Higher CCL22+ Cell Infiltration is Associated with Poor Prognosis in Cervical Cancer Patients

 ,
,  and
and
Abstract
:1. Introduction
2. Results
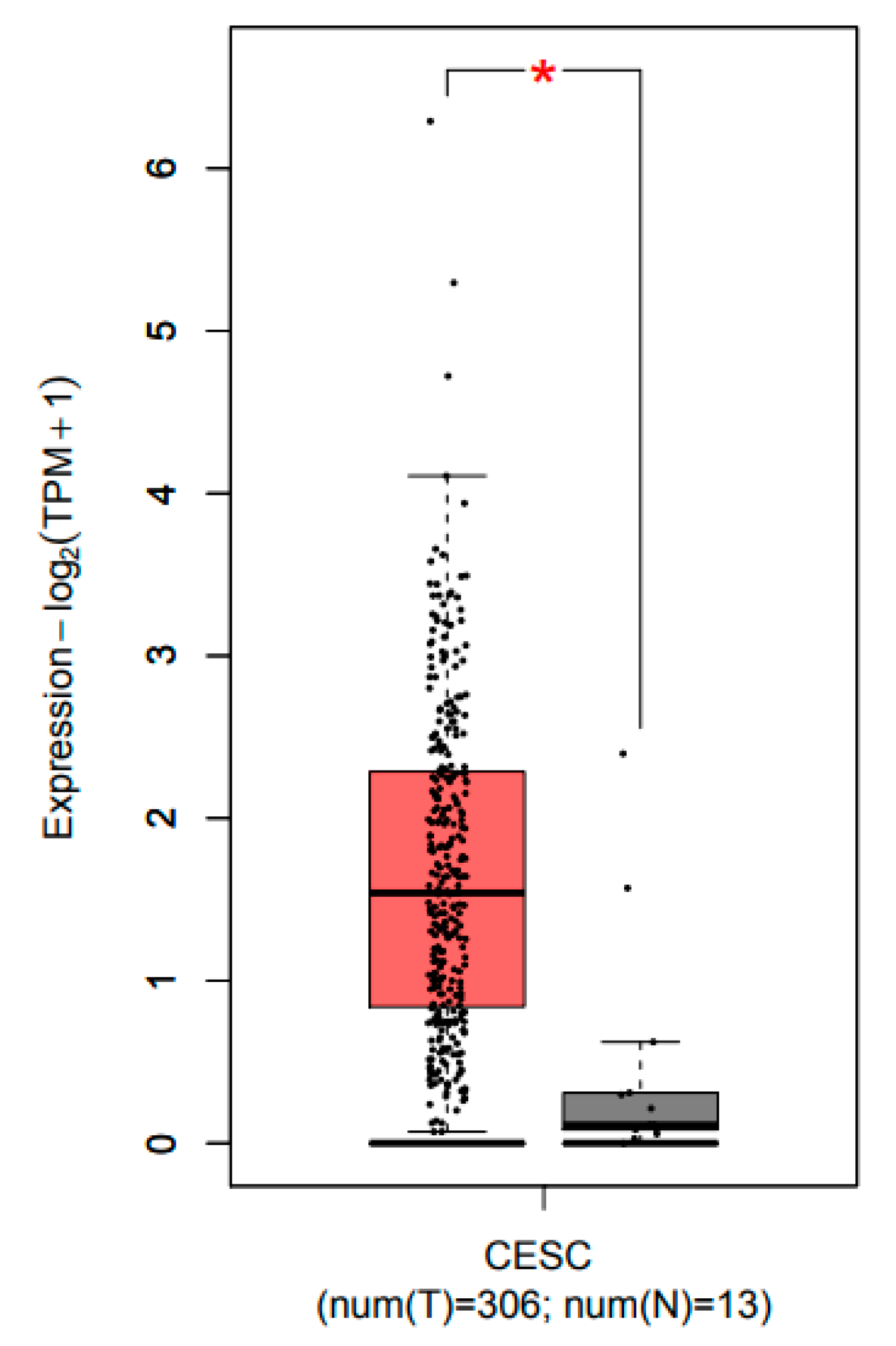
2.1. CCL22 Was Overexpressed in Cervical Squamous Cell Carcinoma and Endocervical Adenocarcinoma (CESC)
2.2. The Association between the IRS of CCL22 in CC Cells, Infiltrating CCL22+ Cell and FOXP3+ Cell Counts with Clinical Characteristics
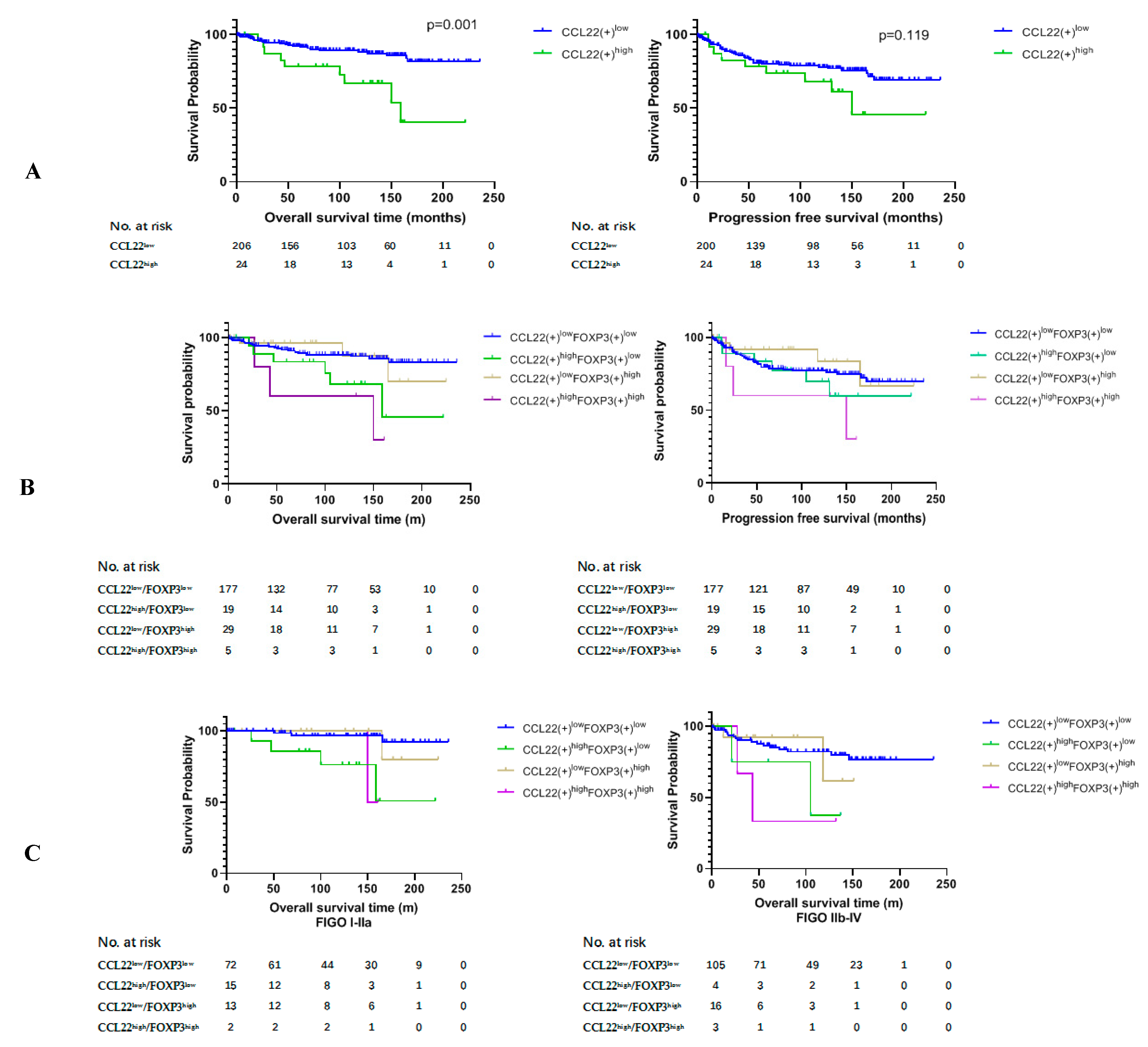
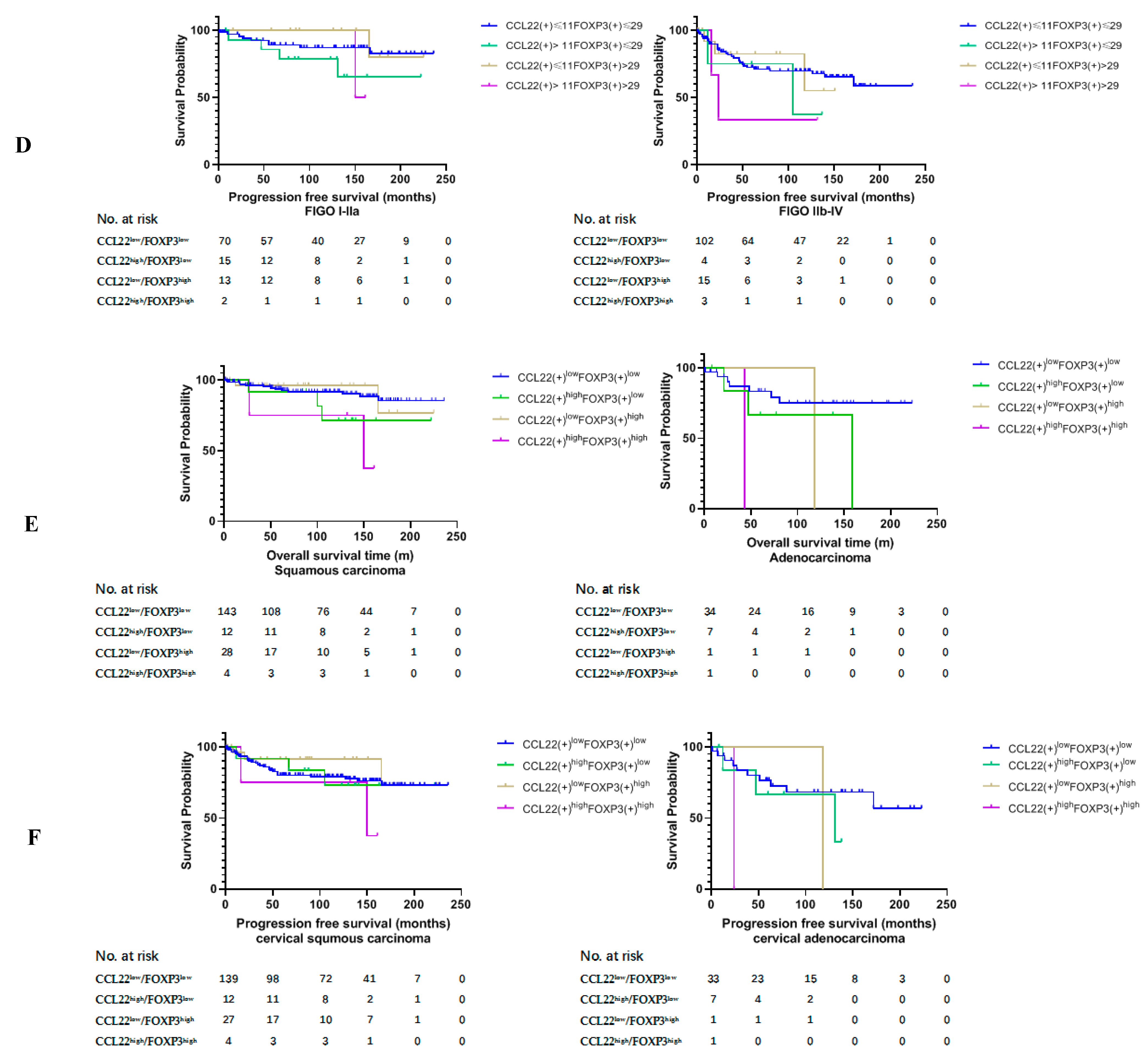
2.3. Higher Number of CCL22+ Cells Predicts Poor Prognosis of Patients with Cervical Cancer
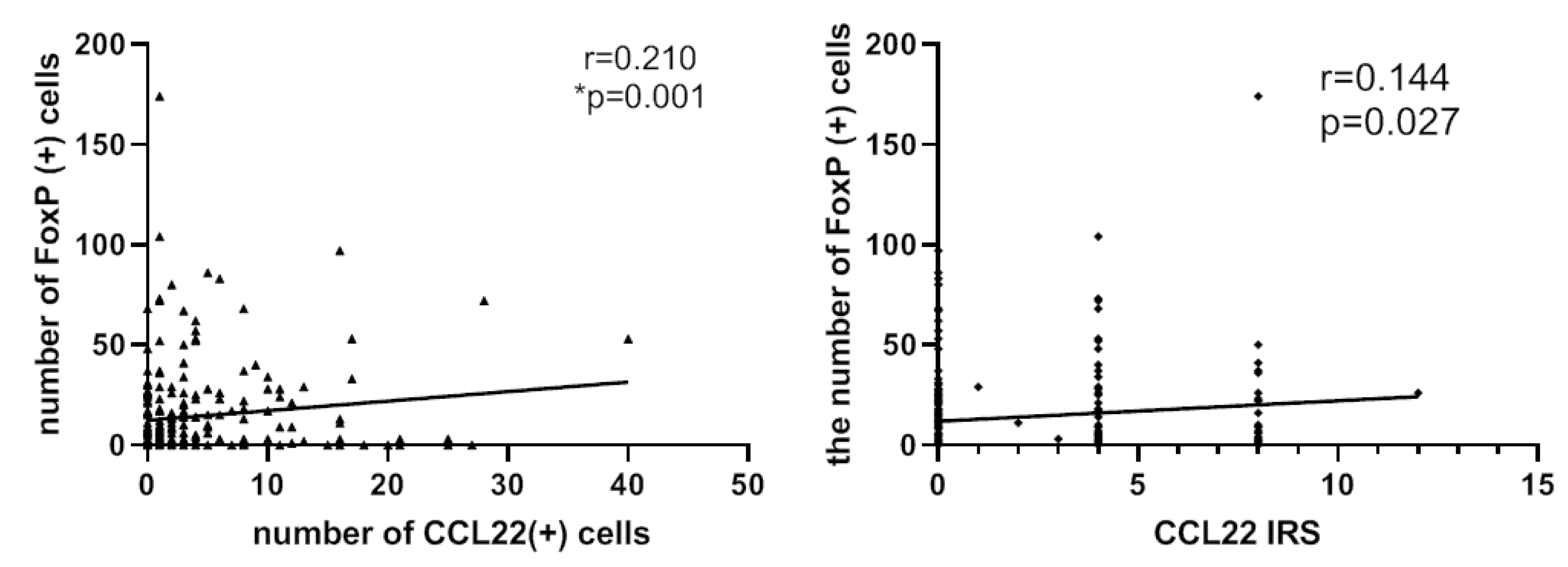
2.4. Correlation Analysis between CCL22+ Expression and FOXP3+ Cells
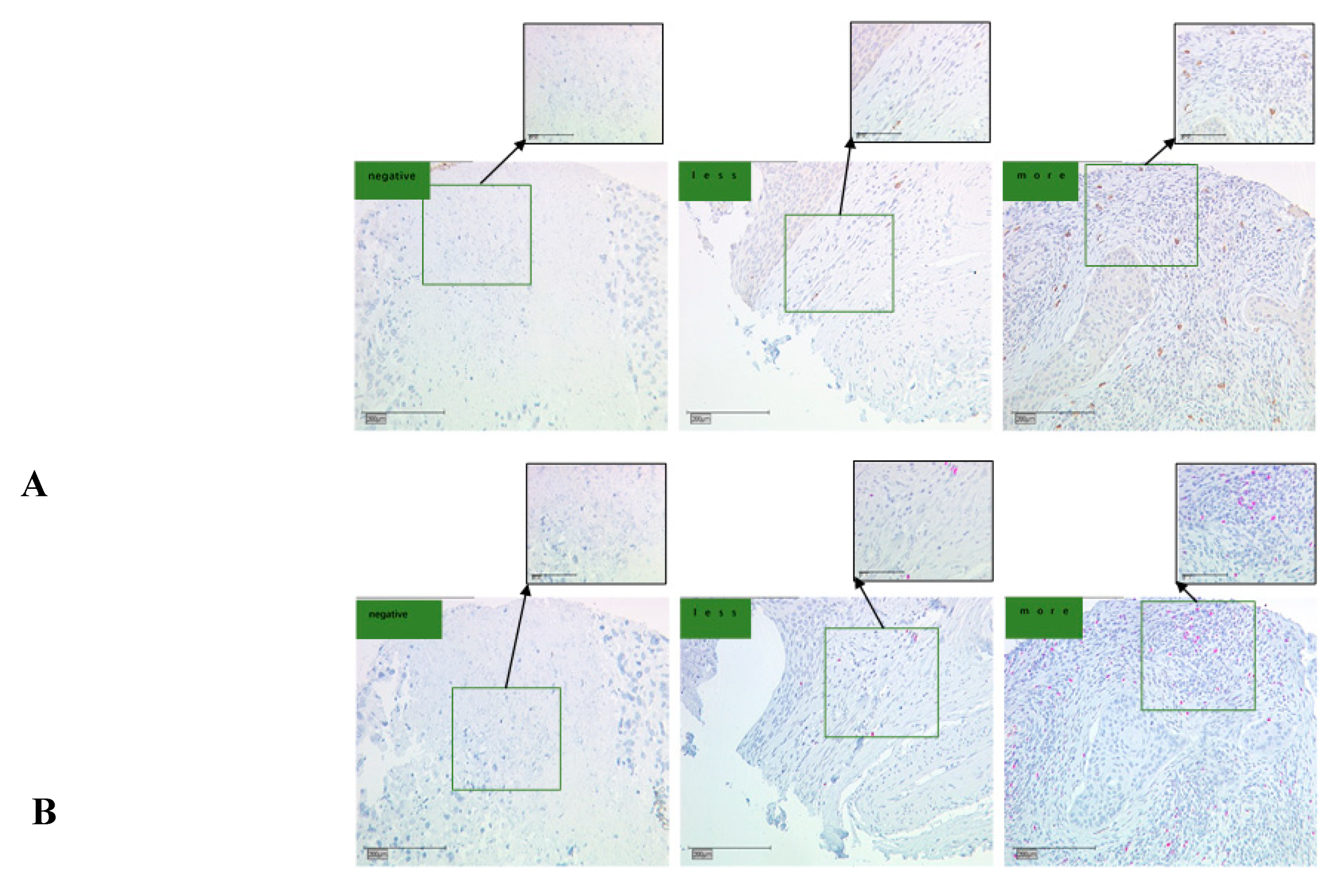
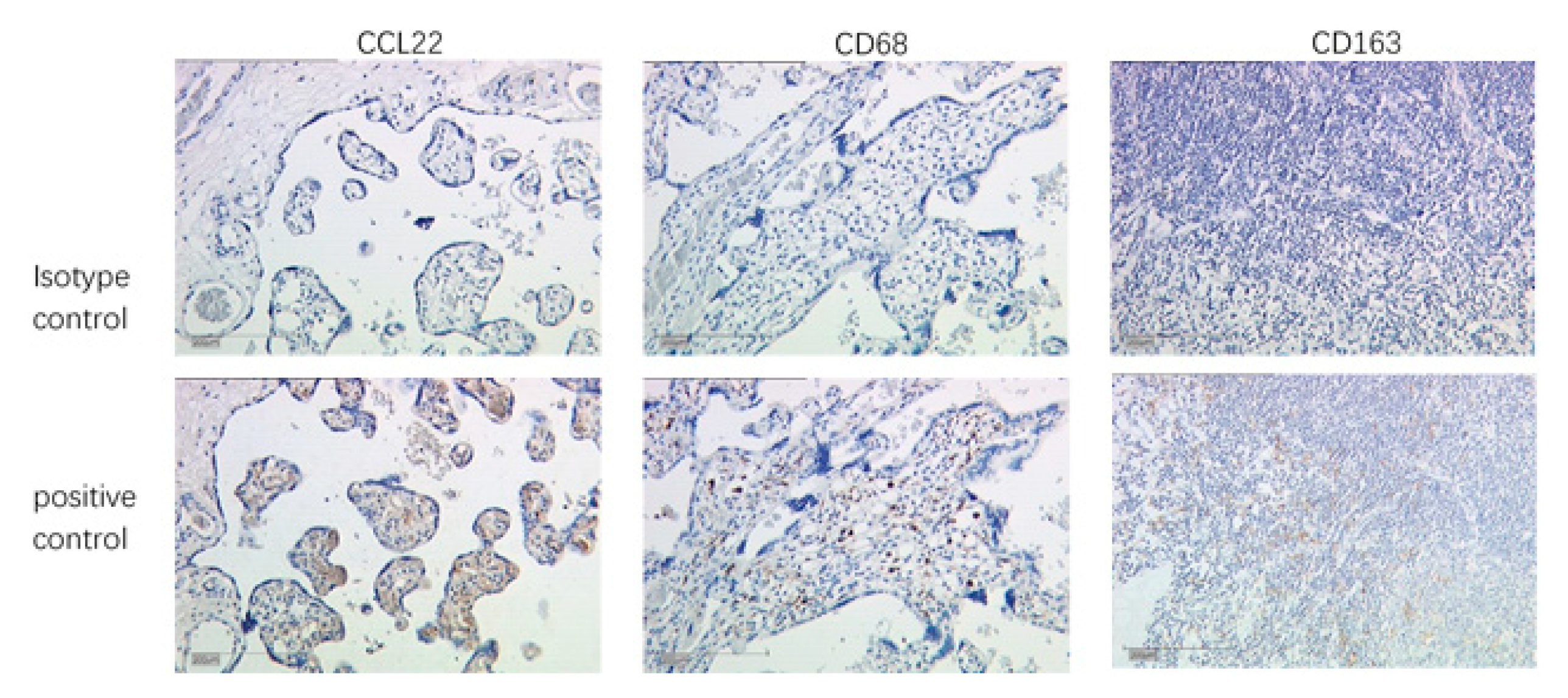
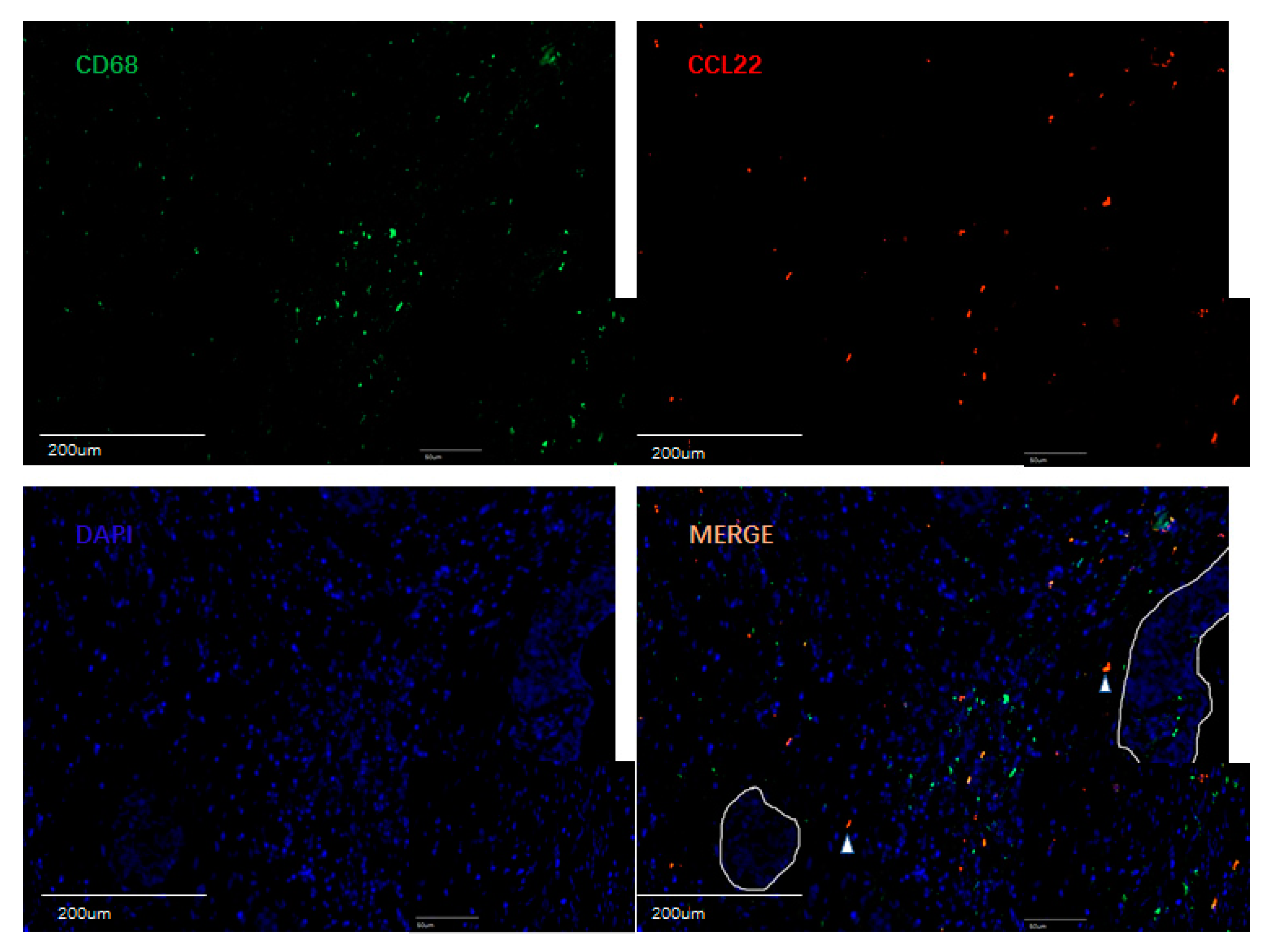
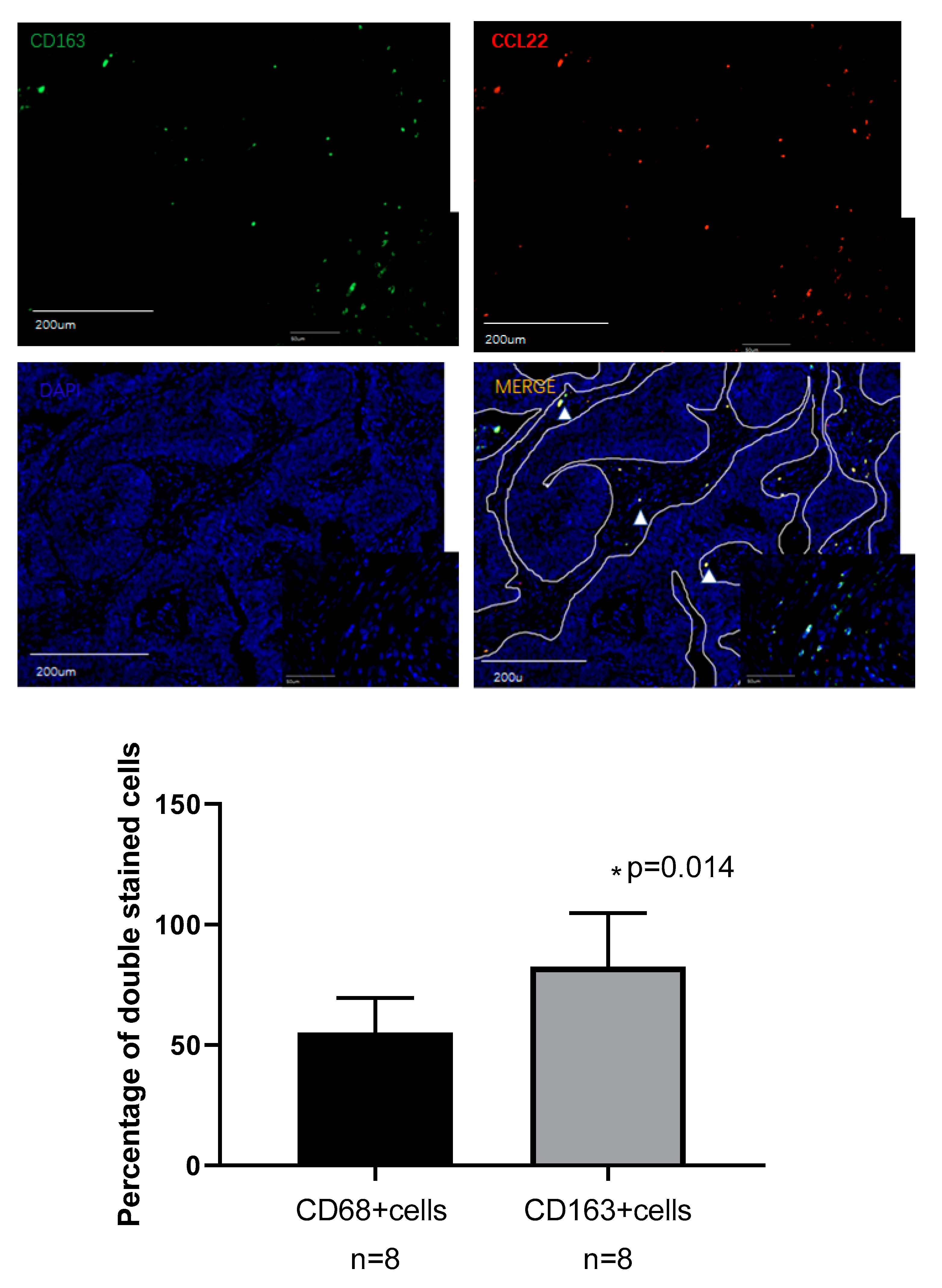
2.5. Identification of CCL22 Expressing Cells in Cervical Cancer
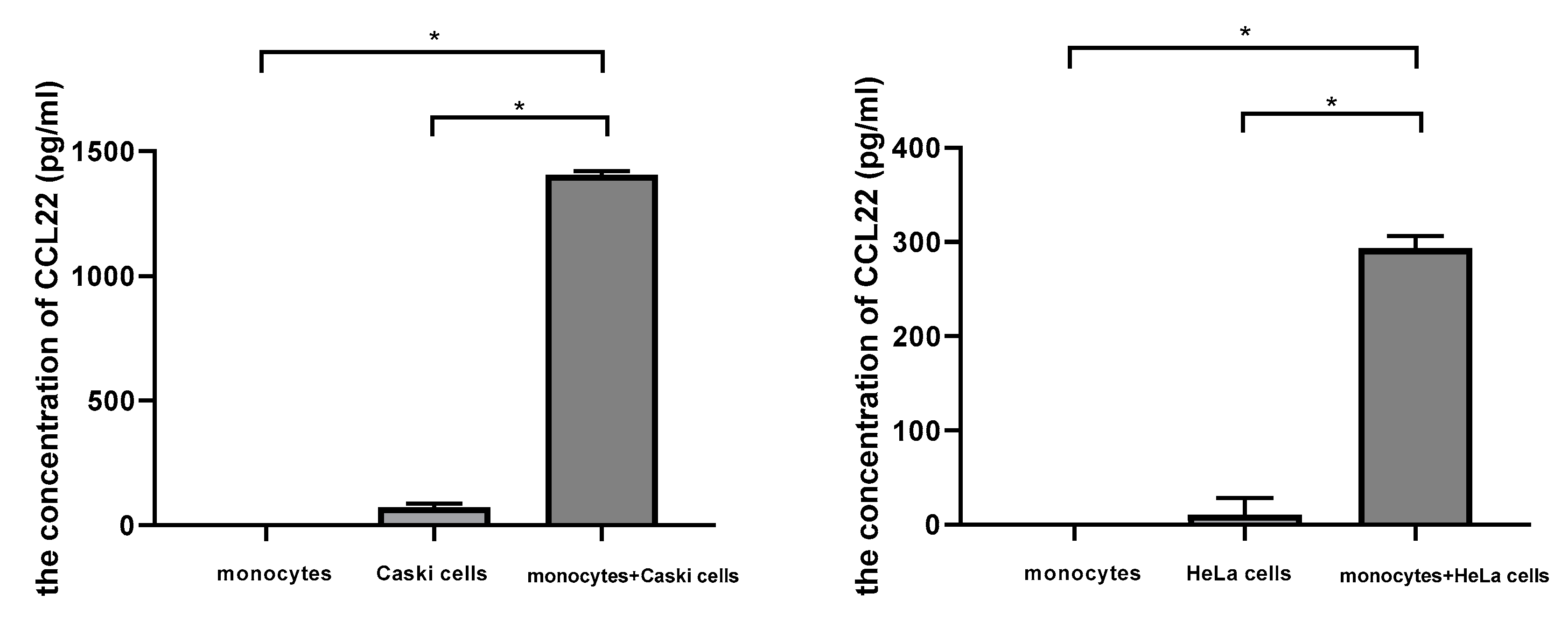
2.6. Cervical Cancer Cells Induced CCL22 in Monocytes
3. Materials and Methods
3.1. Bioinformatics
3.2. Clinical Sample
3.3. Tissue Microarray Construction
3.4. Immunohistochemistry
3.4.1. Evaluation of CCL22+ Cells as Macrophages
3.4.2. Evaluation of CCL22+ Cells as M2-Like Macrophages
3.5. Cell Coculture
3.6. Elisa
3.7. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ch, P.N.; Gurram, L.; Chopra, S.; Mahantshetty, U. The management of locally advanced cervical cancer. Curr. Opin. Oncol. 2018, 30, 323–329. [Google Scholar] [CrossRef]
- Kessler, T.A. Cervical Cancer: Prevention and Early Detection. Semin. Oncol. Nurs. 2017, 33, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Walboomers, J.M.; Jacobs, M.V.; Manos, M.M.; Bosch, F.X.; Kummer, J.A.; Shah, K.V.; Snijders, P.J.F.; Peto, J.; Meijer, C.J.L.M.; Munoz, N. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J. Pathol. 1999, 189, 12–19. [Google Scholar] [CrossRef]
- Clifford, G.M.; Tully, S.; Franceschi, S. Carcinogenicity of Human Papillomavirus (HPV) Types in HIV-Positive Women: A Meta-Analysis From HPV Infection to Cervical Cancer. Clin. Infect. Dis. 2017, 64, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Feldman, C.H.; Liu, J.; Feldman, S.; Solomon, D.H.; Kim, S.C. Risk of high-grade cervical dysplasia and cervical cancer in women with systemic lupus erythematosus receiving immunosuppressive drugs. Lupus 2017, 26, 682–689. [Google Scholar] [CrossRef]
- Duenas-Gonzalez, A.; Campbell, S. Global strategies for the treatment of early-stage and advanced cervical cancer. Curr. Opin. Obs. Gynecol. 2016, 28, 11–17. [Google Scholar] [CrossRef]
- Borcoman, E.; Le Tourneau, C. Pembrolizumab in cervical cancer: Latest evidence and clinical usefulness. Adv. Med. Oncol. 2017, 9, 431–439. [Google Scholar] [CrossRef]
- Toker, A.; Ohashi, P.S. Expression of costimulatory and inhibitory receptors in FOXP3(+) regulatory T cells within the tumor microenvironment: Implications for combination immunotherapy approaches. Adv. Cancer Res. 2019, 144, 193–261. [Google Scholar] [CrossRef]
