Differing Roles of Hyaluronan Molecular Weight on Cancer Cell Behavior and Chemotherapy Resistance


Abstract
1. Introduction
2. Hyaluronan
2.1. HA Synthesis, Degradation and Conformation
2.2. HA in the Tumor Microenvironment
3. Role of Molecular Weight of Hyaluronan in Normal Biology
4. Role of Molecular Weight of Hyaluronan in Cancer Biology
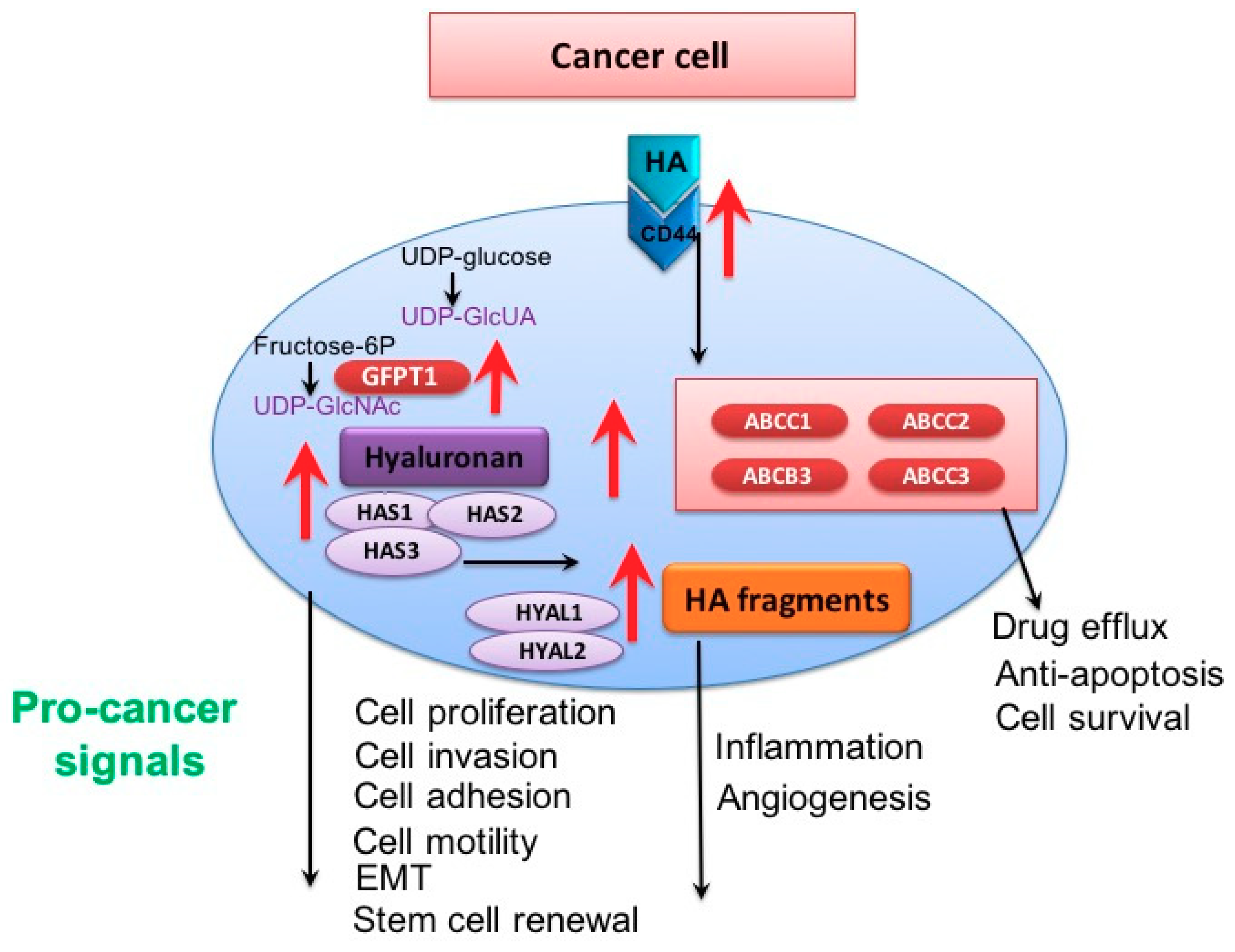
4.1. Cancer Stem Cells, Hyaluronan and Therapy Resistance
4.2. Hyaluronan, ABC Transporters and Therapy Resistance
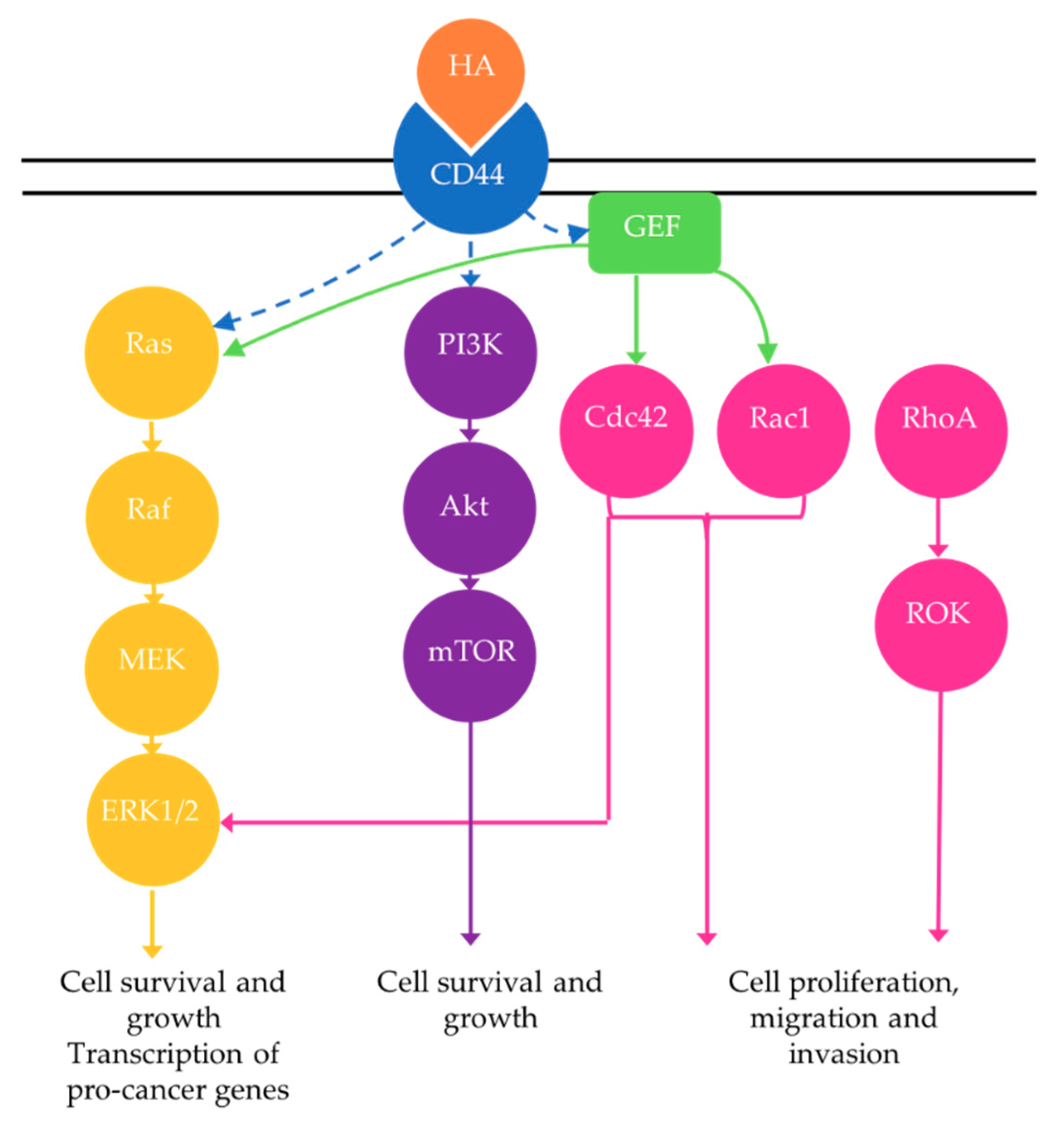
4.3. Hyaluronan, Receptor Tyrosine Kinase Pathways and Therapy Resistance
4.3.1. Rho GTPase Signaling
4.3.2. Phosphoinositide 3-Kinase (PI3K) Pathway
4.3.3. MAPK Pathway
5. Targeting HA in Cancer
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef]
- Longley, D.B.; Johnston, P.G. Molecular mechanisms of drug resistance. J. Pathol. 2005, 205, 275–292. [Google Scholar] [CrossRef]
- Tsuruo, T.; Naito, M.; Tomida, A.; Fujita, N.; Mashima, T.; Sakamoto, H.; Haga, N. Molecular targeting therapy of cancer: Drug resistance, apoptosis and survival signal. Cancer Sci. 2003, 94, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, L.; de Sousa e Melo, F.; Richel, D.J.; Medema, J.P. The developing cancer stem-cell model: Clinical challenges and opportunities. Lancet Oncol. 2012, 13, e83–89. [Google Scholar] [CrossRef]
- Chanmee, T.; Ontong, P.; Kimata, K.; Itano, N. Key roles of hyaluronan and its cd44 receptor in the stemness and survival of cancer stem cells. Front. Oncol. 2015, 5, e180. [Google Scholar] [CrossRef] [PubMed]
- Toole, B.P.; Slomiany, M.G. Hyaluronan: A constitutive regulator of chemoresistance and malignancy in cancer cells. Semin. Cancer Biol. 2008, 18, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Toole, B.P. Hyaluronan: From extracellular glue to pericellular cue. Nat. Rev. Cancer 2004, 4, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Ricciardelli, C.; Ween, M.P.; Lokman, N.A.; Tan, I.A.; Pyragius, C.E.; Oehler, M.K. Chemotherapy-induced hyaluronan production: A novel chemoresistance mechanism in ovarian cancer. BMC Cancer 2013, 13, 476. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.; Palmer, J.W. The polysaccharide of the vitreous humor. J. Biol. Chem. 1934, 107, 629–634. [Google Scholar]
- Dianhua, J.; Jiurong, L.; Paul, W.N. Hyaluronan as an immune regulator in human diseases. Physiol. Rev. 2011, 91, 221–264. [Google Scholar]
- Vigetti, D.; Viola, M.; Karousou, E.; De Luca, G.; Passi, A. Metabolic control of hyaluronan synthases. Matrix Biol. 2014, 35, 8–13. [Google Scholar] [CrossRef]
- Itano, N.; Sawai, T.; Yoshida, M.; Lenas, P.; Yamada, Y.; Imagawa, M.; Shinomura, T.; Hamaguchi, M.; Yoshida, Y.; Ohnuki, Y.; et al. Three isoforms of mammalian hyaluronan synthases have distinct enzymatic properties. J. Biol. Chem. 1999, 274, 25085–25092. [Google Scholar] [CrossRef]
- Maharjan, A.S.; Pilling, D.; Gomer, R.H. High and low molecular weight hyaluronic acid differentially regulate human fibrocyte differentiation. PLoS ONE 2011, e6. [Google Scholar] [CrossRef]
- Stern, R. Devising a pathway for hyaluronan catabolism: Are we there yet? Glycobiology 2003, 13, 105R–115R. [Google Scholar] [CrossRef]
- Erickson, M.; Stern, R. Chain gangs: New aspects of hyaluronan metabolism. Biochem. Res. Int. 2012, 2012, e893947. [Google Scholar] [CrossRef]
- Bohaumilitzky, L.; Huber, A.-K.; Stork, E.M.; Wengert, S.; Woelfl, F.; Boehm, H. A trickster in disguise: Hyaluronan’s ambivalent roles in the matrix. Front. Oncol. 2017, 7, 242. [Google Scholar] [CrossRef]
- Tian, X.; Azpurua, J.; Hine, C.; Vaidya, A.; Myakishev-Rempel, M.; Ablaeva, J.; Mao, Z.; Nevo, E.; Gorbunova, V.; Seluanov, A. High-molecular-mass hyaluronan mediates the cancer resistance of the naked mole rat. Nature 2013, 499, 346–349. [Google Scholar] [CrossRef]
- Seluanov, A.; Hine, C.; Azpurua, J.; Feigenson, M.; Bozzella, M.; Mao, Z.; Catania, K.C.; Gorbunova, V. Hypersensitivity to contact inhibition provides a clue to cancer resistance of naked mole-rat. Proc. Natl. Acad. Sci. USA 2009, 106, 19352–19357. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, G.; Liu, H.; Han, J.; Liu, Z.; Ma, H. Molecular weight impact on the mechanical forces between hyaluronan and its receptor. Carbohydr. Polym. 2018, 197, 326–336. [Google Scholar] [CrossRef]
- Wolny, P.M.; Banerji, S.; Gounou, C.; Brisson, A.R.; Day, A.J.; Jackson, D.G.; Richter, R.P. Analysis of cd44-hyaluronan interactions in an artificial membrane system insight into the distinct binding properties of high and low molecular weight hyaluronan. J. Biol. Chem. 2010, 285, 30170–30180. [Google Scholar] [CrossRef]
- Lesley, J.; Hascall, V.C.; Tammi, M.; Hyman, R. Hyaluronan binding by cell surface cd44. J. Biol. Chem. 2000, 275, 26967–26975. [Google Scholar] [CrossRef]
- Yang, C.X.; Cao, M.L.; Liu, H.; He, Y.Q.; Xu, J.; Du, Y.; Liu, Y.W.; Wang, W.J.; Cui, L.; Hu, J.J.; et al. The high and low molecular weight forms of hyaluronan have distinct effects on cd44 clustering. J. Biol. Chem. 2012, 287, 43094–43107. [Google Scholar] [CrossRef]
- Weigel, P.H.; Baggenstoss, B.A. What is special about 200 kda hyaluronan that activates hyaluronan receptor signaling? Glycobiology 2017, 27, 868–877. [Google Scholar] [CrossRef]
- Kultti, A.; Li, X.; Jiang, P.; Thompson, C.B.; Frost, G.I.; Shepard, H.M. Therapeutic targeting of hyaluronan in the tumor stroma. Cancers (Basel) 2012, 4, 873–903. [Google Scholar] [CrossRef]
- McCarthy, J.B.; El-Ashry, D.; Turley, E.A. Hyaluronan, cancer-associated fibroblasts and the tumor microenvironment in malignant progression. Front. Cell Dev. Biol. 2018, 6, 48. [Google Scholar] [CrossRef]
- Evanko, S.P.; Potter-Perigo, S.; Petty, L.J.; Workman, G.A.; Wight, T.N. Hyaluronan controls the deposition of fibronectin and collagen and modulates tgf-beta1 induction of lung myofibroblasts. Matrix Biol. 2015, 42, 74–92. [Google Scholar] [CrossRef]
- Chanmee, T.; Ontong, P.; Itano, N. Hyaluronan: A modulator of the tumor microenvironment. Cancer Lett. 2016, 375, 20–30. [Google Scholar] [CrossRef]
- Monslow, J.; Govindaraju, P.; Pure, E. Hyaluronan—A functional and structural sweet spot in the tissue microenvironment. Front. Immunol. 2015, 6, e231. [Google Scholar] [CrossRef]
