; Calcagnotto, A.; Beelman, R.B. Mushrooms: A rich source of the antioxidants ergothioneine and glutathione. Food Chem. 2017, 233, 429–433. [Google Scholar] [CrossRef]
- Tepwong, P.; Giri, A.; Ohshima, T. Effect of mycelial morphology on ergothioneine production during liquid fermentation of Lentinula edodes. Mycoscience 2012, 53, 102–112. [Google Scholar] [CrossRef]
- Lin, S.Y.; Chien, S.C.; Wang, S.Y.; Mau, J.L. Nonvolatile taste components and antioxidant properties of fruiting body and mycelium with high ergothioneine content from the culinarymedicinal golden oyster mushroom Pleurotus citrinopileatus (Agaricomycetes). Int. J. Med. Mushrooms 2016, 18, 689–698. [Google Scholar] [CrossRef]
- Stastny, J.; Marsik, P.; Tauchen, J.; Bozik, M.; Mascellani, A.; Havlik, J.; Landa, P.; Jablonsky, I.; Treml, J.; Herczogova, P.; et al. Antioxidant and anti-inflammatory activity of five medicinal mushrooms of the genus Pleurotus. Antioxidants 2022, 11, 1569. [Google Scholar] [CrossRef] [PubMed]
- Roda, E.; Priori, E.C.; Ratto, D.; De Luca, F.; Di Iorio, C.; Angelone, P.; Locatelli, C.A.; Desiderio, A.; Goppa, L.; Savino, E.; et al. Neuroprotective metabolites of Hericium erinaceus promote neuro-healthy aging. Int. J. Mol. Sci. 2021, 22, 6379. [Google Scholar] [CrossRef]
- Falandysz, J.; Borovička, J. Macro and trace mineral constituents and radionuclides in mushrooms: Health benefits and risks. Appl. Microbiol. Biotechnol. 2013, 97, 477–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haro, A.; Trescastro, A.; Lara, L.; Fernández-Fígares, I.; Nieto, R.; Seique, I. Mineral elements content of wild growing edible mushrooms from the southeast of Spain. J. Food Compost. Anal. 2020, 91, 103504. [Google Scholar] [CrossRef]
- Marklund, S.R.; Westman, N.G.; Lundgren, E.; Ross, G. Copper and zinc-containing superoxide-dismutase, manganese-containing superoxide dismutase, catalase and gtutathione peroxidase in normal and neoplastic human cell lines and normal tissue. Cancer Res. 1982, 42, 1955–1961. [Google Scholar] [PubMed]
- Marreiro, D.D.; Cruz, K.J.; Morais, J.B.; Beserra, J.B.; Severo, J.S.; deOliveira, A.R.S. Zinc and oxidative stress: Current mechanisms. Antioxidants 2017, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Gałgowska, M.; Pietrzak-Fiecko, R. Mineral Composition of three popular wild mushrooms from Poland. Molecules 2020, 25, 3588. [Google Scholar] [CrossRef] [PubMed]
- Giannaccini, G.; Betti, L.; Palego, L.; Mascia, G.; Schmid, L.; Lanza, M.; Mela, A.; Fabbrini, L.; Biondi, L.; Lucacchini, A. The trace element content of top-soil and wild edible mushroom samples collected in Tuscany, Italy. Environ. Monit. Assess. 2012, 184, 7579–7585. [Google Scholar] [CrossRef] [PubMed]
- Gençcelep, H.; Uzun, Y.; Tunçtürk, Y.; Demirel, K. Determination of mineral contents of wild-grown edible mushrooms. Food Chem. 2009, 113, 1033–1036. [Google Scholar] [CrossRef]
- Györfi, J.; Geösel, A.; Vetter, J. Mineral composition of different strains of edible medicinal mushroom Agaricus subrufescens Peck. J. Med. Food. 2010, 13, 1510–1514. [Google Scholar] [CrossRef]
- Vetter, J.; Hajdú, J.; Györfi, J.; Maszlavér, P. Mineral composition of the cultivated mushrooms Agaricus bisporus, Pleurotus ostreatus and Lentinula eodes. Acta Alim. 2005, 34, 441–451. [Google Scholar] [CrossRef]
