Simple Summary
Ovarian cancer is the leading cause of death from gynecological malignancies. Recent studies have focused on ovarian cancer-associated microRNAs that play strong regulatory roles in various cellular processes. While miRNAs have been shown to participate in regulation of tumorigenesis and drug responses through modulating the DNA damage response (DDR), little is known about their potential influence on sensitivity to chemotherapy. The main objective of this review is to summarize recent findings on the utility of miRNAs as ovarian cancer biomarkers and their regulation of DDR or modified replication stress response proteins.
Abstract
Genomic alterations and aberrant DNA damage signaling are hallmarks of ovarian cancer (OC), the leading cause of mortality among gynecological cancers worldwide. Owing to the lack of specific symptoms and late-stage diagnosis, survival chances of patients are significantly reduced. Poly (ADP-ribose) polymerase (PARP) inhibitors and replication stress response inhibitors present attractive therapeutic strategies for OC. Recent research has focused on ovarian cancer-associated microRNAs (miRNAs) that play significant regulatory roles in various cellular processes. While miRNAs have been shown to participate in regulation of tumorigenesis and drug responses through modulating the DNA damage response (DDR), little is known about their potential influence on sensitivity to chemotherapy. The main objective of this review is to summarize recent findings on the utility of miRNAs as cancer biomarkers, in particular, ovarian cancer, and their regulation of DDR or modified replication stress response proteins. We further discuss the suppressive and promotional effects of various miRNAs on ovarian cancer and their participation in cell cycle disturbance, response to DNA damage, and therapeutic functions in multiple cancer types, with particular focus on ovarian cancer. Improved understanding of the mechanisms by which miRNAs regulate drug resistance should facilitate the development of effective combination therapies for ovarian cancer.
1. Introduction
Ovarian cancer is the leading cause of death from gynecological malignancies. The high fatality rate is linked to the complexity of the disease and consequent difficulty in making an accurate diagnosis. At the initial stages of disease progression, patients present with non-specific symptoms [1]. The majority of cases are diagnosed at the third or fourth stage of clinical advancement following the spread of disease to other organs. At present, standard treatment for ovarian cancer involves total removal of tumor mass and any tissue that may pose risk of spread. In the case of highly advanced tumors, radical surgery is not possible [2]. Debulking surgery is performed, followed by adjuvant treatment with drugs containing platinum compounds or taxane-based chemotherapy, which has shown success in improving the survival rates of patients in the fourth stage of the disease. However, the five-year survival rate for advanced-stage cases remains below 30% [3]. Ovarian cancer cells often carry BRCA1 or BRCA2 (BRCA1/2), germline or somatic mutations in ataxia telangiectasia and Rad3-related protein (ATR) or checkpoint kinase 1 (CHK1) genes of the homologous recombination (HR) pathway [4]. Poly (ADP ribose) polymerase inhibitors (PARPis) have been identified as the most promising targeted therapy for ovarian cancer. In the United States, the FDA approved olaparib for maintenance treatment of patients with BRCAMUT advanced epithelial ovarian cancer (EOC) showing complete or partial response to first-line platinum-based chemotherapy in 2018 [5].
Elucidation of molecular alterations in serous ovarian carcinoma cells is necessary to identify novel targets for early detection and treatment. Recent studies have focused on ovarian cancer-associated microRNAs (miRNAs) that play strong regulatory roles in various cellular processes. Initial findings support a potential correlation between miRNAs and cancer development. Even low-level disruption in expression patterns of individual miRNAs can lead to significant pathological changes, such as neoplasia. Alterations in miRNA expression are widely reported in multiple cancer types, especially ovarian cancer resistant to chemotherapy [6]. Knowledge of the specific associations between miRNAs and DNA damage response (DDR) or DNA repair should aid in expanding applications of miRNAs in cancer therapy. Cells detect DNA damage and coordinate an appropriate response involving activation of repair pathways, such as nucleotide excision repair (NER) and HR. If damage is too excessive for repair, an apoptotic response is initiated through activating death receptors or triggering intrinsic apoptosis.
In this review, earlier findings on the direct effects of miRNAs on sensitivity of ovarian cancer cells to replication stress response (RSR) inhibitors are described. The therapeutic potential of the miRNAs regulating DDR/DNA repair is discussed, along with the mechanisms by which miRNAs affect sensitivity to PARP, ATR, and CHK1 inhibitor therapy. The identification of mediator miRNAs that improve response to treatment with checkpoint inhibitors would increase the proportion of patients benefiting from therapy.
2. Participation of miRNAs in Pathogenesis and Development of Neoplastic Diseases
MiRNAs are a class of small, endogenous, and noncoding RNAs that post-transcriptionally regulate gene expression. MiRNA is transcripted in the nucleus, usually by RNA polymerase II, to produce the primary miRNA (pri-miRNA). Pri-miRNA is identified and cleaved by the Drosha, an RNase III enzyme, and its cofactor DGCR8 (Pasha), which form a hairpin precursor miRNA (pre-mRNA). The hairpin precursor is exported out of the nucleus by Exportin 5, where Dicer (RNase III enzyme) cleaves double stranded mi-RNA and creates miRNA as transient 21–24 nucleotide duplex miRNA. The strand of mature sequence is then transported onto Argonaute (Ago) and is loaded into a protein complex called RISC. MiRNA recognizes their target sequences based on complementarity to the 3′untranslated region (3′UTR) of mRNA transcripts, leading to translational inhibition and/or mRNA degradation [7,8], Figure 1.
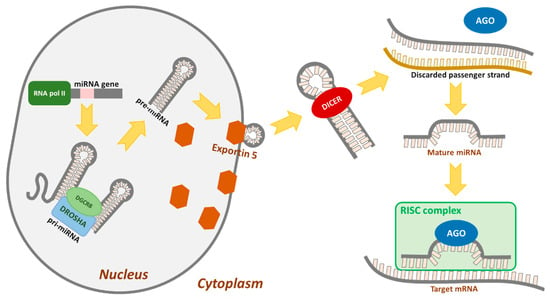
Figure 1.
The main molecular steps involved in miRNA formation and organization.