- Camenisch, T.D.; Spicer, A.P.; Brehm-Gibson, T.; Biesterfeldt, J.; Augustine, M.L.; Calabro, A.; Kubalak, S.; Klewer, S.E.; McDonald, J.A. Disruption of hyaluronan synthase-2 abrogates normal cardiac morphogenesis and hyaluronan-mediated transformation of epithelium to mesenchyme. J. Clin. Investig. 2000, 106, 349–360. [Google Scholar] [CrossRef]
- Auvinen, P.; Tammi, R.; Kosma, V.M.; Sironen, R.; Soini, Y.; Mannermaa, A.; Tumelius, R.; Uljas, E.; Tammi, M. Increased hyaluronan content and stromal cell cd44 associate with her2 positivity and poor prognosis in human breast cancer. Int. J. Cancer 2013, 132, 531–539. [Google Scholar] [CrossRef]
- Anttila, M.A.; Tammi, R.H.; Tammi, M.I.; Syrjanen, K.J.; Saarikoski, S.V.; Kosma, V.M. High levels of stromal hyaluronan predict poor disease outcome in epithelial ovarian cancer. Cancer Res. 2000, 60, 150–155. [Google Scholar]
- Lipponen, P.; Aaltomaa, S.; Tammi, R.; Tammi, M.; Agren, U.; Kosma, V.M. High stromal hyaluronan level is associated with poor differentiation and metastasis in prostate cancer. Eur. J. Cancer 2001, 37, 849–856. [Google Scholar] [CrossRef]
- Knudson, W.; Biswas, C.; Toole, B.P. Interactions between human-tumor cells and fibroblasts stimulate hyaluronate synthesis. Proc. Natl. Acad. Sci. Biol. 1984, 81, 6767–6771. [Google Scholar] [CrossRef]
- Asplund, T.; Versnel, M.A.; Laurent, T.C.; Heldin, P. Human mesothelioma cells produce factors that stimulate the production of hyaluronan by mesothelial cells and fibroblasts. Cancer Res. 1993, 53, 388–392. [Google Scholar]
- Auvinen, P.; Rilla, K.; Tumelius, R.; Tammi, M.; Sironen, R.; Soini, Y.; Kosma, V.M.; Mannermaa, A.; Viikari, J.; Tammi, R. Hyaluronan synthases (has1-3) in stromal and malignant cells correlate with breast cancer grade and predict patient survival. Breast Cancer Res. Treat. 2014, 143, 277–286. [Google Scholar] [CrossRef]
- Wang, S.J.; Earle, C.; Wong, G.; Bourguignon, L.Y. Role of hyaluronan synthase 2 to promote cd44-dependent oral cavity squamous cell carcinoma progression. Head Neck 2013, 35, 511–520. [Google Scholar] [CrossRef]
- McAtee, C.O.; Barycki, J.J.; Simpson, M.A. Emerging roles for hyaluronidase in cancer metastasis and therapy. Adv. Cancer Res. 2014, 123, 1–34. [Google Scholar]
- Heldin, P.; Lin, C.-Y.; Kolliopoulos, C.; Chen, Y.-H.; Skandalis, S.S. Regulation of hyaluronan biosynthesis and clinical impact of excessive hyaluronan production. Matrix Biol. 2018. [Google Scholar] [CrossRef]
- Nykopp, T.K.; Rilla, K.; Sironen, R.; Tammi, M.I.; Tammi, R.H.; Hamalainen, K.; Heikkinen, A.M.; Komulainen, M.; Kosma, V.M.; Anttila, M. Expression of hyaluronan synthases (has1-3) and hyaluronidases (hyal1-2) in serous ovarian carcinomas: Inverse correlation between hyal1 and hyaluronan content. BMC Cancer 2009, 9, e143. [Google Scholar] [CrossRef]
- Nakamura, K.; Yokohama, S.; Yoneda, M.; Okamoto, S.; Tamaki, Y.; Ito, T.; Okada, M.; Aso, K.; Makino, I. High, but not low, molecular weight hyaluronan prevents t-cell-mediated liver injury by reducing proinflammatory cytokines in mice. J. Gastroenterol. 2004, 39, 346–354. [Google Scholar] [CrossRef]
- Campo, G.M.; Avenoso, A.; Campo, S.; D’Ascola, A.; Nastasi, G.; Calatroni, A. Molecular size hyaluronan differently modulates toll-like receptor-4 in lps-induced inflammation in mouse chondrocytes. Biochimie 2010, 92, 204–215. [Google Scholar] [CrossRef]
- Albano, G.D.; Bonanno, A.; Cavalieri, L.; Ingrassia, E.; Di Sano, C.; Siena, L.; Riccobono, L.; Gagliardo, R.; Profita, M. Effect of high, medium, and low molecular weight hyaluronan on inflammation and oxidative stress in an in vitro model of human nasal epithelial cells. Eur. Respir. J. 2016, 48, e8727289. [Google Scholar]
- De la Motte, C.; Nigro, J.; Vasanji, A.; Rho, H.; Kessler, S.; Bandyopadhyay, S.; Danese, S.; Fiocchi, C.; Stern, R. Platelet-derived hyaluronidase 2 cleaves hyaluronan into fragments that trigger monocyte-mediated production of proinflammatory cytokines. Am. J. Pathol. 2009, 174, 2254–2264. [Google Scholar] [CrossRef]
- Dong, Y.; Arif, A.; Olsson, M.; Cali, V.; Hardman, B.; Dosanjh, M.; Lauer, M.; Midura, R.J.; Hascall, V.C.; Brown, K.L.; et al. Endotoxin free hyaluronan and hyaluronan fragments do not stimulate tnf-α, interleukin-12 or upregulate co-stimulatory molecules in dendritic cells or macrophages. Sci. Rep.-UK 2016, 6, 36928. [Google Scholar] [CrossRef]
- Krejcova, D.; Pekarova, M.; Safrankova, B.; Kubala, L. The effect of different molecular weight hyaluronan on macrophage physiology. Neuro Endocrinol. Lett. 2009, 30 (Suppl. 1), 106–111. [Google Scholar]
- Ebid, R.; Lichtnekert, J.; Anders, H.-J. Hyaluronan is not a ligand but a regulator of toll-like receptor signaling in mesangial cells: Role of extracellular matrix in innate immunity. ISRN Nephrol. 2014, 2014, 714081. [Google Scholar] [CrossRef]
- Shiedlin, A.; Bigelow, R.; Christopher, W.; Arbabi, S.; Yang, L.; Maier, R.V.; Wainwright, N.; Childs, A.; Miller, R.J. Evaluation of hyaluronan from different sources: Streptococcus zooepidemicus, rooster comb, bovine vitreous, and human umbilical cord. Biomacromolecules 2004, 5, 2122–2127. [Google Scholar] [CrossRef]
- Rayahin, J.E.; Buhrman, J.S.; Zhang, Y.; Koh, T.J.; Gemeinhart, R.A. High and low molecular weight hyaluronic acid differentially influence macrophage activation. ACS Biomater. Sci. Eng. 2015, 1, 481–493. [Google Scholar] [CrossRef]
- Hodge-Dufour, J.; Noble, P.W.; Horton, M.R.; Bao, C.; Wysoka, M.; Burdick, M.D.; Strieter, R.M.; Trinchieri, G.; Puré, E. Induction of il-12 and chemokines by hyaluronan requires adhesion-dependent priming of resident but not elicited macrophages. J. Immunol. 1997, 159, 2492–2500. [Google Scholar]
- Pandey, M.S.; Baggenstoss, B.A.; Washburn, J.; Harris, E.N.; Weigel, P.H. The hyaluronan receptor for endocytosis (hare) activates nf-κb-mediated gene expression in response to 40-400-kda, but not smaller or larger, hyaluronans. J. Boil. Chem. 2013, 288, 14068–14079. [Google Scholar] [CrossRef]
- Scheibner, K.A.; Lutz, M.A.; Boodoo, S.; Fenton, M.J.; Powell, J.D.; Horton, M.R. Hyaluronan fragments act as an endogenous danger signal by engaging tlr2. J. Immunol. 2006, 177, 1272–1281. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Leu, S.-W.; Shi, L.; Dedaj, R.; Zhao, G.; Garg, H.G.; Shen, L.; Lien, E.; Fitzgerald, K.A.; Shiedlin, A.; et al. Tlr4 is a negative regulator in noninfectious lung inflammation. J. Immunol. 2010, 184, 5308–5314. [Google Scholar] [CrossRef]
