Proteomic Analysis of Secretomes of Oncolytic Herpes Simplex Virus-Infected Squamous Cell Carcinoma Cells

Abstract
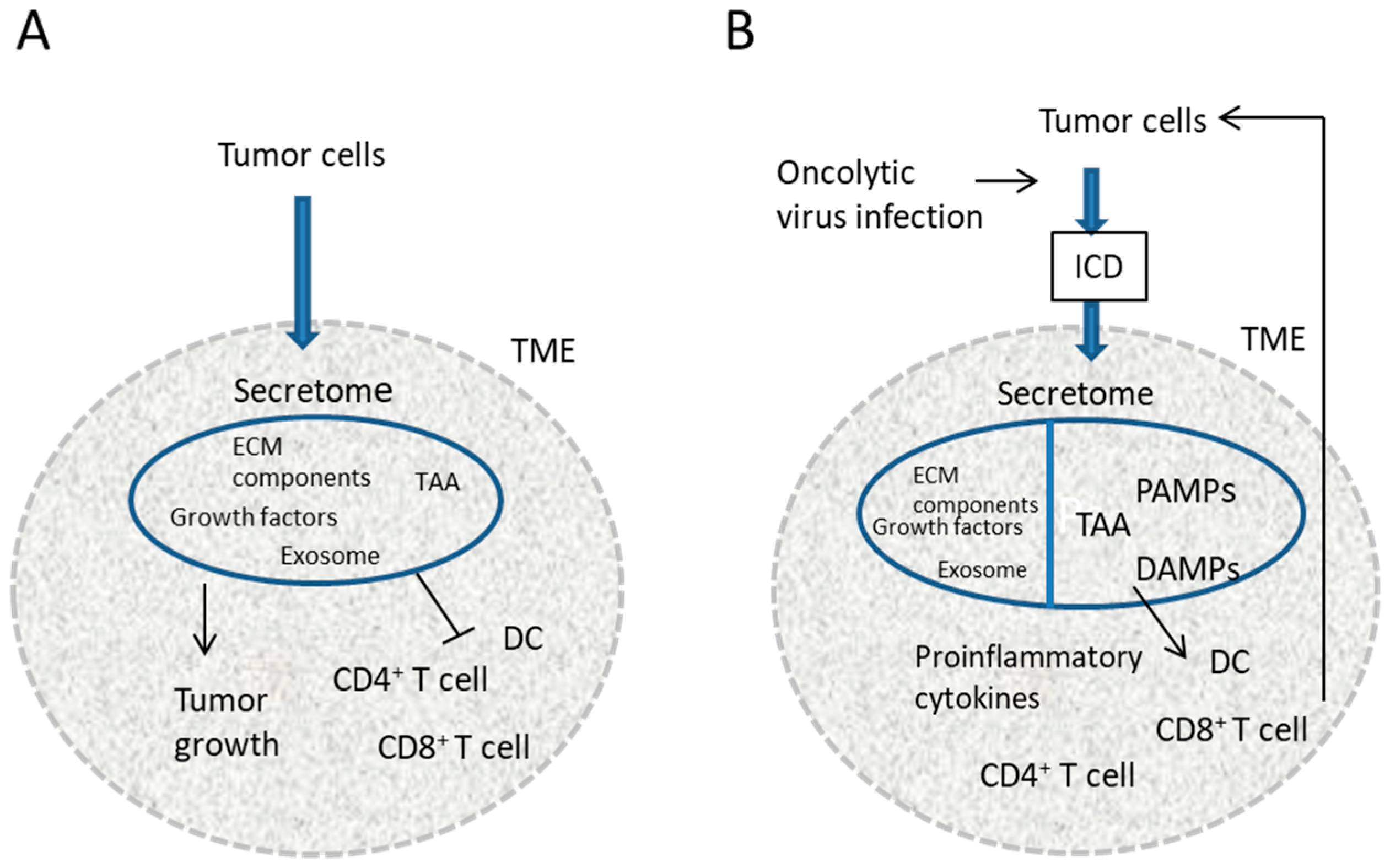
1. Introduction
2. Results
2.1. Classification of Released Proteins of Med24 and MedC
2.2. Alteration of DAMPs and DAMPs-Related Proteins in the Secretomes
2.3. Viral Proteins in the Secretomes
3. Discussion
4. Materials and Methods
4.1. Cells and Virus
4.2. Preparation of Concentrated Supernatants of RH2-Infected Cells
4.3. Proteomic Analysis
4.4. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Thomas, D.L.; Fraser, N.W. HSV-1 therapy of primary tumors reduces the number of metastases in an immune-competent model of metastatic breast cancer. Mol. Ther. 2003, 8, 543–551. [Google Scholar] [CrossRef]
- Benencia, F.; Courrèges, M.C.; Conejo-García, J.R.; Mohammed-Hadley, A.; Coukos, G. Direct vaccination with tumor cells killed with ICP4-deficient HSVd120 elicits effective antitumor immunity. Cancer Biol. Ther. 2006, 5, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.S.; Liu, Z.; Bartlett, D.L. Oncolytic immunotherapy: Dying the right way is a key to eliciting potent antitumor immunity. Front. Oncol. 2014, 4, 74. [Google Scholar] [CrossRef] [PubMed]
- Yura, Y.; Hamada, M. Development of oncolytic virothearpy for the treatment of oral cancer–At the waiting stage for clinical use. Oral Sci. Int. 2017, 14, 1–12. [Google Scholar] [CrossRef]
- Bartlett, D.L.; Liu, Z.; Sathaiah, M.; Ravindranathan, R.; Guo, Z.; He, Y.; Guo, Z.S. Oncolytic viruses as therapeutic cancer vaccines. Mol. Cancer 2013, 12, 103. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.J.; Peng, K.W.; Bell, J.C. Oncolytic virotherapy. Nat. Biotechnol. 2012, 30, 658–670. [Google Scholar] [CrossRef] [PubMed]
- Harrington, K.J.; Puzanov, I.; Hecht, J.R.; Hodi, F.S.; Szabo, Z.; Murugappan, S.; Kaufman, H.L. Clinical development of talimogene laherparepvec (T-VEC): A modified herpes simplex virus type-1-derived oncolytic immunotherapy. Expert Rev. Anticancer Ther. 2015, 15, 1389–1403. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C. Immunogenicity signals 1,2,3... and 0. Immunol. Today 1989, 10, 283–286. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Coyne, C.B.; Zeh, H.J.; Lotze, M.T. PAMPs and DAMPs: Signal 0s that spur autophagy and immunity. Immunol. Rev. 2012, 249, 158–175. [Google Scholar] [CrossRef] [PubMed]
- Kono, H.; Rock, K.L. How dying cells alert the immune system to danger. Nat. Rev. Immunol. 2008, 8, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Sakai, R.; Ouchi, M.; Onimatsu, H.; Hioki, M.; Kagawa, S.; Uno, F.; Watanabe, Y.; Urata, Y.; Tanaka, N.; et al. Virus-mediated oncolysis induces danger signal and stimulates cytotoxic T-lymphocyte activity via proteasome activator upregulation. Oncogene 2008, 27, 2375–2381. [Google Scholar] [CrossRef] [PubMed]