Almost half the miRNA genes are located in fragile sites of the genome where chromosome fragments are lost or rearranged with high frequency. Mutations in these areas are often linked to cancer development, implicating microRNAs in the formation and progression of neoplasms [7,8,9]. Expression of miRNA genes located near these regions is commonly disrupted. One example is miR-15a and miR-16-1 genes located on the long arm of chromosome 13 in region 14.2 where deletions are frequent. Reduced levels or complete absence of miR-15a and miR-16-1 are detected in many patients with B-cell chronic lymphocytic leukemia, ovarian and prostate cancer, mantle cell lymphoma, and multiple myeloma [10,11,12]. MiRNAs are also secreted from both normal and cancer cells in exosomes, small vesicles that play a key role in cell-to-cell communication in the body [13]. In addition, variable environmental factors, such as low pH and hypoxia (characteristic of most solid tumors, including ovarian cancer), affect miRNA expression and promote exosome secretion. Several indications support the potential utility of miRNAs circulating in the bloodstream as biomarkers of cancer. One hypothesis is that miRNAs appear in the bloodstream through two mechanisms, one associated with tissue damage, as demonstrated in earlier studies (e.g., miR-208 is observed in serum after cardiac muscle damage) [14], and the second related to so-called microbubbles (exosomes) involved in tumor-associated immunosuppression, metastasis, and angiogenesis that are derived directly from the cytoplasmic membrane and reflect the antigenic composition of parent cells [15]. Secreted miRNAs can also be an additional source of information about defects in the DNA repair system, including those related to replication stress (miR-200c, miR-214 [16], miR-185-5p [17], miR-126, miR-17, miR-92a [18], and miR-34a [19]). In 2008, Taylor et al. first demonstrated that eight exosomal miRNAs (miR-21, miR-141, miR-200a, miR-200b, miR-200c, miR-203, miR-205, and miR-214) were elevated in the serum of ovarian cancer patients, even in the case of patients with early stages of the disease. Very importantly, the miRNA profiles observed in exosomes were similar to those in the originating tumor cells. Circulating miRNA profiles accurately reflect the tumor profiles, which make them potential biomarkers and relevant for ovarian cancer diagnosis prognosis and therapeutics [16,20,21]. Since that time, other studies confirmed other circulating miRNA profiles in plasma samples of ovarian cancer as possible biomarkers of which some miRNA were significantly increased, e.g., hsa-miR-106a-5p, hsa-let-7d-5p, hsa-miR-93-5p [17], miR-1274a, miR-625-3p, and miR-720 [18], and others significantly decreased, e.g., hsa-miR-122-5p, hsa-miR-185-5p and hsa-miR-99b-5p [17], miR-106b, miR-126, miR-150, miR-17, miR-20a, and miR-92a [18]. Maeda et al. recently described the potential role of serum miR-34a in early diagnosis of ovarian cancer and for histological subtyping of EOC [19]. In the case of ovarian cancer patients, the elevated level of miRNA (in comparison to healthy) was reported not only in serum exosomes, but also, e.g., in ascites: miR-21, miR-23b, or miR-29a [22], or in urine: miR-30-5p [23]. It is considered that these exosomes are responsible for inducing more aggressive disease, so it confirms that they also might serve as a promising diagnostic and therapeutic targets [21,22]. The correlation between the increased levels of miR-200b and miR-200c with the main marker of ovarian cancer-CA125, commonly used in diagnosis [24], was also observed. Additionally, studies performed by Kapetanakis et al. [25] demonstrated that miR-200b was able to predict the sensitivity to treatment in much more sensitive manner than CA125. After primary treatment (surgery and chemotherapy) of the group of 33 ovarian cancer patients, CA125 very quickly (even after 1 month after treatment) returned to a normal level in almost all patients, whereas there was a difference in the level of miR-200b between individuals. The patients with a negative miR-200b variation had a longer progression-free survival (PFS), than those patients with a positive variation. Increased levels of specific mRNAs characteristic of certain cancer types (breast, lung, ovary, prostate, pancreas, liver, and colon cancer and chronic myeloid leukemia) are often associated with tumor invasiveness or metastasis. The miRNA molecules known to inhibit the processes of migration and invasion of neoplastic cells include miR-149 (breast cancer), miR-138 (ovarian and kidney cancer), miR-126 (lung and stomach cancer), and miR-206 (melanoma and cervical cancer), among others. Additionally, miR-373 and miR-520c are associated with the invasive and metastatic ability of cancer cells. These molecules directly inhibit expression of the CD44 surface receptor responsible for binding hyaluronan (the main component of the extracellular matrix), an intermediary for several stimulatory processes, such as migration and proliferation. Moreover, miRNAs are involved in epithelial-to-mesenchymal transition (EMT), a necessary step in metastasis. Increased levels of specific miRNAs are additionally associated with the occurrence of epigenetic abnormalities in cancer cells [26,27] Molecular functions of miRNAs in ovarian cancer acting as oncogenes or suppressors are presented in Table 1.

Table 1.
Molecular functions of miRNAs in ovarian cancer.
3. Aberrant Expression Profiles of miRNAs in Ovarian Cancer
Recent studies revealed differences in the miRNA expression profiles in tissues from patients with ovarian cancer and healthy individuals. Increased miRNA levels were not detected in patients with benign ovarian disease [73]. Further comparison of expression levels of miRNAs in ovarian cancer revealed distinct roles of different miRNAs. Expression of miR-200a, miR-200b, and miR-200c was significantly higher than that in normal tissues, whereas mir-199a, miR-140, miR-145, and miR-125b1 displayed low expression in ovarian cancer tissues [74]. MiR-10a and miR-10b suppressed proliferation of granulosa cells in the ovary. The miR-10 family suppressed expression of several key genes in the transforming growth factor beta (TGF-β) pathway, suggesting a negative feedback loop between the miR-10 family and TGF-β pathway [34]. Numerous studies performed on various tissue types have validated the utility of miRNAs as a prognostic marker of ovarian cancer [75,76,77].
3.1. MiRNAs as Tumor Suppressors
Suppressor genes, also known as anti-oncogenes, encode proteins that inhibit the processes of cell growth and differentiation and maintain genetic stability of the cell. Mutations in these genes can lead to uncontrolled cell proliferation and, consequently, development of cancer. The effect of miRNAs in the case of their activation or deactivation will lead to an insufficient number of target genes or their overexpression, respectively. Target miRNA transcripts determine whether the miRNA should be considered an oncogene or a tumor suppressor [78,79]. Biological functions of miRNAs depend on the cellular context, tumor molecular subtype, stage of tumor progression, or interactions with therapy [80]. It was observed that miR-200c and miR-141 produce resistance to carboplatin while sensitizing MES-OV/TP cells to paclitaxel. The authors suggest that the effects of these miRNAs on drug sensitivity are cell context dependent [6]. Higher miR-200c levels were also associated with better progression-free survival in stage I epithelial ovarian cancer [81]. Liu et al. found that miR-200b and miR-200c increased cisplatin sensitivity through downregulation of DNA methyltransferases (DNMT3A/DNMT3B) and the indirect downregulation of DNMT1 by targeting Sp1 transcription factor [26]. Based on 220 ovarian cancer patients’ analysis, it was observed that overexpression of miR-200c correlated with poor or good outcome depending on the cellular localization of HuR (RNA binding protein). MiR-200c can act either as a suppressor or enhancer of the aggressive phenotype, depending upon the localization of HuR. Suppressor genes contribute to drug resistance of several types of solid tumors [82]. Tumor suppressor miRNAs prevent tumor development through negative regulation of genes that control cell differentiation or apoptosis. To date, a number of miRNAs have been identified as tumor suppressors [83]. For instance, miR-29 significantly reduces migration of highly metastatic ovarian cancer cells [36]. Expression of miR-29 alone or in combination with cisplatin could effectively reduce tumorigenicity of CP70 ovarian cancer cells [35].
