Integrative Metabolomic and Transcriptomic Analysis for the Study of Bladder Cancer

 , , ,
, , ,  ,
,  , , and
, , and
Abstract
1. Introduction
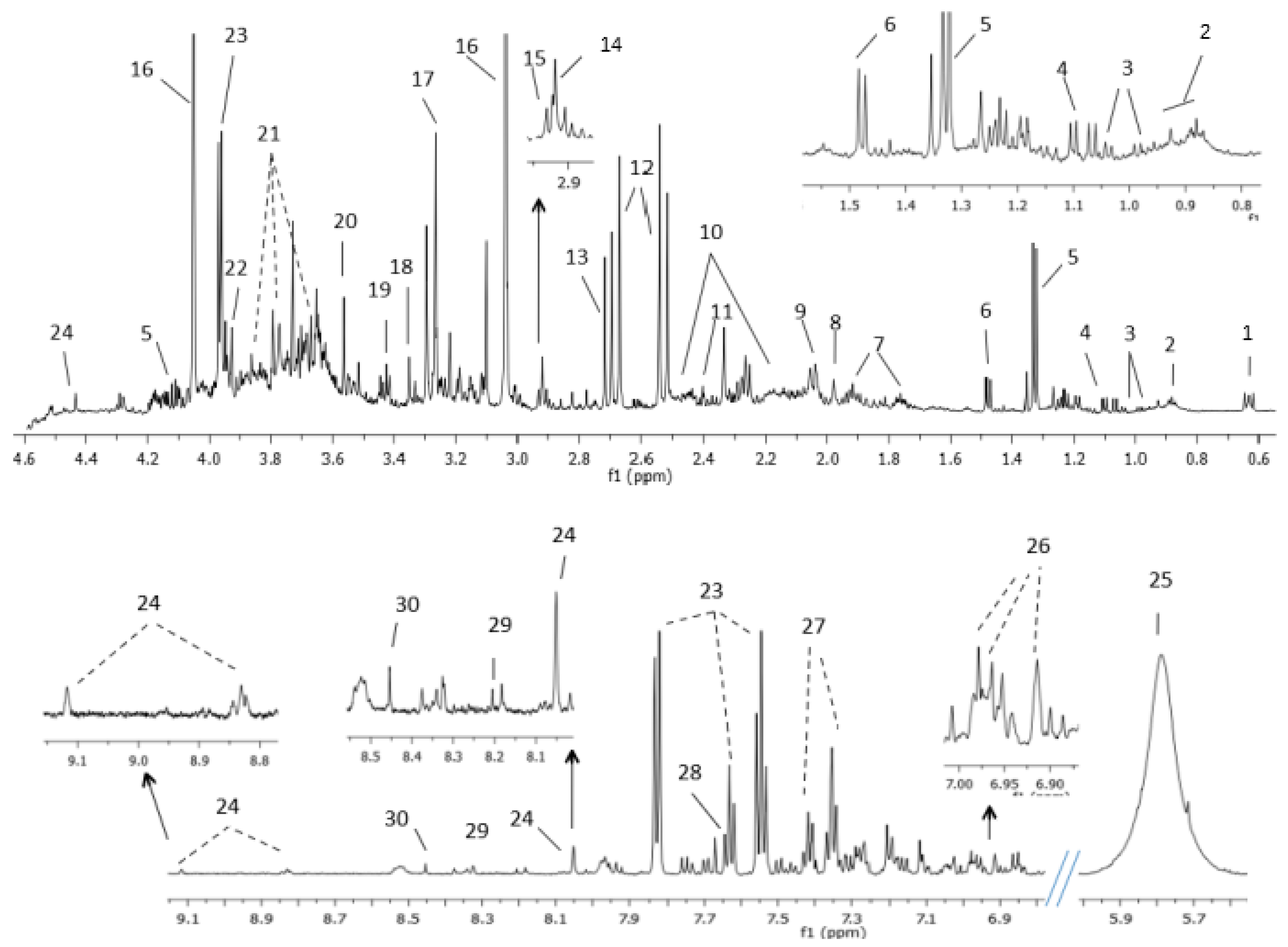
2. Results
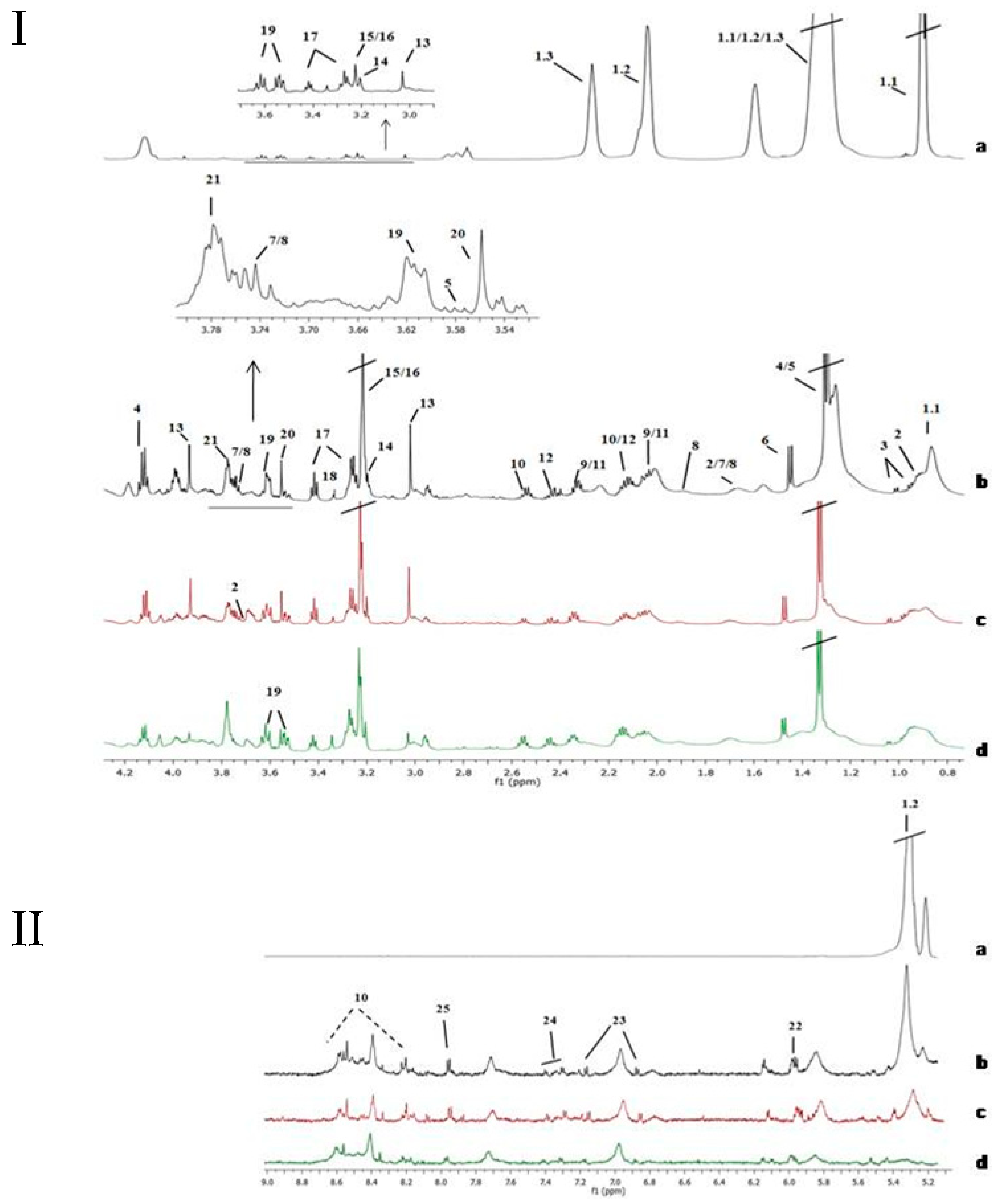
2.1. Tissue NMR Profile
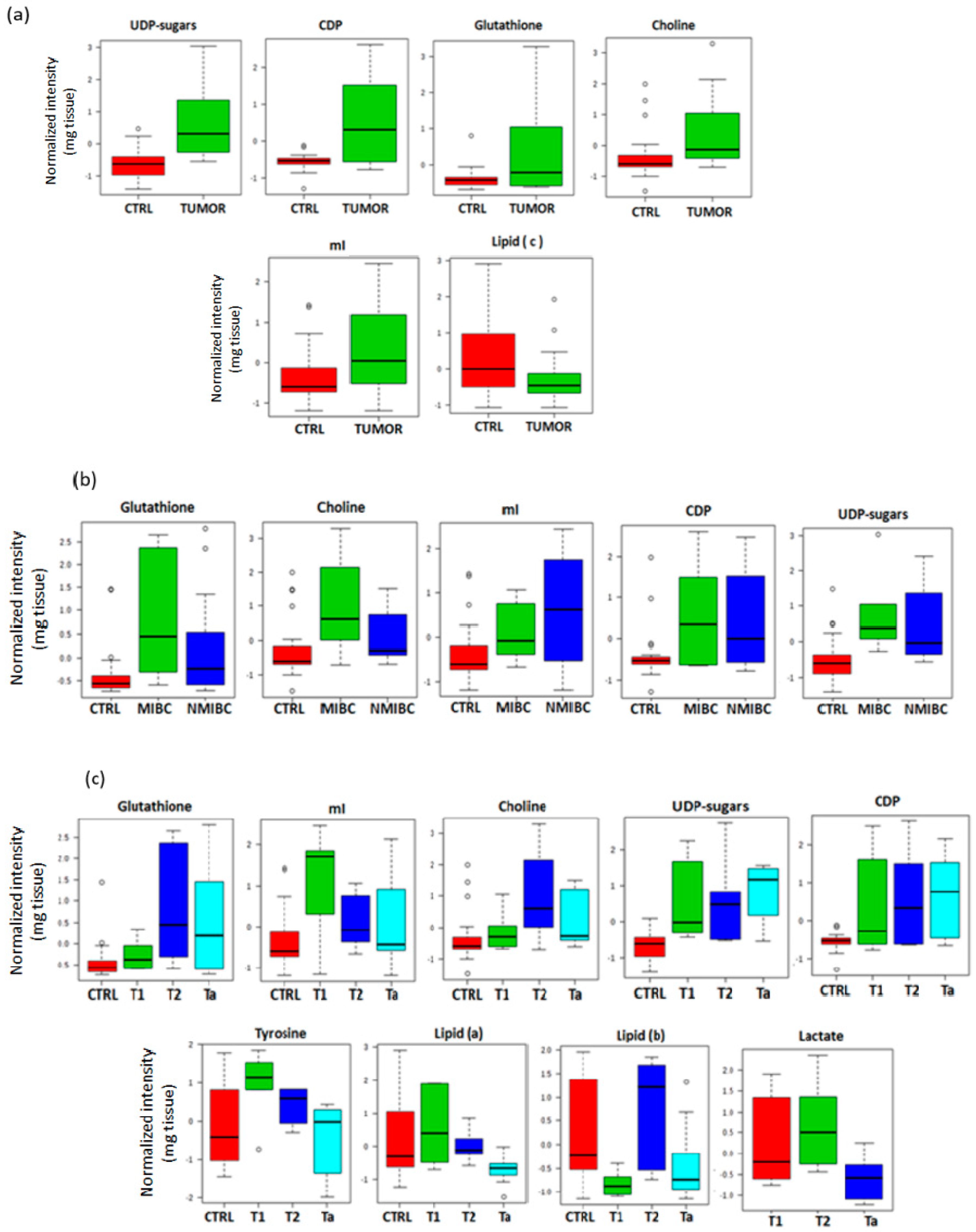
2.2. Mean Comparison of Metabolites Detected in Tissue
2.3. PLS-DA Analysis in NMR Tissue Data
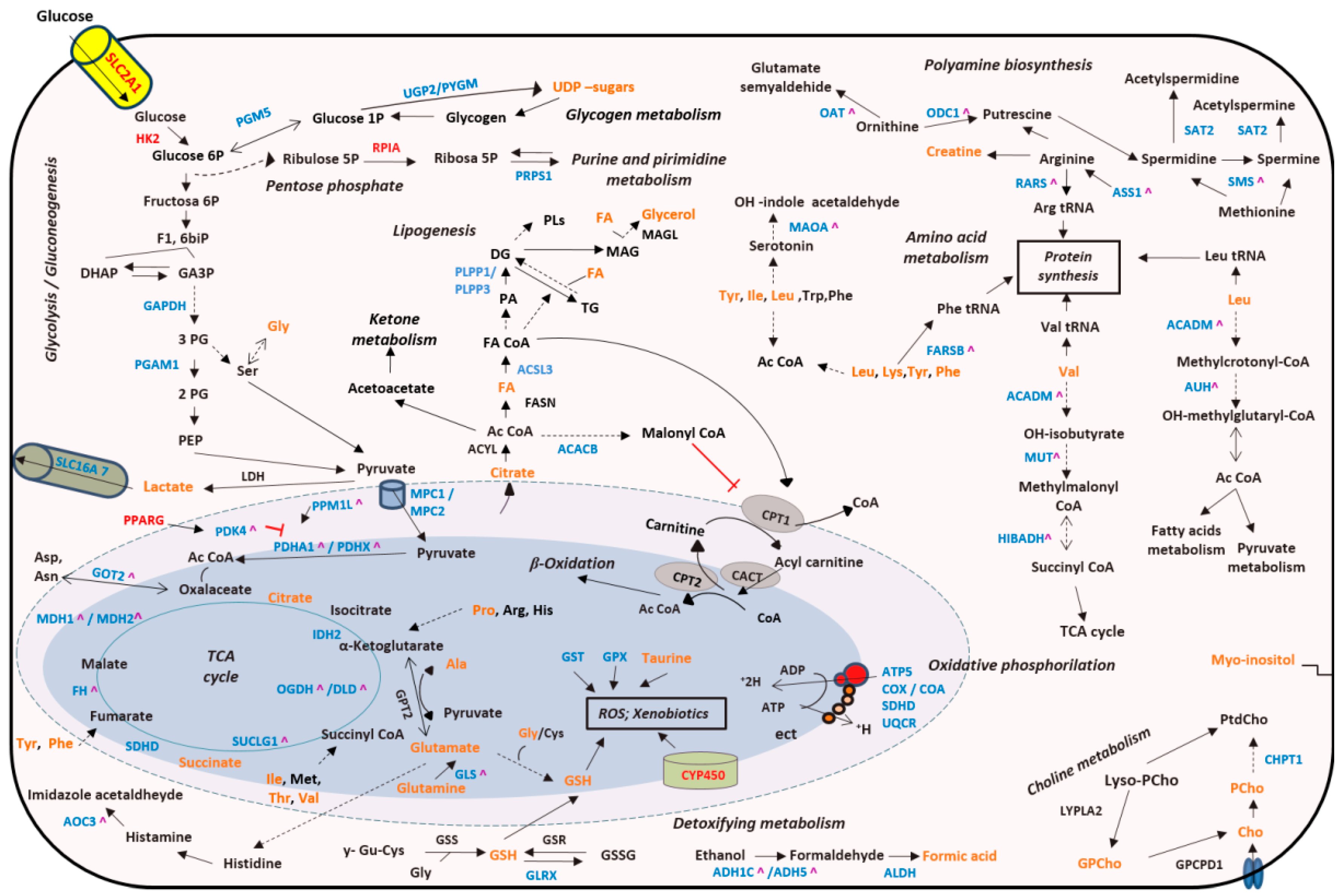
2.4. Transcriptomic Analysis in Tissue Samples
2.5. Metabolomic Analysis in Urine Samples
2.6. PLS-DA Analysis in NMR Urine Data
3. Discussion
4. Materials and Methods
4.1. Patient Selection and Sample Collection
4.2. Tissue NMR Experiments
4.3. Tissue Transcriptomic Experiments
4.4. Urine NMR Experiments
4.5. Tissue NMR Data Pre-Processing and Analysis
4.6. Tissue Transcriptomic Data Analysis
4.7. Urine NMR Data Pre-Processing and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leal, J.; Luengo-Fernandez, R.; Sullivan, R.; Witjes, J.A. Economic Burden of Bladder Cancer Across the European Union. Eur. Urol. 2016, 69, 438–447. [Google Scholar] [CrossRef]
- Burger, M.; Catto, J.W.F.; Dalbagni, G.; Grossman, H.B.; Herr, H.; Karakiewicz, P.; Kassouf, W.; Kiemeney, L.A.; La Vecchia, C.; Shariat, S.; et al. Epidemiology and Risk Factors of Urothelial Bladder Cancer. Eur. Urol. 2013, 63, 234–241. [Google Scholar] [CrossRef]
- Park, J.C.; Citrin, D.E.; Agarwal, P.K.; Apolo, A.B. Multimodal management of muscle-invasive bladder cancer. Curr. Probl. Cancer 2014, 38, 80–108. [Google Scholar] [CrossRef] [PubMed]