- Zheng, L.; Liu, M.; Zhai, G.Y.; Ma, Z.; Wang, L.Q.; Jia, L. Antioxidant and anti-ageing activities of mycelia zinc polysaccharide from Pholiota nameko SW-03. J. Sci. Food Agric. 2015, 95, 3117–3126. [Google Scholar] [CrossRef]
- Reeves, M.A.; Hoffmann, P.R. The human selenoproteome: Recent insights into functions and regulation. Cell. Mol. Life Sci. 2009, 66, 2457–2478. [Google Scholar] [CrossRef] [Green Version]
- Papp, L.V.; Holmgren, A.; Khanna, K.K. Selenium and selenoproteins in health and disease. Antioxid. Redox Signal 2010, 12, 793–795. [Google Scholar] [CrossRef]
- Cai, Z.; Zhang, J.; Li, H. Selenium, aging and aging-related diseases. Aging Clin. Exp. Res. 2019, 31, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Prange, A.; Sari, M.; von Ameln, S.; Hajdu, C.; Hambitzer, R.; Ellinger, S.; Hormes, J. Characterization of selenium speciation in selenium-enriched button mushrooms (Agaricus bisporus) and selenized yeasts (dietary supplement) using X-ray absorption near-edge structure (XANES) spectroscopy. J. Trace Elem. Med. Biol. 2019, 51, 164–168. [Google Scholar] [CrossRef]
- Gagandeep, K.; Anu, K.; Harpreet, S. Selenium bio-fortification of Pleurotus species and its effect on yield, phytochemical profiles and protein chemistry of fruiting bodies. J. Food Biochem. 2018, 42, e12467. [Google Scholar]
- Dong, Z.; Xiao, Y.; Wu, H. Selenium accumulation, speciation, and its effect on nutritive value of Flammulina velutipes (Golden needle mushroom). Food Chem. 2021, 350, 128667. [Google Scholar] [CrossRef] [PubMed]
- Pickart, L.; Vasquez-Soltero, J.M.; Margolina, A. The human tripeptide GHK-Cu in prevention of oxidative stress and degenerative conditions of aging: Implications for cognitive health. Oxid. Med. Cell Longev. 2012, 2012, 324832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, Y.; Lee, A.; Zhu, L.; Morton, J.; Ladiges, W. The potential of GHK as an anti-aging peptide. Aging Pathobiol. Ther. 2020, 2, 58–61. [Google Scholar] [CrossRef] [Green Version]
- Tuzen, M. Determination of heavy metals in soil, mushroom and plant samples by atomic adsorption spectrometry. Microchem. J. 2003, 74, 289–297. [Google Scholar] [CrossRef]
- Chen, X.H.; Zhou, H.B.; Qiu, G.Z. Analysis of several heavy metals in wild edible mushrooms from regions of China. Bull. Environ. Contam. Toxicol. 2009, 83, 280–285. [Google Scholar] [CrossRef]
- Keles, A.; Genccelep, H.; Demirel, K. Elemental composition of naturally growing wild edible mushroom. J. Nat. Prod.Plant Resour. 2017, 7, 37–44. [Google Scholar]
- Fu, Z.; Liu, G.; Wang, L. Assessment of potential human health risk of trace element in wild edible mushroom species collected from Yunnan Province, China. Environ. Sci. Pollut. Res. 2020, 27, 29218–29227. [Google Scholar] [CrossRef]
- Winterbourn, C.C. Revisiting the reactions of superoxide with glutathione and other thiols. Arch. Biochem. Biophys. 2016, 595, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Braidy, N.; Zarka, M.; Peter, C.; Welch, J.; Bridge, W. Therapeutic approaches to modulating glutathione levels as a pharmacological strategy in Alzheimer’s disease. Curr. Alzheimer Res. 2015, 12, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Homma, T.; Fujii, J. Application of glutathione as anti-oxidative and anti-aging drugs. Curr. Drug. Metab. 2015, 16, 560–571. [Google Scholar] [CrossRef] [PubMed]