- West, D.C.; Kumar, S. The effect of hyaluronate and its oligosaccharides on endothelial cell proliferation and monolayer integrity. Exp. Cell Res. 1989, 183, 179–196. [Google Scholar] [CrossRef]
- Slevin, M.; Kumar, S.; Gaffney, J. Angiogenic oligosaccharides of hyaluronan induce multiple signaling pathways affecting vascular endothelial cell mitogenic and wound healing responses. J. Biol. Chem. 2002, 277, 41046–41059. [Google Scholar] [CrossRef] [PubMed]
- Deed, R.; Rooney, P.; Kumar, P.; Norton, J.D.; Smith, J.; Freemont, A.J.; Kumar, S. Early-response gene signalling is induced by angiogenic oligosaccharides of hyaluronan in endothelial cells. Inhibition by non-angiogenic, high-molecular-weight hyaluronan. Int. J. Cancer 1998, 71, 251–256. [Google Scholar] [CrossRef]
- Gao, F.; Yang, C.X.; Mo, W.; Liu, Y.W.; He, Y.Q. Hyaluronan oligosaccharides are potential stimulators to angiogenesis via rhamm mediated signal pathway in wound healing. Clin. Investig. Med. 2008, 31, 106–116. [Google Scholar] [CrossRef]
- Wang, Y.; Han, G.; Guo, B.; Huang, J. Hyaluronan oligosaccharides promote diabetic wound healing by increasing angiogenesis. Pharmacol. Rep. 2016, 68, 1126–1132. [Google Scholar] [CrossRef]
- Tolg, C.; Telmer, P.; Turley, E. Specific sizes of hyaluronan oligosaccharides stimulate fibroblast migration and excisional wound repair. PLoS ONE 2014, 9, e88479. [Google Scholar] [CrossRef]
- Craig, E.A.; Parker, P.; Camenisch, T.D. Size-dependent regulation of snail2 by hyaluronan: Its role in cellular invasion. Glycobiology 2009, 19, 890–898. [Google Scholar] [CrossRef]
- Gouëffic, Y.; Guilluy, C.; Guérin, P.; Patra, P.; Pacaud, P.; Loirand, G. Hyaluronan induces vascular smooth muscle cell migration through rhamm-mediated pi3k-dependent rac activation. Cardiovasc. Res. 2006, 72, 339–348. [Google Scholar] [CrossRef]
- Zhu, R.; Huang, Y.H.; Tao, Y.; Wang, S.C.; Sun, C.; Piao, H.L.; Wang, X.Q.; Du, M.R.; Li, D.J. Hyaluronan up-regulates growth and invasion of trophoblasts in an autocrine manner via pi3k/akt and mapk/erk1/2 pathways in early human pregnancy. Placenta 2013, 34, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Tolg, C.; Yuan, H.; Flynn, S.M.; Basu, K.; Ma, J.; Tse, K.C.K.; Kowalska, B.; Vulkanesku, D.; Cowman, M.K.; McCarthy, J.B.; et al. Hyaluronan modulates growth factor induced mammary gland branching in a size dependent manner. Matrix Biol. 2017, 63, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Rosines, E.; Schmidt, H.J.; Nigam, S.K. The effect of hyaluronic acid size and concentration on branching morphogenesis and tubule differentiation in developing kidney culture systems: Potential applications to engineering of renal tissues. Biomaterials 2007, 28, 4806–4817. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Cao, M.; He, Y.; Liu, Y.; Yang, C.; Du, Y.; Wang, W.; Gao, F. A novel role of low molecular weight hyaluronan in breast cancer metastasis. FASEB J. 2015, 29, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.Y.; Zhu, H.; Zhou, B.; Diedrich, F.; Singleton, P.A.; Hung, M.C. Hyaluronan promotes cd44v3-vav2 interaction with grb2-p185(her2) and induces rac1 and ras signaling during ovarian tumor cell migration and growth. J. Biol. Chem. 2001, 276, 48679–48692. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.Y.; Gilad, E.; Rothman, K.; Peyrollier, K. Hyaluronan-cd44 interaction with iqgap1 promotes cdc42 and erk signaling, leading to actin binding, elk-1/estrogen receptor transcriptional activation, and ovarian cancer progression. J. Biol. Chem. 2005, 280, 11961–11972. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.Y.; Peyrollier, K.; Xia, W.; Gilad, E. Hyaluronan-cd44 interaction activates stem cell marker nanog, stat-3-mediated mdr1 gene expression, and ankyrin-regulated multidrug efflux in breast and ovarian tumor cells. J. Biol. Chem. 2008, 283, 17635–17651. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.Y.; Singleton, P.A.; Zhu, H.; Diedrich, F. Hyaluronan-mediated cd44 interaction with rhogef and rho kinase promotes grb2-associated binder-1 phosphorylation and phosphatidylinositol 3-kinase signaling leading to cytokine (macrophage-colony stimulating factor) production and breast tumor progression. J. Biol. Chem. 2003, 278, 29420–29434. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.Y.; Xia, W.; Wong, G. Hyaluronan-mediated cd44 interaction with p300 and sirt1 regulates beta-catenin signaling and nfkappab-specific transcription activity leading to mdr1 and bcl-xl gene expression and chemoresistance in breast tumor cells. J. Biol. Chem. 2009, 284, 2657–2671. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.Y.; Wong, G.; Earle, C.; Krueger, K.; Spevak, C.C. Hyaluronan-cd44 interaction promotes c-src-mediated twist signaling, microRNA-10b expression, and rhoa/rhoc up-regulation, leading to rho-kinase-associated cytoskeleton activation and breast tumor cell invasion. J. Biol. Chem. 2010, 285, 36721–36735. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.Y.W.; Wong, G.; Earle, C.A.; Xia, W. Interaction of low molecular weight hyaluronan with cd44 and toll-like receptors promotes the actin filament-associated protein 110-actin binding and myd88-nfκb signaling leading to proinflammatory cytokine/chemokine production and breast tumor invasion. Cytoskeleton 2011, 68, 671–693. [Google Scholar] [CrossRef]
- Zhao, Y.F.; Qiao, S.P.; Shi, S.L.; Yao, L.F.; Hou, X.L.; Li, C.F.; Lin, F.H.; Guo, K.; Acharya, A.; Chen, X.B.; et al. Modulating three-dimensional microenvironment with hyaluronan of different molecular weights alters breast cancer cell invasion behavior. ACS Appl. Mater. Interfaces 2017, 9, 9327–9338. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.Y.; Gilad, E.; Brightman, A.; Diedrich, F.; Singleton, P. Hyaluronan-cd44 interaction with leukemia-associated rhogef and epidermal growth factor receptor promotes rho/ras co-activation, phospholipase c epsilon-Ca2+ signaling, and cytoskeleton modification in head and neck squamous cell carcinoma cells. J. Biol. Chem. 2006, 281, 14026–14040. [Google Scholar] [CrossRef]
