Dietary Phytochemicals Promote Health by Enhancing Antioxidant Defence in a Pig Model


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Plant Extracts
2.3. Total Phenolic Content of Plant Extracts
2.4. Prediction of Human Absorption Kinetics of Plant Extracts
2.5. Dietary Intervention Using an Animal Model
2.5.1. Animals and Background Diet
2.5.2. Cephalic Vein Catheterisation Procedure
2.5.3. Experimental Design and Procedure
2.5.4. Plasma Total Antioxidant Capacity Assay
2.5.5. Plasma Glutathione Peroxidase Activity
2.6. Hydrogen Peroxide Production of Plant Extracts in Pig Plasma In Vitro
2.7. Data Analysis
3. Results
3.1. Predicted Human Absorption as Functional Fingerprints of Plant Extracts
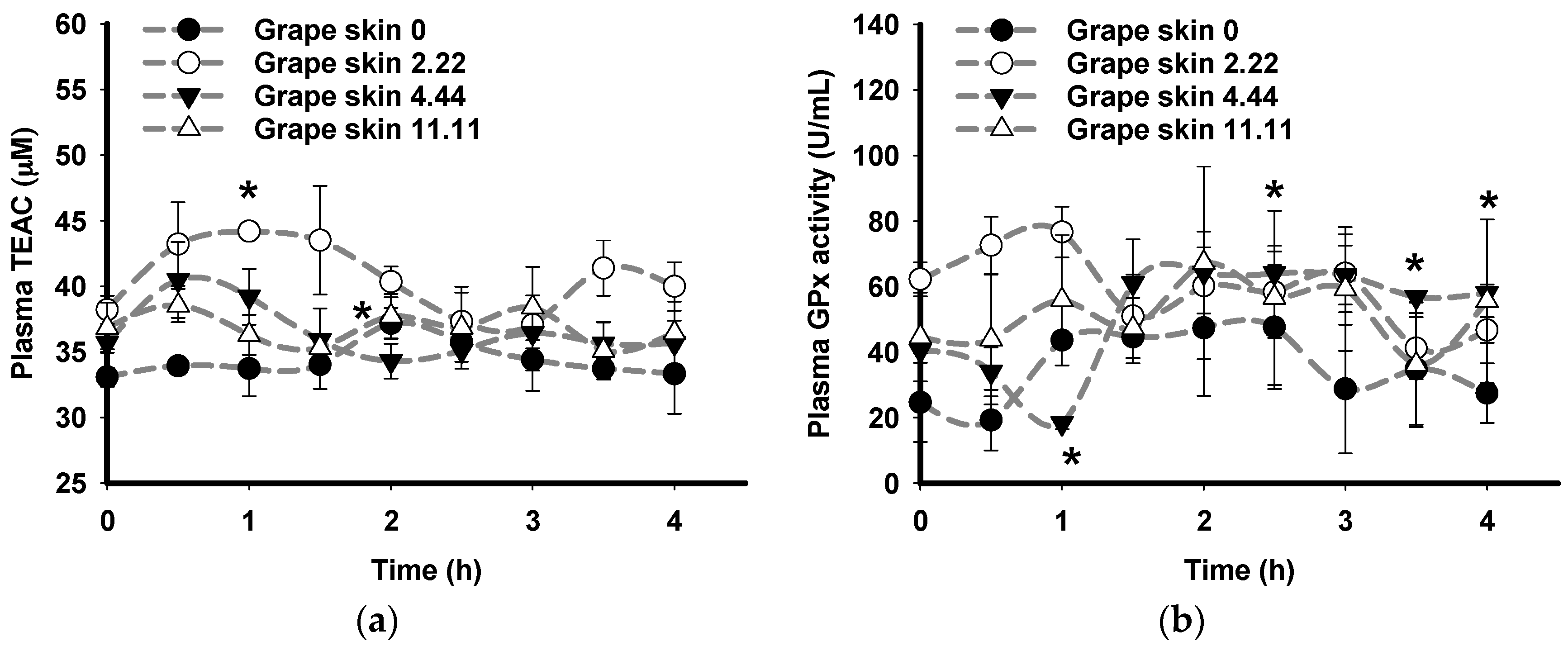
3.2. Time Course Effects of Oral Consumption of Plant Extracts on Plasma Antioxidant Status Ex Vivo of Pigs
3.3. Effects of Plant Extracts on Plasma Total Antioxidant Capacity and Plasma Hydrogen Peroxide Concentration In Vitro
3.4. Effects of Phytochemical Dose and Their H2O2 Production Capacity In Vitro on Plasma Antioxidant Status of Pigs Ex Vivo
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Key, T.J. Fruit and vegetables and cancer risk. Br. J. Cancer 2011, 104, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Dauchet, L.; Amouyel, P.; Dallongeville, J. Fruits, vegetables and coronary heart disease. Nat. Rev. Cardiol. 2009, 6, 599–608. [Google Scholar] [CrossRef] [PubMed]
- D’Onofrio, G.; Sancarlo, D.; Ruan, Q.; Yu, Z.; Panza, F.; Daniele, A.; Greco, A.; Seripa, D. Phytochemicals in the treatment of alzheimer’s disease: A systematic review. Curr. Drug Targets 2016, 17. [Google Scholar] [CrossRef]
- Burton-Freeman, B. Postprandial metabolic events and fruit-derived phenolics: A review of the science. Br. J. Nutr. 2010, 104, S1–S14. [Google Scholar] [CrossRef] [PubMed]
- Van der Merwe, M.; Bloomer, R.J. The influence of methylsulfonylmethane on inflammation-associated cytokine release before and following strenuous exercise. J. Sports Med. 2016, 2016, 7498359. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Albers, R.; Antoine, J.M.; Blum, S.; Bourdet-Sicard, R.; Ferns, G.A.; Folkerts, G.; Friedmann, P.S.; Frost, G.S.; Guarner, F.; et al. Inflammatory disease processes and interactions with nutrition. Br. J. Nutr. 2009, 101, S1–S45. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Kryston, T.B.; Georgiev, A.B.; Pissis, P.; Georgakilas, A.G. Role of oxidative stress and DNA damage in human carcinogenesis. Mutat. Res. Fund. Mol. Mech. Mut. 2011, 711, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.C. Antimicrobial reactive oxygen and nitrogen species: Concepts and controversies. Nat. Rev. Microbiol. 2004, 2, 820–832. [Google Scholar] [CrossRef] [PubMed]