- Miyara, M.; Sakaguchi, S. Natural regulatory T cells: Mechanisms of suppression. Trends Mol. Med. 2007, 13, 108–116. [Google Scholar] [CrossRef]
- Loddenkemper, C.; Hoffmann, C.; Stanke, J.; Nagorsen, D.; Baron, U.; Olek, S.; Huehn, J.; Ritz, J.-P.; Stein, H.; Kaufmann, A.M.; et al. Regulatory (FOXP3+) T cells as target for immune therapy of cervical intraepithelial neoplasia and cervical cancer. Cancer Sci. 2009, 100, 1112–1117. [Google Scholar] [CrossRef]
- Visser, J.; Nijman, H.W.; Hoogenboom, B.N.; Jager, P.; Van Baarle, D.; Schuuring, E.; Abdulahad, W.; Miedema, F.; Van Der Zee, A.G.; Daemen, T. Frequencies and role of regulatory T cells in patients with (pre)malignant cervical neoplasia. Clin. Exp. Immunol. 2007, 150, 199–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heusinkveld, M.; De Vos van Steenwijk, P.J.; Goedemans, R.; Ramwadhdoebe, T.H.; Gorter, A.; Welters, M.J.; Van Hall, T.; Van der Burg, S.H. M2 macrophages induced by prostaglandin E2 and IL-6 from cervical carcinoma are switched to activated M1 macrophages by CD4+ Th1 cells. J. Immunol. 2011, 187, 1157–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierra-Filardi, E.; Nieto, C.; Dominguez-Soto, A.; Barroso, R.; Sanchez-Mateos, P.; Puig-Kroger, A.; Lopez-Bravo, M.; Joven, J.; Ardavin, C.; Rodrguez-Fernandez, J.L.; et al. CCL2 shapes macrophage polarization by GM-CSF and M-CSF: Identification of CCL2/CCR2-dependent gene expression profile. J. Immunol. 2014, 192, 3858–3867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, J.K.; Takeda, K.; Akira, S.; Schreiber, R.D. Interleukin-10 receptor signaling through the JAK-STAT pathway. Requirement for two distinct receptor-derived signals for anti-inflammatory action. J. Biol. Chem. 1999, 274, 16513–16521. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Bae, J.S. Tumor-Associated Macrophages and Neutrophils in Tumor Microenvironment. Mediat. Inflamm. 2016, 2016, 6058147. [Google Scholar] [CrossRef] [Green Version]
- Davidson, B.; Goldberg, I.; Kopolovic, J. Inflammatory response in cervical intraepithelial neoplasia and squamous cell carcinoma of the uterine cervix. Pathol. Res. Pract. 1997, 193, 491–495. [Google Scholar] [CrossRef]
- Rigo, A.; Gottardi, M.; Zamo, A.; Mauri, P.; Bonifacio, M.; Krampera, M.; Damiani, E.; Pizzolo, G.; Vinante, F. Macrophages may promote cancer growth via a GM-CSF/HB-EGF paracrine loop that is enhanced by CXCL12. Mol. Cancer 2010, 9, 273. [Google Scholar] [CrossRef] [Green Version]
- Punt, S.; Thijssen, V.L.; Vrolijk, J.; De Kroon, C.D.; Gorter, A.; Jordanova, E.S. Galectin-1, -3 and -9 Expression and Clinical Significance in Squamous Cervical Cancer. PLoS ONE 2015, 10, e0129119. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.J.; Han, L.F.; Wu, X.G.; Wei, W.F.; Wu, L.F.; Yi, H.Y.; Yan, R.-M.; Bai, X.; Zhong, M.; Yu, Y.; et al. Clinical Significance of CD163+ and CD68+ Tumor-associated Macrophages in High-risk HPV-related Cervical Cancer. J. Cancer 2017, 8, 3868–3875. [Google Scholar] [CrossRef] [Green Version]