- Torre, C.; Wang, S.J.; Xia, W.; Bourguignon, L.Y. Reduction of hyaluronan-cd44-mediated growth, migration, and cisplatin resistance in head and neck cancer due to inhibition of rho kinase and pi-3 kinase signaling. Arch. Otolaryngol. Head Neck Surg. 2010, 136, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Shiina, M.; Bourguignon, L.Y. Selective activation of cancer stem cells by size-specific hyaluronan in head and neck cancer. Int. J. Cell Biol. 2015, 2015, e989070. [Google Scholar] [CrossRef]
- Bourguignon, L.Y.; Wong, G.; Shiina, M. Up-regulation of histone methyltransferase, dot1l, by matrix hyaluronan promotes microRNA-10 expression leading to tumor cell invasion and chemoresistance in cancer stem cells from head and neck squamous cell carcinoma. J. Biol. Chem. 2016, 291, 10571–10585. [Google Scholar] [CrossRef]
- Mascaro, M.; Pibuel, M.A.; Lompardia, S.L.; Diaz, M.; Zotta, E.; Bianconi, M.I.; Lago, N.; Otero, S.; Jankilevich, G.; Alvarez, E.; et al. Low molecular weight hyaluronan induces migration of human choriocarcinoma jeg-3 cells mediated by rhamm as well as by pi3k and mapk pathways. Histochem. Cell Biol. 2017, 148, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Lompardia, S.L.; Papademetrio, D.L.; Mascaro, M.; Alvarez, E.M.D.; Hajos, S.E. Human leukemic cell lines synthesize hyaluronan to avoid senescence and resist chemotherapy. Glycobiology 2013, 23, 1463–1476. [Google Scholar] [CrossRef]
- Nowell, P.C. The clonal evolution of tumor cell populations. Science 1976, 194, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Arya, R.K.; Maheshwari, S.; Singh, A.; Meena, S.; Pandey, P.; Dormond, O.; Datta, D. Tumor heterogeneity and cancer stem cell paradigm: Updates in concept, controversies and clinical relevance. Int. J. Cancer 2015, 136, 1991–2000. [Google Scholar] [CrossRef]
- Chanmee, T.; Ontong, P.; Mochizuki, N.; Kongtawelert, P.; Konno, K.; Itano, N. Excessive hyaluronan production promotes acquisition of cancer stem cell signatures through the coordinated regulation of twist and the transforming growth factor β (tgf-β)-snail signaling axis. J. Boil. Chem. 2014, 289, 26038–26056. [Google Scholar] [CrossRef]
- Vaidyanath, A.; Mahmud, H.B; Khayrani, A.C.; KoKo Oo, A.; Seno, A. Hyaluronic acid mediated enrichment of cd44 expressing glioblastoma stem cells in u251mg xenograft mouse model. J. Stem Cell Res. Ther. 2017, 7, e384. [Google Scholar] [CrossRef]
- Choudhary, M.; Zhang, X.; Stojkovic, P.; Hyslop, L.; Anyfantis, G.; Herbert, M.; Murdoch, A.P.; Stojkovic, M.; Lako, M. Putative role of hyaluronan and its related genes, has2 and rhamm, in human early preimplantation embryogenesis and embryonic stem cell characterization. Stem Cells 2007, 25, 3045–3057. [Google Scholar] [CrossRef]
- Ahmed, N.; Abubaker, K.; Findlay, J.K. Ovarian cancer stem cells: Molecular concepts and relevance as therapeutic targets. Mol. Asp. Med. 2014, 39, 110–125. [Google Scholar] [CrossRef]
- Zinzi, L.; Contino, M.; Cantore, M.; Capparelli, E.; Leopoldo, M.; Colabufo, N.A. ABC transporters in CSCs membranes as a novel target for treating tumor relapse. Front. Pharmacol. 2014, 5, 163. [Google Scholar] [CrossRef]
- Kryczek, I.; Liu, S.L.; Roh, M.; Vatan, L.; Szeliga, W.; Wei, S.; Banerjee, M.; Mao, Y.J.; Kotarski, J.; Wicha, M.S.; et al. Expression of aldehyde dehydrogenase and cd133 defines ovarian cancer stem cells. Int. J. Cancer 2012, 130, 29–39. [Google Scholar] [CrossRef]
- Patel, S.S.; Shah, K.A.; Shah, M.J.; Kothari, K.C.; Rawal, R.M. Cancer stem cells and stemness markers in oral squamous cell carcinomas. Asian Pac. J. Cancer Prev. 2014, 15, 8549–8556. [Google Scholar] [CrossRef]
- Mansoori, M.; Madjd, Z.; Janani, L.; Rasti, A. Circulating cancer stem cell markers in breast carcinomas: A systematic review protocol. Syst. Rev. 2017, 6, 262. [Google Scholar] [CrossRef]
- Liu, M.; Mor, G.; Cheng, H.; Xiang, X.; Hui, P.; Rutherford, T.; Yin, G.; Rimm, D.L.; Holmberg, J.; Alvero, A.; et al. High frequency of putative ovarian cancer stem cells with cd44/ck19 coexpression is associated with decreased progression-free intervals in patients with recurrent epithelial ovarian cancer. Reprod. Sci. 2013, 20, 605–615. [Google Scholar] [CrossRef]
- Meng, E.H.; Long, B.; Sullivan, P.; McClellan, S.; Finan, M.A.; Reed, E.; Shevde, L.; Rocconi, R.P. Cd44+/cd24-ovarian cancer cells demonstrate cancer stem cell properties and correlate to survival. Clin. Exp. Metastas 2012, 29, 939–948. [Google Scholar] [CrossRef]
- Fan, Z.N.; Li, M.X.; Chen, X.B.; Wang, J.; Liang, X.Y.; Wang, H.F.; Wang, Z.; Cheng, B.; Xia, J. Prognostic value of cancer stem cell markers in head and neck squamous cell carcinoma: A meta-analysis. Sci. Rep.-UK 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Seo, A.N.; Lee, H.J.; Kim, E.J.; Jang, M.H.; Kim, Y.J.; Kim, J.H.; Kim, S.W.; Ryu, H.S.; Park, I.A.; Im, S.A.; et al. Expression of breast cancer stem cell markers as predictors of prognosis and response to trastuzumab in her2-positive breast cancer. Br. J. Cancer 2016, 114, 1109–1116. [Google Scholar] [CrossRef]
- Horimoto, Y.; Arakawa, A.; Sasahara, N.; Tanabe, M.; Sai, S.; Himuro, T.; Saito, M. Combination of cancer stem cell markers cd44 and cd24 is superior to aldh1 as a prognostic indicator in breast cancer patients with distant metastases. PLoS ONE 2016, 11, e0165253. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.Y.; Sun, B.C. The prognostic role of the cancer stem cell marker aldehyde dehydrogenase 1 in head and neck squamous cell carcinomas: A meta-analysis. Oral. Oncol. 2014, 50, 1144–1148. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Q.; Zhou, J.D.; Lu, J.; Xiong, H.; Shi, X.L.; Gong, L. Significance of cd44 expression in head and neck cancer: A systemic review and meta-analysis. BMC Cancer 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, V.; Rezende, N.C.; Scotland, K.B.; Shaffer, S.M.; Persson, J.L.; Gudas, L.J.; Mongan, N.P. Regulation of stem cell pluripotency and differentiation involves a mutual regulatory circuit of the nanog, oct4, and sox2 pluripotency transcription factors with polycomb repressive complexes and stem cell microRNAs. Stem Cells Dev. 2009, 18, 1093–1108. [Google Scholar] [CrossRef]