- Bøhn, S.K.; Myhrstad, M.C.; Thoresen, M.; Holden, M.; Karlsen, A.; Tunheim, S.H.; Erlund, I.; Svendsen, M.; Seljeflot, I.; Moskaug, J.Ø.; et al. Blood cell gene expression associated with cellular stress defense is modulated by antioxidant-rich food in a randomised controlled clinical trial of male smokers. BMC Med. 2010, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Krishnaiah, D.; Sarbatly, R.; Nithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233. [Google Scholar] [CrossRef]
- Lee, J.; Koo, N.; Min, D.B. Reactive oxygen species, aging, and antioxidative nutraceuticals. Compr. Rev. Food Sci. Food Saf. 2004, 3, 21–33. [Google Scholar] [CrossRef]
- Seifried, H.E.; Anderson, D.E.; Fisher, E.I.; Milner, J.A. A review of the interaction among dietary antioxidants and reactive oxygen species. J. Nutr. Biochem. 2007, 18, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox. Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Lotito, S.B.; Frei, B. Consumption of flavonoid-rich foods and increased plasma antioxidant capacity in humans: Cause, consequence, or epiphenomenon? Free Radic. Biol. Med. 2006, 41, 1727–1746. [Google Scholar] [CrossRef] [PubMed]
- Kong, A.N.; Yu, R.; Lei, W.; Mandlekar, S.; Tan, T.H.; Ucker, D.S. Differential activation of MAPK and ICE/CED-3 protease in chemical-induced apoptosis. The role of oxidative stress in the regulation of mitogen-activated protein kinases (MAPKs) leading to gene expression and survival or activation of caspases leading to apoptosis. Restor. Neurol. Neurosci. 1998, 12, 63–70. [Google Scholar] [PubMed]
- Kong, A.N.; Owuor, E.; Yu, R.; Hebbar, V.; Chen, C.; Hu, R.; Mandlekar, S. Induction of xenobiotic enzymes by the MAP kinase pathway and the antioxidant or electrophile response element (ARE/EpRE). Drug Metab. Rev. 2001, 33, 255–271. [Google Scholar] [CrossRef] [PubMed]
- Finley, J.W.; Kong, A.N.; Hintze, K.J.; Jeffery, E.H.; Ji, L.L.; Lei, X.G. Antioxidants in foods: State of the science important to the food industry. J. Agric. Food Chem. 2011, 59, 6837–6846. [Google Scholar] [CrossRef] [PubMed]
- Drew, J.E. Cellular defense system gene expression profiling of human whole blood: Opportunities to predict health benefits in response to diet. Adv. Nutr. 2012, 3, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.L.; Huang, J.Y.; Shyur, L.F. Phytoagents for cancer management: Regulation of nucleic acid oxidation, ROS, and related mechanisms. Oxid. Med. Cell. Longev. 2013, 2013, 925804. [Google Scholar] [CrossRef] [PubMed]
- Manjunatha, H.; Srinivasan, K. Protective effect of dietary curcumin and capsaicin on induced oxidation of low-density lipoprotein, iron-induced hepatotoxicity and carrageenan-induced inflammation in experimental rats. FEBS J. 2006, 273, 4528–4537. [Google Scholar] [CrossRef] [PubMed]