- Ghatalia, P.; Zibelman, M.; Geynisman, D.M.; Plimack, E. Approved checkpoint inhibitors in bladder cancer: which drug should be used when? Ther. Adv. Med. Oncol. 2018, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Jiang, H.; Tao, T.; Kang, X.; Zhang, X.; Kang, D.; Li, S.; Li, C.; Wang, H.; Yang, Z.; et al. Hope and challenge: Precision medicine in bladder cancer. Cancer Med. 2019, 8, 1806–1816. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.H.; Chang, S.S. Management of carcinoma in situ of the bladder: best practice and recent developments. Ther. Adv. Urol. 2015, 7, 351–364. [Google Scholar] [CrossRef]
- Yang, B.; Liao, G.-Q.; Wen, X.-F.; Chen, W.-H.; Cheng, S.; Stolzenburg, J.-U.; Ganzer, R.; Neuhaus, J. Nuclear magnetic resonance spectroscopy as a new approach for improvement of early diagnosis and risk stratification of prostate cancer. J. Zhejiang Univ. Sci. B 2017, 18, 921–933. [Google Scholar] [CrossRef]
- Tilki, D.; Burger, M.; Dalbagni, G.; Grossman, H.B.; Hakenberg, O.W.; Palou, J.; Reich, O.; Rouprêt, M.; Shariat, S.F.; Zlotta, A.R. Urine Markers for Detection and Surveillance of Non–Muscle-Invasive Bladder Cancer. Eur. Urol. 2011, 60, 484–492. [Google Scholar] [CrossRef]
- Robertson, A.G.; Kim, J.; Al-Ahmadie, H.; Bellmunt, J.; Guo, G.; Cherniack, A.D.; Hinoue, T.; Laird, P.W.; Hoadley, K.A.; Akbani, R.; et al. Comprehensive Molecular Characterization of Muscle-Invasive Bladder Cancer. Cell 2017, 171, 540–556. [Google Scholar] [PubMed]
- Network, T.C.G.A.R. Comprehensive molecular characterization of urothelial bladder carcinoma. Nature 2014, 507, 315–322. [Google Scholar]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Martín-Martín, N.; Carracedo, A.; Torrano, V. Metabolism and Transcription in Cancer: Merging Two Classic Tales. Front. Cell Dev. Biol. 2017, 5, 119. [Google Scholar] [CrossRef] [PubMed]