One of the most well-characterized tumor suppressors of the miRNA family in ovarian cancer is Let-7, which belongs to a family of highly homologous members. Ten mature subtypes of the human let-7 family have been identified to date, whereby mature let-7a and let-7f are processed from precursor sequences (let-7a-1, let-7a-2, let-7a-3; let-7f-1, and let-7f-2) [38,39,40]. Let-7 inhibits cell proliferation and increases apoptosis by inhibiting expression of proto-oncogenes, such as the small GTPase RAS, high mobility group AT-hook 2 (HMGA 2), c-Myc, cell division cycle homolog 25A (CDC25A), cdk 6, and cyclin 2 [39,84]. Overexpression of let-7g miRNA in OVCAR3 and HEY-A8 EOC cells induced cell cycle arrest, slowed progression of EMT, and significantly improved cell response to cis-platinum treatment. Let-7g worked through vimentin and reduced the expression of Snail and Slug (the protein product of snail family transcriptional repressor 2) [39]. Other studies have demonstrated overexpression of miR-16 in ovarian cancer tissues, including SKOV3 and OVCAR3 cell lines, compared with normal ovarian epithelial cells. MiR-16 is reported to exert suppressive effects on cell migration and invasion by inactivation of the Wnt/β-catenin signaling pathway through decreasing expression of matrix metallopeptidase MMP2 and MMP9. Additionally, miR-16 regulates the expression of mesenchymal markers (cadherin 1 and 2, snail 1 and 2, vimentin, and twist family BHLH transcription factor) [28]. MiR-31 is another microRNA with biological significance. This miRNA is expressed at low levels in serous ovarian cancer cells and tissues and induces suppression of cell proliferation, clonogenic potential, and cell migration and invasion [43]. Recent research indicates that miR503HG interacts with and promotes methylation of miR-31-5p that play a role in inhibition of ovarian cancer cell invasion and migration [42]. MiR-506-3p inhibits proliferation and promotes apoptosis via inactivation of the NAD-dependent protein deacetylase sirtuin-1 (SIRT1)/AKT/Forkhead box protein 3a (FOXO3a) signaling pathway [45]. Myotubularin-related protein 6 (MTMR6) has been identified as another functional target of miR-506-3p. Several recent studies indicate that malignant biological behaviors are regulated by the myotubularin (MTM) protein family [85]. Other miRNAs acting as suppressors include miR-424-5p and miR-503-5p that directly target the 3′UTR of KIF23 (kinesin-6, a plus-end-directed motor protein in mitosis) to suppress its expression and inhibit ovarian cancer cell proliferation and migration [47]. Additionally, miR-199a-5p is reported to function as a suppressor of ovarian cancer (HO-8910 and ES-2) cell proliferation and invasion through inhibiting NF-κB1 expression. Notably, expression patterns of matrix metalloproteinases (MMP-2 and MMP-9) are altered in a similar manner as NF-κB1 upon exogenous expression of miR-199a-5p [51]. The anti-oncomiR list includes miRNAs from the miR-34 family that inhibit oncogenes, such as c-MYC and c-MET, or promote mitosis CDKs [53] and miR-340-5p. Deficiency of miR-340-5p promotes expression of serine/threonine-protein kinase B-raf (BRAF), NF-kB and ATP-binding cassette sub-family B member 5, also known as P-glycoprotein (ABCB5), resulting in development of drug resistance [55].
3.2. MiRNAs as Oncogenes
Alterations in expression of several miRNAs are observed in many cancer types [81,86,87]. Mutation in a single allele of proto-oncogenes can trigger transformation into oncogenes. These genes promote cancer development by negatively regulating the tumor genes responsible for cell differentiation or apoptosis [88]. Several miRNAs in tumor cells exhibit oncogenic traits and promote tumorigenesis. Notably, almost all members of the miR-200 family (miR-200a, miR-200b, miR-200c, miR-141, and miR-429) are upregulated in ovarian cancer [89]. Different miRNA types, including miR-182 and the miR-200 family (specifically, miR-200a, miR-200b, and miR-200c), are highly overexpressed in high-grade serous epithelial ovarian cancer (SEOC). The miR-200 family participates in EMT through regulating E-cadherin by inhibiting zinc-finger E-box-binding homeobox 1 (ZEB1) and zinc-finger E-box-binding homeobox 2 (ZEB2) [30] and improves response to paclitaxel (PTX) due to repression of the miR-200c target, ZEB1. The transcription factor, Grainyhead-like 2n (GRHL2), acts as a pivotal gatekeeper of EMT in EOC via miR-200-ZEB1 [31]. The miR-200 family also sensitizes ovarian cancer cells to PTX through downregulation of TUBB3/class III beta-tubulin, a component of microtubules that binds paclitaxel [90]. Moreover, in PTX resistant cells (A2780/1A9, MES-OV, OVCAR-3, ES-2), miR-200b and miR-200c were downregulated and associated with EMT, with increased vimentin, fibronectin1, MMP2, or MMP9 [90]. MiR-200a is reported to enhance sensitivity to PTX-induced reactive oxygen species production. Overexpression of miR-200a-3p markedly promotes proliferation, colony formation, and invasion of ovarian cancer cells. Expression of this miRNA in ovarian cancer tissues is significantly negatively correlated with that of Protocadherin-9, a potential tumor suppressor, in a variety of cancers [32]. Moreover, the miR-200 family plays a major role in regulating EMT and sensitivity to carboplatin and PTX in OVCAR-3 and MES-OV cells. Inhibition of miR-200c and miR-141 resulted in the downregulation of E-cadherin and the upregulation of vimentin and fibronectin [33].