- Hartwig, T.; Montinaro, A.; Von Karstedt, S.; Sevko, A.; Surinova, S.; Chakravarthy, A.; Taraborrelli, L.; Draber, P.; Lafont, E.; Vargas, F.A.; et al. The TRAIL-Induced Cancer Secretome Promotes a Tumor-Supportive Immune Microenvironment via CCR2. Mol. Cell 2017, 65, 730–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepique, A.P.; Daghastanli, K.R.; Cuccovia, I.M.; Villa, L.L. HPV16 tumor associated macrophages suppress antitumor T cell responses. Clin. Cancer Res. 2009, 15, 4391–4400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilders, C.G.; Munoz, I.M.; Nooyen, Y.; Fleuren, G.J. Altered HLA expression by metastatic cervical carcinoma cells as a factor in impaired immune surveillance. Gynecol. Oncol. 1995, 57, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Walentowicz-Sadlecka, M.; Koper, A.; Krystyna, G.; Koper, K.; Basta, P.; Mach, P.; Skret-Magierlo, J.; Dutsch-Wicherek, M.; Sikora, J.; Grabiec, M.; et al. The analysis of metallothionein immunoreactivity in stromal fibroblasts and macrophages in cases of uterine cervical carcinoma with respect to both the local and distant spread of the disease. Am. J. Reprod. Immunol. 2013, 70, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Melichar, B.; Solichova, D.; Freedman, R.S. Neopterin as an indicator of immune activation and prognosis in patients with gynecological malignancies. Int. J. Gynecol. Cancer 2006, 16, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Bolpetti, A.; Silva, J.S.; Villa, L.L.; Lepique, A.P. Interleukin-10 production by tumor infiltrating macrophages plays a role in Human Papillomavirus 16 tumor growth. BMC Immunol. 2010, 11, 27. [Google Scholar] [CrossRef] [Green Version]
- Godiska, R.; Chantry, D.; Raport, C.J.; Sozzani, S.; Allavena, P.; Leviten, D.; Mantovani, A.; Gray, P.W. Human macrophage-derived chemokine (MDC), a novel chemoattractant for monocytes, monocyte-derived dendritic cells, and natural killer cells. J. Exp. Med. 1997, 185, 1595–1604. [Google Scholar] [CrossRef] [Green Version]
- Chang, M.; McNinch, J.; Elias, C., 3rd; Manthey, C.L.; Grosshans, D.; Meng, T.; Boone, T.; Andrew, D. Molecular cloning and functional characterization of a novel CC chemokine, stimulated T cell chemotactic protein (STCP-1) that specifically acts on activated T lymphocytes. J. Biol. Chem. 1997, 272, 25229–25237. [Google Scholar] [CrossRef] [Green Version]
- Schaniel, C.; Pardali, E.; Sallusto, F.; Speletas, M.; Ruedl, C.; Shimizu, T.; Seidl, T.; Andersson, J.; Melchers, F.; Rolink, A.G.; et al. Activated murine B lymphocytes and dendritic cells produce a novel CC chemokine which acts selectively on activated T cells. J. Exp. Med. 1998, 188, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Gray, P.A.; Van Damme, J.; Sozzani, S. Macrophage-derived chemokine (MDC). J. Leukoc. Biol. 2000, 68, 400–404. [Google Scholar]