- Hirschey, M.D.; DeBerardinis, R.J.; Diehl, A.M.E.; Drew, J.E.; Frezza, C.; Green, M.F.; Jones, L.W.; Ko, Y.H.; Le, A.; Lea, M.A.; et al. Dysregulated metabolism contributes to oncogenesis. Semin. Cancer Biol. 2015, 35 Suppl, 129–150. [Google Scholar] [CrossRef]
- Loras, A.; Trassierra, M.; Sanjuan-Herráez, D.; Martínez-Bisbal, M.C.; Castell, J.V.; Quintás, G.; Ruiz-Cerdá, J.L. Bladder cancer recurrence surveillance by urine metabolomics analysis. Sci. Rep. 2018, 8, 9172. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Yang, X.; Deng, X.; Zhang, X.; Li, P.; Tao, J.; Qin, C.; Wei, J.; Lu, Q. Metabolomics in bladder cancer: a systematic review. Int. J. Clin. Exp. Med. 2015, 8, 11052–11063. [Google Scholar] [PubMed]
- Piyarathna, D.W.B.; Rajendiran, T.M.; Putluri, V.; Vantaku, V.; Soni, T.; von Rundstedt, F.-C.; Donepudi, S.R.; Jin, F.; Maity, S.; Ambati, C.R.; et al. Distinct Lipidomic Landscapes Associated with Clinical Stages of Urothelial Cancer of the Bladder. Eur. Urol. Focus 2017, 17, 30107–30114. [Google Scholar] [CrossRef]
- Martínez-Bisbal, M.C.; Martí-Bonmatí, L.; Piquer, J.; Revert, A.; Ferrer, P.; Llácer, J.L.; Piotto, M.; Assemat, O.; Celda, B. 1H and 13C HR-MAS spectroscopy of intact biopsy samples ex vivo and in vivo 1H MRS study of human high grade gliomas. NMR Biomed. 2004, 17, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, 90–97. [Google Scholar] [CrossRef]
- Yu, M.; Yongzhi, H.; Chen, S.; Luo, X.; Lin, Y.; Zhou, Y.; Jin, H.; Hou, B.; Deng, Y.; Tu, L.; et al. The prognostic value of GLUT1 in cancers: a systematic review and meta-analysis. Oncotarget 2017, 8, 43356–43367. [Google Scholar] [CrossRef]
- Hu, J.; Locasale, J.W.; Bielas, J.H.; O’Sullivan, J.; Sheahan, K.; Cantley, L.C.; Heiden, M.G.; Vander; Vitkup, D. Heterogeneity of tumor-induced gene expression changes in the human metabolic network. Nat. Biotechnol. 2013, 31, 522–529. [Google Scholar] [CrossRef]