- Bourguignon, L.Y.W.; Earle, C.; Shiina, M. Activation of matrix hyaluronan-mediated cd44 signaling, epigenetic regulation and chemoresistance in head and neck cancer stem cells. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Li, Y.; Cao, X.; Du, J.; Huang, X. Nanog regulates epithelial-mesenchymal transition and chemoresistance in ovarian cancer. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Hou, Y.; Huang, Z.; Cai, J.; Wang, Z. Sox2 is required to maintain cancer stem cells in ovarian cancer. Cancer Sci. 2017, 108, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Comisso, E.; Scarola, M.; Rosso, M.; Piazza, S.; Marzinotto, S.; Ciani, Y.; Orsaria, M.; Mariuzzi, L.; Schneider, C.; Schoeftner, S.; et al. Oct4 controls mitotic stability and inactivates the rb tumor suppressor pathway to enhance ovarian cancer aggressiveness. Oncogene 2017, 36, 4253–4266. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Liu, G.; Huang, B.; Sun, J.; Wu, D. Prognostic significance of sox2 in head and neck cancer: A meta-analysis. Int. J. Clin. Exp. Med. 2014, 7, 5010–5020. [Google Scholar] [PubMed]
- You, L.; Guo, X.; Huang, Y. Correlation of cancer stem-cell markers oct4, sox2, and nanog with clinicopathological features and prognosis in operative patients with rectal cancer. Yonsei Med. J. 2018, 59, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Rasti, A.; Mehrazma, M.; Madjd, Z.; Abolhasani, M.; Saeednejad Zanjani, L.; Asgari, M. Co-expression of cancer stem cell markers oct4 and nanog predicts poor prognosis in renal cell carcinomas. Sci. Rep. 2018, 8, 11739. [Google Scholar] [CrossRef] [PubMed]
- Gwak, J.M.; Kim, M.; Kim, H.J.; Jang, M.H.; Park, S.Y. Expression of embryonal stem cell transcription factors in breast cancer: Oct4 as an indicator for poor clinical outcome and tamoxifen resistance. Oncotarget 2017, 8, 36305–36318. [Google Scholar] [CrossRef]
- Silva, I.A.; Bai, S.M.; McLean, K.; Yang, K.; Griffith, K.; Thomas, D.; Ginestier, C.; Johnston, C.; Kueck, A.; Reynolds, R.K.; et al. Aldehyde dehydrogenase in combination with cd133 defines angiogenic ovarian cancer stem cells that portend poor patient survival. Cancer Res. 2011, 71, 3991–4001. [Google Scholar] [CrossRef] [PubMed]
- Abubaker, K.; Latifi, A.; Luwor, R.; Nazaretian, S.; Zhu, H.; Quinn, M.A.; Thompson, E.W.; Findlay, J.K.; Ahmed, N. Short-term single treatment of chemotherapy results in the enrichment of ovarian cancer stem cell-like cells leading to an increased tumor burden. Mol. Cancer 2013, 12, 24. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.Q.; Mou, L.S.; Cheng, K.W.; Wang, C.Y.; Deng, X.S.; Chen, J.R.; Fan, Z.B.; Ni, Y. Hepatocellular carcinoma stem cell-like cells are enriched following low-dose 5-fluorouracil chemotherapy. Oncol. Lett. 2016, 12, 2511–2516. [Google Scholar] [CrossRef]
- Hu, X.; Ghisolfi, L.; Keates, A.C.; Zhang, J.; Xiang, S.; Lee, D.-k.; Li, C.J. Induction of cancer cell stemness by chemotherapy. Cell Cycle 2012, 11, 2691–2698. [Google Scholar] [CrossRef]
- Bourguignon, L.Y.; Spevak, C.C.; Wong, G.; Xia, W.; Gilad, E. Hyaluronan-cd44 interaction with protein kinase c(epsilon) promotes oncogenic signaling by the stem cell marker nanog and the production of microRNA-21, leading to down-regulation of the tumor suppressor protein pdcd4, anti-apoptosis, and chemotherapy resistance in breast tumor cells. J. Biol. Chem. 2009, 284, 26533–26546. [Google Scholar]
- Bourguignon, L.Y.; Earle, C.; Wong, G.; Spevak, C.C.; Krueger, K. Stem cell marker (nanog) and stat-3 signaling promote microRNA-21 expression and chemoresistance in hyaluronan/cd44-activated head and neck squamous cell carcinoma cells. Oncogene 2012, 31, 149–160. [Google Scholar] [CrossRef]
- Bourguignon, L.Y.; Wong, G.; Earle, C.; Chen, L. Hyaluronan-cd44v3 interaction with oct4-sox2-nanog promotes mir-302 expression leading to self-renewal, clonal formation, and cisplatin resistance in cancer stem cells from head and neck squamous cell carcinoma. J. Biol. Chem. 2012, 287, 32800–32824. [Google Scholar] [CrossRef] [PubMed]
- Shibue, T.; Weinberg, R.A. Emt, cscs, and drug resistance: The mechanistic link and clinical implications. Nat. Rev. Clin. Oncol. 2017, 14, 611–629. [Google Scholar] [CrossRef] [PubMed]
- Giordano, A.; Gao, H.; Anfossi, S.; Cohen, E.; Mego, M.; Lee, B.N.; Tin, S.; De Laurentiis, M.; Parker, C.A.; Alvarez, R.H.; et al. Epithelial-mesenchymal transition and stem cell markers in patients with her2-positive metastatic breast cancer. Mol. Cancer Ther. 2012, 11, 2526–2534. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Chen, Y.W.; Hsu, H.S.; Tseng, L.M.; Huang, P.I.; Lu, K.H.; Chen, D.T.; Tai, L.K.; Yung, M.C.; Chang, S.C.; et al. Aldehyde dehydrogenase 1 is a putative marker for cancer stem cells in head and neck squamous cancer. Biochem. Biophys. Res. Commun. 2009, 385, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Pang, R.; Law, W.L.; Chu, A.C.Y.; Poon, J.T.; Lam, C.S.C.; Chow, A.K.M.; Ng, L.; Cheung, L.W.H.; Lan, X.R.; Lan, H.Y.; et al. A subpopulation of cd26(+) cancer stem cells with metastatic capacity in human colorectal cancer. Cell Stem Cell 2010, 6, 603–615. [Google Scholar] [CrossRef]
- Wellner, U.; Schubert, J.; Burk, U.C.; Schmalhofer, O.; Zhu, F.; Sonntag, A.; Waldvogel, B.; Vannier, C.; Darling, D.; zur Hausen, A.; et al. The emt-activator zeb1 promotes tumorigenicity by repressing stemness-inhibiting micrornas. Nat. Cell Biol. 2009, 11, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, B.T.; Gonzalez-Angulo, A.M.; Stemke-Hale, K.; Gilcrease, M.Z.; Krishnamurthy, S.; Lee, J.S.; Fridlyand, J.; Sahin, A.; Agarwal, R.; Joy, C.; et al. Characterization of a naturally occurring breast cancer subset enriched in epithelial-to-mesenchymal transition and stem cell characteristics. Cancer Res. 2009, 69, 4116–4124. [Google Scholar] [CrossRef] [PubMed]
- Porsch, H.; Bernert, B.; Mehic, M.; Theocharis, A.D.; Heldin, C.H.; Heldin, P. Efficient tgfbeta-induced epithelial-mesenchymal transition depends on hyaluronan synthase has2. Oncogene 2013, 32, 4355–4365. [Google Scholar] [CrossRef]
- Zoltan-Jones, A.; Huang, L.; Ghatak, S.; Toole, B.P. Elevated hyaluronan production induces mesenchymal and transformed properties in epithelial cells. J. Biol. Chem. 2003, 278, 45801–45810. [Google Scholar] [CrossRef]
- Nguyen, N.; Kumar, A.; Chacko, S.; Ouellette, R.J.; Ghosh, A. Human hyaluronic acid synthase-1 promotes malignant transformation via epithelial-to-mesenchymal transition, micronucleation and centrosome abnormalities. Cell Commun. Signal 2017, 15, 48. [Google Scholar] [CrossRef]