- Dogan, H.; Coteli, E.; Karatas, F. Determination of glutathione, selenium, and malondialdehyde in different edible mushroom species. Biol. Trace Elem. Res. 2016, 174, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Ir, R.; Jeewon, R.; Doble, M.; Hyde, K.D.; Kaliappan, I.; Jeyaraman, R.; Reddi, R.N.; Krishnan, J.; Li, M.; et al. A Mechanistic Review on Medicinal Mushrooms-Derived Bioactive Compounds: Potential Mycotherapy Candidates for Alleviating Neurological Disorders. Planta Med. 2020, 86, 1161–1175. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, R.W.; Macharia, J.M.; Wagara, I.N.; Bence, R.L. The antioxidant potential of different edible and medicinal mushrooms. Biomed. Pharmacother. 2022, 147, 112621. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Van Griensven, L.J.L.D. Pro-and antioxidative properties of medicinal mushroom extracts. Int. J. Med. Mushrooms 2008, 10, 315–324. [Google Scholar] [CrossRef]
- Ferreira, I.C.F.R.; Barros, L.; Abreu, R.M.V. Antioxidants in wild mushrooms. Curr. Med. Chem. 2009, 16, 1543–1560. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Luo, J.; Qi, J.; Zhao, X.; An, P.; Luo, Y.; Wang, G. The role and mechanism of polysaccharides in anti-aging. Nutrients 2022, 14, 5330. [Google Scholar] [CrossRef]
- Li, S.; Liu, H.; Wang, W.; Wang, X.; Zhang, C.; Zhang, J.; Jing, H.; Ren, Z.; Gao, Z.; Song, X.; et al. Antioxidant and anti-aging effects of acidic-extractable polysaccharides by Agaricus bisporus. Int. J. Biol. Macromol. 2018, 106, 1297–1306. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, Z.; Li, S.; Gao, S.; Ren, Z.; Jing, H.; Jia, L. Purification, characterization, antioxidation, and antiaging properties of exopolysaccharides and endopolysaccharides of the royal sun medicinal mushroom, Agaricus brasiliensis (Agaricomycetes). Int. J. Med. Mushrooms 2016, 18, 1071–1081. [Google Scholar] [CrossRef]
- Jing, H.; Li, J.; Zhang, J.; Wang, W.; Li, S.; Ren, Z.; Gao, Z.; Song, X.; Wang, X.; Jia, L. The antioxidative and anti-aging effects of acidic- and alkalic-extractable mycelium polysaccharides by Agrocybe aegerita (Brig). Sing. Int. J. Biol. Macromol. 2018, 106, 1270–1278. [Google Scholar] [CrossRef]
- Liu, M.; Jing, H.; Zhang, J.; Che, G.; Zhou, M.; Gao, Z.; Li, S.; Ren, Z.; Hao, L.; Liu, Y.; et al. Optimization of mycelia selenium polysaccharide extraction from Agrocybe cylindracea SL-02 and assessment of their antioxidant and anti-ageing activities. PLoS ONE 2016, 11, e0160799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Li, J.; Zhang, J.; Wang, X.; Hao, L.; Jia, L. Purification, in vitro antioxidant and in vivo anti-aging activities of exopolysaccharides by Agrocybe cylindracea. Int. J. Biol. Macromol. 2017, 102, 351–357. [Google Scholar] [CrossRef]