MiR-205 expression is significantly increased with a simultaneous decrease in transcription factor 21 (TCF21, a tumor suppressor gene) in epithelial ovarian carcinoma compared to normal ovarian cells. Thus, miR-205 is regarded as an oncogene in ovarian cancer that plays critical roles in tumor invasion and metastasis [41]. MiRNA-126-3p is also implicated in cancer progression and inflammation. Overexpression of miR-126-3p in OVCAR3 cells is reported to suppress cell proliferation and invasion as well as phosphorylation of serine/threonine-specific protein kinase B (AKT) and extracellular signal-regulated kinases ½ (ERK1/2) [44]. MiR-183 exerts tumor-promoting effects in ovarian cancer by regulating one of the transcription factor proteins, Mothers against decapentaplegic homolog 4 (Smad 4), via the TGF-β/Smad4 pathway. MiR-183 is upregulated in OC tissues and cell lines. Downregulation of miR-183 via cell transfection inhibited proliferation and invasion and induced apoptosis in SKOV3 and OVCAR3 cells [46]. Expression of miR-760 is markedly upregulated in association with an aggressive phenotype of OC and poor prognosis [48,49]. Additionally, miR-151 plays an oncogenic role in carcinogenesis and progression of ovarian cancer by activating AKT/mTOR signaling through effects on the Rho guanine nucleotide dissociation inhibitor (RhoGDIA). MiR-151 activates Ras-related C3 botulinum toxin substrate 1 (Rac1), Cdc42, and Rho GTPase by directly targeting the 3-UTR of RhoGDIA, a metastasis suppressor [50]. Examples of oncomiRs include miR-21-5p, which controls the suppressor gene phosphatase and tensin homolog (PTEN, an inhibitor of the Akt kinase pathway) [52], miR-106a, which regulates the p21 protein level, and miR-195, which controls WEE1 kinase, an inhibitor of cell division [54]. MiR-222 is overexpressed in EOC cases and promotes proliferation through downregulation of target cyclin-dependent kinase inhibitor p27Kip1 [57]. An earlier study reported upregulation of miR-221 in 63 samples of ovarian cancer. A negative correlation between expression of apoptosis protease activator 1 (APAF1) protein and miR-221 in 5 of 63 ovarian cancer tissues and six cell lines was observed, including A2780, OVCAR3, SKOV3, and 3AO5 [60]. An in vitro cell viability assay showed that downregulation of miR-221/222 sensitized A2780/CP cells to cisplatin-induced cytotoxicity [58]. Another identified oncomir shown to promote proliferation of SKOV3, Hey, and OVCAR3 cells is miR-520b, which targets the ring finger protein 216 (RNF216) gene to promote cell growth. The negative correlation between miR-520b and RNF216 may present a new strategy for ovarian cancer [62]. In addition, numerous studies have shown that oncomirs play an important role in the acquisition of the ability to invade and form metastases by cancer cells. Overexpression of miR-10b in ovarian cancer has been reported in association with reduced amounts of transcription factor, HOXD10, in altered cells, leading to an increase in the levels of ras homolog family member C (RhoC) and matrix metallopeptidase 14 (MMP14), which are responsible for metastasis [64].
4. MiRNA Functions in Cancer Based on Regulation of DDR
The DNA damage response is a complex network involving proteins that are activated to facilitate detection of DNA damage and determine the survival or death of cells exposed to stress via stimulation of the signal transduction cascade [91]. Activation of the DDR pathway triggers cell cycle checkpoint activation and dividing alternation, in turn preventing the transfer of damaged DNA to daughter cells. Simultaneously, DNA repair mechanisms are activated. Upon repair of damage, cell cycle and division resume, allowing survival and continuation of function. If repair is not possible due to an excessive number of lesions, cells are eliminated by triggering programmed cell death or cellular aging, irreversible cell cycle arrest, and division processes [92], as presented in Figure 2. DDR modulates miRNA expression in transcriptional and post-transcriptional levels and involves miRNA degradation [66,93,94]. On the other hand, miRNAs may directly modulate the expression of multiple proteins in the DDR pathways.
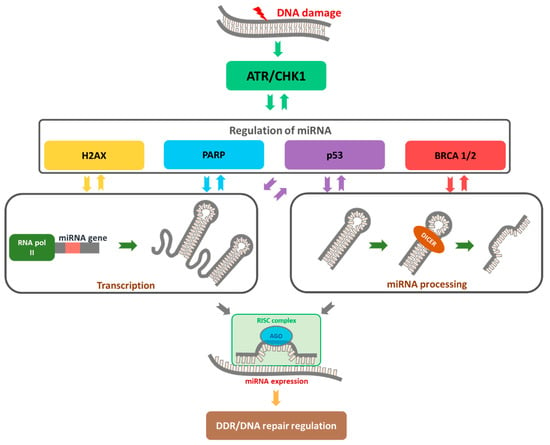
Figure 2.
The role of miRNAs as a regulator of DDR. DNA damage triggers the activation of the ATR/CHK1 pathway, which is responsible for substrates phosphorylation (i.e., H2AX, p53, and BRCA 1/2) to repair DNA damage. Single-strand DNA damage is identified and repaired by poly (ADP-ribose) polymerase (PARP) pathway activation, predominantly through base excision repair (BER). With continuous PARP inhibition, ssDNA breaks are converted to dsDNA breaks during DNA replication in which replication forks stall at the point of DNA damage. Over-expressed or down-regulated miRNAs may affect DDR by changing the expression of repair genes.
ATR and Ataxia telangiectasia mutated (ATM) kinases belonging to the phosphatidylinositol 3-kinase-related kinases (PIKK) family play central roles in activation of the DDR pathway [95]. Histone H2AX, one of the first known substrates for ATR and ATM kinases, is expressed in high-grade SOC, mucinous adenocarcinomas, and clear cell carcinomas. Significant changes in the gene and protein levels of H2AX have been reported in OC, supporting its predictive value as a biomarker [96].
4.1. MiRNAs Are Involved in Cell Cycle Disruption
Imbalances in activities of miRNA molecules significantly affect cell cycle regulation, leading to excessive proliferation. Disruption of this process is often associated with direct interactions of miRNAs with key regulatory molecules of signaling pathways underlying proliferation, e.g., PTEN, Myc, Ras, and V-abl Abelson murine leukemia viral oncogene homolog 1 (ABL1), as well as proteins from the Rb pathway, cyclin-CDK complexes, or cell cycle inhibitors from families of inhibitors of CDK4 (INK4) and CDK-interacting protein/kinase inhibitory protein (Cip/Kip) [97]. Examples include miR-21, which is overexpressed in breast, ovary, and liver cancer, and a group of miR-17-92 members that inhibit PTEN phosphatase activity. Suppression of the gene encoding PTEN promotes cell proliferation. Another miRNA that influences the cell cycle is miR-15b, Figure 3. Decreased expression of miR-15b leads to an increase in cellular cyclin E1, resulting in lack of control during the transition from G1 to S phase. Ectopic expression of miR-192/215 induces cell cycle arrest and targets a number of transcripts that regulate G1/S and G2/M checkpoints [98,99]. These miRNAs are transcriptional targets of p53 and also upregulate p53 by downregulating Murine Double Minute gene 2 protein (MDM2) [100].
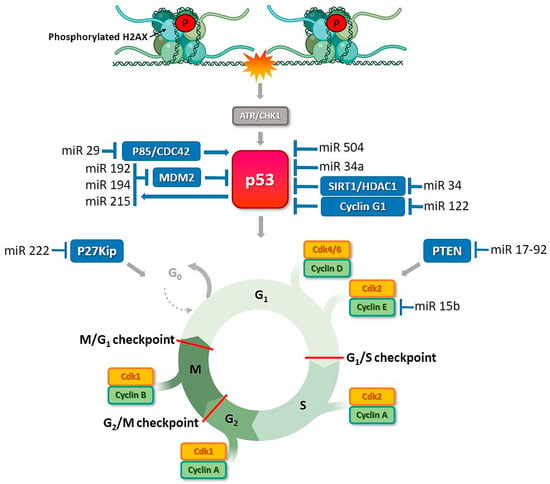
Figure 3.
