Simple Summary
Transforming growth factor beta (TGF-β) is a multifunctional cytokine that can restrict cancer onset but also promote cancer progression at late stages of cancer. The ability of TGF-β in producing diverse and sometimes opposing effects relies on its potential to control different cellular signalling and gene expression in distinct cell types, and environmental settings. The tumour promoting role of TGF-β is primarily mediated through its effects on the local tumour microenvironment (TME) of the cancer cells. In this review, we discuss the most recent research on the role and regulation of TGF-β, with a specific focus on its functions on promoting cancer progression through targeting different immune cells in the TME as well as its therapeutic perspectives.
Abstract
Transforming growth factor-β (TGF-β) was originally identified as an anti-tumour cytokine. However, there is increasing evidence that it has important roles in the tumour microenvironment (TME) in facilitating cancer progression. TGF-β actively shapes the TME via modulating the host immunity. These actions are highly cell-type specific and complicated, involving both canonical and non-canonical pathways. In this review, we systemically update how TGF-β signalling acts as a checkpoint regulator for cancer immunomodulation. A better appreciation of the underlying pathogenic mechanisms at the molecular level can lead to the discovery of novel and more effective therapeutic strategies for cancer.
1. Introduction
Transforming growth factor-beta (TGF-β) is a secretory cytokine that has pleiotropic roles in cancer progression through controlling cell proliferation, differentiation, apoptosis, and migration. The TGF-β family of cytokines consists of three different isoforms, TGF-β1, TGF-β2, and TGF-β3, with each of them having a unique expression mode and executing distinct functions. For example, TGF-β2 can deplete interleukin 6 (IL-6) function and induce apoptosis [1], TGF-β3 affects the differentiation of mesenchymal stromal cells (MSCs) [2], while TGF-β1 plays an important role in cancer progression and tumour microenvironment (TME) development [3,4].
TGF-β executes its functions through canonical and non-canonical pathways (Figure 1). In the canonical TGF-β/Smad pathway, TGF-β ligands bind to a heterotetrameric TGF-β receptor complex, composed of dimers of type I (TGFBR1) and type II (TGFBR2) TGF-β receptors. The ligated TGF-β and receptor complexes then become phosphorylated and activated before recruiting and phosphorylating Smads, including Smad2 and Smad3. Smad2/3 phosphorylation induces the subsequent recruitment of Smad4 to the Smad2/3 complex which then translocates to the nucleus. Smad3 is the transcription factor (TF) and the component of the complex that directly binds to DNA, and the Smad complexes then cooperate with other cofactors to regulate multiple downstream mechanisms through directing transcriptional activation and suppression [5]. For example, TGF-β mediates the transcriptional regulation of effector genes including the cyclin-dependent kinase (CDK) inhibitors p15INK4 [6] and p21Cip1 [7] to modulate cell cycle arrest. Besides, the TGF-β/Smad axis also promotes the stemness and epithelial–mesenchymal transition (EMT) of cancer cells by restoring mesenchymal phenotypes and upregulating the expression of genes, such as Snail and Vim [8,9]. Meanwhile, the TGF-β/Smad pathway also has a negative feedback mechanism mediated through Smad7 competitive binding to TGFBR1 and blocking the TGF-β/Smad pathway signalling [10]. For the non-canonical TGF-β pathway, the activated TGF-β crosstalks with other signalling pathways, such as Rho, phosphoinositide 3-kinase (PI3K), and mitogen-activated protein kinase (MAPK) signalling cascades, to promote EMT [11], cancer invasion [12], and angiogenesis [13]. In consequence, both the canonical and non-canonical TGF-β pathways play an important role in cancer progression [14].
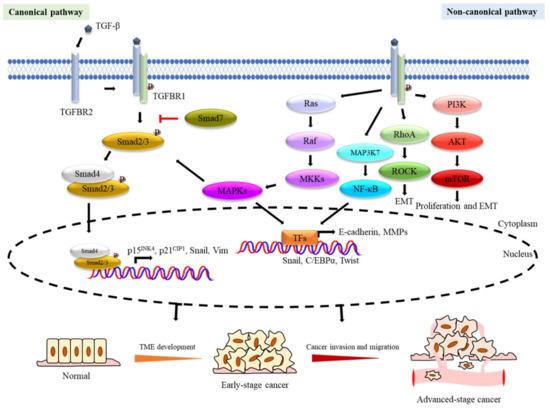
Figure 1.
Transforming growth factor-β (TGF-β) signalling pathways in tumorigenesis. The dual roles of TGF-β signalling pathways have been demonstrated in tumorigenesis. TGF-β is a tumour suppressor in TME development of early-stage cancer and a tumour promoter in malignancy processes of advanced-stage cancer. Schematic diagramme (above) showing TGF-β signalling and its role in cancer tumorigenesis and progression as well as tumour suppression. TGF-β binds to TGFBR2 which then complexes with TGFBR1 to activate downstream signalling. TGF-β can activate both Smad-dependent canonical and Smad-independent non-canonical signalling cascades. The TGF-β activated TGFBR1 phosphorylates the Smad2/3 complex which then associates Smad4, before translocating to the nucleus to regulate the transcription of different targeted genes involved in tumour suppression during tumorigenesis (e.g., p15INK4b and p21Cip1), as well as tumour progression (e.g., Snail and Vimentin). The Smad-dependent TGF-β canonical signalling pathway can also be antagonised by Smad7 through inhibiting the binding of the Smad2/3 complex with Smad4. In the Smad-independent non-canonical signalling cascades, the activated TGFBR1/2 receptors induce downstream signalling through the Ras/Raf/MAPKs (JNK/p38/ERK), PI3K/AKT, MAP3K7(TAK1)/NF-κB, and Rho family of small GTPase-dependent signalling pathways. The TGF-β activated JNK/p38/ERK-MAPKs also crosstalk with Smad2/3/4 to modulate downstream signalling to influence cancer development. The Rho family of small GTPase-dependent signalling pathways (e.g., Rho/ROCK) are involved in epithelial–mesenchymal transition (EMT). The activated PI3K/AKT pathway induces mTOR to promote cell proliferation and EMT. The TGF-β signals also activate NF-κB signalling to modulate inflammatory response.
Accumulating evidence has indicated that TGF-β has a dual function in cancer progression, with the different TGF-β signalling pathways switching between the two phenotypes of tumour suppression and tumour promotion [15]. More importantly, the role of TGF-β in the immunosuppressive TME and immune cells has attracted increasing attention. In this review, we will evaluate the complex role of TGF-β in cancer progression, focusing primarily on its function in the regulation of immune responses and TME development.
2. TGF-β in Cancer Initiation and Progression
2.1. Role of TGF-β as Tumour Suppressor
TGF-β plays multiple roles in the early stages of carcinogenesis, one of which is to modulate cell proliferation arrest. For example, TGF-β can induce cell cycle arrest via the cAMP response element-binding protein (CREB) to mediate histone acetylation and transcriptional activation of plasminogen activator inhibitor type-1 in a p53/Smad-dependent manner to maintain the anti-proliferative effects to suppress cancer tumorigenesis [16]. Besides, TGF-β can also activate the forkhead box O1 (FOXO1) transcription factor to induce the expression of the cyclin-dependent kinase inhibitor p21Cip1 at the transcriptional level to enforce a G0/G1 phase cell cycle arrest. This is mediated via the TGF-β/FOXO1/p21Cip1 axis to keep cells in a quiescent or senescent state [17]. TGF-β can also promote normal cell differentiation as an indirect tumour suppressive antiproliferative mechanism by promoting the activity of the stress-activated p38MAPK signalling pathway [18]. In agreement, haematopoietic stem and progenitor cells (HSPCs) expressing high levels of active TGF-β1 protein and p38MAPK activity lose their haematopoietic stem cell (HSC) self-renewal and multi-lineage capacity and differentiate into progenitor cells [18].
Mechanistically, the role of TGF-β as a tumour suppressor during early tumorigenesis is mediated primarily through its function to cause cell cycle and proliferative arrest, to induce differentiation and apoptosis, and to block paracrine factor production [17]. The anti-proliferative tumour suppressive function of TGF-β is mediated primarily through the induction of CDK inhibitor expression and the suppression of c-Myc expression, at the transcriptional level, to arrest cell cycle progression at the G1 and S phases of the cell cycle. For example, in normal epithelial cells, TGF-β induces the expression of the cyclin-dependent kinase inhibitor (CKI) p15INK4b, which competes with cyclin Ds for the formation of cyclin D complexes with CDK4/6, and of p21Cip1, which inhibits the activity of the cyclin E/A-CDK2 complexes directly. Specifically, the Smad3/4 complexes associate and cooperate with FOXO transcription factors to transcriptionally activate the promoters of the CDKN2B gene, which encodes p15INK4b, and of CDKN1A, which encodes p21Cip1. The induced CKI p15INK4b can also displace p27Kip1 from the cyclin D-CDK4/6 complexes, releasing the Cip/Kip CKI (e.g., p21Cip1, p27Kip1, and p57Kip2) to inhibit the cyclin E/A-CDK2 complexes to restrict cell cycle progression through late G1 and S phases of the cell cycle. Indeed, TGF-β has also been shown to stimulate the expression of CKIs, such as p15INK4b, p21Cip1, p27Kip1, and p57Kip2, in a cell type- and context-dependent manner. For example, TGF-β induces the expression of p21CIP1 in T cells, p57Kip2 in haematopoietic stem/progenitor cells, and p15INK4b and p21Cip1 in astrocytes, neural progenitor cells, and epithelial cells to cause cell cycle and proliferation arrest. The TGF-β-induced Smad3/4-containing protein complexes also repress the expression of transcription of c-Myc, a potent oncogene which plays a pivotal role in cell cycle entry and proliferation. Besides cell cycle progression and proliferation, TGF-β also limits proliferation indirectly through promoting differentiation. For instance, TGF-β signalling also represses the expression of Id proteins (inhibitor of differentiation/DNA binding), which inhibit some key differentiation pathways. In addition, TGF-β signalling can also trigger apoptosis, programmed cell death to restrict cell proliferation. Consistently, TGF-β can promote apoptosis through inducing the expression of modulators of programmed cell death, including the death-associated protein kinase (DAPK), the growth arrest and DNA damage 45β factor (GADD45β), the death receptor FAS, etc., and often in a cell type-dependent manner. Lastly, TGF-β can also restrict epithelial cell proliferation and tumorigenesis by inhibiting the production of mitotic growth factors, such as hepatocyte growth factor (HGF), by the fibroblasts and immune cells in the local TME. Together, these findings suggest that TGF-β targets different gene targets via discrete signalling pathways in distinct cell types and settings to function as a pleiotropic suppressor of cancer onset [17].
2.2. Role of TGF-β as Tumour Progression Promoter
Conversely, TGF-β can also function as an oncogene and promote cancer cell progression by activating the PI3K/AKT/mTOR pathway [19]. Appropriately, TGF-β is associated with poor prognosis and cancer progression in osteosarcoma [19]. Alternatively, TGF-β1 has also been shown to activate Golgi membrane protein 1 (GOLM1; also called GP73) through lipid rafts to suppress Smad-dependent tumour-suppressive signals and, at the same time, induce the extracellular signal-regulated kinase (ERK) MAPK signalling pathway to promote tumorigenesis and progression in liver cancer [20]. In addition, TGF-β also regulates tissue infiltration and migration by enhancing bFGF signalling through the FGFR/FRS2/ERK axis in paediatric medulloblastoma [21]. The importance of MAPK in mediating the oncogenic function of TGF-β is exemplified by the fact that the Smad4 phosphatase Wip1 can restrain the TGF-β-induced cell growth arrest, migration, and invasion and enhances the tumorigenicity of cancer cells by repressing Smad4 activity via antagonising MAPK function. Mechanistically, Wip1 selectively dephosphorylates Smad4 at a specific MAPK phosphorylation site to inhibit its nuclear accumulation and stability [22].
Increasing evidence has indicated that TGF-β can aggravate EMT, a key process involved in metastasis during cancer progression. For example, the TGF-β/Smad pathways can promote EMT and contribute to lung cancer progression by driving the expression of the DNA binding protein family member ID1 and EMT-related transcriptional factor Snail. Besides that, TGF-β can also induce EMT via the PI3K/AKT axis in a tuberous sclerosis protein complex-dependent manner [23]. Furthermore, TGF-β also promotes EMT and enhances cancer cell invasion and migration via activating Rho and Rho-associated protein kinases (ROCKs), whereas the malignant phenotypes induced by TGF-β via the Rho/ROCK signalling pathway can be blocked by miR-335-5p and farnesyl pyrophosphate synthase inhibitors in non-small cell lung cancer (NSCLC) [24,25]. The activation of MAPKs and downstream genes is another crucial mechanism by which TGF-β mediates cellular responses. TGF-β induces Ras and recruits Raf to the plasma membrane. The TGF-β-activated Ras and Raf then transmit signals via MAP kinase kinases (MKKs) to MAPKs to promote EMT. Cho et al. have found that the activated TGF-β signals induce the degradation of downstream Raf kinase inhibitory protein and promote the activation of the MAPK signalling pathway, which in turn results in the transcriptional suppression of p53 to facilitate EMT [26]. Moreover, the TGF-β pathway can also crosstalk with NF-κB to increase the expression of Twist to enhance EMT, and this cooperation between TGF-β and NF-κB pathways is again mediated by MAP kinase kinase kinase 7 (MAP3K7), an upstream MAPK family member [27,28]. Recently, CCAAT enhancer-binding protein alpha (C/EBPα) has been identified as a “gatekeeper” gene in TGF-β-induced EMT. More specifically, the TGF-β-induced downregulation of C/EBPα allows epithelial cells to undergo malignant transformation of breast epithelial cells, while abundant C/EBPα expression can effectively prevent breast cancer tumourigenesis [29].
Nevertheless, there is also evidence that TGF-β can also suppress tumour progression by generating a lethal EMT. Specifically, David et al. have found that TGF-β induces the expression of SRY-box transcription factor 4 (SOX4) to promote pro-apoptotic events via Smad4-mediated repression of kruppel-like factor 5 (KLF5). This in turn induces the lethal effects and phenotype transformation of EMT in TGF-β-sensitive pancreatic ductal adenocarcinoma (PDA) cells [30]. In agreement, the pro-apoptotic regulator WT1 is another gene that has been reported to be induced during TGF-β/Smad4-derived lethal EMT [31].
2.3. Role of TGF-β in Normal and Cancerous Cells
Overall, these findings suggest that TGF-β plays distinct and sometimes contrasting roles during cancer initiation and development. In general, in non-cancerous cells, TGF-β normally functions as a tumour suppressor to limit cell proliferation and promote apoptosis in order to exert growth inhibition during early stages of carcinogenesis. During later stages of cancer development, TGF-β often switches to a cancer promoting role, which is essential for tumour progression and metastasis. In concordance with this conjecture, one transcription factor that plays a key role during this TGF-β functional switch is distal-less homeobox 2 (Dlx2) [32]. In normal mammary epithelial cells, Dlx2 neutralises the TGFβ-induced cell cycle arrest and apoptosis by multiple mechanisms. Essentially, Dlx2 functions as a transcriptional repressor of TGFBR2 gene expression, restricting the downstream canonical Smad-dependent TGF-β signalling and expression of the cell cycle inhibitors, such as p21Cip1, and enhancing the expression of the potent mitogenic transcription factor c-Myc. Conversely, Dlx2 can also directly drive the expression of the epidermal growth factor (EGF) family member betacellulin to promote cell survival by stimulating EGF receptor (EGFR) signalling. Moreover, Dlx2 also further supports tumour growth and metastasis. These results establish Dlx2 as a critical switch in shifting TGF-β from its tumour-suppressive functions in early tumorigenesis to its tumour-promoting functions during cancer progression [32].