- Yuan, F.; Gao, Z.; Liu, W.; Li, H.; Zhang, Y.; Feng, Y.; Song, X.; Wang, W.; Zhang, J.; Huang, C.; et al. Characterization, antioxidant, anti-aging and organ protective effects of sulfated polysaccharides from Flammulina velutipes. Molecules 2019, 24, 3517. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Li, Y.; Pei, G. Polysaccharides from Ganoderma lucidum attenuate microgliamediated neuroinflammation and modulate microglial phagocytosis and behavioural response. J. Neuroinflammation 2017, 14, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, C.S.W.; Yu, M.S.; Yuen, W.H.; So, K.F.; Zee, S.Y.; Chang, R.C.C. Antagonizing beta-amyloid peptide neurotoxicity of the anti-aging fungus Ganoderma lucidum. Brain Res. 2008, 1190, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Lai, G.; Guo, Y.; Chen, D.; Tang, X.; Shuai, O.; Yong, T.; Wang, D.; Xiao, C.; Zhou, G.; Xie, Y.; et al. Alcohol extracts from Ganoderma lucidum delay the progress of Alzheimer’s disease by regulating DNA methylation in rodents. Front. Pharmacol. 2019, 10, 272–298. [Google Scholar] [CrossRef]
- Zhou, Y.; Qu, Z.; Zeng, Y.; Lin, Y.; Chung, P. Neuroprotective effect of preadministration with Ganoderma lucidum spore on rat hippocampus. Exp. Toxicol. Pathol. 2012, 64, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Xu, S.; Cai, Y.; Zhou, M.; Zuo, X.; Chan, P. Ganoderma lucidum protects dopaminergic neuron degeneration through inhibition of microglial activation. Evid. Based Complement. Alternat. Med. 2011, 2011, 156810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Tang, Y.; Liu, A.; Jin, X.; Zhu, J.; Lu, X. Oral administration of Grifola frondosa polysaccharides improves memory impairment in aged rats via antioxidant action. Mol. Nutr. Food Res. 2017, 61, 1700313. [Google Scholar] [CrossRef]
- Li, H.; Zhao, H.; Liu, W.; Feng, Y.; Zhang, Y.; Yuan, F.; Jia, L. Liver and brain protective effect of sulfated polysaccharides from residue of lion’s mane medicinal mushroom, Hericium erinaceus (Agaricomycetes), on D-Galactose-Induced Aging Mice. Int. J. Med. Mushrooms 2021, 23, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, C.; Gao, X.; Xu, N.; Lin, L.; Zhao, H.; Jia, S.; Jia, L. Purification, characterization and anti-aging capacity of mycelia zinc polysaccharide by Lentinus edodes SD-08. BMC Complement. Altern. Med. 2015, 15, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, W.; Liu, N.; Xie, Y.; Zhao, Y.; Song, X.; Zhong, W. Antioxidant and anti-aging activities of mycelial polysaccharides from Lepista sordida. Int. J. Biol. Macromol. 2013, 60, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhao, H.; Gao, Z.; Song, X.; Wang, W.; Yuan, F.; Feng, Y.; Zhang, Y.; Zhang, J.; Zhan, S.; et al. The antioxidant and anti-aging effects of acetylated mycelia polysaccharides from Pleurotus Djamor. Molecules 2019, 24, 2698. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.J.; Guo, J.Y.; Cheng, H.; Lin, L.; Liu, Y.; Shi, Y.; Xu, J.; Yu, H.T. Protective effects of the king oyster culinary-medicinal mushroom, Pleurotus eryngii (Agaricomycetes), Polysaccharides on β-amyloid-induced neurotoxicity in PC12 cells and aging rats, in vitro and in vivo studies. Int. J. Med. Mushrooms 2020, 22, 325–333. [Google Scholar] [CrossRef]
- Zhang, C.; Song, X.; Cui, W.; Yang, Q. Antioxidant and anti-ageing effects of enzymatic polysaccharide from Pleurotus eryngii residue. Int. J. Biol. Macromol. 2021, 173, 341–350. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, X.; Jin, G.; Yang, X.; Zhang, Y. Polysaccharides from Pleurotus ostreatus alleviate cognitive impairment in a rat model of Alzheimer’s disease. Int. J. Biol. Macromol. 2016, 92, 935–941. [Google Scholar] [CrossRef]