- Galluzzi, L.; Senovilla, L.; Vitale, I.; Michels, J.; Martins, I.; Kepp, O.; Castedo, M.; Kroemer, G. Molecular mechanisms of cisplatin resistance. Oncogene 2012, 31, 1869–1883. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.I.; Haber, M.; Henderson, M.J.; Norris, M.D. Abc transporters in cancer: More than just drug efflux pumps. Nat. Rev. Cancer 2010, 10, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Ween, M.P.; Armstrong, M.A.; Oehler, M.K.; Ricciardelli, C. The role of ABC transporters in ovarian cancer progression and chemoresistance. Crit. Rev. Oncol. Hematol. 2015, 96, 220–256. [Google Scholar] [CrossRef] [PubMed]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of abc transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Gros, P.; Neriah, Y.B.; Croop, J.M.; Housman, D.E. Isolation and expression of a complementary DNA that confers multidrug resistance. Nature 1986, 323, 728–731. [Google Scholar] [CrossRef] [PubMed]
- Veneroni, S.; Zaffaroni, N.; Daidone, M.G.; Benini, E.; Villa, R.; Silvestrini, R. Expression of p-glycoprotein and in vitro or in vivo resistance to doxorubicin and cisplatin in breast and ovarian cancers. Eur. J. Cancer 1994, 30, 1002–1007. [Google Scholar] [CrossRef]
- Misra, S.; Ghatak, S.; Toole, B.P. Regulation of mdr1 expression and drug resistance by a positive feedback loop involving hyaluronan, phosphoinositide 3-kinase, and erbb2. J. Biol. Chem. 2005, 280, 20310–20315. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, R.; Takahashi, F.; Cui, R.; Yoshioka, M.; Gu, T.; Sasaki, S.; Tominaga, S.; Nishio, K.; Tanabe, K.K.; Takahashi, K. Interaction between cd44 and hyaluronate induces chemoresistance in non-small cell lung cancer cell. Cancer Lett. 2007, 252, 225–234. [Google Scholar] [CrossRef]
- Cordo Russo, R.I.; García, M.G.; Alaniz, L.; Blanco, G.; Alvarez, E.; Hajos, S.E. Hyaluronan oligosaccharides sensitize lymphoma resistant cell lines to vincristine by modulating p-glycoprotein activity and pi3k/akt pathway. Int. J. Cancer 2008, 122, 1012–1018. [Google Scholar] [CrossRef]
- Gilg, A.G.; Tye, S.L.; Tolliver, L.B.; Wheeler, W.G.; Visconti, R.P.; Duncan, J.D.; Kostova, F.V.; Bolds, L.N.; Toole, B.P.; Maria, B.L. Targeting hyaluronan interactions in malignant gliomas and their drug-resistant multipotent progenitors. Clin. Cancer Res. 2008, 14, 1804–1813. [Google Scholar] [CrossRef]
- Slomiany, M.G.; Dai, L.; Tolliver, L.B.; Grass, G.D.; Zeng, Y.; Toole, B.P. Inhibition of functional hyaluronan-cd44 interactions in cd133-positive primary human ovarian carcinoma cells by small hyaluronan oligosaccharides. Clin. Cancer Res. 2009, 15, 7593–7601. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.M.; Ridley, A.J. Rho gtpases in cancer cell biology. FEBS Lett. 2008, 582, 2093–2101. [Google Scholar] [CrossRef] [PubMed]
- Hanna, S.; El-Sibai, M. Signaling networks of rho gtpases in cell motility. Cell Signal 2013, 25, 1955–1961. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J. Rho gtpase signalling in cell migration. Curr. Opin. Cell Boil. 2015, 36, 103–112. [Google Scholar] [CrossRef]
- Sharma, S.; Santiskulvong, C.; Rao, J.Y.; Gimzewski, J.K.; Dorigo, O. The role of rho gtpase in cell stiffness and cisplatin resistance in ovarian cancer cells. Integr. Biol.-UK 2014, 6, 611–617. [Google Scholar] [CrossRef]
- Ohta, T.; Takahashi, T.; Shibuya, T.; Amita, M.; Henmi, N.; Takahashi, K.; Kurachi, H. Inhibition of the rho/rock pathway enhances the efficacy of cisplatin through the blockage of hypoxia-inducible factor-1alpha in human ovarian cancer cells. Cancer Biol. Ther. 2012, 13, 25–33. [Google Scholar] [CrossRef]
- Toulany, M.; Rodemann, H.P. Phosphatidylinositol 3-kinase/akt signaling as a key mediator of tumor cell responsiveness to radiation. Semin. Cancer Boil. 2015, 35, 180–190. [Google Scholar] [CrossRef]
- Engelman, J.A.; Luo, J.; Cantley, L.C. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat. Rev. Genet. 2006, 7, 606–619. [Google Scholar] [CrossRef]
- Engelman, J.A. Targeting pi3k signalling in cancer: Opportunities, challenges and limitations. Nat. Rev. Cancer 2009, 9, 550–562. [Google Scholar] [CrossRef]
- Datta, S.R.; Dudek, H.; Tao, X.; Masters, S.; Fu, H.; Gotoh, Y.; Greenberg, M.E. Akt phosphorylation of bad couples survival signals to the cell-intrinsic death machinery. Cell 1997, 91, 231–241. [Google Scholar] [CrossRef]
- Tang, E.D.; Nunez, G.; Barr, F.G.; Guan, K.L. Negative regulation of the forkhead transcription factor FKHR by akt. J. Biol. Chem. 1999, 274, 16741–16746. [Google Scholar] [CrossRef] [PubMed]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef]
- Takaishi, H.; Konishi, H.; Matsuzaki, H.; Ono, Y.; Shirai, Y.; Saito, N.; Kitamura, T.; Ogawa, W.; Kasuga, M.; Kikkawa, U.; et al. Regulation of nuclear translocation of forkhead transcription factor afx by protein kinase b. Proc. Natl. Acad. Sci. USA 1999, 96, 11836–11841. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, L.; Greshock, J.; Colligon, T.A.; Wang, Y.; Ward, R.; Katsaros, D.; Lassus, H.; Butzow, R.; Godwin, A.K.; et al. Frequent genetic abnormalities of the pi3k/akt pathway in primary ovarian cancer predict patient outcome. Genes Chromosom. Cancer 2011, 50, 606–618. [Google Scholar] [CrossRef]
- Gao, N.; Flynn, D.C.; Zhang, Z.; Zhong, X.S.; Walker, V.; Liu, K.J.; Shi, X.; Jiang, B.H. G1 cell cycle progression and the expression of g1 cyclins are regulated by pi3k/akt/mtor/p70s6k1 signaling in human ovarian cancer cells. Am. J. Physiol. Cell Physiol. 2004, 287, C281–291. [Google Scholar] [CrossRef]
- Bates, R.C.; Edwards, N.S.; Burns, G.F.; Fisher, D.E. A cd44 survival pathway triggers chemoresistance via lyn kinase and phosphoinositide 3-kinase/akt in colon carcinoma cells. Cancer Res. 2001, 61, 5275–5283. [Google Scholar]
- Yang, X.; Fraser, M.; Moll, U.M.; Basak, A.; Tsang, B.K. Akt-mediated cisplatin resistance in ovarian cancer: Modulation of p53 action on caspase-dependent mitochondrial death pathway. Cancer Res. 2006, 66, 3126–3136. [Google Scholar] [CrossRef]
- Gagnon, V.; Mathieu, I.; Sexton, É.; Leblanc, K.; Asselin, E. Akt involvement in cisplatin chemoresistance of human uterine cancer cells. Gynecol. Oncol. 2004, 94, 785–795. [Google Scholar] [CrossRef]
- Yu, H.-G.; Ai, Y.-W.; Yu, L.-L.; Zhou, X.-D.; Liu, J.; Li, J.-H.; Xu, X.-M.; Liu, S.; Chen, J.; Liu, F.; et al. Phosphoinositide 3-kinase/akt pathway plays an important role in chemoresistance of gastric cancer cells against etoposide and doxorubicin induced cell death. Int. J. Cancer 2007, 122, 433–443. [Google Scholar] [CrossRef]