- Curiel, T.J.; Coukos, G.; Zou, L.; Alvarez, X.; Cheng, P.; Mottram, P.; Evdemon-Hogan, M.; Conejo-Garcia, J.R.; Zhang, L.; Burow, M.; et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat. Med. 2004, 10, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Yang, C.; Wang, S.; Shi, D.; Zhang, C.; Lin, X.; Xiong, B. M2 macrophages confer resistance to 5-fluorouracil in colorectal cancer through the activation of CCL22/PI3K/AKT signaling. OncoTargets Ther. 2019, 12, 3051–3063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Li, Y.; Wei, X.; Zhang, Q.; Jia, H.; Quan, S.; Cao, D.; Wang, L.; Yang, T.; Zhao, J.; et al. Negative immune factors might predominate local tumor immune status and promote carcinogenesis in cervical carcinoma. Virol. J. 2017, 14, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedraza-Brindis, E.J.; Sanchez-Reyes, K.; Hernandez-Flores, G.; Bravo-Cuellar, A.; Jave-Suarez, L.F.; Aguilar-Lemarroy, A.; Gomez-Lomeli, P.; Lpez-Lopez, B.A.; Ortiz-Lazareno, P.C. Culture supernatants of cervical cancer cells induce an M2 phenotypic profile in THP-1 macrophages. Cell. Immunol. 2016, 310, 42–52. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Vissers, J.L.; Hartgers, F.C.; Lindhout, E.; Teunissen, M.B.; Figdor, C.G.; Adema, G.J. Quantitative analysis of chemokine expression by dendritic cell subsets in vitro and in vivo. J. Leukoc. Biol. 2001, 69, 785–793. [Google Scholar]
- Jiang, S.; Yang, Y.; Fang, M.; Li, X.; Yuan, X.; Yuan, J. Co-evolution of tumor-associated macrophages and tumor neo-vessels during cervical cancer invasion. Oncol. Lett. 2016, 12, 2625–2631. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Q.; Liu, F.F.; Zhang, X.M.; Guo, X.J.; Ren, M.J.; Fu, L. Tumor secretion of CCL22 activates intratumoral Treg infiltration and is independent prognostic predictor of breast cancer. PLoS ONE 2013, 8, e76379. [Google Scholar] [CrossRef]
- Zhou, M.; Bracci, P.M.; McCoy, L.S.; Hsuang, G.; Wiemels, J.L.; Rice, T.; Zheng, S.; Kelsey, K.T.; Wrensch, M.R.; Wiencke, J.K. Serum macrophage-derived chemokine/CCL22 levels are associated with glioma risk, CD4 T cell lymphopenia and survival time. Int. J. Cancer 2015, 137, 826–836. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; He, H.; Liu, H.; Cao, Y.; Li, R.; Zhang, H.; Li, H.; Shen, Z.; Qin, J.; Xu, J. C-C motif chemokine 22 predicts postoperative prognosis and adjuvant chemotherapeutic benefits in patients with stage II/III gastric cancer. Oncoimmunology 2018, 7, e1433517. [Google Scholar] [CrossRef]
- Huang, Y.H.; Chang, C.Y.; Kuo, Y.Z.; Fang, W.Y.; Kao, H.Y.; Tsai, S.T.; Wu, L.-W. Cancer-associated fibroblast-derived interleukin-1beta activates the pro-tumor CCL22 signaling in head and neck cancer. Cancer Sci. 2019, 110, 2783–2793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furudate, S.; Fujimura, T.; Kambayashi, Y.; Kakizaki, A.; Hidaka, T.; Aiba, S. Immunomodulatory Effect of Imiquimod Through CCL22 Produced by Tumor-associated Macrophages in B16F10 Melanomas. Anticancer Res. 2017, 37, 3461–3471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.Y.; Zhao, J.; Yang, T.; Wang, L.; Pei, M.L.; Tian, S.J.; Yu, Y.; Yang, X.-F. Aberrant Expressions of Immune Factors Facilitate the Disequilibrium of Immune Status in Cervical Cancer. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2016, 38, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Nanbu, U.; Noguchi, H.; Harada, Y.; Kumamoto, K.; Sasaguri, Y.; Nakayama, T. Macrophage CCL22 expression in the tumor microenvironment and implications for survival in patients with squamous cell carcinoma of the tongue. J. Oral Pathol. Med. 2019, 48, 677–685. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Characteristics | Total (%) | IRS of CCL22 in CC Cells | p | Quantity of CCL22+ (n%) | p | Quantity of FOXP3+ (n%) | p | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Low (IRS < 4) | High (IRS ≥ 4) | Small (n ≤ 11) | Large (n > 11) | Small (n ≤ 29) | Large (n > 29) | |||||
| All cases | 230 | 134 (58.3%) | 96 (41.7%) | − | 206 (89.6%) | 24 (10.4%) | − | 196 (85.2%) | 34 (14.8%) | − |
| Age (year) | ||||||||||
| ≤50 | 132 (57.4%) | 80 (59.7%) | 52 (54.2%) | 0.403 | 114 (55.3%) | 18 (75.0%) | 0.065 | 115 (58.7%) | 17 (50.0%) | 0.345 |
| >50 | 98 (42.6%) | 54 (40.3%) | 44 (45.8%) | 92 (44.7%) | 6 (25.0%) | 81 (41.3%) | 17 (50.0%) | |||
| Tumor Size (cm) | ||||||||||
| <2 | 2 (0.9%) | 2 (1.5%) | 0 (0%) | 0.080 | 2 (1.0%) | 0 (0%) | 0.090 | 2 (1.0%) | 0 (0%) | 0.905 |
| 2–4 | 120 (52.2%) | 75 (56%) | 45 (46.9%) | 103 (50.0%) | 17 (70.8%) | 102 (52%) | 18 (52.9%) | |||
| >4 | 107 (46.5%) | 57 (42.5%) | 51 (53.1%) | 101 (49.0%) | 7 (29.2%) | 92 (46.9%) | 16 (47.1%) | |||
| PN | ||||||||||
| Without lymph node metastasis | 139 (60.4%) | 81 (60.4%) | 58 (60.4%) | 0.996 | 118 (57.3%) | 21 (87.5%) | 0.004 * | 122 (62.2%) | 17 (50.0%) | 0.178 |
| With lymph node metastasis | 91 (39.6%) | 53 (39.6%) | 38 (39.6%) | 88 (42.7%) | 3 (12.5%) | 74 (37.8%) | 17 (50.0%) | |||
| PM | ||||||||||
| Without metastasis | 219 (95.2%) | 127 (94.8%) | 92 (95.8%) | 0.709 | 195 (94.7%) | 24 (100%) | 0.115 | 187 (95.4%) | 32 (94.1%) | 0.752 |
| With metastasis | 11 (4.8%) | 7 (5.2%) | 4 (4.2%) | 11 (5.3%) | 0 (0%) | 9 (4.6%) | 2 (5.9%) | |||
| FIGO | ||||||||||
| I–IIA | 102 (44.3%) | 62 (46.3%) | 40 (41.7%) | 0.488 | 85 (41.3%) | 17 (70.8%) | 0.006 * | 87 (60.2%) | 15 (50.0%) | 0.265 |
| IIB–IV | 128 (55.7%) | 55 (53.7%) | 40 (58.3%) | 121 (58.7%) | 7 (29.2%) | 109 (39.8%) | 19 (50.0%) | |||
| Grade | ||||||||||
| 1 | 19 (8.3%) | 10 (7.8%) | 9 (9.5%) | 0.676 | 17 (8.9%) | 2 (6.1%) | 0.653 | 13 (6.7%) | 4 (12.1%) | 0.243 |
| 2 | 129 (56.1%) | 72 (56.3%) | 58 (60.0%) | 112 (58.9%) | 17 (51.5%) | 123 (63.1%) | 12 (36.4%) | |||
| 3 | 75 (32.6%) | 46 (35.9%) | 29 (30.5%) | 61 (32.1%) | 14 (42.4%) | 59 (30.3%) | 17 (51.50%) | |||
| NA | 7 (3.0%) | |||||||||
| Subtype | ||||||||||
| Squamous carcinoma | 187 (81.3%) | 95 (70.9%) | 92 (95.8%) | 0.001 * | 171 (83.0%) | 16 (66.7%) | 0.091 | 155 (79.1%) | 32 (94.1%) | 0.038 * |
| Adenocarcinoma | 43 (18.7%) | 39 (29.1%) | 4 (4.2%) | 35 (17.0%) | 8 (33.3%) | 41 (20.9%) | 2 (5.9%) | |||
| Progression | ||||||||||
| No | 175 (76.1%) | 99 (75.0%) | 76 (80.0%) | 0.377 | 160 (78.8%) | 15 (62.5%) | 0.072 | 149 (76.8%) | 26 (78.8%) | 0.802 |
| Yes | 52 (22.6%) | 33 (25.0%) | 19 (20.0%) | 43 (21.2%) | 9 (37.5%) | 45 (23.2%) | 7 (21.2%) | |||
| NA | 3 (1.3%) | |||||||||
| Therapeutic Strategy | ||||||||||
| Surgery | 96 (41.7%) | 59 (44.0%) | 37 (38.5%) | 0.675 | 79 (38.3%) | 17 (70.8%) | 0.007 * | 83 (42.3%) | 13 (38.2%) | 0.494 |