- Krysko, D.V.; Garg, A.D.; Kaczmarek, A.; Krysko, O.; Agostinis, P.; Vandenabeele, P. Immunogenic cell death and DAMPs in cancer therapy. Nat. Rev. Cancer 2012, 12, 860–875. [Google Scholar] [CrossRef] [PubMed]
- Takasu, A.; Masui, A.; Hamada, M.; Imai, T.; Iwai, S.; Yura, Y. Immunogenic cell death by oncolytic herpes simplex virus type 1 in squamous cell carcinoma cells. Cancer Gene Ther. 2016, 23, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Meshii, N.; Takahashi, G.; Okunaga, S.; Hamada, M.; Iwai, S.; Takasu, A.; Ogawa, Y.; Yura, Y. Enhancement of systemic tumor immunity for squamous cell carcinoma cells by an oncolytic herpes simplex virus. Cancer Gene Ther. 2013, 20, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Pavlou, M.P.; Diamandis, E.P. The cancer cell secretome: A good source for discovering biomarkers? J. Proteom. 2010, 73, 1896–1906. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, J.J.; Matikainen, S.; Nyman, T.A. Global secretome characterization of herpes simplex virus 1-infected human primary macrophages. J. Virol. 2012, 86, 12770–12778. [Google Scholar] [CrossRef] [PubMed]
- Taylor, T.J.; Knipe, D.M. Proteomics of herpes simplex virus replication compartments: Association of cellular DNA replication, repair, recombination, and chromatin remodeling proteins with ICP8. J. Virol. 2004, 78, 5856–5866. [Google Scholar] [CrossRef] [PubMed]
- Fontaine-Rodriguez, E.C.; Taylor, T.J.; Olesky, M.; Knipe, D.M. Proteomics of herpes simplex virus infected cell protein 27: Association with translation initiation factors. Virology 2004, 330, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Suk, H.; Knipe, D.M. Proteomic analysis of the herpes simplex virus 1 virion protein 16 transactivator protein in infected cells. Proteomics 2015, 15, 1957–1967. [Google Scholar] [PubMed]
- Prasannan, L.; Misek, D.E.; Hinderer, R.; Michon, J.; Geiger, J.D.; Hanash, S.M. Identification of beta-tubulin isoforms as tumor antigens in neuroblastoma. Clin. Cancer Res. 2000, 6, 3949–3956. [Google Scholar] [PubMed]
- Jalbout, M.; Bel Hadj Jrad, B.; Bouaouina, N.; Gargouri, J.; Yacoub, S.; Zakhama, A.; Khlifa, R.; Chouchane, L. Autoantibodies to tubulin are specifically associated with the young age onset of the nasopharyngeal carcinoma. Int. J. Cancer 2002, 101, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Adachi-Hayama, M.; Adachi, A.; Shinozaki, N.; Matsutani, T.; Hiwasa, T.; Takiguchi, M.; Saeki, N.; Iwadate, Y. Circulating anti-filamin C autoantibody as a potential serum biomarker for low-grade gliomas. BMC Cancer 2014, 14, 452. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, G.; Pfeiffer, S.; Simons, K.; Matlin, K. Exit of newly synthesized membrane proteins from the trans cisterna of the Golgi complex to the plasma membrane. J. Cell Biol. 1985, 101, 949–964. [Google Scholar] [CrossRef] [PubMed]