- Cordo-Russo, R.I.; Alaniz, L.D.; Saccodossi, N.; Lompardia, S.; Blanco, G.; Alvarez, E.; Garcia, M.G.; Hajos, S.E. Hyaluronan induces migration of multidrug-resistant lymphoma cell lines in vitro through tiam1 activation by a pi3k-dependent mechanism. Leuk. Res. 2010, 34, 1525–1532. [Google Scholar] [CrossRef]
- Ghatak, S.; Misra, S.; Toole, B.P. Hyaluronan oligosaccharides inhibit anchorage-independent growth of tumor cells by suppressing the phosphoinositide 3-kinase/akt cell survival pathway. J. Biol. Chem. 2002, 277, 38013–38020. [Google Scholar] [CrossRef] [PubMed]
- Santarpia, L.; Lippman, S.M.; El-Naggar, A.K. Targeting the mapk-ras-raf signaling pathway in cancer therapy. Expert Opin. Ther. Targets 2012, 16, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, S.M.; Paplomata, E.; Peake, B.M.; Sanabria, E.; Chen, Z.; Nahta, R. P38 mapk contributes to resistance and invasiveness of her2-overexpressing breast cancer. Curr. Med. Chem. 2014, 21, 501–510. [Google Scholar] [CrossRef] [PubMed]
- McGivern, N.; El-Helali, A.; Mullan, P.; McNeish, I.A.; Harkin, D.P.; Kennedy, R.D.; McCabe, N. Activation of mapk signalling results in resistance to saracatinib (azd0530) in ovarian cancer. Oncotarget 2018, 9, 4722–4736. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.C.; Chen, C.H.; Wang, J.Y.; Lin, Y.S.; Chang, J.K.; Ho, M.L. Hyaluronan size alters chondrogenesis of adipose-derived stem cells via the cd44/erk/sox-9 pathway. Acta Biomater. 2018, 66, 224–237. [Google Scholar] [CrossRef]
- Park, G.B.; Ko, H.S.; Kim, D. Sorafenib controls the epithelial-mesenchymal transition of ovarian cancer cells via egf and the cd44-ha signaling pathway in a cell type-dependent manner. Mol. Med. Rep. 2017, 16, 1826–1836. [Google Scholar] [CrossRef]
- Wang, S.J.; Bourguignon, L.Y.W. Hyaluronan and the interaction between cd44 and epidermal growth factor receptor in oncogenic signaling and chemotherapy resistance in head and neck cancer. Arch. Otolaryngol. 2006, 132, 771–778. [Google Scholar] [CrossRef]
- Kultti, A.; Pasonen-Seppanen, S.; Jauhiainen, M.; Rilla, K.J.; Karna, R.; Pyoria, E.; Tammi, R.H.; Tammi, M.I. 4-methylumbelliferone inhibits hyaluronan synthesis by depletion of cellular udp-glucuronic acid and downregulation of hyaluronan synthase 2 and 3. Exp. Cell Res. 2009, 315, 1914–1923. [Google Scholar] [CrossRef]
- Urakawa, H.; Nishida, Y.; Wasa, J.; Arai, E.; Zhuo, L.; Kimata, K.; Kozawa, E.; Futamura, N.; Ishiguro, N. Inhibition of hyaluronan synthesis in breast cancer cells by 4-methylumbelliferone suppresses tumorigenicity in vitro and metastatic lesions of bone in vivo. Int. J. Cancer 2012, 130, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Yates, T.J.; Lopez, L.E.; Lokeshwar, S.D.; Ortiz, N.; Kallifatidis, G.; Jordan, A.; Hoye, K.; Altman, N.; Lokeshwar, V.B. Dietary supplement 4-methylumbelliferone: An effective chemopreventive and therapeutic agent for prostate cancer. J. Natl. Cancer Inst. 2015, 107. [Google Scholar] [CrossRef] [PubMed]
- Lokeshwar, V.B.; Lopez, L.E.; Munoz, D.; Chi, A.; Shirodkar, S.P.; Lokeshwar, S.D.; Escudero, D.O.; Dhir, N.; Altman, N. Antitumor activity of hyaluronic acid synthesis inhibitor 4-methylumbelliferone in prostate cancer cells. Cancer Res. 2010, 70, 2613–2623. [Google Scholar] [CrossRef] [PubMed]
- Karalis, T.T.; Heldin, P.; Vynios, D.H.; Neill, T.; Buraschi, S.; Iozzo, R.V.; Karamanos, N.K.; Skandalis, S.S. Tumor-suppressive functions of 4-mu on breast cancer cells of different er status: Regulation of hyaluronan/has2/cd44 and specific matrix effectors. Matrix Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Tamura, R.; Yokoyama, Y.; Yoshida, H.; Imaizumi, T.; Mizunuma, H. 4-methylumbelliferone inhibits ovarian cancer growth by suppressing thymidine phosphorylase expression. J. Ovarian Res. 2014, 7, 94. [Google Scholar] [CrossRef] [PubMed]
- Edward, M.; Quinn, J.A.; Pasonen-Seppanen, S.M.; McCann, B.A.; Tammi, R.H. 4-methylumbelliferone inhibits tumour cell growth and the activation of stromal hyaluronan synthesis by melanoma cell-derived factors. Br. J. Dermatol. 2010, 162, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Matsumoto, S.; Fujita, Y.; Kuroda, A.; Menju, T.; Sonobe, M.; Kondo, N.; Torii, I.; Nakano, T.; Lara, P.N.; et al. Trametinib plus 4-methylumbelliferone exhibits antitumor effects by erk blockade and cd44 downregulation and affects pd-1 and pd-l1 in malignant pleural mesothelioma. J. Thorac. Oncol. 2017, 12, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Teranishi, F.; Takahashi, N.; Gao, N.; Akamo, Y.; Takeyama, H.; Manabe, T.; Okamoto, T. Phosphoinositide 3-kinase inhibitor (wortmannin) inhibits pancreatic cancer cell motility and migration induced by hyaluronan in vitro and peritoneal metastasis in vivo. Cancer Sci. 2009, 100, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhao, S.J.; Karnad, A.; Freeman, J.W. The biology and role of cd44 in cancer progression: Therapeutic implications. J. Hematol. Oncol. 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Lokeshwar, V.B.; Selzer, M.G. Hyaluronidase: Both a tumor promoter and suppressor. In Hyaluronan in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 189–206. [Google Scholar]
- Thompson, C.B.; Shepard, H.M.; Connor, P.M.; Kadhim, S.; Jiang, P.; Osgood, R.J.; Bookbinder, L.H.; Li, X.; Sugarman, B.J.; Connor, R.J.; et al. Enzymatic depletion of tumor hyaluronan induces antitumor responses in preclinical animal models. Mol. Cancer Ther. 2010, 9, 3052–3064. [Google Scholar] [CrossRef]
- Hingorani, S.R.; Zheng, L.; Bullock, A.J.; Seery, T.E.; Harris, W.P.; Sigal, D.S.; Braiteh, F.; Ritch, P.S.; Zalupski, M.M.; Bahary, N.; et al. Halo 202: Randomized phase ii study of pegph20 plus nab-paclitaxel/gemcitabine versus nab-paclitaxel/gemcitabine in patients with untreated, metastatic pancreatic ductal adenocarcinoma. J. Clin. Oncol. 2018, 36, 359–366. [Google Scholar] [CrossRef]
- Ramanathan, R.K.; McDonough, S.; Philip, P.A.; Hingorani, S.R.; Lacy, J.; Kortmansky, J.S. A phase ib/ii randomized study of mfolfirinox (mffox) plus pegylated recombinant human hyaluronidase (pegph20) versus mffox alone in patients with good performance status metastatic pancreatic adenocarcinoma (mpc): Swog s1313 (nct #01959139). J. Clin. Oncol. 2018, 36, 208. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cell Type | HA (kDa) | Functional role and effects of HA | Reference |