- Saccenti., E.; Hoefsloot, H.C.J.; Smilde, A.K.; Westerhuis, J.A.; Hendriks, M.M.W.B. Reflections on univariate and multivariate analysis of metabolomics data. Metabolomics 2014, 10, 361–374. [Google Scholar] [CrossRef]
- Sajnani, K.; Islam, F.; Smith, R.A.; Gopalan, V.; Lam, A.K.-Y. Genetic alterations in Krebs cycle and its impact on cancer pathogenesis. Biochimie 2017, 135, 164–172. [Google Scholar] [CrossRef]
- Pavlova, N.N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef]
- Lew, C.R.; Guin, S.; Theodorescu, D. Targeting glycogen metabolism in bladder cancer. Nat. Rev. Urol. 2015, 12, 383–391. [Google Scholar] [CrossRef]
- Patra, K.C.; Wang, Q.; Bhaskar, P.T.; Miller, L.; Wang, Z.; Wheaton, W.; Chandel, N.; Laakso, M.; Muller, W.J.; Allen, E.L.; et al. Hexokinase 2 Is Required for Tumor Initiation and Maintenance and Its Systemic Deletion Is Therapeutic in Mouse Models of Cancer. Cancer Cell 2013, 24, 213–228. [Google Scholar] [CrossRef]
- Conde, V.R.; Oliveira, P.F.; Nunes, A.R.; Rocha, C.S.; Ramalhosa, E.; Pereira, J.A.; Alves, M.G.; Silva, B.M. The progression from a lower to a higher invasive stage of bladder cancer is associated with severe alterations in glucose and pyruvate metabolism. Exp. Cell Res. 2015, 335, 91–98. [Google Scholar] [CrossRef]
- Inamoto, T.; Shah, J.B.; Kamat, A.M. Friend or foe? Role of peroxisome proliferator-activated receptor-γ in human bladder cancer. Urol. Oncol. Semin. Orig. Investig. 2009, 27, 585–591. [Google Scholar] [CrossRef]
- Annicotte, J.-S.; Culine, S.; Fajas, L. Rôle de PPARγ dans la croissance du cancer de la prostate: de nouvelles pistes thérapeutiques. Bull. Cancer 2007, 94, 135–137. [Google Scholar]