- Seliger, B.; Dressler, S.P.; Massa, C.; Recktenwald, C.V.; Altenberend, F.; Bukur, J.; Marincola, F.M.; Wang, E.; Stevanovic, S.; Lichtenfels, R. Identification and characterization of human leukocyte antigen class I ligands in renal cell carcinoma cells. Proteomics 2011, 11, 2528–2541. [Google Scholar] [CrossRef] [PubMed]
- Liou, A.K.; Willison, K.R. Elucidation of the subunit orientation in CCT (chaperonin containing TCP1) from the subunit composition of CCT micro-complexes. EMBO J. 1997, 16, 4311–4316. [Google Scholar] [CrossRef] [PubMed]
- Sternlicht, H.; Farr, G.W.; Sternlicht, M.L.; Driscoll, J.K.; Willison, K.; Yaffe, M.B. The t-complex polypeptide 1 complex is a chaperonin for tubulin and actin in vivo. Proc. Natl. Acad. Sci. USA 1993, 90, 9422–9426. [Google Scholar] [CrossRef] [PubMed]
- Guest, S.T.; Kratche, Z.R.; Bollig-Fischer, A.; Haddad, R.; Ethier, S.P. Two members of the TRiC chaperonin complex, CCT2 and TCP1 are essential for survival of breast cancer cells and are linked to driving oncogenes. Exp. Cell Res. 2015, 332, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Yokota, S.; Yamamoto, Y.; Shimizu, K.; Momoi, H.; Kamikawa, T.; Yamaoka, Y.; Yanagi, H.; Yura, T.; Kubota, H. Increased expression of cytosolic chaperonin CCT in human hepatocellular and colonic carcinoma. Cell Stress Chaperon. 2001, 6, 345–350. [Google Scholar] [CrossRef]
- Coghlin, C.; Carpenter, B.; Dundas, S.R.; Lawrie, L.C.; Telfer, C.; Murray, G.I. Characterization and over-expression of chaperonin t-complex proteins in colorectal cancer. J. Pathol. 2006, 210, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Zheng, M.; Sun, S.; Wang, H.; Yue, Z.; Zhu, Y.; Han, X.; Yang, J.; Zhou, Y.; Cai, Y.; et al. Chaperonin containing TCP1 subunit 5 is a tumor associated antigen of non-small cell lung cancer. Oncotarget 2017, 8, 64170–64179. [Google Scholar] [CrossRef] [PubMed]
- Höfflin, S.; Prommersberger, S.; Uslu, U.; Schuler, G.; Schmidt, C.W.; Lennerz, V.; Dörrie, J.; Schaft, N. Generation of CD8(+) T cells expressing two additional T-cell receptors (TETARs) for personalised melanoma therapy. Cancer Biol. Ther. 2015, 16, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Murshid, A.; Gong, J.; Stevenson, M.A.; Calderwood, S.K. Heat shock proteins and cancer vaccines: Developments in the past decade and chaperoning in the decade to come. Expert Rev. Vaccines 2011, 10, 1553–1568. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Galluzzi, L.; Kepp, O.; Zitvogel, L. Immunogenic cell death in cancer therapy. Annu. Rev. Immunol. 