| Preoperative chemoradiotherapy | 6 (2.6%) | 3 (2.2%) | 3 (3.1%) | 6 (2.9%) | 0 (0.0%) | 4 (2.0%) | 2 (5.9%) | |||
| Chemoradiotherapy | 128 (55.7%) | 72 (53.7%) | 56 (58.3%) | 121 (58.7%) | 7 (29.2%) | 109 (55.6%) | 19 (55.9%) | |||
| Survival State | ||||||||||
| Alive | 198 (86.1%) | 114 (85.1%) | 84 (87.5%) | 0.600 | 183 (88.8%) | 15 (62.5%) | 0.002 * | 170 (86.7%) | 28 (82.4%) | 0.507 |
| Death | 32 (13.9%) | 20 (14.9%) | 12 (12.5%) | 23 (11.2%) | 9 (37.5%) | 26 (13.3%) | 6 (17.6%) | |||
| Group | CCL22low FOXP3low | CCL22high FOXP3low | CCL22low FOXP3high | CCL22high FOXP3high | |||||
|---|---|---|---|---|---|---|---|---|---|
| X2 | p-Value | X2 | p-Value | X2 | p-Value | X2 | p-Value | ||
| Log Rank (Mantel–Cox) | CCL22low FOXP3low | − | − | 5.250 | 0.022 * | 0.005 | 0.946 | 8.732 | 0.003 * |
| CCL22high FOXP3low | 5.250 | 0.022 * | − | − | 2.466 | 0.116 | 0.469 | 0.494 | |
| CCL22low FOXP3high | 0.005 | 0.946 | 2.466 | 0.116 | − | − | 5.486 | 0.019 * | |
| CCL22high FOXP3high | 8.732 | 0.003 * | 0.469 | 0.494 | 5.486 | 0.019 * | − | − | |
| Group | CCL22low FOXP3low | CCL22high FOXP3low | CCL22low FOXP3high | CCL22high FOXP3high | |||||
|---|---|---|---|---|---|---|---|---|---|
| X2 | p-Value | X2 | p-Value | X2 | p-Value | X2 | p-Value | ||
| Log Rank (Mantel–Cox) | CCL22low FOXP3low | − | − | 0.519 | 0.471 | 0.490 | 0.484 | 3.320 | 0.068 |
| CCL22high FOXP3low | 0.519 | 0.471 | − | − | 1.431 | 0.232 | 0.353 | 0.552 | |
| CCL22low FOXP3high | 0.490 | 0.484 | 1.431 | 0.232 | − | − | 4.069 | 0.044 * | |
| CCL22high FOXP3high | 3.320 | 0.068 | 0.353 | 0.552 | 4.069 | 0.044 * | − | − | |
| Group | CCL22low FOXP3low | CCL22high FOXP3low | CCL22low FOXP3high | CCL22high FOXP3high | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| X2 | p-Value | X2 | p-Value | X2 | p-Value | X2 | p-Value | |||
| I–IIa | Log Rank (Mantel–Cox) | CCL22low FOXP3low | − | − | 10.547 | 0.001 * | 0.263 | 0.608 | 6.107 | 0.013 * |
| CCL22high FOXP3low | 10.547 | 0.001 * | − | − | 3.122 | 0.077 | 0.010 | 0.920 | ||
| CCL22low FOXP3high | 0.263 | 0.608 | 3.122 | 0.077 | − | − | 3.000 | 0.083 | ||
| CCL22high FOXP3high | 6.107 | 0.013 * | 0.010 | 0.920 | 3.000 | 0.083 | − | − | ||
| IIb-IV | Log Rank (Mantel–Cox) | CCL22low FOXP3low | − | − | 2.575 | 0.109 | 0.013 | 0.911 | 4.761 | 0.029 * |
| CCL22high FOXP3low | 2.575 | 0.109 | − | − | 0.896 | 0.344 | 0.119 | 0.730 | ||
| CCL22low FOXP3high | 0.013 | 0.911 | 0.896 | 0.344 | − | − | 1.891 | 0.169 | ||
| CCL22high FOXP3high | 4.761 | 0.029 * | 0.119 | 0.730 | 1.891 | 0.169 | − | − | ||
| Group | CCL22low FOXP3low | CCL22high FOXP3low | CCL22low FOXP3high | CCL22high FOXP3high | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| X2 | p-Value | X2 | p-Value | X2 | p-Value | X2 | p-Value | |||
| I–IIa | Log Rank (Mantel–Cox) | CCL22low FOXP3low | − | − | 1.967 | 0.161 | 0.301 | 0.583 | 1.413 | 0.235 |
| CCL22high FOXP3low | 1.967 | 0.161 | − | − | 2.888 | 0.089 | 0.029 | 0.864 | ||
| CCL22low FOXP3high | 0.301 | 0.583 | 2.888 | 0.089 | − | − | 3.000 | 0.083 | ||
| CCL22high FOXP3high | 1.413 | 0.235 | 0.029 | 0.864 | 3.000 | 0.083 | − | − | ||