2.4. Regulation and Role of TGF-β in the Tumour Microenvironment
The TME provides the structures and materials for tumour maintenance and progression, and comprises the cellular and extracellular materials surrounding the tumour mass. Essentially, TME serves as a platform for cancer cells to reprogramme infiltrating stromal cells, thereby promoting tumorigenesis, as well as cancer invasion, metastasis, and resistance to therapy. TGF-β is secreted in a latent form as a large latent complex (LLC) which is primarily embedded in extracellular matrix (ECM) and is partially anchored on the cell surface [33,34]. It will remain as an inactivated complex until it is processed further to release the active TGF-β. Within the TME, a myriad of cell types can produce TGF-β, and the generation and processing of this important cytokine is mediated through diverse and complex mechanisms. For example, TGF-β produced by epithelial cells is induced via matrix metalloprotease (MMP) stimulation and EGFR activation [35]. In addition, TGF-β secretion in macrophages is regulated by the Notch signalling pathways [36]. Moreover, signal transducer and activator of transcription 6 (STAT6) and Furin can also cooperate to induce the production of TGF-β by T lymphocytes [37]. Furthermore, inactive forms of TGF-β can be stimulated and released by integrin-mediated activation, protease cleavage, and latency-associated peptide (LAP) dissociation upon X-ray irradiation [38,39,40]. It is also worth noting that a recent study has suggested a novel integrin-activated form of TGF-β that does not require release from the latent TGF-β complex [41]. Together, these observations illustrate the complexity and diversity of the mechanisms by which TGF-β are generated and activated. Moreover, diverse cell types are involved in modulating the effects of TGF-β on the development and the reprogramming of the TME, as well as during cancer progression. For example, migratory dendritic cells (DCs) can activate TGF-β via an integrin-dependent manner and promote naïve CD8+ T cell differentiation into epithelial resident memory T cells (eTRM) [42]. Notably, the interactions between non-immune cells and immune cells are also essential for TGF-β activation and function. It has been found that keratinocytes help to preserve epithelial-resident DCs and eTRM by inducing integrin expression and thereby TGF-β activation [43]. Besides that, tumour-initiating cells also release IL-33 to induce macrophage differentiation, which ultimately promotes TGF-β secretion and signalling to cancer stem cells (CSCs) to create a CSC niche for the maintenance of a stem cell pool within the TME [44]. In concordance, Takasaka et al. have also observed that integrin αvβ8 is highly expressed on the cancer cell surface and that the integrin αvβ8-expressing tumour cells can evade host immunity by regulating TGF-β activation in immune cells [45]. On the contrary, integrin β1-mediated TGF-β activation may also drive tumor suppression. In a human melanoma xenograft model, cell surface integrin β1-activation can increase TGF-β activity, which culminates in stromal activation and angiogenesis but also an accumulation of intra-tumoral CD8+ T cell infiltration within the TME. The latter recruitment of CD8+ T lymphocytes can result in an attenuation of tumour growth and long-term survival, suggesting a role of TGF-β in immune surveillance against tumours [46].
Besides targeting cancer cells, TGF-β also exerts its effect on stromal cells of the TME through controlling their differentiation, angiogenesis, and metabolic reprogramming during tumourigenesis and cancer progression. For example, amphiregulin expressed by macrophages can induce TGF-β activation to mediate pericyte differentiation into myofibroblasts to direct tissue restoration during inflammation [47]. In addition, TGF-β-licensed mesenchymal stromal cells (TGF-β MSCs) can contribute to immunosuppression by altering the phenotypes of macrophages and by promoting regulatory T cell (Treg) expansion [48]. Previous studies have also showed a strong association between stromal TGF-β levels and poor prognosis in hepatocellular carcinoma (HCC), as well as colorectal cancer (CRC), suggesting an important role for stromal TGF-β in promoting an immunosuppressive TME [49,50]. Moreover, TGF-β is able to stimulate myofibroblasts and other stromal cells to boost the synthesis of collagen crosslinked enzymes, especially lysyl oxidases (LOs) and matrix metallopeptidases (MMPs), to improve collagen crosslinking during the early stages of carcinogenesis [51]. Collagen crosslinking mediated by the lysyl oxidase family of enzymes (LOX, LOXL1-4) contributes to the pathogenesis of idiopathic pulmonary fibrosis (IPF), a progressive scarring lung disease which predisposes patients to lung cancer, mostly NSCLC. A previous study has also shown that the novel lysyl hydroxylase 3 receptor recruits MMP9 to the surface of fibroblasts and activated TGF-β and actin alpha 2 (α-SMA) functions during fibroblast differentiation [52]. At the same time, TGF-β can also downregulate the synthesis and expression of MMP family members, such as MMP2, MMP7, and MMP8, to facilitate the regulation of the ECM [53]. In addition, increased matrix protein synthesis and reduced matrix proteinase activity due to increased TGF-β activity can also contribute to the tumour ECM remodelling and result in desmoplasia, which is commonly found in many types of tumours, particularly pancreatic and renal cell carcinomas, as well as sarcomas [54]. In agreement, bone marrow-derived mesenchymal stem cells (BM-MSCs) have been shown to inhibit the TGF-β/SMAD pathway to repress hypoxia-inducible factor 1 subunit alpha (HIF-1α) and VEGF protein expression in the ECM to restrict liver fibrosis and early HCC tumourigenesis [55]. Similarly, pharmaceutical inhibition of the TGF-β signalling can promote the epithelial differentiation of human adipose-derived mesenchymal stem cells (ADSCs) during EMT through the downregulation of mesenchymal genes (e.g., Slug, zinc finger E-box binding homeobox 1 (ZEB1), integrin α5 (ITGA5), and vimentin (VIM)) and upregulation of epithelial genes (e.g., E-cadherin, epithelial cell adhesion molecule (EpCAM), zonula occludens-1 (ZO-1), occludin, deltaN p63 (δNp63), transcription factor 4 (TCF4), and Twist family bHLH transcription factor (TWIST)) to reverse EMT [56]. Moreover, cancer-associated fibroblasts (CAFs) are an abundant and active population of stromal cells and play a major role in conditioning the TME. TGF-β1 can drive the conversion of normal fibroblasts to CAFs in the TME. For example, CAFs have a key role in the tumourigenesis of oral squamous cell carcinoma (OSCC), the most common cancer of the oral cavity. In this context, TGF-β can induce the conversion of normal oral fibroblasts (NOFs) into CAFs, which are fibronectin type III domain-containing 1 (FNDC1), serpin peptidase inhibitor type 1 (SERPINE1), stanniocalcin 2 (STC2), and type I collagen, to promote the proliferation and invasion of OSCC cells, resulting in a more aggressive tumour phenotypes [57]. The transformation of fibroblasts, which is promoted by the TGF-β/Smad pathway, is also involved in the process of pulmonary fibrosis and early lung cancer tumourigenesis. Consistently, it has been shown that TGF-β1 can induce lung fibroblast transition to myofibroblasts and the expression of α-SMA via the Rho/ROCK and TGF-β/Smad pathways in lung fibroblast–myofibroblast transformation [58]. Furthermore, CAFs also interact with immune cells in the TME, including macrophages, mast cells, natural killer (NK) cells, DCs, myeloid-derived suppressor cells (MDSCs), tumour-associated neutrophils (TANs), and T lymphocytes, to promote tumour progression and immune evasion [59,60]. For example, CAFs and tumour-associated macrophages (TAMs) have been shown to cooperate in driving neuroblastoma progression and invasion [59]. Similarly, CAFs from the HCC TME can enhance the generation of regulatory DCs, to facilitate tumour progression and immune evasion through the production of high levels of TGF-β, impairment of T cell proliferation, and expansion of the Treg population [59]. In addition, the activation of TGF-β signalling in CAFs has been linked to immunosuppression mediated through the induction of a unique set of ECM genes. Moreover, the immunosuppression triggered by TGF-β signalling in CAFs is caused by immune checkpoint blockade and has been shown to be able to lead to anticancer immunotherapy failure in melanoma and bladder cancer [61]. Together, these findings provide solid evidence that TGF-β plays an unparalleled role in mediating tumour growth, metastasis, and chemoresistance and immune evasion through the TME and is therefore a promising target for anticancer intervention.
Besides deregulation of TGF-β expression and activity itself, changes in the expression of components of the TGF-β downstream signalling pathways can also modulate tumourigenesis, cancer progression, and chemoresistance. DC-STAMP domain-containing 1-antisense 1 (DCST1-AS1) and annexin A1 are two genes functioning downstream of TGF-β that play key roles in EMT and cancer chemotherapy resistance. DCST1-AS1 regulates the expression of EMT-related genes including MMP2, MMP9, and E-cadherin. DCST1-AS1 has been found to bind to annexin A1 directly in triple-negative breast cancer (TNBC) cells to modulate the transcription of its downstream regulatory genes involved in EMT and cancer drug resistance [62]. In addition, the TGF-β/MAPK pathway also participates in the transcriptional activation of ETS1 to modulate TGF-β-mediated chemoresistance in liver cancer [63]. Conversely, Ma et al. have found that DNA methyltransferase inhibitor treatment can restore the decreased expression of TGFBR2 caused by hypermethylation on the gene promoter, and this treatment can successfully promote cell cycle arrest and inhibit cancer proliferation in esophageal squamous cell carcinoma (ESCC) [64]. Moreover, the epigenetic silencing of the TGF-β gene, TGFB1, also contributes to trastuzumab resistance in Her2+ breast cancer [65]. Another point worth mentioning is the function of non-coding RNAs in TGF-β pathway regulation and their effects on tumourigenesis. The microRNA miR-495 is one that inhibits the expression of homeobox C6 and the function of TGF-β1. Its overexpression in CSCs contributes to EMT reversion, apoptosis, and the decreased proliferation and migration of CSCs [66]. The miRNA-200 family is another important group of microRNAs that promote cyst formation and ovarian cancer spread through targeting TGF-β expression [67]. Moreover, some recent studies show that circular RNAs, such as circCACTIN and circCCDC66, also affect cancer progression by regulating the TGF-β pathway [68,69], and circular RNA cESRP1 can increase the chemosensitivity of cancer cells by inducing Smad7 and blocking the TGF-β pathway [70].
3. TGF-β in Immune Surveillance
TGF-β is produced not only by cancer cells but also different types of immune cells in the TME [71]. Accordingly, based on immunohistochemistry staining, Ohtani et al. have observed that the latent form of TGF-β1 is mainly localised in immune cells instead of cancer cells, with macrophages, DCs, and T cells being the predominant sources of latent TGF-β1 in cancer stroma [71]. In fact, both the autocrine and the paracrine pathways play an important role in mediating the effects of TGF-β on tumour growth and progression [72]. For example, autocrine TGF-β1 in T cells promotes Th17 cell differentiation [73], and many types of immune cells, including Tregs and macrophages, can perform a similar function by releasing TGF-β1 anchored on the cell surface to prime paracrine signalling [74]. In consequence, TGF-β plays a pivotal role in the immunomodulation of TME and different immune cells (Figure 2). The main functions of TGF-β in cancer immunity are listed in Table 1.
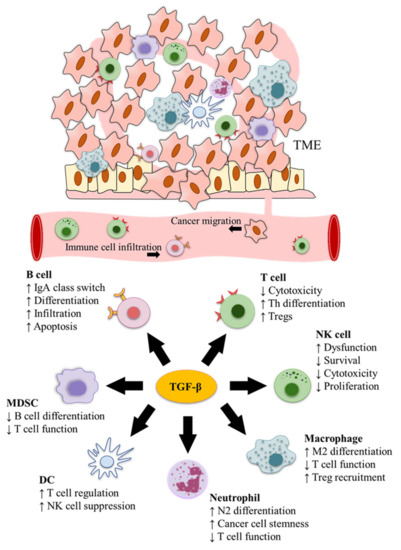
Figure 2.
TGF-β functions in cancer immune surveillance. TGF-β modulates cancer immunity and has pivotal roles in the immunomodulation of the tumour microenvironment (TME) and different immune cells.

Table 1.
Mechanistic effectors of the TGF-β pathway in the TME.
3.1. T Cells
TGF-β affects the survival, activation, and differentiation of different lineages of T cells. These effects are not only caused by TGF-β-induced cell cycle arrest and differentiation in CD4+ and CD8+ T cells directly [101], but the TGF-β-stimulated stromal cells can also affect T cell functions. For example, MSCs can inhibit the activation of T cells by enhancing the expression of latent TGF-β1 complexes on the cell surface [102]. In bone metastasis of castration-resistant prostate cancer, increased TGF-β levels also prevent Th1 lineage development. Combining the TGF-β1 blockade with immune checkpoint blockade therapy can effectively reverse the immunosuppressive state by increasing the number of Th1 and CD8+ T cells to achieve significant tumour regression and improve patient survival [75]. In addition, the blockade of different isoforms of TGF-β, including TGF-β1 and TGF-β2, has been shown to enhance tumour immunity through increasing the immune response from the Th1 population and the production of interferon gamma (IFN-γ), which is more efficient under programmed cell death 1 (PD-1) blockade [76]. It is worth noting that combining TGF-β with other cytokines may stimulate cytotoxic T cell differentiation to produce even more powerful anti-tumour functions. Specifically, IL-4 and TGF-β have been reported to be indispensable for the cell priming and differentiation of IL-9-producing CD4+ Th9 cells, which are a subset of CD4+ T helper cells with a powerful anti-tumour capacity [77]. Moreover, TGF-β can directly promote the differentiation of T helper 17 (Th17) cells to drive cancer progression [103]. Specifically, TGF-β1 increases the population of IL-22-producing Th17 cells via activation of PI3K signalling and thereby promotes tumour growth, aggressiveness, and treatment resistance through the subsequent uncontrolled high levels of IL-22 [78]. In addition, TGF-β signalling also drives the transdifferentiation of Th17 cells into Foxp3+ regulatory T cells (Tregs), and this process directly affects immune responses and results in immune tolerance and immunosuppression in the TME [104,105]. Accordingly, the induction of Tregs will suppress the development of cytotoxic CD8+ T cells and help tumour cells to escape from immune surveillance [106]. TGF-β also promotes the generation of antigen-specific in vitro-induced Tregs (iTregs) by inducing the expression of Foxp3 directly in the presence of IL2. The TGF-β-induced iTregs suppress the proliferation of naïve effector T cells, inhibit the function of Th17, and prevent antigen presentation by DCs [79]. Indeed, the immunosuppression functions mediated by iTregs have shown great potential in ameliorating autoimmune diseases, such as arthritis and stromal keratitis, by promoting differentiation of Th17 and impairing CD4+ T cell function [107,108]. However, these immunosuppressive properties exerted by iTregs can also interfere with cancer treatment and present a barrier to cancer immunotherapy. In concordance, antigen-specific iTregs have been demonstrated to suppress the immunotherapy effects of cancer vaccines and promote cancer growth and progression [109,110]. Not surprisingly, silencing TGF-β1 is also associated with decreased levels of Tregs and enhanced activation of effector T cells in the TME, which has a synergistic effect with VEGFA depletion using siRNA in cancer immunotherapies [80]. In addition, knocking out other genes downstream of TGF-β signalling also suppresses Tregs and enhances cancer immunity. For example, TGFBR2 knockout chimeric antigen receptor-modified T cells can induce Treg conversion and promote tumour regression in vivo [111]. Increasing Tregs induced by TGF-β signalling in turn increases the expression and activation of TGF-β1 [112].
3.2. B Cells
TGF-β can also affect B cell development by regulating the differentiation, proliferation, and apoptosis of B cells [113]. In addition, it is also one of the cytokines that dictate the immunoglobulin IgM to IgA class switching in B cells. In Peyer’s patches, IgA switching requires the interaction between B cells and DCs because the integrin αvβ8 expressed on DCs activates TGF-β to induce B cell maturation and IgA production [114]. In concordance, the B cells located in the vicinity of tumour cells are IgA positive and exert immunosuppressive effects on the TME. In CRC, IgA-producing B cells also express the high levels of programmed death ligand 1 (PD-L1) and TGF-β, which restrict the production and activation of cytotoxic CD8+ T cells and enhance immunosuppression [81]. Besides facilitating the B cell immunoglobulin class switch to IgA, TGF-β/Smad signalling also modulates DNA methylation to block B cell terminal differentiation into plasma cells. In particular, downregulation of TGF-β1 induces the generation of a CD23-negative cell subpopulation during the B cell to plasma cell transition, a process caused by the hydroxymethylation of PR domain 1 (PRDM1), a gene essential for plasma cell fate determination [115]. Additionally, TGF-β1 can inhibit the generation of B lymphoid progenitor colonies by decreasing stromal IL-7 production to limit lymphopoiesis [116], and it inhibits T cell and NK cell proliferation and anti-tumour effects via promoting the function and infiltration of tumour-educated B cells [82]. Moreover, in combination with cyclic adenosine monophosphate, TGF-β1 induces the peripheral blood resting human B cells to undergo apoptosis via a B cell lymphoma 2-independent mechanism [117].