- Srihari, T.; Sengottuvelan, M.; Nalini, N. Dose-dependent effect of oregano (origanum vulgare L.) on lipid peroxidation and antioxidant status in 1,2-dimethylhydrazine-induced rat colon carcinogenesis. J. Pharm. Pharmacol. 2008, 60, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Ku, C.S.; Pham, T.X.; Park, Y.; Martin, D.A.; Xie, L.; Taheri, R.; Lee, J.; Bolling, B.W. Aronia melanocarpa (chokeberry) polyphenol-rich extract improves antioxidant function and reduces total plasma cholesterol in apolipoprotein E knockout mice. Nutr. Res. 2013, 33, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.A.; Afifi, M.; Abdelazim, A.M.; Mosleh, Y.Y. Quercetin and omega 3 ameliorate oxidative stress induced by aluminium chloride in the brain. J. Mol. Neurosci. 2014, 53, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Gourineni, V.; Shay, N.F.; Chung, S.; Sandhu, A.K.; Gu, L. Muscadine grape (vitis rotundifolia) and wine phytochemicals prevented obesity-associated metabolic complications in C57BL/6J mice. J. Agric. Food Chem. 2012, 60, 7674–7681. [Google Scholar] [CrossRef] [PubMed]
- Belviranli, M.; Gokbel, H.; Okudan, N.; Basarali, K. Effects of grape seed extract supplementation on exercise-induced oxidative stress in rats. Br. J. Nutr. 2012, 108, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Natella, F.; Belelli, F.; Gentili, V.; Ursini, F.; Scaccini, C. Grape seed proanthocyanidins prevent plasma postprandial oxidative stress in humans. J. Agric. Food Chem. 2002, 50, 7720–7725. [Google Scholar] [CrossRef] [PubMed]
- Serafini, M.; Bugianesi, R.; Maiani, G.; Valtuena, S.; De Santis, S.; Crozier, A. Plasma antioxidants from chocolate. Nature 2003, 424, 1013. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.B.; Kyle, J.; Jenkinson, A.M.; Gardner, P.T.; McPhail, D.B.; Duthie, G.G. Effects of blueberry and cranberry juice consumption on the plasma antioxidant capacity of healthy female volunteers. Eur. J. Clin. Nutr. 2000, 54, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Nah, J.; Chun, S.; Park, H.; Yang, S.E.; Min, W.K. In vivo antioxidant effect of green tea. Eur. J. Clin. Nutr. 2000, 54, 527–529. [Google Scholar] [CrossRef] [PubMed]
- Guarrera, S.; Sacerdote, C.; Fiorini, L.; Marsala, R.; Polidoro, S.; Gamberini, S.; Saletta, F.; Malaveille, C.; Talaska, G.; Vineis, P.; et al. Expression of DNA repair and metabolic genes in response to a flavonoid-rich diet. Br. J. Nutr. 2007, 98, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Siegmund, K.; Tseng, C.C.; Lee, A.S.; Wu, A.H. Soy food supplementation, dietary fat reduction and peripheral blood gene expression in postmenopausal women—A randomized, controlled trial. Mol. Nutr. Food Res. 2011, 55, S264–S277. [Google Scholar] [CrossRef] [PubMed]
- Bakker, G.C.; van Erk, M.J.; Pellis, L.; Wopereis, S.; Rubingh, C.M.; Cnubben, N.H.; Kooistra, T.; van Ommen, B.; Hendriks, H.F. An antiinflammatory dietary mix modulates inflammation and oxidative and metabolic stress in overweight men: A nutrigenomics approach. Am. J. Clin. Nutr. 2010, 91, 1044–1059. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef] [PubMed]
- Long, L.H.; Clement, M.V.; Halliwell, B. Artifacts in cell culture: Rapid generation of hydrogen peroxide on addition of (−)-epigallocatechin, (−)-epigallocatechin gallate, (+)-catechin, and quercetin to commonly used cell culture media. Biochem. Biophys. Res. Commun. 2000, 273, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Chai, P.C.; Long, L.H.; Halliwell, B. Contribution of hydrogen peroxide to the cytotoxicity of green tea and red wines. Biochem. Biophys. Res. Commun. 2003, 304, 650–654. [Google Scholar] [CrossRef]