|---|---|---|---|
| Inflammation | |||
| Mouse knee chondrocytes | 50, 1000, 5000 | 50 kDa promoted and 5000 kDa reduced inflammatory response | [41] |
| Nasal epithelium RPMI 256 | 370, 900, 1600 | 1600 kDa decreased the inflammatory effects and 900kDa reduced ROS production | [42] |
| BALB/c mice | 250, 470, 780, 900, 1200 | 780, 900 and 1200 kDa reduced and 250 kDa increased liver injury. 900 kDa HA reduced inflammation | [40] |
| RAW 264.7 and MHS macrophages | 11, 52, 87, 250, 970 | No molecular weight of HA stimulated macrophage inflammatory response | [45] |
| Primary mesangial cells | 1.5, 3 | HA have no effect and HYAL (containing endotoxins) stimulated an inflammatory response | [46] |
| BMDM and BMDC | 10, 28, 243, 1680 Rooster comb Umbilical cord | Only umbilical cord HA and HYAL stimulated BMDM and BMDC due to endotoxin contamination | [44] |
| J774A, ATCC, TIB-67 murine macrophages | oHA, 5, 60, 800, 3000 | oHA, 5 and 60 kDa were pro-inflammatory, 800 and 3000 kDa anti-inflammatory | [48] |
| Peripheral blood mononuclear cells (PBMC) | ~2000, ~80–800 | 2000 kDa HA promotes and 80–800 kDa HA inhibits the differentiation of PBMC cells to fibrocytes | [13] |
| Angiogenesis and Wound Repair | |||
| BAEC (Bovine aortic endothelial cells) | oHA (3–16 disaccharides) Rooster comb HA | oHA promotes BAEC proliferation and angiogenensis, inhibited by rooster comb HA | [53] |
| BAEC | Rooster comb HA oHA | oHA via MAPK promotes BAEC proliferation and wound healing | [54] |
| BAEC | oHA (unspecified biological source) | oHA promotes and native HA inhibits BAEC proliferation and expression of angiogenesis early response genes | [55] |
| Porcine vascular endothelial cells | oHA (2–10 disaccharides) Umbilical cord HA | oHA promoted cell proliferation, wound healing and migration via MAPK and RHAMM. Native HA promoted cell migration | [56] |
| Human umbilical vein endothelial cells Sprague-Dawley | oHA (2–10 disaccharides) | oHA promotes cell proliferation, tube formation and in vivo and in vitro wound healing | [57] |
| Arterial Smooth Muscle Cells | 2100 | HA promotes cell migration via RhoA and Rac kinases, PI3K activates Rac | [60] |
| NIH-3T3 Mouse fibroblasts | 980, 132, 31, 2.3 | 980 kDa HA promotes cell invasion via NFκB activity and Snail2 expression | [59] |
| Sprague Dawley rats dermal fibroblasts | 5, 40, 500, oHA: 4, 6, 8 and 10 mer | 6 mer & 8 mer oHA promoted wound healing in vivo & in vitro via CD44 and RHAMM, 6mer oHA recruits M1 and M2 macrophages | [58] |
| Human umbilical vein endothelial cells Sprague-Dawley rat | oHA (2–10 disaccharides) | oHA promotes in vivo and in vitro cell proliferation and wound healing | [57] |
| Embryogenesis and Gland Branching | |||
| Primary Trophoblasts | HMW-HA MMW-HA LMW-HA (unspecified weights) | HMW-HA and MMW-HA promote cell proliferation, invasion and survival. LMW-HA has no effect | [61] |
| Mammary epithelial cell line (Ep-H4) | 6–21, 50, 240, 500 | 240 kDa and 500 kDa HA inhibit and 6–25 kDa HA promote mammary gland branching | [62] |
| Holtzman rat kidneys | 234.4, 132.3, 64, 17, 6.55 | 234.4 kDa and 132.3 kDa HA inhibit ureteric gland morphogenesis, 17 kDa and 6.5 kDa stimulates branching morphogenesis. | [63] |
| Cell Type | HA (kDa) | Functional role and effects of HA | Reference |
|---|---|---|---|
| Ovarian cancer | |||
| SKOV-3 | 1000 | HA promotes cell migration and growth via Rac1 and Ras | [65] |
| SKOV-3 | 500 | HA promotes cell migration via CDC42 and ERK1 | [66] |
| SKOV-3 OV-90 OVCAR-3 OVCAR-5 | Umbilical Cord HA oHA (6–10) | HA promotes resistance to carboplatin via ABCB3, ABCC2, ABCC3 and ABCC1 expression. oHA abrogated HA effect | [8] |
| SKOV-3 | 500 | HA increases REX1, SOX2 and MDR1 expression, promoting drug resistance | [67] |
| Breast cancer | |||
| MDA-MB-231 | 1000 | HA promotes cell growth and invasion via RhoA | [68] |
| MCF-7 | 500 | HA increases REX1, SOX2 and MDR1 expression, promoting drug resistance | [67] |
| MCF-7 | 500 | HA promotes MDR1 and Bcl-xL (anti-apoptotic) expression, cell growth and invasion | [69] |
| MDA-MB-231 | 400–500 | HA promotes cell growth and invasion via RhoA, RhoC and ROK | [70] |
| MDA-MB-231 | 3–5 500–1000 | 3–5 kDa promotes cell invasion | [71] |
| 4T-1 SKBR-3 | 35, 117 | 35 kDa promotes cell migration and invasion | [72] |
| Head and neck squamous cell carcinoma | |||
| HSC-3 | 500 Rooster Comb | HA via CD44 complexes with leukemia associated RhoGEF (LARG) and epidermal growth factor receptor (EGFR). Promotes cell migration and growth | [73] |
| HSC-3 | 500 | HA promotes cell migration, proliferation and cisplatin resistance via PI3K and ROK | [74] |
| HSC-3 | 5, 20, 200, 700 | 200 kDa HA promotes stemness and cisplatin resistance | [75] |
| HSC-3 | 500–700 | HA promoted expression of CSC markers, sphere and clone formation, cell growth and invasion, cisplatin resistance via RhoC | [76] |
| Choriocarcinoma | |||
| JEG-3 | LMW-HA (unspecified weight) 1500–1800 | LMW-HA promotes cell migration via RHAMM, PI3K and MAPK | [77] |
| Leukemia | |||
| K562 Vincristine sensitive Kv562 Resistant | 1500–1800 oHA | HA promotes cell proliferation via CD44, inhibited by 4-methylumbelliferone (4MU) and oHA (in K562) | [78] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Price, Z.K.; Lokman, N.A.; Ricciardelli, C. Differing Roles of Hyaluronan Molecular Weight on Cancer Cell Behavior and Chemotherapy Resistance. Cancers 2018, 10, 482. https://doi.org/10.3390/cancers10120482
Price ZK, Lokman NA, Ricciardelli C. Differing Roles of Hyaluronan Molecular Weight on Cancer Cell Behavior and Chemotherapy Resistance. Cancers. 2018; 10(12):482. https://doi.org/10.3390/cancers10120482
Chicago/Turabian StylePrice, Zoe K., Noor A. Lokman, and Carmela Ricciardelli. 2018. "Differing Roles of Hyaluronan Molecular Weight on Cancer Cell Behavior and Chemotherapy Resistance" Cancers 10, no. 12: 482. https://doi.org/10.3390/cancers10120482
APA StylePrice, Z. K., Lokman, N. A., & Ricciardelli, C. (2018). Differing Roles of Hyaluronan Molecular Weight on Cancer Cell Behavior and Chemotherapy Resistance. Cancers, 10(12), 482. https://doi.org/10.3390/cancers10120482