3.3. Natural Killer (NK) Cells
NK cells are a major cellular component functional during innate immune responses and they can kill tumour cells without any priming or prior activation. TGF-β has been found to be able to modulate NK cell function and activity in several ways (Figure 3). Mamessier et al. have identified TGF-β as an important stromal factor involved in tumour-induced dysfunction and enhanced anti-tumour immunity of NK cells in breast cancer [118]. Specifically, high levels of circulating TGF-β1 in the peripheral blood of HCC patients can disturb the balance between CD96, CD226, and T cell immunoglobulin and immunoreceptor tyrosine-based inhibitory motifdomain levels in NK cells, which causes NK cell dysfunction. Patients with a higher cumulative percentage of CD96+ NK cells within tumours also exhibit poorer disease-free survival, suggesting that TGF-β1 regulates the cytotoxic potential and cell immunity of NK cells against tumour cells [119]. TGF-β induces the conversion of NK cells into intermediate type 1 innate lymphoid cells (intILCs) in the TME to restrict innate immune surveillance. This type of innate lymphoid cell (ILC)-like phenotype loses the metastasis-restraining function of NK cells through the expression of CD69, CD49a, and TNF superfamily member 10 (TNFSF10), and the loss of the T-box transcription factor Eomesodermin (EOMES) due to constitutively active TGF-β signalling. In support of this observation, NK cell-deficient mice were unable to restrict lung metastasis in the presence of TGF-β-activated ILCs [120]. Moreover, Smad4 can hamstring this cell conversion by restricting the function of the non-canonical TGF-β pathway in NK cells [121]. Different recognition receptors of NK cells can be triggered by adverse environmental conditions in the TME to activate NK cell cytotoxicity towards tumour cells, while TGF-β1 secreted by the tumour cells demonstrated immunosuppressive effects on NK cells. TGF-β1 probably causes the conversion of tumour-associated NK cells to a non-cytotoxic phenotype in prostate cancer by impairing NK cell receptor D (NKG2D) and DNAX accessory molecule-1 (DNAM-1) [83]. Meanwhile, the loss of phenotype in circulating NK cells with activating receptors, including NKp30, NKp46, NKG2D, and DNAM-1, showed a positive association with tumour progression in gastric cancer. As observed in the previous prostate cancer study, TGF-β1 represses the expression of activating receptors. It is encouraging that the addition of TGFBR1 inhibitors can restore the cytotoxic phenotype of NK cells, which highlights the potential of TGFBR1 and the TGF-β pathway in cancer treatment [122]. Furthermore, the blockade of IL-15, a cytokine responsible for the survival and development of NK cells, can also lead to depletion of the activating receptor NKG2D. Equally, TGF-β1 can also suppress the induction of mTOR activity mediated by IL-15 in murine models of melanoma, breast cancer, and prostate carcinoma. In particular, TGF-β1 inhibits the proliferation of recognition receptors, the production of cytotoxicity cytokines, and the anti-metastatic functions of NK cells [84]. In addition, TGF-β1 can also exert a negative effect on NK cell-mediated immune surveillance via the Smad3/E4BP4 axis to suppress NK cell proliferation and their cancer-killing effects. Accordingly, TGF-β1 suppresses E4BP4 expression in NK cells in a Smad3-dependent manner, and depletion of E4BP4 in NK cells significantly promotes tumour growth [85].
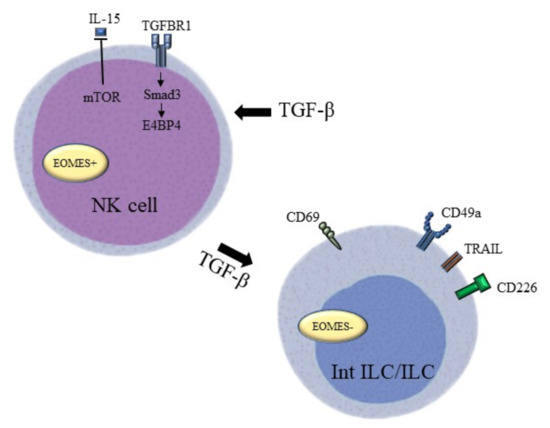
Figure 3.
TGF-β targeting factors that “lock” NK cell functions. TGF-β impairs the balance between activating signals and inhibitory signals on NK cells to block the activation of NK cells. Their anti-tumour activity is reduced by downregulating the mTOR pathway and upregulating the Smad3/E4BP4 pathway. The conversion of NK cell to innate lymphoid cell under the high levels of TGF-β leads to the loss of the metastasis-restraining function of NK cells through the expression of CD69, CD49a, and TRAIL, and the loss of EOMES.
3.4. Myeloid-Derived Suppressor Cells (MDSCs)
MDSCs play an essential role in tumour immune evasion by regulating the anti-tumour immune responses during cancer progression. In NSCLC, the TGF-β pathway can stimulate the expression of CD39 and CD73 in MDSCs through activation of HIF-1α to inhibit T cell and NK cell function, thus helping cancer cells to avoid immune surveillance [86]. Additionally, TGF-β1 secreted by MDSCs can accelerate tumour growth and increase tumour burden in lung cancer in response to lung microenvironment changes induced by carbon nanomaterial [123]. Wang et al. have also reported a potential mechanism of TGF-β1 in MDSC-derived immunosuppressive function. In this scenario, MDSC-derived TGF-β1 inhibits IL-7 function, blocks signal transducer and activator of transcription 5A (STAT5A) phosphorylation, and impairs B cell differentiation [87]. In addition, MDSCs educated under TGF-β stimulation can suppress CD8+ T lymphocyte cancer-killing effects by secreting nitric oxide synthase 2or arginase 1 [88]. At odds with most previous findings, Jayaraman et al. have also uncovered that although cancer-induced MDSCs play a key role in tumour immune evasion, they can be conditioned with TGF-β1 to acquire a novel immune-stimulatory phenotype, losing the ability to inhibit T cell proliferation and acquiring an enhanced antigen-presenting capability. Thus, these TGF-β1-induced TGFβ-MDSCs show decreased immunosuppressive effects, enhanced antigen-presenting ability, and increased cancer-killing activity [89].
3.5. Macrophages
Macrophages are phagocytes that are the first line of defence against pathogenic tissue damage. They have heterogeneous populations and are typically defined as the classically activated M1 macrophages or the alternatively activated M2 macrophages [124]. TAMs are important components of the TME and are strongly associated with poor prognosis in solid tumours, while M2-like TAMs have also been suggested to serve as prominent metastasis promoters with anti-inflammatory functions in the development of TME [125,126].
TGF-β is an important cytokine for TAM function, and it stimulates monocyte recruitment and macrophage differentiation. This cytokine has been shown to inhibit tumour regression by hindering the activation of MHC-II+ TAMs and production of IFN-α. In consequence, the blockade of the TGF-β pathway or its functions may improve IFN-α-mediated cancer therapy [127]. Moreover, TGF-β1 has a well-established role in the M2 phenotype differentiation of TAMs. Zhang et al. have found that TGF-β1 promotes the expression of anti-inflammatory cytokine IL-10 and suppresses the pro-inflammatory cytokines, including tumour necrosis factor alpha (TNF-α) and IL-12, through the TGF-β/Snail signalling pathway [90]. In addition, the expression of TGF-β is also regulated by other pro-inflammatory cytokines, including IL-6 and IL-17, in TAMs [91,92,93]. In previous studies of gastric cancer, IL-6 has been shown to induce the M2 differentiation and increase the expression of TGF-β and IL-10 in TAMs via promoting STAT3 phosphorylation and activation, and this phenomenon augments the proliferation and migration of cancer cells [91]. Similar mechanisms were also found in the progression of NSCLC, where highly expressed plasminogen activator inhibitor 1 (PAI-1) enhances the expression of cytokines, including C-C motif chemokine ligand 17 (CCL17), CCL22, IL-6, and TGF-β1, in TAMs to promote cancer progression [92]. The upregulated TGF-β1 also participates in M2 differentiation of TAMs in glioma in a hypoxia-mediated manner via the AKT/PI3K pathway [128]. Together, these findings underscore the immense therapeutic value of TGF-β1 in cancer immunosuppression.
Additionally, macrophage-secreted TGF-β also affects the development of TME by regulating other immune cells. For example, TGF-β secreted by macrophages can suppress cytotoxicity of T cells in malignant pleural effusion, and TAM-derived TGF-β has the ability to induce the expression of CCL22 and promote the recruitment of Tregs. The TGF-β-activated Tregs can also secrete IL-8 in the TME to stimulate further TGF-β production in a positive feed-forward loop [129]. Interestingly, TAMs can also modulate TGF-β functions and promote cancer progression by secreting pro-tumour exosomal miRNAs. The microRNA miRNA-501-3p is one of the exosomal miRNAs which are involved in PDA formation and metastasis. In these cases, TGF-β functions through inhibiting type III TGF-β receptor (TGFBR3) expression and activating the TGF-β pathway [130].
3.6. Dendritic Cells (DCs)
TGF-β also regulates the adaptive immune response through modulating the function of DCs and their interactions with other immune cells, such as Tregs and NK cells. In the TME, tumour-derived TGF-β inhibits IFN-α, TNF-α, and IL-6 production in tumour-associated plasmacytoid dendritic cells (pDCs) to foster their immune tolerance functions [131]. In addition, TGF-β produced by the TME can also alter DC functions. Enhanced TGF-β signalling within the TME upregulates both the immunoregulatory enzyme indoleamine 2,3-dioxygenase (IDO) in pDCs and the CCL22 chemokine in myeloid DCs (mDCs). The TGF-β-mediated changes in these different DC populations in the TME can direct Treg infiltration and the suppression of anti-tumour immunity [94]. In addition, TGF-β1 also induces the expression of PD-L1 and TNFSF18 in DCs to enhance Treg expansion in lung cancer [95]. TGF-β signalling activation in cancer cells may inhibit DC functions and regulate immune tolerance in the TME. For example, Zhong et al. have found that high levels of TGF-β1 in HCC cells suppress the production and function of DCs and augment immune tolerance [96]. Moreover, blocking signal transduction of TGF-β pathways in self-differentiated DCs increases the IFN-γ production in CD3+ T cells and enhances their cancer-killing effects [132]. These results provide important evidence for DC-mediated immune regulation in the TME.
On the other hand, the TGF-β expressed by DCs also has immunomodulatory effects on the TME. Previous studies have shown that CD11b+CD103- tumour-infiltrating DCs expressing high levels of TGF-β1 and IL-23 can induce tumour-promoting FOXP3-CD4+ Tregs in PDA. These IL-10+IL-17+IFN-γ+ Tregs can confer immunotherapy resistance and poor survival in cancer patients [133]. Besides, DCs educate naïve CD8+ T cells for tissue-resident memory through secreting TGF-β1 [42]. DCs also suppress NK cell activities via TGF-β1, lymphotoxin alpha, IL-12 secretion, and STAT3 phosphorylation [134]. All these studies reveal the diverse functions of TGF-β in immunomodulation and highlight the potential of DC-based cancer immunotherapy. In addition to affecting immune cell infiltration and TME development, DCs can also achieve immunomodulation of TGF-β signalling pathways through secreting antigen-specific exosomes. Exosomes derived from α-fetoprotein+ DCs have been used in novel cell-free vaccines in HCC treatment and they have successfully suppressed tumour growth and increased survival in a tumour-bearing mice model through increasing IFN-γ+CD8+ T cells, IFN-γ, and IL-2, and decreasing CD25+FOXP3+ Tregs, as well as IL-10, and TGF-β production [135]. Some recent studies provided new evidence of TGF-β signalling effects on DCs and tumourigenesis. For example, the single-nucleotide polymorphism (SNP) 509C/T on the TGF-β1 gene promoter region can influence the infiltration of DCs at the invasive margin of the tumour, with the T-allele of the TGFB1 -509C/T SNP having a protective factor for the development of CRC. DCs have always been recognised as the key cellular component of the inflammatory TME and the infiltration of tumours by DCs is associated with favourable prognosis and fewer metastases [136].
3.7. Neutrophils
Neutrophils represent 60–80% of the circulating leucocytes and play a pivotal role in host defence. They have high mobility and display a strong activation to release cytokines, defensins, and reactive oxygen species into the TME [137]. TGF-β1 derived from platelets stimulates the recruitment of neutrophils to tumours and promotes metastasis at the early stages of cancer in tumour-bearing mice, hinting at a crucial role of TGF-β1 in shaping the neutrophil responses and the related tumour biology that promotes metastasis [138]. Fridlender et al. have uncovered that tumour-associated neutrophils (TANs) are programmed to induce cell populations with either an anti-tumourigenic N1 or pro-tumourigenic N2 phenotype, and that the TGF-β pathway also regulates this type of polarisation. In the TME, N2 neutrophils are found to have a TGF-β-activated phenotype that releases pro-tumour chemokines and proteases, including CCL2, CCL5, neutrophil elastase, and cathepsin G, that support immunosuppression. By contrast, TGF-β also suppresses N1 neutrophils that express anti-tumour chemokines, modulate arginase reduction, and provide cancer-killing activities [139]. By characterising the development of neutrophil populations into mature high-density neutrophils, mature low-density neutrophils, and immature low-density neutrophils in the TME, Sagiv et al. have further demonstrated that TGF-β treatment forces the N1-like high-density neutrophils to converse into N2-like low-density neutrophils. This conversion has been proven to result in a subsequent positive association between TANs and tumour burden [140]. Neutrophils found in various cancer types, including breast, lung, and colorectal cancer, are often associated with unfavourable patient outcomes [141,142]. Haider et al., have revealed that the high degree of neutrophil recruitment and the poor survival rate at the advanced stage of HCC are correlated with TGF-β1 stimulation. Functional and mechanical analyses in HCC models have also shown that long-term TGF-βl exposure can promote C-X-C motif chemokine ligand 5 (CXCL5) expression to induce a high frequency of neutrophil recruitment [97]. Once the neutrophil numbers increase, the oncogenic hepatocytes can further raise the activity of the TGF-β pathway to maintain the neutrophil recruitment process [98]. Besides that, TANs can also secret TGF-β2 to trigger miR-301-3p-related stem cell characteristics in the oncogenic hepatocytes, which will then secrete a higher level of CXCL5 and recruit more neutrophils to infiltrate the tumour and generate a pro-tumour TME. These interactions between TGF-β2 and neutrophils in the TME form a positive feedback loop to suppress anti-tumour gene expression and promote angiogenesis in HCC [143]. In addition, the TGF-β2-activated neutrophils can further facilitate tumour proliferation by modulating the immune system to lead to differentiation and response changes in other immune cells. It has been proposed that the TGF-β signalling through TGFBR1 in neutrophils participates in the production of TGF-β2 to trigger immunosuppressive activities that can attenuate the T cell response and thereby create a metastatic niche in CRC [99]. Both mouse tumour neutrophils and CRC patient-derived neutrophils have been shown to be able to further inhibit T cell activation by releasing TGF-β1 under high levels of MMP9 within the TME [100]. Collectively, these results suggest that tumour progression in the TME involves the recruitment, differentiation, and activation of neutrophils that are tightly controlled by TGF-β.
4. Therapeutic Implications
Cancers develop adaptive mechanisms to suppress the host immune system to facilitate tumour growth and progression. Incidentally, these cancer-evolved immunosuppressive mechanisms are also involved in cancer immunotherapy resistance. Cancers frequently express TGF-β, which drives immune dysfunction in the TME by promoting Tregs and inhibiting the cytotoxic CD8+ and helper Th1 T cells. As TGF-β contributes to immunosuppression and immunotherapy resistance, it is therefore a pivotal target for cancer immunotherapy (Table 2). Based on one of the more substantial pan-cancer transcriptional analyses of ECM gene dysregulation, TGF-β activation in CAFs is a crucial indicator for ECM dysfunction in cancer and this CAF-driven ECM dysfunction is identified as a single important event responsible for the failure of PD-1 blockade cancer immunotherapy [61]. In addition, TGF-β has also been shown to induce PD-L1 expression in cancer cells to further hinder the immunotherapy effects from PD-L1 blockade treatment [144]. As a result, TGF-β inhibition is a promising strategy for overcoming immune checkpoint blockade therapy resistance. Current therapeutic approaches for blocking TGF-β signalling in cancer immunotherapy involve antibodies, proteins, peptides, small molecule inhibitors, and some natural compounds.
Table 2.
Cancer immunotherapeutic strategies based on the regulation of the TGF-β pathway.