P53-induced miRNAs control cell cycle and cell survival in ovarian cancer. DNA damage stimulates ATR or DNA-PK kinases that activate TP53 to directly induce many kinds of miRNAs that repress cell-cycle regulators or allow to DNA repair. ATR transduces the DDR signal by phosphorylation of the checkpoint kinase CHK1, which results in cell cycle arrest and DNA repair. MiRNAs, e.g., miR-34a, a direct transcriptional target of TP53, participate in the regulation of TP53 activity. Activated TP53 translocates into the nucleus where it induces the transcription of several targets involved in cell cycle control, DNA repair, or apoptosis. DDR-related proteins that are MiRNAs recognizable are shown in blue box.
4.2. Functional miRNAs in Activation of the “Response Track” to DNA Damage and the Role of H2AX Histone
In response to DNA damage, H2AX is phosphorylated by DNA-dependent protein kinase, catalytic subunit (DNA-PKc), which is also a member of the PIKK family. The histone is phosphorylated at serine 139 (known as γH2AX) and initiates attachment of subsequent elements of the signaling pathway [101]. At the same time, histone H2AX is dephosphorylated at tyrosine 142 and constitutively phosphorylated under conditions of no DNA damage [102]. Dephosphorylation promotes direct attachment of the mediator of DNA damage checkpoint protein 1 (MDC1) protein to γH2AX. Anchoring of MDC1 at the site of damage is a platform for activation of other proteins belonging to the DDR pathway and the MRN (MRE11, Rad50, NBS1)/ATM complex. This enhances local ATM kinase activity and extension of the H2AX phosphorylation region to include nucleosomes adjacent to DNA damage [103,104]. The clusters favor extensive formation of γH2AX, which plays an important role in accumulation and maintenance of components of the DDR pathway, such as MRN, and proteins related to DNA repair, including BRCA1 and p53-binding protein 1 (53BP1). Binding of phosphorylated MDC1 to γH2AX facilitates attachment of E3, RNF8 (E3 ubiquitin-protein ligase), and RNF168 ubiquitin ligases to the lesion site, which promote association of BRCA1 and 53BP1 via ubiquitination of chromatin [91,105,106,107,108]. Downregulation of ubiquitin ligase RNF8, which is necessary for γH2AX to recruit DNA repair proteins to DNA damage sites, via miR-214, induces chromosomal instability in ovarian cancer [109], Figure 4. Thus, H2AX histone appears to play a pivotal role as an early indicator protein for DDR. Previous reports showed that miR-24 and miR-138 regulate H2AX via 3’-UTR attachment. Overexpression of miR-138 inhibited homologous recombination and enhanced cellular sensitivity to multiple DNA damage agents (cisplatin, camptothecin, and ionizing radiation) [98]. MiR-138 was recently identified as an effective tumor suppressor in multiple malignancies including ovarian cancer [56]. MiR-24 mediates suppression of H2AX in terminally differentiated blood cells, which renders them hypersensitive to gamma-irradiation, deficient in DSB repair, and susceptible to chromosomal instability [110]. Another study reported that overexpression of miR-24-insensitive CHEK1 does not rescue the DNA repair phenotype induced by miR-24 [111]. Moreover, γH2AX has been shown to regulate miR-3196 gene expression. H3K27 trimethylation in the miR-3196 promoter region regulated via H2AX phosphorylation at Ser139 is a key step in H2AX-mediated apoptosis [112]. Furthermore, Fra-1 transcriptional factor and miR-134 are upregulated in ovarian cancer tissues. MiR-134 enhances H2AX S139 phosphorylation via activation of c-Jun NH2 kinase (JNK) and promotes DNA repair through non-homologous end-joining (NHEJ) [113].
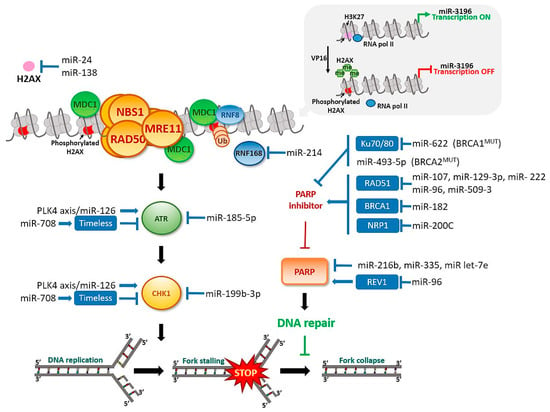
Figure 4.
MiRNAs’ involvement in the DNA damage response in ovarian cancer. DNA synthesis inhibition or damage induces checkpoint responses. PARP and checkpoint proteins controlled by the ATR–CHK1 pathway prevent fork collapse, replication stress, and genome instability. DDR-related proteins that are MiRNAs recognizable are shown in blue box.
4.3. MiRNAs Contributes to DSB DNA Damage Repair System
The primary function of the DDR pathway is to identify DNA damage and, where possible, initiate repair processes. The majority of DNA damage is repaired by the triggering of catalytic event sequences involving multiple proteins, including base excision repair (BER), NER, mismatch repair (MMR), HR, and NHEJ. Two types of nucleotide excision repair pathways exist. One is active during transcription (transcription coupled repair, TCR), while the other is independent of transcription (Global Genomic Repair, GGR) [114,115]. Activation of a specific mechanism depends on the type of DNA damage. BER, NER, and MMR pathways play key roles in repairing damage such as single DNA strand breaks (SSB), replication errors, insertions, deletions, and adducts [116,117].
Double-strand breaks (DSB) are one of the most dangerous types of DNA damage, and a single unrepaired DSB is sufficient to trigger apoptosis [118]. Two processes are involved in repair of double-strand breaks, specifically, HR and NHEJ. Homologous recombination can occur in the S and G2 phases of the cell cycle. On the other hand, repair of damage by non-homologous recombination is possible at any phase of the cell cycle, including G0 [119,120,121]. In HR repair, proteins of the MRN complex and BRCA1 C-terminal Interacting Protein (CtIP) play a key role. These proteins are involved in formation of short sections of single-stranded DNA (ssDNA), which initiate repair of damage through homologous recombination. With the aid of BRCA1, BRCA2, and RAD51 proteins, short sections of single-stranded DNA are joined to the undamaged template. In conjunction with the activities of polymerase, nuclease, helicase, and other proteins, damage is repaired. HR is also involved in resumption of replication caused by blockage of replication forks [122,123]. One of the key proteins of the MRN complex is RAD51. In an earlier study, upregulation of miR-210 significantly suppressed expression of RAD51, while upregulation of miR-373 inhibited RAD52 (which recognizes double-strand breaks and adheres to the free ends of the break) [110]. Another study by Moskwa et al. [124] consistently showed that miR-182 downregulates BRCA1 expression. MiR-182 enhances BRCA1 protein levels and protects against irradiation-induced cell death, while its overexpression reduces BRCA1 protein, impairs homologous recombination-mediated repair, and renders cells hypersensitive to irradiation. Subsequently, ability of HR to stimulate DSB repair is significantly decreased [124].