- Liu, M.; Dai, Y.; Song, C.; Wang, J.; Liu, Y.; Wang, Q. Structural characterization of a Pleurotus sajor-caju polysaccharide and its neuroprotection related to the inhibition of oxidative stress. Nutrients 2022, 14, 4047. [Google Scholar] [CrossRef]
- Shen, T.; Duan, C.; Chen, B.; Li, M.; Ruan, Y.; Xu, D.; Shi, D.; Yu, D.; Li, J.; Wang, C. Tremella fuciformis polysaccharide suppresses hydrogen peroxide-triggered injury of human skin fibroblasts via upregulation of SIRT1. Mol. Med. Rep. 2017, 16, 1340–1346. [Google Scholar] [CrossRef] [Green Version]
- Ding, Q.; Yang, D.; Zhang, W.; Lu, Y.; Zhang, M.; Wang, L.; Li, X.; Zhou, L.; Wu, Q.; Pan, W.; et al. Antioxidant and anti-aging activities of the polysaccharide TLH-3 from Tricholoma lobayense. Int. J. Biol. Macromol. 2016, 85, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.H.; Tsai, C.L.; Lien, Y.Y.; Lee, M.S.; Sheu, S.C. High molecular weight of polysaccharides from Hericium erinaceus against amyloid beta-induced neurotoxicity. BMC Complement. Altern. Med. 2016, 16, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; An, S.; Hu, W.; Teng, M.; Wang, X.; Qu, Y.; Liu, Y.; Yuan, Y.; Wang, D. The Neuroprotective Properties of Hericium erinaceus in glutamate-damaged differentiated PC12 cells and an Alzheimer’s disease mouse model. Int. J. Mol. Sci. 2016, 17, 1810. [Google Scholar] [CrossRef] [PubMed]
- Kushairi, N.; Phan, C.W.; Sabaratnam, V.; David, P.; Naidu, M. Lion’s Mane Mushroom, Hericium erinaceus (Bull.: Fr.) Pers. Suppresses H2O2-induced oxidative damage and LPS-induced inflammation in HT22 hippocampal neurons and BV2 microglia. Antioxidants 2019, 8, 261. [Google Scholar] [CrossRef] [Green Version]
- An, S.; Lu, W.; Zhang, Y.; Yuan, Q.; Wang, D. Pharmacological basis for use of Armillaria mellea polysaccharides in Alzheimer’s Disease: Antiapoptosis and antioxidation. Oxid. Med. Cell. Longev. 2017, 2017, 4184562. [Google Scholar] [CrossRef] [Green Version]
- Tripodi, F.; Falletta, E.; Leri, M.; Angeloni, C.; Beghelli, D.; Giusti, L.; Milanesi, R.; Sampaio-Marques, B.; Ludovico, P.; Goppa, L.; et al. Anti-aging and neuroprotective properties of Grifola frondosa and Hericium erinaceus extracts. Nutrients 2022, 14, 4368. [Google Scholar] [CrossRef]
- Rizzo, G.; Goggi, S.; Giampieri, F.; Baroni, L. A review of mushrooms in human nutrition and health. Trends Food Sci. Technol. 2021, 117, 60–73. [Google Scholar] [CrossRef]
- Mori, K.; Inatomi, S.; Ouchi, K.; Azumi, Y.; Tuchida, T. Improving effects of the mushroom yamabushitake (Hericium erinaceus) on mild cognitive impairment: A double-blind placebo-controlled clinical trial. Phytother. Res. 2009, 23, 367–372. [Google Scholar] [CrossRef]
- Feng, L.; Cheah, I.K.M.; Ng, M.M.X.; Li, J.; Chan, S.M.; Lim, S.L.; Mahendran, R.; Kua, E.H.; Halliwell, B. The association between mushroom consumption and mild cognitive impairment: A community-based cross-sectional study in Singapore. J. Alzheimers Dis. 2019, 68, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Tsuk, S.; Lev, Y.H.; Rotstein, A.; Carasso, R.; Zeev, A.; Netz, Y.; Dwolatski, T.; Steiner, G. Clinical effects of a commercial supplement of Ophiocordyceps sinensis and Ganoderma lucidum on cognitive function of healthy young volunteers. Int. J. Med. Mushrooms 2017, 19, 667–673. [Google Scholar] [CrossRef]