As a direct TGF-β-targeting agent, the high-affinity artificial TGF-β1 antibody SRK-181 has been employed successfully in anti-PD-1 cancer treatments to override immunotherapy resistance. Equally, the co-administration of SRK-181 and anti-PD-1 has also demonstrated increased anti-tumour effects and survival benefits in bladder cancer-bearing mice by increasing infiltering CD8+ T cells and decreasing immunosuppressive myeloid cells [145]. Another chemical TGF-β inhibitor, tranilast, which was originally developed as an antiallergic drug, has also demonstrated proven anti-fibrosis and anti-tumour effects. Panagi et al. have shown that combining tranilast and the cytotoxic chemotherapy drug Doxil significantly inhibits tumour progression and improves the efficacy of anti-PD-1 and anti-cytotoxic T lymphocyte-associated protein 4 (CTLA-4) in TNBC [146]. Besides, immunotherapy using antibodies targeting PD-1, TIM-3, and LAG-3 in combination with anti-IL-6 and -TGF-β1 antibodies can enhance treatment outcomes of the E7-TM mRNA vaccine against SOX2 in metastatic lung cancer [147]. Furthermore, a recent study in a lung tumour-bearing mouse model has also shown that fusing a tumour antigen peptide to annexin A5 (AnxA5) significantly enhances its immunogenicity and anti-tumour efficacy when administered after chemotherapy. In this context, cisplatin treatment can activate TGF-β to create an immunosuppressive TME and annexin A5 works as an immune checkpoint inhibitor to reprogramme the TME and restore tumour sensitivity by blocking the expression of TGF-β3 [148]. To circumvent the resistance to checkpoint blockade immunotherapy and target multiple cancer specific pathways concurrently, multifunctional fusion proteins have also been designed and developed as novel cancer immunotherapeutic strategies. Bintrafusp alfa, also known as M7824, is a fusion protein that contains the extracellular domain of TGFBR2 fused to an IgG1 antibody of PD-L1, which traps TGF-β and targets PD-L1 at the same time [149]. This antibody–drug conjugate (ADC) designed to simultaneously target the two immunosuppressive pathways of TGF-β and PD-L1 has shown some encouraging results in some early clinical studies. In an ongoing phase I clinical trial (NCT02517398), bintrafusp alfa has demonstrated good clinical response, safety, and tolerance in advanced head and neck squamous cell carcinoma (HNSCC) that progressed after platinum therapy [150]. In addition, a joint targeting of CXCR1/2, TGF-β1, and PD-L1 using the CXCR1/2 inhibitor SX-682 and bintrafusp alfa also reduces EMT, enhances the infiltration of effector T cells and promotes the blockade of granulocytic myeloid cells in breast and lung cancer mouse models in vivo [151]. Similarly, bifunctional fusion antibodies that simultaneously target the immune checkpoint and disable TGF-β signalling can significantly enhance the efficacy of cancer immunotherapy [152]. Accordingly, compared with traditional CTLA-4 antibody (ipilimumab)- and PD-L1 antibody (atezolizumab or avelumab)-mediated immunotherapy, bifunctional antibody–ligand traps comprising an antibody targeting CTLA-4 or PD-L1 fused to a TGFBR2 ectodomain sequence are more effective in reducing tumour-infiltrating Tregs and inhibiting tumour progression in the TME [152]. Overall, these multifunctional TGF-β pathway-targeting fusion proteins exhibit a good efficacy and manageable safety profile in patients with a range of advanced solid tumours and can therefore have the potential to be developed into effective cancer immunotherapeutic agents [153,154].
Furthermore, due to the immunosuppressive effects of TGF-β on immune cells, targeting TGF-β can restore anti-tumour activity of immune cells. For instance, galunisertib, an inhibitor of TGFBR1, blocks the function TGF-β signalling and enhances the anti-tumour effects of CD133 and Her2 chimeric antigen receptor (CAR) T cells against both glioma and breast cancer [155]. Moreover, SD-208, a novel inhibitor of TGFBR1, induces the infiltration of immune cells, including macrophages, CD8+ T cells, and NK cells, to enhance the immunogenicity of glioma cells and inhibit tumour progression [156]. Furthermore, SD-208 also increases the viability of CD4+ and CD8+ ROR1-specific CAR T cells to enhance the cancer-killing effects [157]. On the other hand, TGFBR1 inhibitor SB431542 is able to decrease tumour burden by eliminating the immune suppression induced by Tregs and regulatory B cells (Bregs) and restoring normal T cell function [158]. For NK cells, the immunosuppressive effects of TGF-β1 on E4BP4 and its target IFN-γ in natural killer NK-92 cells can be reversed via Smad3 silencing in xenograft mouse models of human hepatoma and melanoma [159]. Consistently, the expression of a chimeric receptor with TGFBR2 extracellular and transmembrane domains and the intracellular domain of NKG2D can also neutralise the high level TGF-β-derived immunosuppressive TME and restore higher killing ability and IFN-γ production capacity in the transferred NK cells. In this context, the hybrid receptor converts the TGF-β1 suppressive signals into activating signals. In further agreement, other studies have shown that the anti-tumour effect of TGF-β1 is associated with TGFBR2 activity and is mediated by regulating the cytotoxicity of NK cells, as tumours grow irrespective of TGFBR2 inhibition in mice that lack NK cells [160]. Moreover, the TGF-β superfamily member activin-A can also weaken human peripheral NK cell cytokine production through inhibitory signalling through the type I and II activin receptors, whereas the blockade of activin-A efficiently downregulates cNK-ILC1-like cell differentiation in the TME by downregulating CD49a and CD69, and upregulating EOMES expression in human NK cells simultaneously [161]. Additionally, promoting the differentiation of tumour stem cells and inhibiting proliferation by regulating the TGF-β pathway also provide feasible cancer treatments. For example, RGFP966, an inhibitor of histone deacetylase 3, can block the TGF-β pathway via negative feedback of Smad7, and induce glioma stem cell differentiation into astrocytes [162]. Meanwhile, both pirfenidone and cabazitaxel can achieve effective inhibition of cancer cell proliferation and viability by targeting the TGF-β pathway [163,164].
In addition, several chemicals and natural compounds have also been demonstrated to facilitate TGF-β1-based cancer immunotherapy. For example, retinoic acid can strengthen the anti-tumour effects of kartogenin in the blockage of the TGF-β pathway and inhibition of the viability of retinoid-resistant tumour cells [165]. Furthermore, Lin et al. have found that chidamide can also attenuate the repression of E-cadherin expression caused by TGF-β, and arrest both EMT and cell migration in lung cancer [166]. Similarly, vanadium can also restrict EMT by regulating the TGF-β pathway. Accordingly, the combination of vanadium and the traditional chemotherapy drug carboplatin induces a more potent G0/G1 cell cycle arrest [167]. Similarly, astragaloside IV also achieves cancer treatment via TGF-β-induced cell cycle arrest in the G0/G1 phase in vulvar squamous cell carcinoma, which enhanced cancer cell apoptosis simultaneously by increasing the expression of apoptotic genes including cleaved caspase-3 [168]. Meanwhile, heteronemin, a natural compound extracted from marine sponges, can reduce the expression of TGF-β1 and intercellular adhesion molecule 1, and inhibits cancer cell adhesion, motility, and proliferation in the bile duct cancer, cholangiocarcinoma [169]. Moreover, a natural bloodroot plant extract sanguinarine can also suppress the proliferation and colony formation of HCC cells via targeting the HIF-1α/TGF-β loop. Primarily, the compound inhibits EMT and the PI3K/AKT pathway and causes, therefore, decreased downstream cancer cell proliferation, migration, and tumour growth [170]. Furthermore, the herbal extract PM014 can also inhibit the generation of pathological lesions in lung diseases, including inflammatory infiltration, fibrosis, and EMT, by repressing TGF-β1 signalling [172]. Furthermore, medical formulas such as “Wenshen zhuanggu” have demonstrated cancer treatment effects via targeting the TGF-β pathway [171]. Some of the above drugs and therapy have been evaluated in clinical trials of cancer treatments. The clinical trials evaluating TGF-β pathway inhibitors are listed in Table 3.
Table 3.
Clinical trials of the TGF-β pathway inhibitors for cancer treatment.
Besides these novel drugs and combinational therapies targeting TGF-β, accurate and effective biomarkers are also required for evaluating and monitoring the effectiveness of these new treatments. Moreover, reliable biomarkers are also necessary for identifying the susceptible and effective population before administering treatment. For example, an increased ratio of CD8+/CD4+ CD3+ T cells and a number of IFN-γ+CD8+ CD3+ T cells can be used to predict enhanced tumour-specific T cell responses in the combination therapy of galunisertib and intestinal microbiota in HCC [173]. Similarly, an increased number of IFN-γ+CD8+ T cells are also an indicator of the successful combination treatment of bintrafusp alfa and radiotherapy [150]. Pu et al. have identified an integrated immune ratio based on the number of infiltrating CD8+ T cells and FOXP3+ Tregs to predict tumour suppression in the combination immunotherapy of CD25, TGF-β, and PD-1 blockade [174]. In melanoma, the treatment mediated by TGFBR1 inhibitor SB505124 in the presence of IL-12 overexpression demonstrates enhanced immune responses, and NK1.1 is a potential biomarker for increased effector cells and enhanced cancer-killing effects [175]. In the combined therapy of galunisertib and paclitaxel, decreased ALDH+ cancer stem-like cell number also reflects treatment efficiency [176].
Radiotherapy is one of the most frequently employed therapeutic approaches for the treatment of cancer, using high doses of ionising radiation to cause cancer cell death and tumour shrinkage. T cells directed to endogenous tumour antigens have been proposed to be crucial mediators of tumour regression [177]. However, ionising radiation also leads to the release of several cytokines, which include TGF-β that has a negative impact on the effectiveness of radiotherapy on tumour regression [178]. Appropriately, a recent study has shown that antibody-mediated TGF-β blockade using the TGFβ-neutralising mAb 1D11 can efficiently enhance radiation-induced cancer treatment through enhancing cytotoxic CD8+ T cell responses in a mouse breast cancer model [177]. Similarly, this beneficial effect of TGF-β blockade can also be observed in cancer radioimmunotherapy. It has been shown that the TGFβ-neutralising mAb 1D11 can markedly enhance the abscopal effects and overall treatment efficacy of combined radio- and immunotherapy in both mouse breast and colorectal cancer models [178]. Again, this synergistic TGF-β blockade and radioimmunotherapy function is mediated through disabling the TGF-β-mediated suppressive function on the anticancer CD8+ T cells [178]. Together, these findings provide strong evidence of the overwhelming positive effects of TGF-β signalling blockade in cancer chemotherapy and radiotherapy.
In summary, TGF-β signalling displays anti-proliferative and tumour suppressive functions during early tumourigenesis, but tumour cells often become refractory to TGF-β-mediated growth inhibition as cancer progresses. Moreover, at late stages of malignancy, TGF-β also targets the vascular, immune, and fibroblastic cellular components of the TME to promote tumour progression and cancer therapy resistance. Conventional chemotherapy and radiotherapy are the most common first-line treatments for the majority of cancers and function primarily by inducing apoptosis, autophagy and cell cycle arrest in tumour cells. However, resistance to these agents often emerges in patients, leading to suboptimal efficacy and disease relapse. As the development of resistance of cancer cells to chemotherapy and radiotherapy is commonly mediated by TGF-β in the TME, chemotherapy and radiotherapy resistance can therefore be reverted by TGF-β inhibition. However, the dual role of TGF-β in cancer development also indicates that long-term treatment strategies with TGF-β inhibitors might promote tumour initiation and also TGF-β-based therapy resistance. Appropriately, current strategies for the use of TGF-β inhibitors as cancer therapeutics involve predominantly short-term and combinatorial therapies. Ultimately, further novel and innovative anticancer immunotherapeutic strategies will stem from a better understanding of the intricate interactions between the malignant transformation, immune system, and the TME, as well as their relationships with TGF-β signalling. In particular, the diversity of TGF-β regulation and functions in different cell types and settings has also rendered anti-TGF-β therapeutic strategies challenging. In consequence, further research into the role and regulation of the TGF-β signalling molecules and target genes in a context-dependent and cell type-specific manner will allow us to dissect and target specific functions of TGF-β to limit cancer initiation, restrict cancer progression and override treatment resistance.
5. Conclusions
Extensive studies conducted over the last three decades have demonstrated that TGF-β is an essential immune-suppressive cytokine. Evidently, TGF-β plays diverse roles in the development and maintenance of the immunosuppressive TME. This implies that targeting TGF-β may lead to effective cancer treatment through modulating immunoresponse in the TME. However, further research is required to fully comprehend the TGF-β signalling pathways and their roles in malignant transformation, the immune system, and the TME. For example, unveiling the comprehensive TGF-β modulation network and cofactors involved in cancer immunity will contribute to the development of novel TGF-β-based cancer immunotherapy strategies.