In the case of NHEJ, DSB are recognized by the heterodimeric Ku70/Ku80 protein complex, which binds DNA-PKc kinase. Subsequently, DNA polymerases and DNA ligase IV, enzymes that process DNA ends, are recruited and activated. In addition, it is possible to repair DNA damage related to joining non-homologous ends. This process, known as alternative NHEJ (alt-NHEJ) or microhomological-mediated end joining (MMEJ), occurs independently of the Ku protein [125,126]. Earlier literature suggests that miR-101 is able to successfully regulate DNA-PKcs and ATM through attaching to their 3’-UTRs. Specifically, upregulation of miR-101 significantly reduced the protein levels of DNA-PKcs and ATM in tumor cells and sensitized them to radiation, both in vitro and in vivo. Thus, miR-101 is a potential option for use in DNA DSB repair gene targeting to optimize the effects of tumor radiotherapy [127].
4.4. MiRNAs Modulate Activity of p53, a Key Protein of the DDR Pathway
The p53 protein is a key suppressor of neoplastic transformation that regulates transcription of numerous genes and interacts directly with multiple proteins. p53 is implicated in a number of critical cell processes, including DNA repair, cell cycle, and programmed cell death. Under conditions where the cell is not exposed to stress factors, the p53 protein level is relatively low. This may be attributed to interactions with (MDM2), which blocks transcriptional activity of p53, leading to its ubiquitination-dependent degradation. MDM2 synthesis is regulated by p53, generating a negative feedback loop leading to a decrease in p53 levels after induction. The imbalance between p53 and MDM2 levels is a critical step in p53 activation [128] and occurs when activated ATM and/or ATR kinase phosphorylates the p53 protein at serine 15 and CHK2 at serine 20. ATM also phosphorylates MDM2 in response to DNA damaging agents. As a result of these modifications, interactions of MDM2 with p53 are blocked, leading to the inhibition of MDM2-dependent degradation and, consequently, accumulation of p53. Thus p53 is activated as a result of post-translational modifications, such as phosphorylation, acetylation, methylation, and ubiquitination. The p53 protein serves as a transcriptional factor to regulate expression of target genes, which also occurs through recruitment of coactivators or corepressors. Among these molecules, acetyltransferases are known to play an important role. Enzymes such as CREB-binding protein (CBP), p300, Tip60, human males absent on the first (Hmof), or P300/CBP-associated factor (PCAF) acetylate p53 and histones alter chromatin conformation, increasing the availability of the DNA template for transcription machinery. In response to DNA damage, CBP and/or p300 acetylate p53 at six lysine residues, which present a target for MDM2 ubiquitination, thereby increasing the stability of p53 and binding ability to DNA [129].
Depending on the type and extent of DNA damage, various post-translational modifications of p53 are initiated, which translate into different cellular responses. Thus, p53 serves as the main decision switch for survival or death. Several groups can be distinguished among the genes regulated by p53 in response to DNA damage. One of these categories is negative regulators/inhibitors of the cell cycle, such as p21, 14-3-3σ, and GADD45α, which trigger cell cycle arrest and division, facilitating repair of DNA damage [130]. In response to DNA damage, p53 is involved in the regulation of processes related to cell metabolism and autophagy. In addition, transcription-independent and miRNA-dependent p53 functions have been reported. MiRNAs either directly target the 3′ UTR of p53 or indirectly regulate p53 activity by modulating proteins associated with p53. Among these microRNAs, miR-504 negatively regulates p53 expression through binding two DNA cis elements located in the 3′ UTR region [131]. DNA damage promotes the p53-dependent upregulation of miR-192, miR-194, and miR-215. Studies also have revealed the existence of a specific p53 binding site around the miR-194/miR-215 cluster [132].
In addition to direct binding to p53, several miRNAs, including miR-34a, miR-29, and miR-122, indirectly modify p53 activity [133,134,135,136]. MiR-34a is a direct transcriptional target of p53 [137,138,139], whereby p53 upregulates miR-34a expression via binding to specific promoter regions. MiR-34a positively regulates p53-dependent apoptosis through another SIRT1 [133]. MiR-34a expression is low in patients with chromosomal abnormalities involving the tumor protein p53 (TP53) gene locus and is associated with poorer prognosis and shorter survival. Mutations or deletions in the 17p13 region of the TP53 gene locus may indirectly lead to reduced miR-34a expression [140]. Another miRNA family involved in p53 regulation is miR-29. Members of this family directly suppress phosphoinositide 3-kinase subunit (P85a) and cell division control protein 42 homolog (CDC42), both of which negatively regulate p53. As a result, miR-29 positively upregulates the p53 level and induces apoptosis and DNA repair in a p53-dependent manner [134].
5. MiRNAs Associated with DNA Repair Checkpoint Proteins: New Options for Optimizing Ovarian Cancer Therapy
5.1. PARP
PARP is an important protein involved in the repair of single-stranded DNA breaks, seen in Figure 3. PARPis have been shown to selectively kill cells with defective HR pathways as a result of synthetic lethality [141]. However, a large proportion of HR-mutated cancers gain resistance to these therapeutic agents. PARPi sensitivity is modulated through downregulation of critical DNA repair genes as a consequence of alterations in miRNA profiles. PARPi resistance may be promoted by miR-622 that modulates the balance of DNA repair through selective inhibition of expression of NHEJ proteins, such as KU70/80, which maintain genome stabilization after treatment with DNA-damaging agents or PARPi. High expression of miR-622 in BRCA1MUT epithelial ovarian cancer is associated with prediction of poorer disease-free and overall survival [69]. The functional impact of miR-493-5p has been characterized in BRCA2MUT cancer cells. MiR-493-5p induces platinum and PARPi resistance by affecting several pathways, including single-strand annealing (SSA), R-loops, and replication fork stability [142]. In contrast, miR-107, miR-129-3p, and miR-222 increase sensitivity to PARP inhibitors and ionizing radiation by causing a reduction in the DNA damage response via impairing the HR pathway based on targeting of RAD51 [143]. Mi182 exerts similar effects and enhances PARPi sensitivity by downregulating BRCA1 [124]. Moreover, expression of miR-96 is increased in many cancer types. This miRNA enhances sensitivity to platinum agents and PARP via downregulation of the DNA repair proteins REV1 and RAD51 [144]. Another study on a patient-derived xenograft (PDX) model of high-grade serous ovarian carcinoma (HGSOC) revealed an essential role of miR-509-3 in tumor suppression and HR signaling, along with increased sensitivity to PARPi treatment [59]. Furthermore, PARP1 expression could be altered by miR-335 or miR-216-b. MiR-335 plays a dual role as either a tumor promoter or suppressor in a wide variety of cancers. However, expression is reduced in ovarian cancer cells and miR-335 shown to effectively increase sensitivity to cisplatin treatment [61]. MiR-216-b regulates apoptosis and autophagy and directly binds to PARP1 mRNA, leading to inhibition of its expression. Lower expression of miR-216b is reported in cisplatin-resistant ovarian cancer cells [145]. Recent studies have additionally demonstrated a role of Neuropilin 1 (NRP1) in response to ovarian cancer therapies. NRP1 is expressed at high levels in resistant cells (SKOV3) and shown to be upregulated in partially sensitive cells (UWB-BRCA) upon prolonged olaparib treatment, resulting in poor drug response. MiR-200c targets and suppresses NRP1 expression in OC cells resistant to therapy, leading to the restoration of olaparib sensitivity [89].