2013, 31, 51–72. [Google Scholar] [CrossRef] [PubMed]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochim. Biophys. Acta 2012, 1820, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Poliakov, A.; Liu, C.; Liu, Y.; Deng, Z.B.; Wang, J.; Cheng, Z.; Shah, S.V.; Wang, G.J.; Zhang, L.; et al. Induction of myeloid-derived suppressor cells by tumor exosomes. Int. J. Cancer 2009, 124, 2621–2633. [Google Scholar] [CrossRef] [PubMed]
- Colunga, A.G.; Laing, J.M.; Aurelian, L. The HSV-2 mutant DeltaPK induces melanoma oncolysis through nonredundant death programs and associated with autophagy and pyroptosis proteins. Gene Ther. 2010, 17, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Workenhe, S.T.; Simmons, G.; Pol, J.G.; Lichty, B.D.; Halford, W.P.; Mossman, K.L. Immunogenic HSV-mediated oncolysis shapes the antitumor immune response and contributes to therapeutic efficacy. Mol. Ther. 2014, 22, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, Y.; Takasu, A.; Yura, Y. Role of autophagy in oncolytic herpes simplex virus type 1-induced cell death in squamous cell carcinoma cells. Cancer Gene Ther. 2017, 24, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F.R.; Capasso, M.; Hagemann, T. The tumor microenvironment at a glance. J. Cell Sci. 2012, 125, 5591–5596. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Clements, D.R.; Sterea, A.M.; Jang, H.W.; Gujar, S.A.; Lee, P.W. Dendritic cells in oncolytic virus-based anti-cancer therapy. Viruses 2015, 7, 6506–6525. [Google Scholar] [CrossRef] [PubMed]
- Werb, Z.; Lu, P. The Role of Stroma in Tumor Development. Cancer J. 2015, 21, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Moehler, M.; Goepfert, K.; Heinrich, B.; Breitbach, C.J.; Delic, M.; Galle, P.R.; Rommelaere, J. Oncolytic virotherapy as emerging immunotherapeutic modality: Potential of parvovirus h-1. Front. Oncol. 2014, 4, 92. [Google Scholar] [CrossRef] [PubMed]
- Lichty, B.D.; Breitbach, C.J.; Stojdl, D.F.; Bell, J.C. Going viral with cancer immunotherapy. Nat. Rev. Cancer 2014, 14, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, G.; Meshii, N.; Hamada, M.; Iwai, S.; Yura, Y. Sequence of a fusogenic herpes simplex virus, RH2, for oncolytic virotherapy. J. Gen. Virol. 2013, 94, 726–737. [Google Scholar] [CrossRef] [PubMed]
- Nagamori, S.; Wiriyasermkul, P.; Guarch, M.E.; Okuyama, H.; Nakagomi, S.; Tadagaki, K.; Nishinaka, Y.; Bodoy, S.; Takafuji, K.; Okuda, S.; et al. Novel cystine transporter in renal proximal tubule identified as a missing partner of cystinuria-related plasma membrane protein rBAT/SLC3A1. Proc. Natl. Acad. Sci. USA 2016, 113, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Lietzén, N.; Ohman, T.; Rintahaka, J.; Julkunen, I.; Aittokallio, T.; Matikainen, S.; Nyman, T.A. Quantitative subcellular proteome and secretome profiling of influenza a virus-infected human primary macrophages. PLoS Pathog. 2011, 7, e1001340. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Accession No. | Protein Name | MW (kDa) | iTRAQ Quantitation b | Fold Change | p-Value | |