Author Contributions
V.W.X., J.Y.-F.C., C.A.G.C., and A.H.-K.C. were responsible for literature research and writing. W.K., K.-T.L., K.-F.T., and H.-Y.L. reviewed the manuscript and E.W.-F.L. made significant revisions to the drafts. P.M.-K.T. supervised and finalised of this work. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Research Grants Council of Hong Kong (GRF 14106518, 14111019, 14111720); Faculty Innovation Award (4620528) and Direct Grant for Research (4054440, 4054510) of The Chinese University of Hong Kong.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ALDH | Aldehyde Dehydrogenases |
| α-SMA | Alpha Smooth Muscle Actin |
| Bregs | Regulatory B Cells |
| CAF | Cancer-associated Fibroblast |
| CAR | Chimeric Antigen Receptor |
| CDK | Cyclin-dependent Kinase |
| CEBPA | CCAAT/Enhancer-binding Protein Alpha |
| CRC | Colorectal Cancer |
| CREB | cAMP-response Element Binding Protein |
| CSCs | Cancer Stem Cells |
| CTLA-4 | Cytotoxic T lymphocyte-associated Protein 4 |
| DAPKs | Death-associated Protein Kinases |
| DCs | Dendritic Cells |
| DCST1-AS1 | DC-STAMP Domain-containing 1-Antisense 1 |
| DNAM-1 | DNAX Accessory Molecule-1 |
| ECM | Extracellular Matrix |
| EGFR | Epidermal Growth Factor Receptor |
| EMT | Epithelial-to-Mesenchymal Transition |
| EOMES | Eomesodermin |
| ESCC | Esophageal Squamous Cell Carcinoma |
| eTRM | Epithelial Resident Memory T cells |
| FOXO1 | Forkhead Box O1 |
| GOLM1 | Golgi Membrane Protein 1 |
| HCC | Hepatocellular Carcinoma |
| HIF-α | Hypoxia Inducible Factor 1 Subunit Alpha |
| HNSCC | Head and Neck Squamous Cell Carcinoma |
| IFN-γ | Interferon-γ |
| IL | Interleukin |
| ILC | Innate Lymphoid Cell |
| intILCs | Intermediate Type 1 Innate Lymphoid Cells |
| iTregs | Induced Regulatory T cells |
| LAP | Latency-associated Peptide |
| LLC | Large Latent Complex |
| MAPK | Mitogen-activated Protein Kinase |
| MDSCs | Myeloid-derived Suppressor Cells |
| MKKs | MAP Kinase Kinases |
| MMP | Matrix Metallopeptidase |
| MSCs | Mesenchymal Stem Cells |
| mTOR | Mammalian Target of Rapamycin |
| NK | Natural Killer |
| NKG2D | NK Cell Receptor D |
| NSCLC | Non-Small Cell Lung Cancer |
| PD-1 | Programmed Cell Death 1 |
| PD-L1 | Programmed Death Ligand 1 |
| PDA | Pancreatic Ductal Adenocarcinoma |
| PI3K | Phosphoinositide 3-kinase |
| PPM1D | Protein Phosphatase 1D |
| pDC | Plasmacytoid Dendritic Cell |
| ROCKs | Rho-associated Protein Kinases |
| SNP | Single-nucleotide Polymorphism |
| SOX4 | SRY-box Transcription Factor 4 |
| TAMs | Tumour-associated Macrophages |
| TANs | Tumour-associated Neutrophils |
| TApDCs | Tumour-associated Plasmacytoid Dendritic Cells |
| TF | Transcription Factor |
| TGF-β | Transforming Growth Factor-β |
| TGFBR1 | Type I TGF-β Receptor |
| TGFBR2 | Type II TGF-β Receptor |
| Th | T Helper Cell |
| TME | Tumour Microenvironment |
| TNBC | Triple-negative Breast Cancer |
| TNF-α | Tumour Necrosis Factor Alpha |
| Tregs | Regulatory T Cells |
| VEGFA | Vascular Endothelial Growth Factor A |
References
- Du, Y.; Sun, J.; Liu, X.; Nan, J.; Qin, X.; Wang, X.; Guo, J.; Zhao, C.; Yang, J. TGF-beta2 antagonizes IL-6-promoted cell survival. Mol. Cell. Biochem. 2019, 461, 119–126. [Google Scholar] [CrossRef]
- Zheng, S.; Zhou, H.; Chen, Z.; Li, Y.; Zhou, T.; Lian, C.; Gao, B.; Su, P.; Xu, C. Type III Transforming Growth Factor-β Receptor RNA Interference Enhances Transforming Growth Factor β3-Induced Chondrogenesis Signaling in Human Mesenchymal Stem Cells. Stem Cells Int. 2018, 2018, 4180857. [Google Scholar] [CrossRef] [PubMed]
- Navarro, R.; Tapia-Galisteo, A.; Martín-García, L.; Tarín, C.; Corbacho, C.; Gómez-López, G.; Sánchez-Tirado, E.; Campuzano, S.; González-Cortés, A.; Yáñez-Sedeño, P.; et al. TGF-β-induced IGFBP-3 is a key paracrine factor from activated pericytes that promotes colorectal cancer cell migration and invasion. Mol. Oncol. 2020, 14, 2609–2628. [Google Scholar] [CrossRef] [PubMed]
- Cave, D.D.; Di Guida, M.; Costa, V.; Sevillano, M.; Ferrante, L.; Heeschen, C.; Corona, M.; Cucciardi, A.; Lonardo, E. TGF-β1 secreted by pancreatic stellate cells promotes stemness and tumourigenicity in pancreatic cancer cells through L1CAM downregulation. Oncogene 2020, 39, 4271–4285. [Google Scholar] [CrossRef] [PubMed]
- Colak, S.; Ten Dijke, P. Targeting TGF-β Signaling in Cancer. Trends Cancer 2017, 3, 56–71. [Google Scholar] [CrossRef]
- Guo, P.; Xing, C.; Fu, X.; He, D.; Dong, J.T. Ras inhibits TGF-β-induced KLF5 acetylation and transcriptional complex assembly via regulating SMAD2/3 phosphorylation in epithelial cells. J. Cell. Biochem. 2020, 121, 2197–2208. [Google Scholar] [CrossRef]
- Oshimori, N.; Oristian, D.; Fuchs, E. TGF-β promotes heterogeneity and drug resistance in squamous cell carcinoma. Cell 2015, 160, 963–976. [Google Scholar] [CrossRef]
- Zhang, B.; Ye, H.; Ren, X.; Zheng, S.; Zhou, Q.; Chen, C.; Lin, Q.; Li, G.; Wei, L.; Fu, Z.; et al. Macrophage-expressed CD51 promotes cancer stem cell properties via the TGF-β1/smad2/3 axis in pancreatic cancer. Cancer Lett. 2019, 459, 204–215. [Google Scholar] [CrossRef]
- Leng, Z.; Li, Y.; Zhou, G.; Lv, X.; Ai, W.; Li, J.; Hou, L. Krüppel-like factor 4 regulates stemness and mesenchymal properties of colorectal cancer stem cells through the TGF-β1/Smad/snail pathway. J. Cell. Mol. Med. 2020, 24, 1866–1877. [Google Scholar] [CrossRef]
- Miyazawa, K.; Miyazono, K. Regulation of TGF-β Family Signaling by Inhibitory Smads. Cold Spring Harb. Perspect. Biol. 2017, 9, a022095. [Google Scholar] [CrossRef]
- Luo, Y.; Ren, Z.; Du, B.; Xing, S.; Huang, S.; Li, Y.; Lei, Z.; Li, D.; Chen, H.; Huang, Y.; et al. Structure Identification of ViceninII Extracted from Dendrobium officinale and the Reversal of TGF-β1-Induced Epithelial–Mesenchymal Transition in Lung Adenocarcinoma Cells through TGF-β/Smad and PI3K/Akt/mTOR Signaling Pathways. Molecules 2019, 24, 144. [Google Scholar] [CrossRef]
- Jin, S.; Gao, J.; Qi, Y.; Hao, Y.; Li, X.; Liu, Q.; Liu, J.; Liu, D.; Zhu, L.; Lin, B. TGF-β1 fucosylation enhances the autophagy and mitophagy via PI3K/Akt and Ras-Raf-MEK-ERK in ovarian carcinoma. Biochem. Biophys. Res. Commun. 2020, 524, 970–976. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhao, X.; Sun, Q.; Jiang, Y.; Zhang, W.; Luo, J.; Li, Y. Synergic effect of PD-1 blockade and endostar on the PI3K/AKT/mTOR-mediated autophagy and angiogenesis in Lewis lung carcinoma mouse model. Biomed. Pharm. 2020, 125, 109746. [Google Scholar] [CrossRef]
- Zajac, O.; Raingeaud, J.; Libanje, F.; Lefebvre, C.; Sabino, D.; Martins, I.; Roy, P.; Benatar, C.; Canet-Jourdan, C.; Azorin, P.; et al. Tumour spheres with inverted polarity drive the formation of peritoneal metastases in patients with hypermethylated colorectal carcinomas. Nat. Cell Biol. 2018, 20, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Pickup, M.; Novitskiy, S.; Moses, H.L. The roles of TGFβ in the tumour microenvironment. Nat. Rev. Cancer 2013, 13, 788–799. [Google Scholar] [CrossRef]
- Kawarada, Y.; Inoue, Y.; Kawasaki, F.; Fukuura, K.; Sato, K.; Tanaka, T.; Itoh, Y.; Hayashi, H. TGF-β induces p53/Smads complex formation in the PAI-1 promoter to activate transcription. Sci. Rep. 2016, 6, 35483. [Google Scholar] [CrossRef] [PubMed]
- Seoane, J.; Gomis, R.R. TGF-β Family Signaling in Tumor Suppression and Cancer Progression. Cold Spring Harb. Perspect. Biol. 2017, 9, a022277. [Google Scholar] [CrossRef]
- Hinge, A.; Xu, J.; Javier, J.; Mose, E.; Kumar, S.; Kapur, R.; Srour, E.F.; Malik, P.; Aronow, B.J.; Filippi, M.D. p190-B RhoGAP and intracellular cytokine signals balance hematopoietic stem and progenitor cell self-renewal and differentiation. Nat. Commun. 2017, 8, 14382. [Google Scholar] [CrossRef]
- Ma, K.; Zhang, C.; Li, W. TGF-β is associated with poor prognosis and promotes osteosarcoma progression via PI3K/Akt pathway activation. Cell Cycle 2020, 19, 2327–2339. [Google Scholar] [CrossRef]
- Yang, X.; Wei, C.; Liu, N.; Wu, F.; Chen, J.; Wang, C.; Sun, Z.; Wang, Y.; Liu, L.; Zhang, X.; et al. GP73, a novel TGF-β target gene, provides selective regulation on Smad and non-Smad signaling pathways. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 588–597. [Google Scholar] [CrossRef]
- Santhana Kumar, K.; Neve, A.; Guerreiro Stucklin, A.S.; Kuzan-Fischer, C.M.; Rushing, E.J.; Taylor, M.D.; Tripolitsioti, D.; Behrmann, L.; Kirschenbaum, D.; Grotzer, M.A.; et al. TGF-β Determines the Pro-migratory Potential of bFGF Signaling in Medulloblastoma. Cell Rep. 2018, 23, 3798–3812. [Google Scholar] [CrossRef] [PubMed]
- Park, D.S.; Yoon, G.H.; Kim, E.Y.; Lee, T.; Kim, K.; Lee, P.C.; Chang, E.J.; Choi, S.C. Wip1 regulates Smad4 phosphorylation and inhibits TGF-β signaling. EMBO Rep. 2020, 21, e48693. [Google Scholar] [CrossRef] [PubMed]
- Thien, A.; Prentzell, M.T.; Holzwarth, B.; Kläsener, K.; Kuper, I.; Boehlke, C.; Sonntag, A.G.; Ruf, S.; Maerz, L.; Nitschke, R.; et al. TSC1 activates TGF-β-Smad2/3 signaling in growth arrest and epithelial-to-mesenchymal transition. Dev. Cell 2015, 32, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Tang, H.; Lei, Z.; Zhu, J.; Zeng, Y.; Liu, Z.; Huang, J.A. miR-335-5p inhibits TGF-β1-induced epithelial-mesenchymal transition in non-small cell lung cancer via ROCK1. Respir. Res. 2019, 20, 225. [Google Scholar] [CrossRef]
- Lin, L.; Li, M.; Lin, L.; Xu, X.; Jiang, G.; Wu, L. FPPS mediates TGF-β1-induced non-small cell lung cancer cell invasion and the EMT process via the RhoA/Rock1 pathway. Biochem. Biophys. Res. Commun. 2018, 496, 536–541. [Google Scholar] [CrossRef]
- Cho, J.H.; Oh, A.Y.; Park, S.; Kang, S.M.; Yoon, M.H.; Woo, T.G.; Hong, S.D.; Hwang, J.; Ha, N.C.; Lee, H.Y.; et al. Loss of NF2 Induces TGFβ Receptor 1-mediated Noncanonical and Oncogenic TGFβ Signaling: Implication of the Therapeutic Effect of TGFβ Receptor 1 Inhibitor on NF2 Syndrome. Mol. Cancer Ther. 2018, 17, 2271–2284. [Google Scholar] [CrossRef]
- Dong, W.; Chen, A.; Chao, X.; Li, X.; Cui, Y.; Xu, C.; Cao, J.; Ning, Y. Chrysin Inhibits Proinflammatory Factor-Induced EMT Phenotype and Cancer Stem Cell-Like Features in HeLa Cells by Blocking the NF-κB/Twist Axis. Cell. Physiol. Biochem. 2019, 52, 1236–1250. [Google Scholar]
- Freudlsperger, C.; Bian, Y.; Contag Wise, S.; Burnett, J.; Coupar, J.; Yang, X.; Chen, Z.; Van Waes, C. TGF-β and NF-κB signal pathway cross-talk is mediated through TAK1 and SMAD7 in a subset of head and neck cancers. Oncogene 2013, 32, 1549–1559. [Google Scholar] [CrossRef]
- Lourenço, A.R.; Roukens, M.G.; Seinstra, D.; Frederiks, C.L.; Pals, C.E.; Vervoort, S.J.; Margarido, A.S.; van Rheenen, J.; Coffer, P.J. C/EBPα is crucial determinant of epithelial maintenance by preventing epithelial-to-mesenchymal transition. Nat. Commun. 2020, 11, 785. [Google Scholar] [CrossRef]
- David, C.J.; Huang, Y.H.; Chen, M.; Su, J.; Zou, Y.; Bardeesy, N.; Iacobuzio-Donahue, C.A.; Massagué, J. TGF-β Tumor Suppression through a Lethal EMT. Cell 2016, 164, 1015–1030. [Google Scholar] [CrossRef]
- Mohd Faheem, M.; Rasool, R.U.; Ahmad, S.M.; Jamwal, V.L.; Chakraborty, S.; Katoch, A.; Gandhi, S.G.; Bhagat, M.; Goswami, A. Par-4 mediated Smad4 induction in PDAC cells restores canonical TGF-β/ Smad4 axis driving the cells towards lethal EMT. Eur. J. Cell Biol. 2020, 99, 151076. [Google Scholar] [CrossRef]
- Yilmaz, M.; Maass, D.; Tiwari, N.; Waldmeier, L.; Schmidt, P.; Lehembre, F.; Christofori, G. Transcription factor Dlx2 protects from TGFβ-induced cell-cycle arrest and apoptosis. EMBO J. 2011, 30, 4489–4499. [Google Scholar] [CrossRef] [PubMed]
- Nüchel, J.; Ghatak, S.; Zuk, A.V.; Illerhaus, A.; Mörgelin, M.; Schönborn, K.; Blumbach, K.; Wickström, S.A.; Krieg, T.; Sengle, G.; et al. TGFB1 is secreted through an unconventional pathway dependent on the autophagic machinery and cytoskeletal regulators. Autophagy 2018, 14, 465–486. [Google Scholar] [CrossRef]
- Tran, D.Q.; Andersson, J.; Wang, R.; Ramsey, H.; Unutmaz, D.; Shevach, E.M. GARP (LRRC32) is essential for the surface expression of latent TGF-beta on platelets and activated FOXP3+ regulatory T cells. Proc. Natl. Acad. Sci. USA 2009, 106, 13445–13450. [Google Scholar] [CrossRef]
- Perng, D.W.; Chang, K.T.; Su, K.C.; Wu, Y.C.; Chen, C.S.; Hsu, W.H.; Tsai, C.M.; Lee, Y.C. Matrix metalloprotease-9 induces transforming growth factor-β(1) production in airway epithelium via activation of epidermal growth factor receptors. Life Sci. 2011, 89, 204–212. [Google Scholar] [CrossRef]
- Zhang, N.; Yang, K.; Bai, J.; Yi, J.; Gao, C.; Zhao, J.; Liang, S.; Wei, T.; Feng, L.; Song, L.; et al. Myeloid-specific blockade of Notch signaling alleviates murine pulmonary fibrosis through regulating monocyte-derived Ly6c(lo) MHCII(hi) alveolar macrophages recruitment and TGF-β secretion. FASEB J. 2020. [Google Scholar] [CrossRef]
- Li, Y.; Liu, W.; Guan, X.; Truscott, J.; Creemers, J.W.; Chen, H.L.; Pesu, M.; El Abiad, R.G.; Karacay, B.; Urban, J.F., Jr.; et al. STAT6 and Furin Are Successive Triggers for the Production of TGF-β by T Cells. J. Immunol. 2018, 201, 2612–2623. [Google Scholar] [CrossRef]
- Katoh, D.; Kozuka, Y.; Noro, A.; Ogawa, T.; Imanaka-Yoshida, K.; Yoshida, T. Tenascin-C Induces Phenotypic Changes in Fibroblasts to Myofibroblasts with High Contractility through the Integrin αvβ1/Transforming Growth Factor β/SMAD Signaling Axis in Human Breast Cancer. Am. J. Pathol. 2020, 190, 2123–2135. [Google Scholar] [CrossRef] [PubMed]