- Zhang, S.; Tomata, Y.; Sugiyama, K.; Sugawara, Y.; Tsuji, I. Mushroom consumption and incident dementia in elderly Japanese: The Ohsaki cohort 2006 study. J. Am. Geriatr. Soc. 2017, 65, 1462–1469. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Arif, M.; Xu, M.; Zhang, J.; Ding, Y.; Lyu, F. Therapeutic values and nutraceutical properties of shiitake mushroom (Lentinula edodes): A review. Trends Food Sci. Technol. 2023, 134, 123–135. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mushroom Species | Bioactive Compounds/Extracts | Mechanism of Action | References |
|---|---|---|---|
| Agaricus bisporus | Acidic-extractable Polysaccharides (AcAPS) | In vitro: hydroxyl and DPPH scavenging activities. In vivo: hepatic and nephric protection by improving serum enzyme activities in aging mice. | [170] |
| Agaricus brasiliensis |
Exopolysaccharides (ExPSs) and endopolysaccharides (EnPSs) | In vitro: hydroxyl and DPPH scavenging activities, reducing power. Improvement of total antioxidant capability, decrease in MDA content. | [171] |
| Agrocybe aegerita | Acidic- and alkalic-extractable polysaccharides (Ac-MPS and Al-MPS) | In vitro: hydroxyl and DPPH scavenging activities. In vivo: increase in SOD, CAT, GPx and total antioxidant capacity; decrease in MDA and lipid peroxidation; reduction of serum levels of triglycerides and total cholesterol in aging mice. | [172] |
| Agrocybe cylindracea | Selenium polysaccharides (SL-02) Exopolysaccharides (EPS) | In vitro: hydroxyl and DPPH scavenging activities, reducing power. In vivo: increase in SOD, GPx, and total antioxidant capacity; decrease in MDA and total cholesterol in aging mice. | [173,174] |
| Flammulina velutipes | Sulfated polysaccharides (SFPS) | In vitro: DPPH, hydroxyl, superoxide and scavenging activities; reducing power; Fe2+-chelating capacity In vivo: increase in the antioxidant enzyme activities; decrease in lipid peroxidation, improvement of the inflammatory response in mice. | [175] |
| Ganoderma lucidum | Polysaccharides | Reduction of amyloid toxicity; decrease in neurotoxicity; increase in GSH, GPx activities; decrease in MDA. Protection of dopaminergic neurons from inflammation. Inhibition of microglial activation; decrease oin TNF-α and IL-1β. | [176,177,178,179,180] |
| Grifola frondosa | Polysaccharides (GFP) Intracellular zinc polysaccharides (IZPS) | In vitro: hydroxyl, superoxide and DPPH scavenging activities, reducing power. In vivo: increase in SOD, CAT, GPx activities, and total antioxidant capacity; decrease in MDA and nitric oxide levels and amelioration of age-associated changes of brain histology. | [181] |
| Hericium erinaceus | Sulfated residue polysaccharides (SHRPs) | In vitro: scavenging activities. In vivo: increase in enzyme activities, decrease in MDA. Improvement of serum biochemical indices and of immunological activity. | [182] |
| Lentinula edodes | Mycelia polysaccharides (MPS) and mycelia zinc polysaccharides (MZPS) | In vitro: hydroxyl and DPPH scavenging activities, reducing power. In vivo: increase in SOD, GPx, and total antioxidant capacity; decrease in MDA in aging mice. | [183] |