| IIb–IV | Log Rank (Mantel–Cox) | CCL22low FOXP3low | − | − | 0.603 | 0.437 | 0.004 | 0.950 | 2.515 | 0.113 |
| CCL22high FOXP3low | 0.603 | 0.437 | − | − | 0.367 | 0.544 | 0.119 | 0.730 | ||
| CCL22low FOXP3high | 0.004 | 0.950 | 0.367 | 0.544 | − | − | 1.160 | 0.282 | ||
| CCL22high FOXP3high | 2.515 | 0.113 | 0.119 | 0.730 | 1.160 | 0.282 | − | − | ||
| Group | CCL22low FOXP3low | CCL22high FOXP3low | CCL22low FOXP3high | CCL22high FOXP3high | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| X2 | p-Value | X2 | p-Value | X2 | p-Value | X2 | p-Value | |||
| Squamous carcinoma | Log Rank (Mantel–Cox) | CCL22low FOXP3low | − | − | 2.133 | 0.144 | 0.011 | 0.917 | 5.399 | 0.020 |
| CCL22high FOXP3low | 2.133 | 0.144 | − | − | 1.387 | 0.239 | 0.208 | 0.648 | ||
| CCL22low FOXP3high | 0.011 | 0.917 | 1.387 | 0.239 | − | − | 5.139 | 0.023 * | ||
| CCL22high FOXP3high | 5.399 | 0.020 * | 0.208 | 0.648 | 5.139 | 0.023 | − | − | ||
| Adenocarcinoma | Log Rank (Mantel–Cox) | CCL22low FOXP3low | − | − | 1.802 | 0.179 | 1.363 | 0.243 | 4.093 | 0.043 * |
| CCL22high FOXP3low | 1.802 | 0.179 | − | − | 0.264 | 0.608 | 1.824 | 0.177 | ||
| CCL22low FOXP3high | 1.363 | 0.243 | 0.264 | 0.608 | − | − | 1.000 | 0.317 | ||
| CCL22high FOXP3high | 4.093 | 0.043 * | 1.824 | 0.177 | 1.000 | 0.317 | − | − | ||
| Group | CCL22low FOXP3low | CCL22high FOXP3low | CCL22low FOXP3high | CCL22high FOXP3high | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| X2 | p-Value | X2 | p-Value | X2 | p-Value | X2 | p-Value | |||
| Squamous carcinoma | Log Rank (Mantel–Cox) | CCL22low FOXP3low | − | − | 0.011 | 0.918 | 0.739 | 0.390 | 1.327 | 0.249 |
| CCL22high FOXP3low | 0.011 | 0.918 | − | − | 0.687 | 0.407 | 0.246 | 0.620 | ||
| CCL22low FOXP3high | 0.739 | 0.390 | 0.687 | 0.407 | − | − | 3.438 | 0.064 | ||
| CCL22high FOXP3high | 1.327 | 0.249 | 0.246 | 0.620 | 3.438 | 0.064 | − | − | ||
| Adenocarcinoma | Log Rank (Mantel–Cox) | CCL22low FOXP3low | − | − | 0.770 | 0.380 | 0.796 | 0.372 | 4.175 | 0.041 * |
| CCL22high FOXP3low | 0.770 | 0.380 | − | − | 0.264 | 0.608 | 1.824 | 0.177 | ||
| CCL22low FOXP3high | 0.796 | 0.372 | 0.264 | 0.608 | − | − | 1.000 | 0.317 | ||
| CCL22high FOXP3high | 4.175 | 0.041 * | 1.824 | 0.177 | 1.000 | 0.317 | − | − | ||
| Clinicopathological Variables | Univariate Analysis | Multivariate Analysis | |||
|---|---|---|---|---|---|
| PFS | OS | PFS | OS (model 1) | OS (model 2) | |
| Age (≤50 years vs. >50 years) | |||||
| HR | 1.384 | 1.804 | - | - | - |
| 95%CI | 0.804–2.385 | 0.897–3.629 | - | - | - |
| p | 0.241 | 0.098 | - | - | - |
| Tumor Size (≤4 cm vs. >4 cm) | |||||
| HR | 3.063 | 4.188 | 2.652 | 5.564 | 5.487 |
| 95%CI | 1.710–5.486 | 1.924–9.112 | 1.063–6.617 | 1.538–20.130 | 1.504–20.011 |
| p | 0.001 * | 0.001 * | 0.037 * | 0.009 * | 0.010 * |
| PN (Without Lymph Node Metastasis vs. Lymph Lode Metastasis) | |||||
| HR | 1.851 | 2.355 | 1.555 | 2.609 | 2.547 |
| 95%CI | 1.065–3.218 | 1.162–4.774 | 0.756–3.200 | 1.032–6.596 | 0.988–6.562 |
| p | 0.029 * | 0.017 * | 0.231 | 0.043 * | 0.053 |