- Long, L.H.; Hoi, A.; Halliwell, B. Instability of, and generation of hydrogen peroxide by, phenolic compounds in cell culture media. Arch. Biochem. Biophys. 2010, 501, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Rodd, A.L.; Ververis, K.; Sayakkarage, D.; Khan, A.W.; Rafehi, H.; Ziemann, M.; Loveridge, S.J.; Lazarus, R.; Kerr, C.; Lockett, T.; et al. RNA sequencing supports distinct reactive oxygen species-mediated pathways of apoptosis by high and low size mass fractions of bay leaf (Lauris nobilis) in HT-29 cells. Food Funct. 2015, 6, 2507–2524. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y. Challenges in providing credible scientific evidence of health benefits of dietary polyphenols. J. Funct. Foods 2013, 5, 524–526. [Google Scholar] [CrossRef]
- Holst, B.; Williamson, G. Nutrients and phytochemicals: From bioavailability to bioefficacy beyond antioxidants. Curr. Opin. Biotechnol. 2008, 19, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Selby-Pham, S.N.B.; Miller, R.B.; Howell, K.; Dunshea, F.; Bennett, L.E. Physicochemical properties of dietary phytochemicals can predict their passive absorption in the human small intestine. Sci. Rep. 2017, 7, 1931. [Google Scholar] [CrossRef] [PubMed]
- Selby-Pham, S.N.B.; Howell, K.S.; Dunshea, F.R.; Ludbey, J.; Lutz, A.; Bennett, L.E. High throughput prediction of human absorption kinetics of plant extracts using LC-MS and statistical modelling. Food Chem. 2017, in press. [Google Scholar]
- Roura, E.; Koopmans, S.J.; Lalles, J.P.; Le Huerou-Luron, I.; de Jager, N.; Schuurman, T.; Val-Laillet, D. Critical review evaluating the pig as a model for human nutritional physiology. Nutr. Res. Rev. 2016, 29, 60–90. [Google Scholar] [CrossRef] [PubMed]
- Flamini, R.; Mattivi, F.; De Rosso, M.; Arapitsas, P.; Bavaresco, L. Advanced knowledge of three important classes of grape phenolics: Anthocyanins, stilbenes and flavonols. Int. J. Mol. Sci. 2013, 14, 19651–19669. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.E.; Terschluesen, A.M.; Rimbach, G. Health promoting effects of brassica-derived phytochemicals: From chemopreventive and anti-inflammatory activities to epigenetic regulation. Oxid. Med. Cell. Longev. 2013, 2013, 964539. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Jiang, X.; Xiao, Z.; Yu, L.; Pham, Q.; Sun, J.; Chen, P.; Yokoyama, W.; Yu, L.L.; Luo, Y.S.; et al. Red cabbage microgreens lower circulating low-density lipoprotein (LDL), liver cholesterol, and inflammatory cytokines in mice fed a high-fat diet. J. Agric. Food Chem. 2016, 64, 9161–9171. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using folin-ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Ostrowska, E.; Cross, R.F.; Muralitharan, M.; Bauman, D.E.; Dunshea, F.R. Effects of dietary fat and conjugated linoleic acid on plasma metabolite concentrations and metabolic responses to homeostatic signals in pigs. Br. J. Nutr. 2002, 88, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Asseburg, H.; Schafer, C.; Muller, M.; Hagl, S.; Pohland, M.; Berressem, D.; Borchiellini, M.; Plank, C.; Eckert, G.P. Effects of grape skin extract on age-related mitochondrial dysfunction, memory and life span in C57BL/6J mice. Neuromol. Med. 2016, 18, 378–395. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.R.; Pires, K.M.; Nalbones-Barbosa, M.N.; Dos Santos Valenca, S.; Resende, A.C.; de Moura, R.S. Grape skin extract-derived polyphenols modify programming-induced renal endowment in prenatal protein-restricted male mouse offspring. Eur. J. Nutr. 2016, 55, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Resende, A.C.; Emiliano, A.F.; Cordeiro, V.S.; de Bem, G.F.; de Cavalho, L.C.; de Oliveira, P.R.; Neto, M.L.; Costa, C.A.; Boaventura, G.T.; de Moura, R.S. Grape skin extract protects against programmed changes in the adult rat offspring caused by maternal high-fat diet during lactation. J. Nutr. Biochem. 2013, 24, 2119–2126. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Marques, S.; Magalhães, L.; Tóth, I.; Segundo, M. Insights on antioxidant assays for biological samples based on the reduction of copper complexes—The importance of analytical conditions. Int. J. Mol. Sci. 2014, 15, 11387–11402. [Google Scholar] [CrossRef] [PubMed]
- Wolff, S.P. Ferrous ion oxidation in presence of ferric ion indicator xylenol orange for measurement of hydroperoxides. Methods Enzymol. 1994, 233, 182–189. [Google Scholar]
- Wolfensohn, S.; Lloyd, M. The larger domestic species. In Handbook of Laboratory Animal Management and Welfare, 3rd ed.; Wolfensohn, S., Lloyd, M., Eds.; Blackwell Publishing: Ames, IA, USA, 2003; pp. 326–364. [Google Scholar]
- Apak, R.; Guclu, K.; Ozyurek, M.; Karademir, S.E.; Altun, M. Total antioxidant capacity assay of human serum using copper(II)-neocuproine as chromogenic oxidant: The CUPRAC method. Free Radic. Res. 2005, 39, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; McLellan, L.I. Glutathione and glutathione-dependent enzymes represent a co-ordinately regulated defence against oxidative stress. Free Radic. Res. 1999, 31, 273–300. [Google Scholar] [CrossRef] [PubMed]
- Leenen, R.; Roodenburg, A.J.; Tijburg, L.B.; Wiseman, S.A. A single dose of tea with or without milk increases plasma antioxidant activity in humans. Eur. J. Clin. Nutr. 2000, 54, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Rein, D.; Lotito, S.; Holt, R.R.; Keen, C.L.; Schmitz, H.H.; Fraga, C.G. Epicatechin in human plasma: In vivo determination and effect of chocolate consumption on plasma oxidation status. J. Nutr. 2000, 130, 2109s–2114s. [Google Scholar] [PubMed]