- Tripathi, P.; Somashekar, B.S.; Ponnusamy, M.; Gursky, A.; Dailey, S.; Kunju, P.; Lee, C.T.; Chinnaiyan, A.M.; Rajendiran, T.M.; Ramamoorthy, A. HR-MAS NMR Tissue Metabolomic Signatures Cross-Validated by Mass Spectrometry Distinguish Bladder Cancer from Benign Disease NIH Public Access. J. Proteome Res. 2013, 5, 3519–3528. [Google Scholar] [CrossRef]
- Glunde, K.; Bhujwalla, Z.M.; Ronen, S.M. Choline metabolism in malignant transformation. Nat. Rev. Cancer 2011, 11, 835–848. [Google Scholar] [CrossRef]
- Gibellini, F.; Smith, T.K. The Kennedy pathway-De novo synthesis of phosphatidylethanolamine and phosphatidylcholine. IUBMB Life 2010, 62, 414–422. [Google Scholar] [CrossRef]
- Nowotarski, S.L.; Woster, P.M.; Casero, R.A., Jr. Polyamines and cancer: implications for chemotherapy and chemoprevention. Expert Rev. Mol. Med. 2013, 15, e3. [Google Scholar] [CrossRef]
- Thomas, T.; Thomas, T.J. Polyamine metabolism and cancer. J. Cell. Mol. Med. 2003, 7, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Cairns, R.A.; Harris, I.S.; Mak, T.W. Regulation of cancer cell metabolism. Nat. Publ. Gr. 2011, 11, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Pendyala, L.; Velagapudi, S.; Toth, K.; Zdanowicz, J.; Glaves, D.; Slocum, H.; Perez, R.; Huben, R.; Creaven, P.J.; Raghavan, D. Translational studies of glutathione in bladder cancer cell lines and human specimens. Clin. Cancer Res. 1997, 3, 793–798. [Google Scholar] [PubMed]
- Singer, K.; Cheng, W.-C.; Kreutz, M.; Ho, P.-C.; Siska, P.J. Immunometabolism in cancer at a glance. Dis. Model. Mech. 2018, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, P. Role of glutathione in immunity and inflammation in the lung. Int. J. Gen. Med. 2011, 4, 105–113. [Google Scholar] [CrossRef]
- Woolbright, B.L.; Ayres, M.; Taylor, J.A. Metabolic changes in bladder cancer. Urol. Oncol. Semin. Orig. Investig. 2018, 36, 327–337. [Google Scholar] [CrossRef]