| PM (Without Metastasis vs. Metastasis) | |||||
| HR | 1.710 | 3.303 | - | - | - |
| 95%CI | 0.531–5.508 | 0.999–10.924 | - | - | - |
| p | 0.369 | 0.050 | - | - | - |
| FIGO (I–IIa vs. IIb–IV) | |||||
| HR | 2.630 | 2.913 | 1.088 | 0.584 | 0.599 |
| 95%CI | 1.439–4.808 | 1.340–6.332 | 0.360–3.288 | 0.134–2.536 | 0.136–2.633 |
| p | 0.002 * | 0.007 * | 0.881 | 0.473 | 0.498 |
| Grade (I vs. II–III) | |||||
| HR | 1.672 | 1.054 | - | - | - |
| 95%CI | 0.520–5.375 | 0.320–3.472 | - | - | - |
| p | 0.388 | 0.931 | - | - | - |
| Disease Subtype (Squamous Carcinoma vs. Adenocarcinoma) | |||||
| HR | 1.901 | 2.882 | 1.918 | 2.824 | 2.830 |
| 95%CI | 1.043–3.466 | 1.408–5.899 | 1.038–3.545 | 1.359–5.869 | 1.361–5.883 |
| p | 0.036 * | 0.004 * | 0.038 * | 0.005 * | 0.005 * |
| Number of CCL22+ cells (CCL22low vs. CCL22high) | |||||
| HR | - | 3.41 | - | 4.985 | - |
| 95%CI | - | 1.567–7.419 | - | 2.206–11.266 | - |
| p | - | 0.002 * | - | 0.0001 * | - |
| Number of CCL22+FOXP3+ cells (CCL22lowFOXP3low vs. CCL22highFOXP3high) | |||||
| HR | 2.806 | 5.355 | 3.018 | - | 5.284 |
| 95%CI | 0.864–9.115 | 1.580–18.154 | 0.923–9.869 | - | 1.513–18.456 |
| p | 0.086 | 0.007 * | 0.068 | - | 0.009 * |
| Antibody | Isotype | Clone | Dilution | Source |
|---|---|---|---|---|
| CCL22 | rabbit IgG | polyclonal | 1:400 in PBS a 1:400 in Dako b | Perprotech; DAKO(S322); Carpentera, CA, USA |
| FoxP3 | mouse IgG | monoklonal | 1:300 in PBS a | Abcam |
| CD68 | mouse IgG | polyclonal | 1:1000 in PBS a 1:1000 in Dako b | Sigma; DAKO(S322); Carpentera, CA, USA |
| CD163 | mouse IgG | monoklonal | 1:800 in PBS a 1:800 in Dako b | Abcam; DAKO(S322); Carpentera, CA, USA |
| Cy-3 b | goat IgG anti-rabbit | polyclonal | 1:500 b in Dako b | Dianova, Hamburg, Germany |
| Cy-2 b | goat IgG anti-mouse | polyclonal | 1:100 b in Dako b | Dianova, Hamburg, Germany |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Schmoeckel, E.; Kost, B.P.; Kuhn, C.; Vattai, A.; Vilsmaier, T.; Mahner, S.; Mayr, D.; Jeschke, U.; Heidegger, H.H. Higher CCL22+ Cell Infiltration is Associated with Poor Prognosis in Cervical Cancer Patients. Cancers 2019, 11, 2004. https://doi.org/10.3390/cancers11122004
Wang Q, Schmoeckel E, Kost BP, Kuhn C, Vattai A, Vilsmaier T, Mahner S, Mayr D, Jeschke U, Heidegger HH. Higher CCL22+ Cell Infiltration is Associated with Poor Prognosis in Cervical Cancer Patients. Cancers. 2019; 11(12):2004. https://doi.org/10.3390/cancers11122004
Chicago/Turabian StyleWang, Qun, Elisa Schmoeckel, Bernd P. Kost, Christina Kuhn, Aurelia Vattai, Theresa Vilsmaier, Sven Mahner, Doris Mayr, Udo Jeschke, and Helene Hildegard Heidegger. 2019. "Higher CCL22+ Cell Infiltration is Associated with Poor Prognosis in Cervical Cancer Patients" Cancers 11, no. 12: 2004. https://doi.org/10.3390/cancers11122004
APA StyleWang, Q., Schmoeckel, E., Kost, B. P., Kuhn, C., Vattai, A., Vilsmaier, T., Mahner, S., Mayr, D., Jeschke, U., & Heidegger, H. H. (2019). Higher CCL22+ Cell Infiltration is Associated with Poor Prognosis in Cervical Cancer Patients. Cancers, 11(12), 2004. https://doi.org/10.3390/cancers11122004