|---|---|---|---|---|---|---|
| Med24 | MedC | |||||
| Q9QXS1 | Plectin | 534 | 79.7 | 54 | 1.48 | 0.057 |
| B7FAU9 | Filamin, alpha | 280 | 73.7 | 49.7 | 1.48 | 0.004 * |
| P68372 | Tubulin beta-4B | 50 | 54.7 | 37 | 1.48 | 0.0027 * |
| Q9JKF1 | Ras GTPase-activating-like protein IQGAP1 | 189 | 47.7 | 33.7 | 1.42 | 0.09 |
| Q02053 | Ubiquitin-like modifier-activating enzyme 1 | 118 | 40 | 28 | 1.43 | 0.058 |
| Q5SXR6 | Clathrin heavy chain | 192 | 40 | 26.3 | 1.51 | 0.023 * |
| P80314 | T-complex protein 1 subunit beta | 57 | 39 | 23.7 | 1.65 | 0.012 * |
| P63038 | 60-kDa heat shock protein, mitochondrial | 61 | 32.4 | 20.3 | 1.59 | 0.22 |
| P42932 | T- complex protein 1 subunit theta | 60 | 31.3 | 20.3 | 1.54 | 0.0013 * |
| Q9CWJ9 | Bifunctional purine biosynthesis | 64 | 27.7 | 17 | 1.63 | 0.021 * |
| protein PURH | ||||||
| P11983 | T-complex protein 1 subunit alpha | 60 | 25.7 | 16.7 | 1.54 | 0.0088 * |
| P56480 | ATP synthase subunit beta, mitochondrial | 56 | 24 | 17 | 1.41 | 0.42 |
| Q64737 | Trifunctional purine biosynthetic protein | 108 | 22.7 | 16 | 1.42 | 0.12 |
| adenosine-3 | ||||||
| Q9JHU4 | Cytoplasmic dynein 1 heavy chain 1 | 532 | 26.7 | 12 | 2.22 | 0.16 |
| D3YW87 | Filamin-C | 289 | 33.3 | 20.7 | 1.61 | 0.033 * |
| P80318 | T-complex protein 1 subunit gamma | 61 | 22.7 | 13.3 | 1.7 | 0.0078 * |
| P80315 | T-complex protein 1 subunit delta | 58 | 19.3 | 13.3 | 1.45 | 0.016 * |
| Q7TPV4 | Myb-binding protein 1A | 152 | 21.3 | 11.3 | 1.56 | 0.064 |
| Q8CGC7 | Bifunctional glutamate/proline-tRNA ligase | 170 | 19.7 | 11.3 | 1.74 | 0.033 * |
| Q03265 | ATP synthase subunit alpha, mitochondrial | 60 | 19.3 | 10.3 | 1.87 | 0.17 |
| P23116 | Eukaryotic translation initiation factor 3 | 162 | 17 | 11.3 | 1.5 | 0.078 |
| subunit A | ||||||
| Q78PY7 | Staphylococcal nuclease domain-containing | 102 | 16.7 | 11 | 1.51 | 0.12 |
| protein1 | ||||||
| B1AX58 | Plastin-3 | 72 | 16 | 11.3 | 1.41 | 0.00015 * |
| Q9D8N0 | Elongation factor 1-gamma | 50 | 16.7 | 10.3 | 1.61 | 0.18 |
| Q8BGQ7 | Alanine--tRNA ligase, cytoplasmic | 107 | 15.7 | 10.3 | 1.51 | 0.21 |
| Q61553 | Fascin | 55 | 15 | 10.3 | 1.45 | 0.049 * |
| Q8CIE6 | Coatomer subunit alpha | 138 | 14.7 | 10 | 1.47 | 0.025 * |
| P21550 | Beta-enolase | 47 | 38 | 25.3 | 1.5 | 0.002 * |
| Q922F4 | Tubulin beta-6 chain | 50 | 27.7 | 17.7 | 1.57 | 0.011 * |
| Accession No. | Protein Name | MW (kDa) | iTRAQ Quantitation b | Fold Change | p-Value | |
|---|---|---|---|---|---|---|
| Med24 | MedC | |||||