| Lepista sordida | Intracellular polysaccharides (CLSP) | In vitro: hydroxyl, superoxide, and DPPH scavenging activities. In vivo: inhibition of MDA formation; increase in SOD and GPx in aging mice. | [184] |
| Pholiota nameko | Zinc-enriched polysaccharides (MZPS) | Improvement of antioxidant status (SOD, total antioxidant capability, MDA and lipid peroxide) in aging mice. | [148] |
| Pleurotus djamor | Acetylated mycelia polysaccharides (AMPS) | In vitro: hydroxyl, superoxide, and DPPH scavenging activities; reducing power. In vivo: increase in SOD, CAT, and GPx activities; decrease in lipid peroxidation and MDA. Improvement of serum biochemical indices and immunological activity in the liver, kidney, and brain of aging mice. | [185] |
| Pleurotus eryngii | Polysaccharides (PEP) Enzymatic residue polysaccharide (PERP) | Neuroprotective actions against β-amyloid-induced neurotoxicity in cultured rat PC12 cells and aging rats. In vitro: hydroxyl, superoxide and DPPH scavenging activities; reducing power. In vivo: increase in SOD, CAT, GPx activities and total antioxidant capacity; suppression of lipid peroxidation. Improvement of organ functions and histopathological damage in brain, liver, kidney, and skin. | [186,187] |
| Pleurotus ostreatus | Polysaccharides (POP) | Improvement of cognitive impairment in a rat model of Alzheimer’s disease. Increase in SOD, CAT, and GPx activities; reduction of MDA levels and AchE activity. | [188] |
| Pleurotus sajor-caju | Polysaccharide PSP2-1 | Improvement of oxidative stress injury, inhibition of apoptosis in H2O2-induced neuronal cells. Improvement of cognition in aging mice. | [189] |
| Tremella fuciformis | Polysaccharide (TFPS) | Improvement of H2O2-induced oxidative stress and inhibition of apoptosis in human skin fibroblasts via upregulation of SIRT1 expression. | [190] |
| Tricholoma lobayense | Polysaccharide TLH-3 | Inhibition of MDA and increase in SOD and CAT activities in liver and serum of aged mice. | [191] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liuzzi, G.M.; Petraglia, T.; Latronico, T.; Crescenzi, A.; Rossano, R. Antioxidant Compounds from Edible Mushrooms as Potential Candidates for Treating Age-Related Neurodegenerative Diseases. Nutrients 2023, 15, 1913. https://doi.org/10.3390/nu15081913
Liuzzi GM, Petraglia T, Latronico T, Crescenzi A, Rossano R. Antioxidant Compounds from Edible Mushrooms as Potential Candidates for Treating Age-Related Neurodegenerative Diseases. Nutrients. 2023; 15(8):1913. https://doi.org/10.3390/nu15081913
Chicago/Turabian StyleLiuzzi, Grazia Maria, Tania Petraglia, Tiziana Latronico, Aniello Crescenzi, and Rocco Rossano. 2023. "Antioxidant Compounds from Edible Mushrooms as Potential Candidates for Treating Age-Related Neurodegenerative Diseases" Nutrients 15, no. 8: 1913. https://doi.org/10.3390/nu15081913
APA StyleLiuzzi, G. M., Petraglia, T., Latronico, T., Crescenzi, A., & Rossano, R. (2023). Antioxidant Compounds from Edible Mushrooms as Potential Candidates for Treating Age-Related Neurodegenerative Diseases. Nutrients, 15(8), 1913. https://doi.org/10.3390/nu15081913