- Michiels, J.; Missotten, J.; Dierick, N.; Fremaut, D.; Maene, P.; De Smet, S. In vitro degradation and in vivo passage kinetics of carvacrol, thymol, eugenol and trans-cinnamaldehyde along the gastrointestinal tract of piglets. J. Sci. Food Agric. 2008, 88, 2371–2381. [Google Scholar] [CrossRef]
- Everette, J.D.; Bryant, Q.M.; Green, A.M.; Abbey, Y.A.; Wangila, G.W.; Walker, R.B. Thorough study of reactivity of various compound classes toward the folin−ciocalteu reagent. J. Agric. Food Chem. 2010, 58, 8139–8144. [Google Scholar] [CrossRef] [PubMed]
- Singha, I.; Das, S.K. Free radical scavenging properties of skin and pulp extracts of different grape cultivars in vitro and attentuation of H2O2-induced oxidative stress in liver tissue ex vivo. Ind. J. Clin. Biochem. 2015, 30, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Gabas, A.L.; Telis-Romero, J.; Menegalli, F.C. Thermodynamic models for water sorption by grape skin and pulp. Drying Technol. 1999, 17, 962–974. [Google Scholar] [CrossRef]
- Singh, J.; Upadhyay, A.K.; Bahadur, A.; Singh, K.P.; Rai, M. Antioxidant phytochemicals in cabbage (Brassica oleracea L. Var. Capitata). Sci. Hort. 2006, 108, 233–237. [Google Scholar] [CrossRef]
- Sivakumaran, S.; Huffman, L.; Sivakumaran, S. The Concise New Zealand Food Composition Tables, 12th ed.; The New Zealand Institute for Plant & Food Research Limited and Ministry of Health: Palmerston North, New Zealand, 2017. [Google Scholar]
- Zeng, C. Effects of different cooking methods on the vitamin c content of selected vegetables. Nutr. Food Sci. 2013, 43, 438–443. [Google Scholar] [CrossRef]
- Tiwari, B.K.; O’Donnell, C.P.O.; Patras, A.; Cullen, P.J. Anthocyanin and ascorbic acid degradation in sonicated strawberry juice. J. Agric. Food Chem. 2008, 56, 10071–10077. [Google Scholar] [CrossRef] [PubMed]
- Alleva, R.; Di Donato, F.; Strafella, E.; Staffolani, S.; Nocchi, L.; Borghi, B.; Pignotti, E.; Santarelli, L.; Tomasetti, M. Effect of ascorbic acid-rich diet on in vivo-induced oxidative stress. Br. J. Nutr. 2012, 107, 1645–1654. [Google Scholar] [CrossRef] [PubMed]
- Cherubini, A.; Beal, M.F.; Frei, B. Black tea increases the resistance of human plasma to lipid peroxidation in vitro, but not ex vivo. Free Radic. Biol. Med. 1999, 27, 381–387. [Google Scholar] [CrossRef]
- Lotito, S.B.; Frei, B. Relevance of apple polyphenols as antioxidants in human plasma: Contrasting in vitro and in vivo effects. Free Radic. Biol. Med. 2004, 36, 201–211. [Google Scholar] [PubMed]
- Kolodziejczyk-Czepas, J.; Nowak, P.; Moniuszko-Szajwaj, B.; Kowalska, I.; Stochmal, A. Free radical scavenging actions of three trifolium species in the protection of blood plasma antioxidant capacity in vitro. Pharm. Biol. 2015, 53, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Sahu, S.C.; Gray, G.C. Interactions of flavonoids, trace metals, and oxygen: Nuclear DNA damage and lipid peroxidation induced by myricetin. Cancer Lett. 1993, 70, 73–79. [Google Scholar] [CrossRef]