| A0A087WR50 | Fibronectin | 263 | 33.7 | 99.0 | 0.34 | 0.00032 |
| P35441 | Thrombospondin-1 | 130 | 15.7 | 48.3 | 0.32 | 0.00045 |
| E9PZ16 | Basement membrane-specific heparan | 470 | 9.3 | 50.0 | 0.19 | 0.0011 |
| sulfate proteoglycan core protein | ||||||
| P97298 | Pigment epithelium-derived factor | 46 | 16.7 | 42.7 | 0.39 | 0.0011 |
| A2AQ53 | Fibrillin-1 | 312 | 10.0 | 41.7 | 0.24 | 0.0000830 |
| Q3UQ28 | Peroxidasin homolog | 165 | 6.7 | 36.3 | 0.18 | 0.0019 |
| P33434 | 72-kDa type IV collagenase | 74 | 11.0 | 31.3 | 0.35 | 0.0074 |
| E9PZ00 | Prosaposin | 61 | 9.0 | 28.3 | 0.32 | 0.0016 |
| D3Z598 | Latent-transforming growth factor beta-binding protein 4 | 167 | 6.0 | 29.7 | 0.2 | 0.00022 |
| Q07797 | Galectin-3-binding protein | 64 | 7.3 | 25.7 | 0.29 | 0.00022 |
| P28653 | Biglycan | 42 | 4.7 | 27.7 | 0.17 | 0.0011 |
| P10605 | Cathepsin B | 37 | 10.0 | 21.3 | 0.47 | 0.023 |
| Q00493 | Carboxypeptidase E | 53 | 10.3 | 21.0 | 0.49 | 0.011 |
| Q8VHY0 | Chondroitin sulfate proteoglycan 4 | 252 | 6.3 | 25.0 | 0.25 | 0.0015 |
| Q06890 | Clusterin | 52 | 6.3 | 23.3 | 0.27 | 0.00044 |
| Q9R118 | Serine protease HTRA1 | 51 | 7.0 | 22.3 | 0.31 | 0.00078 |
| P07214 | SPARC | 34 | 6.3 | 22.7 | 0.28 | 0.00029 |
| Q61398 | Procollagen C | 50 | 4.7 | 24.3 | 0.19 | 0.0000940 |
| Q9QZF2 | Glypican-1 | 61 | 7.3 | 19.7 | 0.37 | 0.0078 |
| Q640N1 | Adipocyte enhancer-binding protein 1 | 128 | 5.7 | 20.7 | 0.27 | 0.0004 |
| Q8BND5 | Sulfhydryl oxidase 1 | 83 | 5.3 | 21.0 | 0.25 | 0.0018 |
| Q02819 | Nucleobindin-1 | 53 | 7.7 | 17.3 | 0.44 | 0.016 |
| Q9R0E2 | Procollagen-lysine, 2-oxoglutarate | 84 | 5.7 | 17.7 | 0.32 | 0.0037 |
| 5-dioxygenase 1 | ||||||
| Q61468 | Mesothelin | 69 | 6.3 | 16.7 | 0.38 | 0.00039 |
| M0QWP1 | Agrin | 217 | 2.7 | 18.3 | 0.15 | 0.00000480 |
| Q61508 | Extracellular matrix protein 1 | 63 | 3.0 | 17.7 | 0.17 | 0.0014 |
| O08665 | Semaphorin-3A | 89 | 5.3 | 15.0 | 0.36 | 0.0054 |
| Q9WVH9 | Fibulin-5 | 50 | 2.3 | 18.0 | 0.13 | 0.00071 |
| Q60963 | Platelet-activating factor acetylhydrolase | 49 | 2.3 | 17.7 | 0.13 | 0.00037 |
| O88325 | Alpha-N-acetylglucosaminidase | 83 | 3.0 | 15.7 | 0.19 | 0.00046 |
| P21460 | Cystatin-C | 16 | 5.3 | 13.3 | 0.34 | 0.0058 |
| P55065 | Phospholipid transfer protein | 54 | 0.7 | 18.0 | 0.04 | 0.00048 |
| P25785 | Metalloproteinase inhibitor 2 | 24 | 5.7 | 12.7 | 0.45 | 0.012 |
| Q8QZR4 | Out at first protein homolog | 32 | 4.7 | 13.7 | 0.34 | 0.0049 |
| G3XA35 | MCG116562, isoform CRA_a | 263 | 4.7 | 13.3 | 0.35 | 0.007 |
| Q8CE08 | Prostatic acid phosphatase | 44 | 2.7 | 15.3 | 0.17 | 0.0000700 |
| P41245 | Matrixmetalloproteinase -9 | 81 | 3.0 | 14.7 | 0.2 | 0.0013 |
| P12023 | Amyloid beta A4 protein | 87 | 3.3 | 13.3 | 0.25 | 0.0042 |