- Rosmarin, A.G.; Resendes, K.K.; Yang, Z.; McMillan, J.N.; Fleming, S.L. GA-binding protein transcription factor: a review of GABP as an integrator of intracellular signaling and protein–protein interactions. Blood Cells, Mol. Dis. 2004, 32, 143–154. [Google Scholar] [CrossRef]
- Lee, J.-S.; Leem, S.-H.; Lee, S.-Y.; Kim, S.-C.; Park, E.-S.; Kim, S.-B.; Kim, S.-K.; Kim, Y.-J.; Kim, W.-J.; Chu, I.-S. Expression signature of E2F1 and its associated genes predict superficial to invasive progression of bladder tumors. J. Clin. Oncol. 2010, 28, 2660–2667. [Google Scholar] [CrossRef]
- Kozlovski, I.; Siegfried, Z.; Amar-Schwartz, A.; Karni, R. The role of RNA alternative splicing in regulating cancer metabolism. Hum. Genet. 2017, 136, 1113–1127. [Google Scholar] [CrossRef]
- Snezhkina, A.V.; Krasnov, G.S.; Zaretsky, A.R.; Zhavoronkov, A.; Nyushko, K.M.; Moskalev, A.A.; Karpova, I.Y.; Afremova, A.I.; Lipatova, A.V.; Kochetkov, D.V.; et al. Differential expression of alternatively spliced transcripts related to energy metabolism in colorectal cancer. BMC Genomics 2016, 17, 1011. [Google Scholar] [CrossRef] [PubMed]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P.; et al. The Human Urine Metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef]
- Cao, M.; Zhao, L.; Chen, H.; Xue, W.; Lin, D. NMR-based metabolomic analysis of human bladder cancer. Anal. Sci. 2012, 28, 451–456. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: the human metabolome database for 2018. Nucleic Acids Res. 2018, 46, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, P.; Vogel, T.; Malusek, A.; Lundquist, P.-O.; Cohen, L.; Dahlqvist, O. MDL—The Magnetic Resonance Metabolomics Database.