- Zheng, L.F.; Wei, Q.Y.; Cai, Y.J.; Fang, J.G.; Zhou, B.; Yang, L.; Liu, Z.L. DNA damage induced by resveratrol and its synthetic analogues in the presence of Cu (II) ions: Mechanism and structure-activity relationship. Free Radic. Biol. Med. 2006, 41, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, L.R.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.S.; Fazal, F.; Rahman, A.; Hadi, S.M.; Parish, J.H. Activities of flavonoids for the cleavage of DNA in the presence of Cu(II): Correlation with generation of active oxygen species. Carcinogenesis 1992, 13, 605–608. [Google Scholar] [CrossRef]
- Tautkus, S.; Irnius, A.; Speiciene, D.; Barkauskas, J.; Kareiva, A. Investigation of distribution of heavy metals between blood plasma and blood cells. Ann. Chim. 2007, 97, 1139–1142. [Google Scholar] [CrossRef]
- Sandstrom, B.E.; Marklund, S.L. Effects of variation in glutathione peroxidase activity on DNA damage and cell survival in human cells exposed to hydrogen peroxide and t-butyl hydroperoxide. Biochem. J. 1990, 271, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Ajila, C.M.; Prasada Rao, U.J. Protection against hydrogen peroxide induced oxidative damage in rat erythrocytes by Mangifera indica L. Peel extract. Food Chem. Toxicol. 2008, 46, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Shui, G.; Bao, Y.M.; Bo, J.; An, L.J. Protective effect of protocatechuic acid from alpinia oxyphylla on hydrogen peroxide-induced oxidative PC12 cell death. Eur. J. Pharmacol. 2006, 538, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Clement, M.V.; Long, L.H. Hydrogen peroxide in the human body. FEBS Lett. 2000, 486, 10–13. [Google Scholar] [CrossRef]
- Ohguro, N.; Fukuda, M.; Sasabe, T.; Tano, Y. Concentration dependent effects of hydrogen peroxide on lens epithelial cells. Br. J. Ophthalmol. 1999, 83, 1064. [Google Scholar] [CrossRef] [PubMed]
- Loo, A.E.; Halliwell, B. Effects of hydrogen peroxide in a keratinocyte-fibroblast co-culture model of wound healing. Biochem. Biophys. Res. Commun. 2012, 423, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Loo, A.E.; Wong, Y.T.; Ho, R.; Wasser, M.; Du, T.; Ng, W.T.; Halliwell, B. Effects of hydrogen peroxide on wound healing in mice in relation to oxidative damage. PLoS ONE 2012, 7, e49215. [Google Scholar] [CrossRef] [PubMed]
- Radak, Z.; Chung, H.Y.; Goto, S. Exercise and hormesis: Oxidative stress-related adaptation for successful aging. Biogerontology 2005, 6, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Pallauf, K.; Giller, K.; Huebbe, P.; Rimbach, G. Nutrition and healthy ageing: Calorie restriction or polyphenol-rich “mediterrasian” diet? Oxid. Med. Cell Longev. 2013, 2013, 707421. [Google Scholar] [CrossRef] [PubMed]
- Covas, G.; Marinho, H.S.; Cyrne, L.; Antunes, F. Activation of Nrf2 by H2O2: De novo synthesis versus nuclear translocation. Methods Enzymol. 2013, 528, 157–171. [Google Scholar] [PubMed]
- Surh, Y.J.; Kundu, J.K.; Na, H.K. Nrf2 as a master redox switch in turning on the cellular signaling involved in the induction of cytoprotective genes by some chemopreventive phytochemicals. Planta Med. 2008, 74, 1526–1539. [Google Scholar] [CrossRef] [PubMed]