| P18242 | Cathepsin D | 45 | 5.0 | 11.7 | 0.43 | 0.034 |
| Q9WV54 | Acid ceramidase | 45 | 2.7 | 14.0 | 0.19 | 0.00071 |
| P08121 | Collagen alpha-I (III) chain | 139 | 2.0 | 13.3 | 0.15 | 0.011 |
| G3UWC2 | N-acetylated alpha-linked acidic dipeptidase 2, isoform CRA_a | 87 | 1.7 | 11.7 | 0.14 | 0.0013 |
| Q62165 | Dystroglycan | 97 | 2.3 | 10.3 | 0.23 | 0.00043 |
| Q62356 | Follistatin-related protein 1 | 35 | 2.3 | 10.0 | 0.23 | 0.0019 |
| P11087 | Collagen alpha-1(I) chain | 138 | 2.0 | 10.3 | 0.19 | 0.011 |
| Q9EPL2 | Calsyntenin-1 | 109 | 0.7 | 11.3 | 0.06 | 0.0015 |
| O88207 | Collagen alpha-1(V) chain | 184 | 0 | 12.0 | 0 | 0.00048 |
| Q04857 | Collagen alpha-1 (VI) chain | 108 | 0 | 12.0 | 0 | 0.0000310 |
| O09159 | Lysosomal alpha-mannosidase | 115 | 0 | 10.0 | 0 | 0.0000652 |
| P47880 | Insulin-like growth factor-biding protein 6 | 25 | 0 | 10.0 | 0 | 0.00098 |
| Accession No. | Protein Name | MW (kDa) | iTRAQ Quantitation a | Fold Change | p-Value | |
|---|---|---|---|---|---|---|
| Med24 | MedC | |||||
| DAMPs | ||||||
| P63017 | Heat shock cognate 71-kDa protein | 71 | 68.0 | 63.7 | 1.07 | 0.41 |
| P11499 | Heat shock protein HSP 90-beta | 83 | 66.7 | 56.7 | 1.18 | 0.046 * |
| P60710 | Actin, cytoplasmic 1 | 42 | 49.0 | 36.0 | 1.36 | 0.027 * |
| Q3U2G2 | Heat shock 70 kDa protein 4 | 94 | 36.0 | 30.7 | 1.17 | 0.071 |
| P07901 | Heat shock protein HSP 90-alpha | 85 | 57.7 | 49.7 | 1.16 | 0.052 |
| P10107 | Annexin A1 | 39 | 30.0 | 29.0 | 1.03 | 0.75 |
| P10853 | Histone H2B type 1-F/J/L | 14 | 27.3 | 29.3 | 0.93 | 0.71 |
| P63038 | 60-kDa heat shock protein, mitochondrial | 61 | 32.3 | 20.3 | 1.59 | 0.22 |
| P16045 | Galectin-1 | 15 | 22.3 | 19 .0 | 1.18 | 0.067 |
| P07356 | Annexin A2 | 39 | 15.3 | 14.3 | 1.07 | 0.72 |
| Q61171 | Peroxiredoxin -2 | 22 | 15.0 | 13.7 | 1.10 | 0.33 |
| O08709 | Peroxiredoxin -6 | 25 | 11.7 | 9.7 | 1.21 | 0.18 |
| Q9JMH6 | Thioredoxin reductase 1, cytoplasmic | 67 | 10.7 | 8.3 | 1.28 | 0.28 |
| Q61699 | Heat shock protein 105 kDa | 96 | 12.3 | 8.0 | 1.54 | 0.25 |
| P16110 | Galectin-3 | 28 | 10.0 | 5.7 | 1.67 | 0.031 * |
| P99029 | Peroxiredoxin-5, mitochondrial | 22 | 9.0 | 6.3 | 1.42 | 0.091 |
| P63158 | High mobility group protein B1 | 25 | 5.7 | 4.7 | 1.22 | 0.66 |
| P15864 | Histone H1.2 | 21 | 3.0 | 7.0 | 0.43 | 0.0081 * |
| P10639 | Thioredoxin | 12 | 5.7 | 3.7 | 1.55 | 0.10 |
| P14211 | Calreticulin | 48 | 3.7 | 3.3 | 1.12 | 0.90 |
| DAMPs-related proteins | ||||||
| Q3V117 | ATP-citrate synthase | 121 | 28.0 | 21.0 | 1.33 | 0.025 * |
| P56480 | ATP synthase subunit beta, mitochondrial | 56 | 24.0 | 17.0 | 1.41 | 0.42 |
| Q03265 | ATP synthase subunit alpha, mitochondrial | 60 | 19.3 | 10.3 | 1.87 | 0.17 |
| Q9DB20 | ATP synthase subunit O, mitochondrial | 23 | 4.7 | 4.0 | 1.17 | 0.80 |