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0--making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, 251–257. [Google Scholar] [CrossRef]
- Savorani, F.; Tomasi, G.; Engelsen, S.B. icoshift: A versatile tool for the rapid alignment of 1D NMR spectra. J. Magn. Reson. 2010, 202, 190–202. [Google Scholar] [CrossRef]
- Liu, J.; Lichtenberg, T.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An Integrated TCGA Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell 2018, 173, 400–416. [Google Scholar] [CrossRef]
- Howe, E.A.; Sinha, R.; Schlauch, D.; Quackenbush, J. RNA-Seq analysis in MeV. Bioinformatics 2011, 27, 3209–3210. [Google Scholar] [CrossRef]
- Frolkis, A.; Knox, C.; Lim, E.; Jewison, T.; Law, V.; Hau, D.D.; Liu, P.; Gautam, B.; Ly, S.; Guo, A.C.; et al. SMPDB: The Small Molecule Pathway Database. Nucleic Acids Res. 2010, 38, 480–487. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
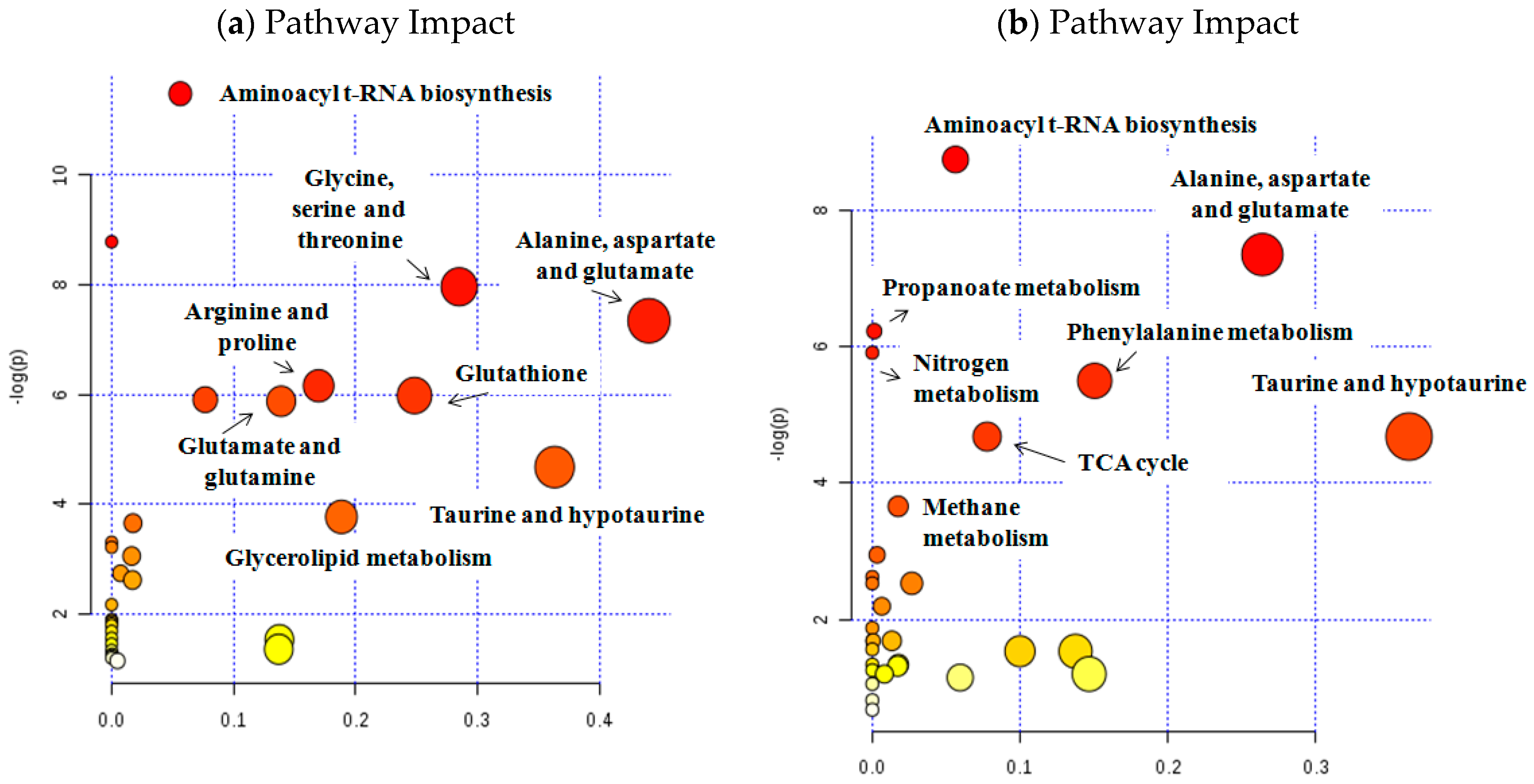
| Metabolic altered Pathways | Sample | Metabolites | p | Impact |
|---|---|---|---|---|
| Alanine, aspartate, glutamate | Tissue | Alanine, glutamine, glutamate | 6.5E-4 | 0.44 |
| Urine | Alanine, glutamine, succinate | 6.5E-4 | 0.26 | |
| Taurine and hypotaurine | Tissue | Taurine, alanine | 9.0E-3 | 0.36 |
| Urine | Taurine, alanine | 9.0E-3 | 0.36 | |
| Aminoacyl-tRNA biosynthesis | Tissue | Proline, glycine, alanine, glutamine, threonine, glutamate | 1.0E-5 | 0.06 |
| Urine | Phenylalanine, glutamine, valine, alanine, lysine | 1.6E-4 | 0.56 | |
| Methane | Tissue | Glycine, methanol | 2.6E-2 | 0.02 |
| Urine | Trimethylamine N-oxide, methanol | 2.5E-2 | 0.02 | |
| Glycine, serine, threonine | Tissue | Glycine, creatine, choline, threonine | 3.5E-4 | 0.29 |
| Glutathione | Tissue | Glutathione, glycine, glutamate | 2.0E-3 | 0.25 |
| Glycerolipid | Tissue | Fatty acid, glycerol | 2.3E-2 | 0.19 |
| Arginine and proline | Tissue | Glutamine, glutamate, proline, creatine | 2.0E-3 | 0.17 |
| Glutamine and glutamate | Tissue | Glutamine, glutamate | 3.0E-3 | 0.14 |
| Glycerophospholipid | Tissue | Choline, glycerophosphocholine, phosphocholine | 3.0E-3 | 0.08 |
| Citrate cycle | Urine | Citrate, succinate | 9.0E-3 | 0.08 |