- Hybertson, B.M.; Gao, B.; Bose, S.K.; McCord, J.M. Oxidative stress in health and disease: The therapeutic potential of Nrf2 activation. Mol. Aspects Med. 2011, 32, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Farombi, E.O.; Shrotriya, S.; Na, H.K.; Kim, S.H.; Surh, Y.J. Curcumin attenuates dimethylnitrosamine-induced liver injury in rats through Nrf2-mediated induction of heme oxygenase-1. Food Chem. Toxicol. 2008, 46, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.H.; Li, Y.Q.; Yang, X.Y. Inhibition of epigallocatechin gallate on orthotopic colon cancer by upregulating the Nrf2-ugt1a signal pathway in nude mice. Pharmacology 2007, 80, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Ben-Dor, A.; Steiner, M.; Gheber, L.; Danilenko, M.; Dubi, N.; Linnewiel, K.; Zick, A.; Sharoni, Y.; Levy, J. Carotenoids activate the antioxidant response element transcription system. Mol. Cancer Ther. 2005, 4, 177–186. [Google Scholar] [PubMed]
- Kode, A.; Rajendrasozhan, S.; Caito, S.; Yang, S.R.; Megson, I.L.; Rahman, I. Resveratrol induces glutathione synthesis by activation of Nrf2 and protects against cigarette smoke-mediated oxidative stress in human lung epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, L478–L488. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Freeman, M.L.; Liebler, D.C. Identification of sensor cysteines in human keap1 modified by the cancer chemopreventive agent sulforaphane. Chem. Res. Toxicol. 2005, 18, 1917–1926. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemical Sources | Plasma TEAC | Plasma H2O2 | ||
|---|---|---|---|---|
| Yield (nmol/µmol GAE) * | Linear Fit R2 | Yield (nmol/µmol GAE) * | Linear Fit R2 | |
| Gallic acid standard | 1606.3 ± 98.1 | 0.99 | 68.7 ± 4.5 | 0.97 |
| Red cabbage extract | 633.2 ± 74.7 | 0.96 | 22.4 ± 1.1 | 0.99 |
| Grape skin extract | 1077.8 ± 120.4 | 0.96 | 44.2 ± 2.1 | 0.99 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selby-Pham, S.N.B.; Cottrell, J.J.; Dunshea, F.R.; Ng, K.; Bennett, L.E.; Howell, K.S. Dietary Phytochemicals Promote Health by Enhancing Antioxidant Defence in a Pig Model. Nutrients 2017, 9, 758. https://doi.org/10.3390/nu9070758
Selby-Pham SNB, Cottrell JJ, Dunshea FR, Ng K, Bennett LE, Howell KS. Dietary Phytochemicals Promote Health by Enhancing Antioxidant Defence in a Pig Model. Nutrients. 2017; 9(7):758. https://doi.org/10.3390/nu9070758
Chicago/Turabian StyleSelby-Pham, Sophie N. B., Jeremy J. Cottrell, Frank R. Dunshea, Ken Ng, Louise E. Bennett, and Kate S. Howell. 2017. "Dietary Phytochemicals Promote Health by Enhancing Antioxidant Defence in a Pig Model" Nutrients 9, no. 7: 758. https://doi.org/10.3390/nu9070758
APA StyleSelby-Pham, S. N. B., Cottrell, J. J., Dunshea, F. R., Ng, K., Bennett, L. E., & Howell, K. S. (2017). Dietary Phytochemicals Promote Health by Enhancing Antioxidant Defence in a Pig Model. Nutrients, 9(7), 758. https://doi.org/10.3390/nu9070758