- Anastasi, C.; Rousselle, P.; Talantikite, M.; Tessier, A.; Cluzel, C.; Bachmann, A.; Mariano, N.; Dussoyer, M.; Alcaraz, L.B.; Fortin, L.; et al. BMP-1 disrupts cell adhesion and enhances TGF-β activation through cleavage of the matricellular protein thrombospondin-1. Sci. Signal. 2020, 13, eaba3880. [Google Scholar] [CrossRef]
- Stachowski, T.; Grant, T.D.; Snell, E.H. Structural consequences of transforming growth factor beta-1 activation from near-therapeutic X-ray doses. J. Synchrotron Radiat. 2019, 26, 967–979. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.G.; Cormier, A.; Ito, S.; Seed, R.I.; Bondesson, A.J.; Lou, J.; Marks, J.D.; Baron, J.L.; Cheng, Y.; Nishimura, S.L. Cryo-EM Reveals Integrin-Mediated TGF-β Activation without Release from Latent TGF-β. Cell 2020, 180, 490–501. [Google Scholar] [CrossRef]
- Mani, V.; Bromley, S.K.; Äijö, T.; Mora-Buch, R.; Carrizosa, E.; Warner, R.D.; Hamze, M.; Sen, D.R.; Chasse, A.Y.; Lorant, A.; et al. Migratory DCs activate TGF-β to precondition naïve CD8(+) T cells for tissue-resident memory fate. Science 2019, 366, eaav5728. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, J.; Beura, L.K.; Bobr, A.; Astry, B.; Chicoine, B.; Kashem, S.W.; Welty, N.E.; Igyártó, B.Z.; Wijeyesinghe, S.; Thompson, E.A.; et al. Stromal cells control the epithelial residence of DCs and memory T cells by regulated activation of TGF-β. Nat. Immunol. 2016, 17, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, S.; Elhance, A.; Van Duzer, A.; Kumar, S.; Leitenberger, J.J.; Oshimori, N. Tumor-initiating cells establish an IL-33-TGF-β niche signaling loop to promote cancer progression. Science 2020, 369, eaay1813. [Google Scholar]
- Takasaka, N.; Seed, R.I.; Cormier, A.; Bondesson, A.J.; Lou, J.; Elattma, A.; Ito, S.; Yanagisawa, H.; Hashimoto, M.; Ma, R.; et al. Integrin αvβ8-expressing tumor cells evade host immunity by regulating TGF-β activation in immune cells. JCI Insight 2018, 3, e122591. [Google Scholar] [CrossRef]
- Ritsma, L.; Dey-Guha, I.; Talele, N.; Sole, X.; Ross, K.N.; Salony; Chowdhury, J.; Ramaswamy, S. Integrin β1 activation induces an anti-melanoma host response. PLoS ONE 2017, 12, e0175300. [Google Scholar] [CrossRef]
- Minutti, C.M.; Modak, R.V.; Macdonald, F.; Li, F.; Smyth, D.J.; Dorward, D.A.; Blair, N.; Husovsky, C.; Muir, A.; Giampazolias, E.; et al. A Macrophage-Pericyte Axis Directs Tissue Restoration via Amphiregulin-Induced Transforming Growth Factor Beta Activation. Immunity 2019, 50, 645–654. [Google Scholar] [CrossRef]
- Lynch, K.; Treacy, O.; Chen, X.; Murphy, N.; Lohan, P.; Islam, M.N.; Donohoe, E.; Griffin, M.D.; Watson, L.; McLoughlin, S.; et al. TGF-β1-Licensed Murine MSCs Show Superior Therapeutic Efficacy in Modulating Corneal Allograft Immune Rejection In Vivo. Mol. Ther. 2020, 28, 2023–2043. [Google Scholar] [CrossRef]
- Benzoubir, N.; Lejamtel, C.; Battaglia, S.; Testoni, B.; Benassi, B.; Gondeau, C.; Perrin-Cocon, L.; Desterke, C.; Thiers, V.; Samuel, D.; et al. HCV core-mediated activation of latent TGF-β via thrombospondin drives the crosstalk between hepatocytes and stromal environment. J. Hepatol. 2013, 59, 1160–1168. [Google Scholar] [CrossRef]
- Calon, A.; Espinet, E.; Palomo-Ponce, S.; Tauriello, D.V.; Iglesias, M.; Céspedes, M.V.; Sevillano, M.; Nadal, C.; Jung, P.; Zhang, X.H.; et al. Dependency of colorectal cancer on a TGF-β-driven program in stromal cells for metastasis initiation. Cancer Cell 2012, 22, 571–584. [Google Scholar] [CrossRef]
- Tjin, G.; White, E.S.; Faiz, A.; Sicard, D.; Tschumperlin, D.J.; Mahar, A.; Kable, E.P.W.; Burgess, J.K. Lysyl oxidases regulate fibrillar collagen remodelling in idiopathic pulmonary fibrosis. Dis. Model. Mech. 2017, 10, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Dayer, C.; Stamenkovic, I. Recruitment of Matrix Metalloproteinase-9 (MMP-9) to the Fibroblast Cell Surface by Lysyl Hydroxylase 3 (LH3) Triggers Transforming Growth Factor-β (TGF-β) Activation and Fibroblast Differentiation. J. Biol. Chem. 2015, 290, 13763–13778. [Google Scholar] [CrossRef] [PubMed]
- Russo, I.; Cavalera, M.; Huang, S.; Su, Y.; Hanna, A.; Chen, B.; Shinde, A.V.; Conway, S.J.; Graff, J.; Frangogiannis, N.G. Protective Effects of Activated Myofibroblasts in the Pressure-Overloaded Myocardium Are Mediated Through Smad-Dependent Activation of a Matrix-Preserving Program. Circ. Res. 2019, 124, 1214–1227. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Q.; Mao, J.; Qin, T.; Sun, Y.; Yang, J.; Han, Y.; Li, L.; Li, Q. Salinomycin suppresses cancer cell stemness and attenuates TGF-β-induced epithelial-mesenchymal transition of renal cell carcinoma cells. Chem. Biol. Interact. 2018, 296, 145–153. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, D.; Li, J.; Yan, X.; Zhu, J.; Xiao, P.; Chen, T.; Xie, X. Effects of Bone Marrow-Derived Mesenchymal Stem Cells on Hypoxia and the Transforming Growth Factor beta 1 (TGFβ-1) and SMADs Pathway in a Mouse Model of Cirrhosis. Med. Sci. Monit. 2019, 25, 7182–7190. [Google Scholar] [CrossRef]
- Setiawan, M.; Tan, X.W.; Goh, T.W.; Hin-Fai Yam, G.; Mehta, J.S. Inhibiting glycogen synthase kinase-3 and transforming growth factor-β signaling to promote epithelial transition of human adipose mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2017, 490, 1381–1388. [Google Scholar] [CrossRef]
- Bagordakis, E.; Sawazaki-Calone, I.; Macedo, C.C.; Carnielli, C.M.; de Oliveira, C.E.; Rodrigues, P.C.; Rangel, A.L.; Dos Santos, J.N.; Risteli, J.; Graner, E.; et al. Secretome profiling of oral squamous cell carcinoma-associated fibroblasts reveals organization and disassembly of extracellular matrix and collagen metabolic process signatures. Tumour Biol. 2016, 37, 9045–9057. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Tang, H.; Lin, H.; Mao, J.; Gao, L.; Liu, J.; Wu, T. Rho/Rock cross-talks with transforming growth factor-β/Smad pathway participates in lung fibroblast-myofibroblast differentiation. Biomed. Rep. 2014, 2, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, O.; Yoshida, M.; Koma, Y.; Yanai, T.; Hasegawa, D.; Kosaka, Y.; Nishimura, N.; Yokozaki, H. Collaboration of cancer-associated fibroblasts and tumour-associated macrophages for neuroblastoma development. J. Pathol. 2016, 240, 211–223. [Google Scholar] [CrossRef]
- Cheng, J.T.; Deng, Y.N.; Yi, H.M.; Wang, G.Y.; Fu, B.S.; Chen, W.J.; Liu, W.; Tai, Y.; Peng, Y.W.; Zhang, Q. Hepatic carcinoma-associated fibroblasts induce IDO-producing regulatory dendritic cells through IL-6-mediated STAT3 activation. Oncogenesis 2016, 5, e198. [Google Scholar] [CrossRef]
- Chakravarthy, A.; Khan, L.; Bensler, N.P.; Bose, P.; De Carvalho, D.D. TGF-β-associated extracellular matrix genes link cancer-associated fibroblasts to immune evasion and immunotherapy failure. Nat. Commun. 2018, 9, 4692. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Chen, Y.; Chen, H.; Jiang, P.; Yan, L.; Mo, D.; Tang, X.; Yan, F. DCST1-AS1 Promotes TGF-β-Induced Epithelial-Mesenchymal Transition and Enhances Chemoresistance in Triple-Negative Breast Cancer Cells via ANXA1. Front. Oncol. 2020, 10, 280. [Google Scholar] [CrossRef] [PubMed]
- Bhagyaraj, E.; Ahuja, N.; Kumar, S.; Tiwari, D.; Gupta, S.; Nanduri, R.; Gupta, P. TGF-β induced chemoresistance in liver cancer is modulated by xenobiotic nuclear receptor PXR. Cell Cycle 2019, 18, 3589–3602. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; He, S.; Gao, A.; Zhang, Y.; Zhu, Q.; Wang, P.; Yang, B.; Yin, H.; Li, Y.; Song, J.; et al. Methylation silencing of TGF-β receptor type II is involved in malignant transformation of esophageal squamous cell carcinoma. Clin. Epigenetics 2020, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Palomeras, S.; Diaz-Lagares, Á.; Viñas, G.; Setien, F.; Ferreira, H.J.; Oliveras, G.; Crujeiras, A.B.; Hernández, A.; Lum, D.H.; Welm, A.L.; et al. Epigenetic silencing of TGFBI confers resistance to trastuzumab in human breast cancer. Breast Cancer Res. 2019, 21, 79. [Google Scholar] [CrossRef] [PubMed]
- You, X.; Zhou, Z.; Chen, W.; Wei, X.; Zhou, H.; Luo, W. MicroRNA-495 confers inhibitory effects on cancer stem cells in oral squamous cell carcinoma through the HOXC6-mediated TGF-β signaling pathway. Stem Cell Res. Ther. 2020, 11, 117. [Google Scholar] [CrossRef]
- Choi, P.-W.; So, W.W.; Yang, J.; Liu, S.; Tong, K.K.; Kwan, K.M.; Kwok, J.S.L.; Tsui, S.K.W.; Ng, S.-K.; Hales, K.H.; et al. MicroRNA-200 family governs ovarian inclusion cyst formation and mode of ovarian cancer spread. Oncogene 2020, 39, 4045–4060. [Google Scholar] [CrossRef]
- Zhang, L.; Song, X.; Chen, X.; Wang, Q.; Zheng, X.; Wu, C.; Jiang, J. Circular RNA CircCACTIN Promotes Gastric Cancer Progression by Sponging MiR-331-3p and Regulating TGFBR1 Expression. Int. J. Biol. Sci. 2019, 15, 1091–1103. [Google Scholar] [CrossRef]
- Xu, G.; Chen, Y.; Fu, M.; Zang, X.; Cang, M.; Niu, Y.; Zhang, W.; Zhang, Y.; Mao, Z.; Shao, M.; et al. Circular RNA CCDC66 promotes gastric cancer progression by regulating c-Myc and TGF-β signaling pathways. J. Cancer 2020, 11, 2759–2768. [Google Scholar] [CrossRef]
- Huang, W.; Yang, Y.; Wu, J.; Niu, Y.; Yao, Y.; Zhang, J.; Huang, X.; Liang, S.; Chen, R.; Chen, S.; et al. Circular RNA cESRP1 sensitises small cell lung cancer cells to chemotherapy by sponging miR-93-5p to inhibit TGF-β signalling. Cell Death Differ. 2020, 27, 1709–1727. [Google Scholar] [CrossRef]
- Ohtani, H.; Terashima, T.; Sato, E. Immune cell expression of TGFβ1 in cancer with lymphoid stroma: Dendritic cell and regulatory T cell contact. Virchows Arch. 2018, 472, 1021–1028. [Google Scholar] [CrossRef]
- Tobin, S.W.; Douville, K.; Benbow, U.; Brinckerhoff, C.E.; Memoli, V.A.; Arrick, B.A. Consequences of altered TGF-β expression and responsiveness in breast cancer: Evidence for autocrine and paracrine effects. Oncogene 2002, 21, 108–118. [Google Scholar] [CrossRef]
- Gutcher, I.; Donkor, M.K.; Ma, Q.; Rudensky, A.Y.; Flavell, R.A.; Li, M.O. Autocrine transforming growth factor-β1 promotes in vivo Th17 cell differentiation. Immunity 2011, 34, 396–408. [Google Scholar] [CrossRef]
- Liénart, S.; Merceron, R.; Vanderaa, C.; Lambert, F.; Colau, D.; Stockis, J.; van der Woning, B.; De Haard, H.; Saunders, M.; Coulie, P.G.; et al. Structural basis of latent TGF-β1 presentation and activation by GARP on human regulatory T cells. Science 2018, 362, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.; Subudhi, S.K.; Aparicio, A.; Ge, Z.; Guan, B.; Miura, Y.; Sharma, P. Differences in Tumor Microenvironment Dictate T Helper Lineage Polarization and Response to Immune Checkpoint Therapy. Cell 2019, 179, 1177–1190. [Google Scholar] [CrossRef]
- Terabe, M.; Robertson, F.C.; Clark, K.; De Ravin, E.; Bloom, A.; Venzon, D.J.; Kato, S.; Mirza, A.; Berzofsky, J.A. Blockade of only TGF-β 1 and 2 is sufficient to enhance the efficacy of vaccine and PD-1 checkpoint blockade immunotherapy. Oncoimmunology 2017, 6, e1308616. [Google Scholar] [CrossRef]
- Xue, G.; Jin, G.; Fang, J.; Lu, Y. IL-4 together with IL-1β induces antitumor Th9 cell differentiation in the absence of TGF-β signaling. Nat. Commun. 2019, 10, 1376. [Google Scholar] [CrossRef]
- Perez, L.G.; Kempski, J.; McGee, H.M.; Pelzcar, P.; Agalioti, T.; Giannou, A.; Konczalla, L.; Brockmann, L.; Wahib, R.; Xu, H.; et al. TGF-β signaling in Th17 cells promotes IL-22 production and colitis-associated colon cancer. Nat. Commun. 2020, 11, 2608. [Google Scholar] [CrossRef]
- Shevach, E.M.; Thornton, A.M. tTregs, pTregs, and iTregs: Similarities and differences. Immunol. Rev. 2014, 259, 88–102. [Google Scholar] [CrossRef]
- Courau, T.; Nehar-Belaid, D.; Florez, L.; Levacher, B.; Vazquez, T.; Brimaud, F.; Bellier, B.; Klatzmann, D. TGF-β and VEGF cooperatively control the immunotolerant tumor environment and the efficacy of cancer immunotherapies. JCI Insight 2016, 1, e85974. [Google Scholar] [CrossRef]
- Liu, R.; Lu, Z.; Gu, J.; Liu, J.; Huang, E.; Liu, X.; Wang, L.; Yang, J.; Deng, Y.; Qian, J.; et al. MicroRNAs 15A and 16-1 Activate Signaling Pathways That Mediate Chemotaxis of Immune Regulatory B cells to Colorectal Tumors. Gastroenterology 2018, 154, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Morgan, R.; Chen, C.; Cai, Y.; Clark, E.; Khan, W.N.; Shin, S.U.; Cho, H.M.; Al Bayati, A.; Pimentel, A.; et al. Mammary-tumor-educated B cells acquire LAP/TGF-β and PD-L1 expression and suppress anti-tumor immune responses. Int. Immunol. 2016, 28, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Pasero, C.; Gravis, G.; Guerin, M.; Granjeaud, S.; Thomassin-Piana, J.; Rocchi, P.; Paciencia-Gros, M.; Poizat, F.; Bentobji, M.; Azario-Cheillan, F.; et al. Inherent and Tumor-Driven Immune Tolerance in the Prostate Microenvironment Impairs Natural Killer Cell Antitumor Activity. Cancer Res. 2016, 76, 2153–2165. [Google Scholar] [CrossRef] [PubMed]
- Viel, S.; Marçais, A.; Guimaraes, F.S.; Loftus, R.; Rabilloud, J.; Grau, M.; Degouve, S.; Djebali, S.; Sanlaville, A.; Charrier, E.; et al. TGF-β inhibits the activation and functions of NK cells by repressing the mTOR pathway. Sci. Signal. 2016, 9, ra19. [Google Scholar] [CrossRef]
- Tang, P.M.; Zhou, S.; Meng, X.M.; Wang, Q.M.; Li, C.J.; Lian, G.Y.; Huang, X.R.; Tang, Y.J.; Guan, X.Y.; Yan, B.P.; et al. Smad3 promotes cancer progression by inhibiting E4BP4-mediated NK cell development. Nat. Commun. 2017, 8, 14677. [Google Scholar] [CrossRef]