Platinum-resistant ovarian tumors display low miR-Let7i expression. Conversely, its gain of function results in restoration of drug sensitivity in chemoresistant ovarian cancer cells [146]. Agomir is a type of specially labelled and chemically modified double-stranded microRNA that can regulate the biological functions of target genes by mimicking endogenous microRNAs. Let-7e agomir suppressed the mRNA levels of PARP1 and insulin-like growth factor I (IGF-1) while its downregulation enhanced PARP1 and IGF-1 expression [37]. Specific miRNA expression profiles could therefore serve as biomarkers in ovarian cancer to predict response to PARPi therapy.
5.2. ATR
The ATR protein belonging to the phosphatidylinositol 3-kinase-related (PI3K) family is involved in the signaling of stalled replication forks and maintaining genomic stability during the S phase, along with its partners ATR interacting protein (ATRIP) and replication protein A (RPA) [147]. A broad spectrum of DNA damage, such as single- and double-stranded DNA breaks, cross-links, and adducts, can lead to the activation of ATR [148]. ATR is referred to as the “master of DDR”, highlighting the relevance of miRNAs implicated in DDR pathways as novel therapeutic targets for ovarian cancer. MiR-383-5p and miR-185-5p have been shown to be associated with ATR kinase. MiRNA-383-5p is predominantly downregulated and acts as a tumor suppressor in several human cancer types, such as gastric, glioma, medulloblastoma, and testicular embryonal carcinomas. In the mammalian ovary, miR-383 plays a functional role in follicle development [63]. MiR-185 suppresses expression of ATR and activation of its downstream effector, CHK1, which are induced by ionizing radiation. Furthermore, miR-185 is reported to induce G1 cell cycle arrest and apoptosis, inhibiting cancer cell proliferation [149].
A serine/threonine-protein kinase, PLK-4, has been identified as a target of miR-126, which is downregulated in various cancers in correlation with tumor progression and poor prognosis. Earlier experiments showed that PLK-4 knockdown led to a decrease in expression of ATR and CHK1, supporting its interactions with the ATR/CHK1 pathway. Moreover, changes in miR-126 expression led to PLK-4, ATR, and CHK1 dysregulation [67]. Based on these findings, it is proposed that miR-126 inhibits cancer progression via regulation of the cell cycle through inducing alterations in the ATR/CHK1 pathway.
MiR-708 overexpression is associated with suppression of the ATR/CHK1 pathway. Timeless was a direct target of miR-708. Total and phosphorylated ATR and CHK1 levels were decreased in cells overexpressing miR-708 after cisplatin treatment [68], supporting the utility of this miRNA as a potential therapeutic target. Overall, the effects of miRNAs on ATR kinase levels signify their potential application as new therapeutic targets for ovarian cancer.
5.3. CHK1
Checkpoint kinase 1 is a serine/threonine kinase encoded by the CHEK1 gene activated in response to DNA damage and replication stress that is proposed to regulate mitotic progression [150]. ATR and CHK1 share the same signaling pathway. However, in addition to ATR-induced activation, CHK1 can be autophosphorylated and activated independently of ATR [151].
Numerous studies have validated the oncogenic association of miR-424. Decreased expression of miR-424-5p is significantly associated with distant metastasis in high-stage (stage III and IV) ovarian cancers [72]. Moreover, downregulation of miR-424 contributes to the progression of cervical cancer via upregulation of target CHEK1 gene expression and phosphorylation of CHK1 protein, while its overexpression inhibits CHK1 expression [152].
Another miRNA downregulated in serous ovarian tumours is miR-195-5p [153]. In lung tumor tissues, miR-195 expression is low and associated with poor survival outcomes, while overexpression of miR-195 results in suppression of cancer cell growth, migration, and invasion. CHK1 has been identified as a direct target of miR-195. Low expression of miR-195 leads to high expression of CHK1, which is associated with poor prognosis in patients with lung tumors [154].
Expression of miR-330-5p regulates the development of different tumor cell types. In cutaneous malignant melanoma, miR-330 suppresses cell proliferation as well as expression of tyrosinase and protein disulfide-isomerase A3 (PDIA3) [155]. Conversely, its overexpression could promote apoptosis of prostate cancer cells through E2F1-mediated suppression of RAC-alpha serine/threonine-protein kinase (Akt) phosphorylation [156]. In esophageal adenocarcinoma, miR-330 was shown to modulate neoadjuvant chemoradiotherapy sensitivity [157], while in non-small cell lung cancer, its overexpression inhibited NIN1/RPN12 binding protein 1 homolog (NOB1) expression and cancer cell growth [158]. On the other hand, downregulation of miR-330-5p is reported in epithelial ovarian cancer tissues [159]. Moreover, the long non-coding RNA LINC01224 modulates expression of miR-330-5p, resulting in the downregulation of CHEK1 in hepatocellular carcinoma [160]. CHEK1 has also been identified as a direct target of miR-497, whereby expression of CHK1 protein is negatively regulated by miR-497 and upregulated under conditions of downregulation of miR-497 [161]. Other miRNAs responsible for suppressing expression of CHK1 and Wee1 are miR-16 and miR-26a. During genotoxic stress, p53 upregulates miR-16 and miR-26a, in turn attenuating expression of Wee1 and CHK1 [29]. These effects promote accumulation of cells in the G1 phase and, consequently, apoptosis. Additionally, miR-199b-3p overexpression in ovarian cancer suppresses E-box binding homeobox (ZEB)1 and CHK1. Moreover, E-cadherin and EMT expression were increased, which led to the conclusion that miRNA-199b-3p may suppress the progression of ovarian cancer via the CHK1/E-cadherin/EMT signaling pathway [162].