| Phenylalanine | Urine | Succinate, phenylalanine, hippuric acid | 4.0E-3 | 0.07 |
| Nitrogen metabolism | Urine | Phenylalanine, taurine, glutamine | 2.7E-3 | 0.05 |
| Propanoate | Urine | Succinate, lactic acid, valine | 2.0E-3 | 0.05 |
| Tissue Samples | Urine Samples | ||
|---|---|---|---|
| PLS-DA Models (BC vs Control) | Calibration (CV) | Validation | Calibration (CV) |
| Patients (male/female) | 17(10/7) | 5 (4/1) | 21 (14/7) |
| Age (mean and standard deviation) | 71 (9) | 63 (11) | 69 (10) |
| Samples (male/female) | 34 (20/14 | 10 (8/2) | 35 (23/11) |
| Tumor samples (BC) | 17 | 5 | 22 |
| Non-tumor samples (Control) | 17 | 5 | 13 |
| Primary/Recurrent BC | 15/2 | 4/1 | 19/3 |
| Tumor stage (Ta, T1, T2) | 7/6/4 | 3/0/2 | 10/6/6 |
| Tumor grade (1/2/3) | 4/3/10 | 0/2/3 | 18/4 |
| Recurrence risk group 5 yr (EORTC)a: L/ L-I/ H-I/ H | 2/4/6/1 | 0/0/3/0 | 2/4/9/1 |
| Progression risk group 5 yr (EORTC) a: L/L-I/H-I/H | 4/2/5/2 | 0/1/2/0 | 4/3/7/2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loras, A.; Suárez-Cabrera, C.; Martínez-Bisbal, M.C.; Quintás, G.; Paramio, J.M.; Martínez-Máñez, R.; Gil, S.; Ruiz-Cerdá, J.L. Integrative Metabolomic and Transcriptomic Analysis for the Study of Bladder Cancer. Cancers 2019, 11, 686. https://doi.org/10.3390/cancers11050686
Loras A, Suárez-Cabrera C, Martínez-Bisbal MC, Quintás G, Paramio JM, Martínez-Máñez R, Gil S, Ruiz-Cerdá JL. Integrative Metabolomic and Transcriptomic Analysis for the Study of Bladder Cancer. Cancers. 2019; 11(5):686. https://doi.org/10.3390/cancers11050686
Chicago/Turabian StyleLoras, Alba, Cristian Suárez-Cabrera, M. Carmen Martínez-Bisbal, Guillermo Quintás, Jesús M. Paramio, Ramón Martínez-Máñez, Salvador Gil, and José Luis Ruiz-Cerdá. 2019. "Integrative Metabolomic and Transcriptomic Analysis for the Study of Bladder Cancer" Cancers 11, no. 5: 686. https://doi.org/10.3390/cancers11050686
APA StyleLoras, A., Suárez-Cabrera, C., Martínez-Bisbal, M. C., Quintás, G., Paramio, J. M., Martínez-Máñez, R., Gil, S., & Ruiz-Cerdá, J. L. (2019). Integrative Metabolomic and Transcriptomic Analysis for the Study of Bladder Cancer. Cancers, 11(5), 686. https://doi.org/10.3390/cancers11050686