- Li, J.; Wang, L.; Chen, X.; Li, L.; Li, Y.; Ping, Y.; Huang, L.; Yue, D.; Zhang, Z.; Wang, F.; et al. CD39/CD73 upregulation on myeloid-derived suppressor cells via TGF-β-mTOR-HIF-1 signaling in patients with non-small cell lung cancer. Oncoimmunology 2017, 6, e1320011. [Google Scholar] [CrossRef]
- Wang, Y.; Schafer, C.C.; Hough, K.P.; Tousif, S.; Duncan, S.R.; Kearney, J.F.; Ponnazhagan, S.; Hsu, H.C.; Deshane, J.S. Myeloid-Derived Suppressor Cells Impair B Cell Responses in Lung Cancer through IL-7 and STAT5. J. Immunol. 2018, 201, 278–295. [Google Scholar] [CrossRef]
- Bodogai, M.; Moritoh, K.; Lee-Chang, C.; Hollander, C.M.; Sherman-Baust, C.A.; Wersto, R.P.; Araki, Y.; Miyoshi, I.; Yang, L.; Trinchieri, G.; et al. Immunosuppressive and Prometastatic Functions of Myeloid-Derived Suppressive Cells Rely upon Education from Tumor-Associated B Cells. Cancer Res. 2015, 75, 3456–3465. [Google Scholar] [CrossRef]
- Jayaraman, P.; Parikh, F.; Newton, J.M.; Hanoteau, A.; Rivas, C.; Krupar, R.; Rajapakshe, K.; Pathak, R.; Kanthaswamy, K.; MacLaren, C.; et al. TGF-β1 programmed myeloid-derived suppressor cells (MDSC) acquire immune-stimulating and tumor killing activity capable of rejecting established tumors in combination with radiotherapy. Oncoimmunology 2018, 7, e1490853. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, H.; Wang, X.; Jiang, G.; Liu, H.; Zhang, G.; Wang, H.; Fang, R.; Bu, X.; Cai, S.; et al. TGF-β induces M2-like macrophage polarization via SNAIL-mediated suppression of a pro-inflammatory phenotype. Oncotarget 2016, 7, 52294–52306. [Google Scholar] [CrossRef]
- Fu, X.L.; Duan, W.; Su, C.Y.; Mao, F.Y.; Lv, Y.P.; Teng, Y.S.; Yu, P.W.; Zhuang, Y.; Zhao, Y.L. Interleukin 6 induces M2 macrophage differentiation by STAT3 activation that correlates with gastric cancer progression. Cancer Immunol. Immunother. 2017, 66, 1597–1608. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Shen, H.; Zhu, L.; Zhao, F.; Shu, Y. Plasminogen Activator Inhibitor 1 Promotes Immunosuppression in Human Non-Small Cell Lung Cancers by Enhancing TGF-Β1 Expression in Macrophage. Cell. Physiol. Biochem. 2017, 44, 2201–2211. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Sun, X.; Pan, B.; Cao, S.; Cao, J.; Che, D.; Liu, F.; Zhang, S.; Yu, Y. IL-17 induces macrophages to M2-like phenotype via NF-κB. Cancer Manag. Res. 2018, 10, 4217–4228. [Google Scholar] [CrossRef] [PubMed]
- Hanks, B.A.; Holtzhausen, A.; Evans, K.S.; Jamieson, R.; Gimpel, P.; Campbell, O.M.; Hector-Greene, M.; Sun, L.; Tewari, A.; George, A.; et al. Type III TGF-β receptor downregulation generates an immunotolerant tumor microenvironment. J. Clin. Investig. 2013, 123, 3925–3940. [Google Scholar] [CrossRef] [PubMed]
- Ni, X.Y.; Sui, H.X.; Liu, Y.; Ke, S.Z.; Wang, Y.N.; Gao, F.G. TGF-β of lung cancer microenvironment upregulates B7H1 and GITRL expression in dendritic cells and is associated with regulatory T cell generation. Oncol. Rep. 2012, 28, 615–621. [Google Scholar] [CrossRef]
- Zhong, M.; Zhong, C.; Cui, W.; Wang, G.; Zheng, G.; Li, L.; Zhang, J.; Ren, R.; Gao, H.; Wang, T.; et al. Induction of tolerogenic dendritic cells by activated TGF-β/Akt/Smad2 signaling in RIG-I-deficient stemness-high human liver cancer cells. BMC Cancer 2019, 19, 439. [Google Scholar] [CrossRef]
- Haider, C.; Hnat, J.; Wagner, R.; Huber, H.; Timelthaler, G.; Grubinger, M.; Coulouarn, C.; Schreiner, W.; Schlangen, K.; Sieghart, W.; et al. Transforming Growth Factor-β and Axl Induce CXCL5 and Neutrophil Recruitment in Hepatocellular Carcinoma. Hepatology 2019, 69, 222–236. [Google Scholar] [CrossRef]
- Yan, C.; Huo, X.; Wang, S.; Feng, Y.; Gong, Z. Stimulation of hepatocarcinogenesis by neutrophils upon induction of oncogenic kras expression in transgenic zebrafish. J. Hepatol. 2015, 63, 420–428. [Google Scholar] [CrossRef]
- Jackstadt, R.; van Hooff, S.R.; Leach, J.D.; Cortes-Lavaud, X.; Lohuis, J.O.; Ridgway, R.A.; Wouters, V.M.; Roper, J.; Kendall, T.J.; Roxburgh, C.S.; et al. Epithelial NOTCH Signaling Rewires the Tumor Microenvironment of Colorectal Cancer to Drive Poor-Prognosis Subtypes and Metastasis. Cancer Cell 2019, 36, 319–336. [Google Scholar] [CrossRef]
- Germann, M.; Zangger, N.; Sauvain, M.O.; Sempoux, C.; Bowler, A.D.; Wirapati, P.; Kandalaft, L.E.; Delorenzi, M.; Tejpar, S.; Coukos, G.; et al. Neutrophils suppress tumor-infiltrating T cells in colon cancer via matrix metalloproteinase-mediated activation of TGFβ. EMBO Mol. Med. 2020, 12, e10681. [Google Scholar] [CrossRef]
- McKarns, S.C.; Schwartz, R.H. Distinct effects of TGF-beta 1 on CD4+ and CD8+ T cell survival, division, and IL-2 production: A role for T cell intrinsic Smad3. J. Immunol. 2005, 174, 2071–2083. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Yue, W.; Le-Le, Z.; Bin, L.; Hu, X. Mesenchymal stem cells inhibit T cell activation by releasing TGF-β1 from TGF-β1/GARP complex. Oncotarget 2017, 8, 99784–99800. [Google Scholar] [CrossRef]
- Pang, N.; Zhang, F.; Ma, X.; Zhu, Y.; Zhao, H.; Xin, Y.; Wang, S.; Chen, Z.; Wen, H.; Ding, J. TGF-β/Smad signaling pathway regulates Th17/Treg balance during Echinococcus multilocularis infection. Int. Immunopharmacol. 2014, 20, 248–257. [Google Scholar] [CrossRef]
- Gagliani, N.; Amezcua Vesely, M.C.; Iseppon, A.; Brockmann, L.; Xu, H.; Palm, N.W.; de Zoete, M.R.; Licona-Limón, P.; Paiva, R.S.; Ching, T.; et al. Th17 cells transdifferentiate into regulatory T cells during resolution of inflammation. Nature 2015, 523, 221–225. [Google Scholar] [CrossRef]
- Xu, L.; Kitani, A.; Fuss, I.; Strober, W. Cutting edge: Regulatory T cells induce CD4+CD25-Foxp3- T cells or are self-induced to become Th17 cells in the absence of exogenous TGF-beta. J. Immunol. 2007, 178, 6725–6729. [Google Scholar] [CrossRef]
- Chen, M.L.; Pittet, M.J.; Gorelik, L.; Flavell, R.A.; Weissleder, R.; von Boehmer, H.; Khazaie, K. Regulatory T cells suppress tumor-specific CD8 T cell cytotoxicity through TGF-beta signals in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Kong, N.; Lan, Q.; Chen, M.; Wang, J.; Shi, W.; Horwitz, D.A.; Quesniaux, V.; Ryffel, B.; Liu, Z.; Brand, D.; et al. Antigen-specific transforming growth factor β-induced Treg cells, but not natural Treg cells, ameliorate autoimmune arthritis in mice by shifting the Th17/Treg cell balance from Th17 predominance to Treg cell predominance. Arthritis Rheum. 2012, 64, 2548–2558. [Google Scholar] [CrossRef] [PubMed]
- Sehrawat, S.; Suvas, S.; Sarangi, P.P.; Suryawanshi, A.; Rouse, B.T. In vitro-generated antigen-specific CD4+ CD25+ Foxp3+ regulatory T cells control the severity of herpes simplex virus-induced ocular immunoinflammatory lesions. J. Virol. 2008, 82, 6838–6851. [Google Scholar] [CrossRef]
- Schreiber, T.H.; Wolf, D.; Bodero, M.; Podack, E. Tumor antigen specific iTreg accumulate in the tumor microenvironment and suppress therapeutic vaccination. Oncoimmunology 2012, 1, 642–648. [Google Scholar] [CrossRef]
- Hegde, U.P.; Chakraborty, N.G. Peripherally induced Tregs or pTregs are the potent tolerance inducer for the growth and metastasis of cancer. Cancer Res. 2018, 78. [Google Scholar] [CrossRef]
- Tang, N.; Cheng, C.; Zhang, X.; Qiao, M.; Li, N.; Mu, W.; Wei, X.F.; Han, W.; Wang, H. TGF-β inhibition via CRISPR promotes the long-term efficacy of CAR T cells against solid tumors. JCI Insight 2020, 5, e133977. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Hong, J.; Yun, C.O. Regulatory T Cells Induce Metastasis by Activating Tgf-Β and Enhancing the Epithelial-Mesenchymal Transition. Cells 2019, 8, 1387. [Google Scholar] [CrossRef] [PubMed]
- Tamayo, E.; Alvarez, P.; Merino, R. TGFβ Superfamily Members as Regulators of B Cell Development and Function-Implications for Autoimmunity. Int. J. Mol. Sci. 2018, 19, 3928. [Google Scholar] [CrossRef] [PubMed]
- Reboldi, A.; Arnon, T.I.; Rodda, L.B.; Atakilit, A.; Sheppard, D.; Cyster, J.G. IgA production requires B cell interaction with subepithelial dendritic cells in Peyer’s patches. Science 2016, 352, aaf4822. [Google Scholar] [CrossRef] [PubMed]
- Caron, G.; Hussein, M.; Kulis, M.; Delaloy, C.; Chatonnet, F.; Pignarre, A.; Avner, S.; Lemarié, M.; Mahé, E.A.; Verdaguer-Dot, N.; et al. Cell-Cycle-Dependent Reconfiguration of the DNA Methylome during Terminal Differentiation of Human B Cells into Plasma Cells. Cell Rep. 2015, 13, 1059–1071. [Google Scholar] [CrossRef]
- Tang, J.; Nuccie, B.L.; Ritterman, I.; Liesveld, J.L.; Abboud, C.N.; Ryan, D.H. TGF-beta down-regulates stromal IL-7 secretion and inhibits proliferation of human B cell precursors. J. Immunol. 1997, 159, 117–125. [Google Scholar] [PubMed]
- Lømo, J.; Blomhoff, H.K.; Beiske, K.; Stokke, T.; Smeland, E.B. TGF-beta 1 and cyclic AMP promote apoptosis in resting human B lymphocytes. J. Immunol. 1995, 154, 1634–1643. [Google Scholar] [PubMed]
- Mamessier, E.; Sylvain, A.; Thibult, M.L.; Houvenaeghel, G.; Jacquemier, J.; Castellano, R.; Gonçalves, A.; André, P.; Romagné, F.; Thibault, G.; et al. Human breast cancer cells enhance self tolerance by promoting evasion from NK cell antitumor immunity. J. Clin. Investig. 2011, 121, 3609–3622. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Huang, Q.; Huang, M.; Wen, H.; Lin, R.; Zheng, M.; Qu, K.; Li, K.; Wei, H.; Xiao, W.; et al. Human CD96 Correlates to Natural Killer Cell Exhaustion and Predicts the Prognosis of Human Hepatocellular Carcinoma. Hepatology 2019, 70, 168–183. [Google Scholar] [CrossRef]
- Gao, Y.; Souza-Fonseca-Guimaraes, F.; Bald, T.; Ng, S.S.; Young, A.; Ngiow, S.F.; Rautela, J.; Straube, J.; Waddell, N.; Blake, S.J.; et al. Tumor immunoevasion by the conversion of effector NK cells into type 1 innate lymphoid cells. Nat. Immunol. 2017, 18, 1004–1015. [Google Scholar] [CrossRef]
- Cortez, V.S.; Ulland, T.K.; Cervantes-Barragan, L.; Bando, J.K.; Robinette, M.L.; Wang, Q.; White, A.J.; Gilfillan, S.; Cella, M.; Colonna, M. SMAD4 impedes the conversion of NK cells into ILC1-like cells by curtailing non-canonical TGF-β signaling. Nat. Immunol. 2017, 18, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Mao, F.Y.; Zhao, Y.L.; Lv, Y.P.; Teng, Y.S.; Duan, M.; Chen, W.; Cheng, P.; Wang, T.T.; Liang, Z.Y.; et al. Altered NKp30, NKp46, NKG2D, and DNAM-1 Expression on Circulating NK Cells Is Associated with Tumor Progression in Human Gastric Cancer. J. Immunol. Res. 2018, 2018, 6248590. [Google Scholar] [CrossRef]
- Shvedova, A.A.; Kisin, E.R.; Yanamala, N.; Tkach, A.V.; Gutkin, D.W.; Star, A.; Shurin, G.V.; Kagan, V.E.; Shurin, M.R. MDSC and TGFβ Are Required for Facilitation of Tumor Growth in the Lungs of Mice Exposed to Carbon Nanotubes. Cancer Res. 2015, 75, 1615–1623. [Google Scholar] [CrossRef]
- Faz-López, B.; Mayoral-Reyes, H.; Hernández-Pando, R.; Martínez-Labat, P.; McKay, D.M.; Medina-Andrade, I.; Olguín, J.E.; Terrazas, L.I. A Dual Role for Macrophages in Modulating Lung Tissue Damage/Repair during L2 Toxocara canis Infection. Pathogens 2019, 8, 280. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Deng, Y.R.; Wang, Z.C.; Wei, W.F.; Zhou, C.F.; Zhang, Y.M.; Yan, R.M.; Liang, L.J.; Zhong, M.; Liang, L.; et al. Hypoxia-induced ZEB1 promotes cervical cancer progression via CCL8-dependent tumour-associated macrophage recruitment. Cell Death Dis. 2019, 10, 508. [Google Scholar] [CrossRef]
- Miyake, M.; Hori, S.; Morizawa, Y.; Tatsumi, Y.; Nakai, Y.; Anai, S.; Torimoto, K.; Aoki, K.; Tanaka, N.; Shimada, K.; et al. CXCL1-Mediated Interaction of Cancer Cells with Tumor-Associated Macrophages and Cancer-Associated Fibroblasts Promotes Tumor Progression in Human Bladder Cancer. Neoplasia 2016, 18, 636–646. [Google Scholar] [CrossRef] [PubMed]
- Guerin, M.V.; Regnier, F.; Feuillet, V.; Vimeux, L.; Weiss, J.M.; Bismuth, G.; Altan-Bonnet, G.; Guilbert, T.; Thoreau, M.; Finisguerra, V.; et al. TGFβ blocks IFNα/β release and tumor rejection in spontaneous mammary tumors. Nat. Commun. 2019, 10, 4131. [Google Scholar] [CrossRef]
- Guo, X.; Xue, H.; Shao, Q.; Wang, J.; Guo, X.; Chen, X.; Zhang, J.; Xu, S.; Li, T.; Zhang, P.; et al. Hypoxia promotes glioma-associated macrophage infiltration via periostin and subsequent M2 polarization by upregulating TGF-beta and M-CSFR. Oncotarget 2016, 7, 80521–80542. [Google Scholar] [CrossRef]
- Wang, D.; Yang, L.; Yue, D.; Cao, L.; Li, L.; Wang, D.; Ping, Y.; Shen, Z.; Zheng, Y.; Wang, L.; et al. Macrophage-derived CCL22 promotes an immunosuppressive tumor microenvironment via IL-8 in malignant pleural effusion. Cancer Lett. 2019, 452, 244–253. [Google Scholar] [CrossRef]
- Yin, Z.; Ma, T.; Huang, B.; Lin, L.; Zhou, Y.; Yan, J.; Zou, Y.; Chen, S. Macrophage-derived exosomal microRNA-501-3p promotes progression of pancreatic ductal adenocarcinoma through the TGFBR3-mediated TGF-β signaling pathway. J. Exp. Clin. Cancer Res. 2019, 38, 310. [Google Scholar] [CrossRef]
- Labidi-Galy, S.I.; Sisirak, V.; Meeus, P.; Gobert, M.; Treilleux, I.; Bajard, A.; Combes, J.D.; Faget, J.; Mithieux, F.; Cassignol, A.; et al. Quantitative and functional alterations of plasmacytoid dendritic cells contribute to immune tolerance in ovarian cancer. Cancer Res. 2011, 71, 5423–5434. [Google Scholar] [CrossRef] [PubMed]