| Accession No. | Protein Name | MW (kDa) | Gene | iTRAQ Quantitation a |
|---|---|---|---|---|
| Med24 | ||||
| F8RG70 | Ribonucleoside-diphosphate reductase large subunit, ICP6 | 124 | UL39 | 33.7 |
| D3YP93 | Single-stranded DNA-binding protein, ICP8 | 128 | UL29 | 32.7 |
| L0N5H2 | DNA polymerase processivity subunit | 51 | UL42 | 19.3 |
| B9VQC5 | Transcriptional regulator ICP4 | 133 | α4 | 7.7 |
| D3YP82 | Major capsid protein, VP5 | 149 | UL19 | 14.3 |
| A0A0B5E4C5 | UL12, Exonuclease | 67 | UL12 | 13.0 |
| D3YPB9 | Multifunctional expression regulator, ICP27 | 55 | UL54 | 13.0 |
| A0A0F7GRC1 | Deoxyuridine triphosphatase | 39 | UL50 | 10.0 |
| F8RCG6 | Tegument protein UL37 | 121 | UL37 | 10.0 |
| A0A0B5E4D5 | Thymidine kinase | 41 | UL23 | 8.3 |
| A0A0B5E4F4 | VP13/VP14 | 74 | UL47 | 7.7 |
| D0V7M2 | Capsid protein VP23 | 34 | UL18 | 7.7 |
| A2I996 | Glycoprotein D | 43 | US6 | 6.7 |
| L0N6C7 | Envelope glycoprotein E | 59 | US8 | 5.3 |
| P10224 | Ribonucleoside-diphosphate reductase small subunit | 38 | UL40 | 6.0 |
| F8RCR4 | Tegument protein VP11/12 | 78 | UL46 | 5.3 |
| F8RDC2 | DNA packaging tegument protein UL25 | 63 | UL25 | 5.0 |
| L0N3F0 | Tegument protein VP22 | 32 | UL49 | 4.3 |
| D3YPB2 | Transactivating tegument protein VP16 | 54 | UL48 | 3.7 |
| A0A0F7CY40 | Nuclear egress membrane protein | 30 | UL34 | 4.7 |
| A0A0F7GTB2 | Capsid maturation protease | 67 | UL26 | 1.7 |
| A0A089YQM2 | Glycoprotein B | 100 | UL27 | 1.7 |
| A0A089YQK4 | UL2, Uracil-DNA glycosylase | 36 | UL2 | 1.3 |
| F8RFA5 | Tegument protein UL21 | 58 | UL21 | 1.3 |
| A0A0B5EE83 | Glycoprotein I | 41 | US7 | 1.0 |
| D3YP85 | Envelope glycoprotein H | 90 | UL22 | 0.7 |
| L0N3E3 | Envelope glycoprotein C | 55 | UL44 | 0.7 |
| L0N6J6 | DNA polymerase | 137 | UL30 | 0.7 |
| A0A089WZ14 | Type II membrane protein | 18 | UL45 | 0.7 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tada, S.; Hamada, M.; Yura, Y. Proteomic Analysis of Secretomes of Oncolytic Herpes Simplex Virus-Infected Squamous Cell Carcinoma Cells. Cancers 2018, 10, 28. https://doi.org/10.3390/cancers10020028
Tada S, Hamada M, Yura Y. Proteomic Analysis of Secretomes of Oncolytic Herpes Simplex Virus-Infected Squamous Cell Carcinoma Cells. Cancers. 2018; 10(2):28. https://doi.org/10.3390/cancers10020028
Chicago/Turabian StyleTada, Shinya, Masakazu Hamada, and Yoshiaki Yura. 2018. "Proteomic Analysis of Secretomes of Oncolytic Herpes Simplex Virus-Infected Squamous Cell Carcinoma Cells" Cancers 10, no. 2: 28. https://doi.org/10.3390/cancers10020028
APA StyleTada, S., Hamada, M., & Yura, Y. (2018). Proteomic Analysis of Secretomes of Oncolytic Herpes Simplex Virus-Infected Squamous Cell Carcinoma Cells. Cancers, 10(2), 28. https://doi.org/10.3390/cancers10020028