- Thepmalee, C.; Panya, A.; Sujjitjoon, J.; Sawasdee, N.; Poungvarin, N.; Junking, M.; Yenchitsomanus, P.T. Suppression of TGF-β and IL-10 receptors on self-differentiated dendritic cells by short-hairpin RNAs enhanced activation of effector T-cells against cholangiocarcinoma cells. Hum. Vaccin. Immunother. 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Barilla, R.M.; Diskin, B.; Caso, R.C.; Lee, K.B.; Mohan, N.; Buttar, C.; Adam, S.; Sekendiz, Z.; Wang, J.; Salas, R.D.; et al. Specialized dendritic cells induce tumor-promoting IL-10(+)IL-17(+) FoxP3(neg) regulatory CD4(+) T cells in pancreatic carcinoma. Nat. Commun. 2019, 10, 1424. [Google Scholar] [CrossRef] [PubMed]
- Sarhan, D.; Palma, M.; Mao, Y.; Adamson, L.; Kiessling, R.; Mellstedt, H.; Österborg, A.; Lundqvist, A. Dendritic cell regulation of NK-cell responses involves lymphotoxin-α, IL-12, and TGF-β. Eur. J. Immunol. 2015, 45, 1783–1793. [Google Scholar] [CrossRef]
- Lu, Z.; Zuo, B.; Jing, R.; Gao, X.; Rao, Q.; Liu, Z.; Qi, H.; Guo, H.; Yin, H. Dendritic cell-derived exosomes elicit tumor regression in autochthonous hepatocellular carcinoma mouse models. J. Hepatol. 2017, 67, 739–748. [Google Scholar] [CrossRef]
- Gulubova, M.; Aleksandrova, E.; Vlaykova, T. Promoter polymorphisms in TGFB1 and IL10 genes influence tumor dendritic cells infiltration, development and prognosis of colorectal cancer. J. Gene Med. 2018, 20, e3005. [Google Scholar] [CrossRef]
- Ethier, J.L.; Desautels, D.; Templeton, A.; Shah, P.S.; Amir, E. Prognostic role of neutrophil-to-lymphocyte ratio in breast cancer: A systematic review and meta-analysis. Breast Cancer Res. 2017, 19, 2. [Google Scholar] [CrossRef]
- Labelle, M.; Begum, S.; Hynes, R.O. Platelets guide the formation of early metastatic niches. Proc. Natl. Acad. Sci. USA 2014, 111, E3053–E3061. [Google Scholar] [CrossRef]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of tumor-associated neutrophil phenotype by TGF-beta: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef]
- Sagiv, J.Y.; Michaeli, J.; Assi, S.; Mishalian, I.; Kisos, H.; Levy, L.; Damti, P.; Lumbroso, D.; Polyansky, L.; Sionov, R.V.; et al. Phenotypic diversity and plasticity in circulating neutrophil subpopulations in cancer. Cell Rep. 2015, 10, 562–573. [Google Scholar] [CrossRef]
- Szczerba, B.M.; Castro-Giner, F.; Vetter, M.; Krol, I.; Gkountela, S.; Landin, J.; Scheidmann, M.C.; Donato, C.; Scherrer, R.; Singer, J.; et al. Neutrophils escort circulating tumour cells to enable cell cycle progression. Nature 2019, 566, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Gentles, A.J.; Newman, A.M.; Liu, C.L.; Bratman, S.V.; Feng, W.; Kim, D.; Nair, V.S.; Xu, Y.; Khuong, A.; Hoang, C.D.; et al. The prognostic landscape of genes and infiltrating immune cells across human cancers. Nat. Med. 2015, 21, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.L.; Yin, D.; Hu, Z.Q.; Luo, C.B.; Zhou, Z.J.; Xin, H.Y.; Yang, X.R.; Shi, Y.H.; Wang, Z.; Huang, X.W.; et al. A Positive Feedback Loop Between Cancer Stem-Like Cells and Tumor-Associated Neutrophils Controls Hepatocellular Carcinoma Progression. Hepatology 2019, 70, 1214–1230. [Google Scholar] [CrossRef]
- Tsutsumi, S.; Saeki, H.; Nakashima, Y.; Ito, S.; Oki, E.; Morita, M.; Oda, Y.; Okano, S.; Maehara, Y. Programmed death-ligand 1 expression at tumor invasive front is associated with epithelial-mesenchymal transition and poor prognosis in esophageal squamous cell carcinoma. Cancer Sci. 2017, 108, 1119–1127. [Google Scholar] [CrossRef]
- Martin, C.J.; Datta, A.; Littlefield, C.; Kalra, A.; Chapron, C.; Wawersik, S.; Dagbay, K.B.; Brueckner, C.T.; Nikiforov, A.; Danehy, F.T.; et al. Selective inhibition of TGFβ1 activation overcomes primary resistance to checkpoint blockade therapy by altering tumor immune landscape. Sci. Transl. Med. 2020, 12, eaay8456. [Google Scholar] [CrossRef]
- Panagi, M.; Voutouri, C.; Mpekris, F.; Papageorgis, P.; Martin, M.R.; Martin, J.D.; Demetriou, P.; Pierides, C.; Polydorou, C.; Stylianou, A.; et al. TGF-β inhibition combined with cytotoxic nanomedicine normalizes triple negative breast cancer microenvironment towards anti-tumor immunity. Theranostics 2020, 10, 1910–1922. [Google Scholar] [CrossRef]
- Bialkowski, L.; Van der Jeught, K.; Bevers, S.; Tjok Joe, P.; Renmans, D.; Heirman, C.; Aerts, J.L.; Thielemans, K. Immune checkpoint blockade combined with IL-6 and TGF-β inhibition improves the therapeutic outcome of mRNA-based immunotherapy. Int. J. Cancer 2018, 143, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.H.; Park, J.H.; Yang, A.; Park, H.J.; Lee, S.E.; Kim, Y.S.; Jang, G.-Y.; Farmer, E.; Lam, B.; Park, Y.-M.; et al. Annexin A5 as an immune checkpoint inhibitor and tumor-homing molecule for cancer treatment. Nat. Commun. 2020, 11, 1137. [Google Scholar] [CrossRef]
- Lan, Y.; Zhang, D.; Xu, C.; Hance, K.W.; Marelli, B.; Qi, J.; Yu, H.; Qin, G.; Sircar, A.; Hernández, V.M.; et al. Enhanced preclinical antitumor activity of M7824, a bifunctional fusion protein simultaneously targeting PD-L1 and TGF-β. Sci. Transl. Med. 2018, 10, eaan5488. [Google Scholar] [CrossRef]
- Cho, B.C.; Daste, A.; Ravaud, A.; Salas, S.; Isambert, N.; McClay, E.; Awada, A.; Borel, C.; Ojalvo, L.S.; Helwig, C.; et al. Bintrafusp alfa, a bifunctional fusion protein targeting TGF-β and PD-L1, in advanced squamous cell carcinoma of the head and neck: Results from a phase I cohort. J. Immunother. Cancer 2020, 8, e000664. [Google Scholar] [CrossRef]
- Horn, L.A.; Riskin, J.; Hempel, H.A.; Fousek, K.; Lind, H.; Hamilton, D.H.; McCampbell, K.K.; Maeda, D.Y.; Zebala, J.A.; Su, Z.; et al. Simultaneous inhibition of CXCR1/2, TGF-β, and PD-L1 remodels the tumor and its microenvironment to drive antitumor immunity. J. Immunother. Cancer 2020, 8, e000326. [Google Scholar] [CrossRef]
- Ravi, R.; Noonan, K.A.; Pham, V.; Bedi, R.; Zhavoronkov, A.; Ozerov, I.V.; Makarev, E.; Artemov, A.V.; Wysocki, P.T.; Mehra, R.; et al. Bifunctional immune checkpoint-targeted antibody-ligand traps that simultaneously disable TGFβ enhance the efficacy of cancer immunotherapy. Nat. Commun. 2018, 9, 741. [Google Scholar] [CrossRef] [PubMed]
- Strauss, J.; Heery, C.R.; Schlom, J.; Madan, R.A.; Cao, L.; Kang, Z.; Lamping, E.; Marté, J.L.; Donahue, R.N.; Grenga, I.; et al. Phase I Trial of M7824 (MSB0011359C), a Bifunctional Fusion Protein Targeting PD-L1 and TGFβ, in Advanced Solid Tumors. Clin. Cancer Res. 2018, 24, 1287–1295. [Google Scholar] [CrossRef] [PubMed]
- Lind, H.; Gameiro, S.R.; Jochems, C.; Donahue, R.N.; Strauss, J.; Gulley, J.M.; Palena, C.; Schlom, J. Dual targeting of TGF-β and PD-L1 via a bifunctional anti-PD-L1/TGF-βRII agent: Status of preclinical and clinical advances. J. Immunother. Cancer 2020, 8, e000433. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, Q.; Risu, N.; Fu, J.; Zou, Y.; Tang, J.; Li, L.; Liu, H.; Zhou, G.; Zhu, X. Galunisertib enhances chimeric antigen receptor-modified T cell function. Eur. J. Histochem. 2020, 64, 3122. [Google Scholar] [CrossRef] [PubMed]
- Uhl, M.; Aulwurm, S.; Wischhusen, J.; Weiler, M.; Ma, J.Y.; Almirez, R.; Mangadu, R.; Liu, Y.W.; Platten, M.; Herrlinger, U.; et al. SD-208, a novel transforming growth factor beta receptor I kinase inhibitor, inhibits growth and invasiveness and enhances immunogenicity of murine and human glioma cells in vitro and in vivo. Cancer Res. 2004, 64, 7954–7961. [Google Scholar] [CrossRef]
- Stüber, T.; Monjezi, R.; Wallstabe, L.; Kühnemundt, J.; Nietzer, S.L.; Dandekar, G.; Wöckel, A.; Einsele, H.; Wischhusen, J.; Hudecek, M. Inhibition of TGF-β-receptor signaling augments the antitumor function of ROR1-specific CAR T-cells against triple-negative breast cancer. J. Immunother. Cancer 2020, 8, e000676. [Google Scholar]
- Premkumar, K.; Shankar, B.S. TGF-βR inhibitor SB431542 restores immune suppression induced by regulatory B-T cell axis and decreases tumour burden in murine fibrosarcoma. Cancer Immunol. Immunother. 2020. [Google Scholar] [CrossRef]
- Wang, Q.M.; Tang, P.M.; Lian, G.Y.; Li, C.; Li, J.; Huang, X.R.; To, K.F.; Lan, H.Y. Enhanced Cancer Immunotherapy with Smad3-Silenced NK-92 Cells. Cancer Immunol. Res. 2018, 6, 965–977. [Google Scholar] [CrossRef]
- Wang, Z.; Guo, L.; Song, Y.; Zhang, Y.; Lin, D.; Hu, B.; Mei, Y.; Sandikin, D.; Liu, H. Augmented anti-tumor activity of NK-92 cells expressing chimeric receptors of TGF-βR II and NKG2D. Cancer Immunol. Immunother. 2017, 66, 537–548. [Google Scholar] [CrossRef]
- Rautela, J.; Dagley, L.F.; de Oliveira, C.C.; Schuster, I.S.; Hediyeh-Zadeh, S.; Delconte, R.B.; Cursons, J.; Hennessy, R.; Hutchinson, D.S.; Harrison, C.; et al. Therapeutic blockade of activin-A improves NK cell function and antitumor immunity. Sci. Signal. 2019, 12, eaat7527. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Wang, Q.; Wang, D.; Zheng, H.; Kalvakolanu, D.V.; Lu, H.; Wen, N.; Chen, X.; Xu, L.; Ren, J.; et al. RGFP966, a Histone deacetylase 3 inhibitor, promotes glioma stem cell differentiation by blocking TGF-β signaling via SMAD7. Biochem. Pharmacol. 2020, 180, 114118. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Rezov, V.; Joensuu, E.; Vartiainen, V.; Rönty, M.; Yin, M.; Myllärniemi, M.; Koli, K. Pirfenidone decreases mesothelioma cell proliferation and migration via inhibition of ERK and AKT and regulates mesothelioma tumor microenvironment in vivo. Sci. Rep. 2018, 8, 10070. [Google Scholar] [CrossRef]
- Cevik, O.; Acidereli, H.; Turut, F.A.; Yildirim, S.; Acilan, C. Cabazitaxel exhibits more favorable molecular changes compared to other taxanes in androgen-independent prostate cancer cells. J. Biochem. Mol. Toxicol. 2020, 34, e22542. [Google Scholar] [CrossRef]
- Duffy, D.J.; Krstic, A.; Halasz, M.; Schwarzl, T.; Konietzny, A.; Iljin, K.; Higgins, D.G.; Kolch, W. Retinoic acid and TGF-β signalling cooperate to overcome MYCN-induced retinoid resistance. Genome Med. 2017, 9, 15. [Google Scholar] [CrossRef]
- Lin, S.H.; Wang, B.Y.; Lin, C.H.; Chien, P.J.; Wu, Y.F.; Ko, J.L.; Chen, J.J. Chidamide alleviates TGF-β-induced epithelial-mesenchymal transition in lung cancer cell lines. Mol. Biol. Rep. 2016, 43, 687–695. [Google Scholar] [CrossRef]
- Petanidis, S.; Kioseoglou, E.; Domvri, K.; Zarogoulidis, P.; Carthy, J.M.; Anestakis, D.; Moustakas, A.; Salifoglou, A. In vitro and ex vivo vanadium antitumor activity in (TGF-β)-induced EMT. Synergistic activity with carboplatin and correlation with tumor metastasis in cancer patients. Int. J. Biochem. Cell Biol. 2016, 74, 121–134. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, L.; Wang, Y.; Dong, S.; Yang, S.; Guan, Y.; Wu, X. Astragaloside IV inhibits cell proliferation in vulvar squamous cell carcinoma through the TGF-β/Smad signaling pathway. Dermatol. Ther. 2019, 32, e12802. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Tey, S.L.; Ho, Y.; Chin, Y.T.; Wang, K.; Whang-Peng, J.; Shih, Y.J.; Chen, Y.R.; Yang, Y.N.; Chen, Y.C.; et al. Heteronemin Induces Anti-Proliferation in Cholangiocarcinoma Cells via Inhibiting TGF-β Pathway. Mar. Drugs 2018, 16, 489. [Google Scholar] [CrossRef]
- Su, Q.; Fan, M.; Wang, J.; Ullah, A.; Ghauri, M.A.; Dai, B.; Zhan, Y.; Zhang, D.; Zhang, Y. Sanguinarine inhibits epithelial–mesenchymal transition via targeting HIF-1α/TGF-β feed-forward loop in hepatocellular carcinoma. Cell Death Dis. 2019, 10, 939. [Google Scholar] [CrossRef]
- Wu, C.; Chen, M.; Sun, Z.; Ye, Y.; Han, X.; Qin, Y.; Liu, S. Wenshen Zhuanggu formula mitigates breast cancer bone metastasis through the signaling crosstalk among the Jagged1/Notch, TGF-β and IL-6 signaling pathways. J. Ethnopharmacol. 2019, 232, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Lee, S.; Lee, H.; Shin, D.; Min, D.; Kim, M.; Ryu, B.; Kim, H.W.; Bae, H. A standardized herbal extract PM014 ameliorates pulmonary fibrosis by suppressing the TGF-β1 pathway. Sci. Rep. 2018, 8, 16860. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Sheng, J.; Wang, M.; Luo, H.; Zhu, J.; Zhang, B.; Liu, Z.; Yang, X. Combination Therapy of TGF-β Blockade and Commensal-derived Probiotics Provides Enhanced Antitumor Immune Response and Tumor Suppression. Theranostics 2019, 9, 4115–4129. [Google Scholar] [CrossRef] [PubMed]
- Pu, N.; Zhao, G.; Yin, H.; Li, J.-A.; Nuerxiati, A.; Wang, D.; Xu, X.; Kuang, T.; Jin, D.; Lou, W.; et al. CD25 and TGF-β blockade based on predictive integrated immune ratio inhibits tumor growth in pancreatic cancer. J. Transl. Med. 2018, 16, 294. [Google Scholar] [CrossRef]
- Jiang, J.; Zhang, Y.; Peng, K.; Wang, Q.; Hong, X.; Li, H.; Fan, G.; Zhang, Z.; Gong, T.; Sun, X. Combined delivery of a TGF-β inhibitor and an adenoviral vector expressing interleukin-12 potentiates cancer immunotherapy. Acta Biomater. 2017, 61, 114–123. [Google Scholar] [CrossRef]
- Bhola, N.E.; Balko, J.M.; Dugger, T.C.; Kuba, M.G.; Sánchez, V.; Sanders, M.; Stanford, J.; Cook, R.S.; Arteaga, C.L. TGF-β inhibition enhances chemotherapy action against triple-negative breast cancer. J. Clin. Investig. 2013, 123, 1348–1358. [Google Scholar] [CrossRef]
- Vanpouille-Box, C.; Diamond, J.M.; Pilones, K.A.; Zavadil, J.; Babb, J.S.; Formenti, S.C.; Barcellos-Hoff, M.H.; Demaria, S. TGFβ Is a Master Regulator of Radiation Therapy-Induced Antitumor Immunity. Cancer Res. 2015, 75, 2232–2242. [Google Scholar] [CrossRef]
- Rodríguez-Ruiz, M.E.; Rodríguez, I.; Mayorga, L.; Labiano, T.; Barbes, B.; Etxeberria, I.; Ponz-Sarvise, M.; Azpilikueta, A.; Bolaños, E.; Sanmamed, M.F.; et al. TGFβ Blockade Enhances Radiotherapy Abscopal Efficacy Effects in Combination with Anti-PD1 and Anti-CD137 Immunostimulatory Monoclonal Antibodies. Mol. Cancer Ther. 2019, 18, 621–631. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).