The collective findings highlight the significance of CHK1 as a key pharmacological target. Inhibition of CHK1 protein induces sensitization of cancer cells to genotoxic therapy and is recognized as beneficial in the treatment of ovarian cancer [163]. Thus, downregulation of CHK1 through targeted miRNAs may present an effective novel therapeutic strategy.
6. Conclusions
Since early detection tools are lacking, ovarian cancer is often diagnosed at late stages, which substantially contributes to the high mortality rates. MiRNAs are implicated in regulating almost every aspect of the DDR, DNA repair, and cell cycle arrest (Figure 2, Figure 3, Figure 4 and Figure 5). MiRNAs may be an alternative method to identify DDR defects in patient therapy. Previously, a miRNA-score was developed that was associated with genome instability and predicted the outcome of ovarian cancer based on mutations in caretaker genes. The authors described 10 miRNAs. Six of them had higher expression than the median value across the dataset and were associated with a high frequency of mutation (miR-151, miR-301b, miR-505, miR-324, miR-502, and miR-421). The other four (let-7a, miR-320, miR-146a, and miR-193a) had lower expression associated with a lower frequency of mutation in the cancer genome [164].
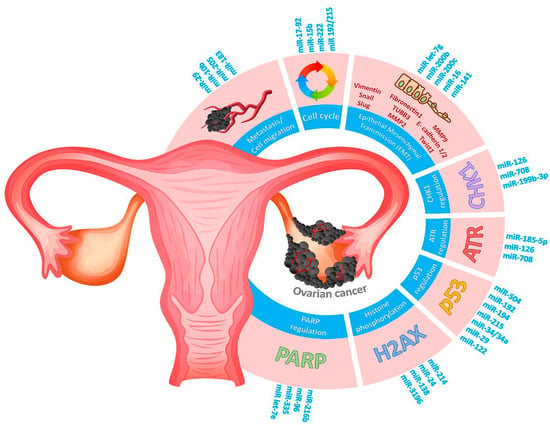
Figure 5.
Schematic diagram representing several hallmarks of cancer contributing to pathogenesis of ovarian cancer. RNA markers involved in DDR signaling (PARP1, histone H2AX, P53, ATR, CHK1 regulation), cell cycle disturbance, metastasis, or epithelial–mesenchymal transmission are highlighted in blue.
Improved understanding of the critical roles of miRNAs in DDR and chemotherapy may therefore provide novel insights with a view to expanding their application as potential tools, biomarkers, or sensitizers in cancer treatment. Promising for increasing the effectiveness of ovarian cancer treatment is the combined therapy with miRNA and chemotherapeutic agents. The role of miRNA in modulating the ovarian cancer cells’ sensitivity to chemotherapeutic agents in multidrug-resistance has been confirmed. It has been revealed that, e.g., decreased resistance to paclitaxel was associated with the upregulation of miR-29b, let-7i, miR-199a, miR-200a, miR-200c, and miR-215, while decreased resistance to platinum agents is related to the upregulation of miR-149, miR-155, miR152, miR-199a, miR200b, miR- 200c, miR-30d, miR-34c, miR-363, miR-497, miR-506, miR-9, and let-7i and to the downregulation of miR-23a and miR-603 [165]. Recent studies demonstrated also that miR-200c significantly enhanced the anticancer efficacy of olaparib in drug-resistant OC cells, which gives hope for optimizing the clinical use of PARPi [89]. Further research is warranted to clarify the correlations among miRNAs, DDR, and ovarian cancer. Continued advancements in miRNA research should allow clarification of the mechanisms’ underlying cancer development, individualization of treatment, and improvement in prognosis for patients with ovarian cancer.
Author Contributions
A.G. and P.G. contributed equally to this work. P.G., A.G., A.M., and A.R. (in order of contribution) wrote this review. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Centre, Poland (Project grant number: Sonata Bis 2019/34/E/NZ7/00056).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ahmed, N.; Kadife, E.; Raza, A.; Short, M.; Jubinsky, P.T.; Kannourakis, G. Ovarian Cancer, Cancer Stem Cells and Current Treatment Strategies: A Potential Role of Magmas in the Current Treatment Methods. Cells 2020, 9, 719. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.J.; Roshid, B.; Pervin, S.; Kabir, S.; Chigurupati, S.; Hasan, M.N. A 35 Year Old Bangladeshi Lady with Hereditary Mucinous Ovarian Cancer, Complicated with Omental Metastasis. Mymensingh Med. J. MMJ 2019, 28, 484–489. [Google Scholar]
- Davidson, B.; Tropé, C.G. Ovarian Cancer: Diagnostic, Biological and Prognostic Aspects. Women’s Health 2014, 10, 519–533. [Google Scholar] [CrossRef]
- Bonadio, R.R.D.C.C.; Fogace, R.N.; Miranda, V.C.; Diz, M.D.P.E. Homologous recombination deficiency in ovarian cancer: A review of its epidemiology and management. Clinics 2018, 73, e450s. [Google Scholar] [CrossRef]
- Pujade-Lauraine, E.; Ledermann, J.A.; Selle, F.; Gebski, V.; Penson, R.T.; Oza, A.M.; Korach, J.; Huzarski, T.; Poveda, A.; Pignata, S.; et al. Olaparib tablets as maintenance therapy in patients with platinum-sensitive, relapsed ovarian cancer and a BRCA1/2 mutation (SOLO2/ENGOT-Ov21): A double-blind, randomised, placebo-controlled, phase 3 trial. Lancet Oncol. 2017, 18, 1274–1284. [Google Scholar] [CrossRef]
- Xu, S.; Fu, G.-B.; Tao, Z.; Ouyang, J.; Kong, F.; Jiang, B.-H.; Wan, X.; Chen, K. MiR-497 decreases cisplatin resistance in ovarian cancer cells by targeting mTOR/P70S6K1. Oncotarget 2015, 6, 26457–26471. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, F.; Wang, X.; Ouyang, G. The roles of microRNAs in the regulation of tumor metastasis. Cell Biosci. 2015, 5, 32. [Google Scholar] [CrossRef]
- Si, W.; Shen, J.; Zheng, H.; Fan, W. The role and mechanisms of action of microRNAs in cancer drug resistance. Clin. Epigenet. 2019, 11, 25. [Google Scholar] [CrossRef]
- Han, W.; Cui, H.; Liang, J.; Su, X. Role of MicroRNA-30c in cancer progression. J. Cancer 2020, 11, 2593–2601. [Google Scholar] [CrossRef]
- Hanlon, K.; Rudin, C.E.; Harries, L.W. Investigating the Targets of MIR-15a and MIR-16-1 in Patients with Chronic Lymphocytic Leukemia (CLL). PLoS ONE 2009, 4, e7169. [Google Scholar] [CrossRef]
- Acunzo, M.; Croce, C.M. Downregulation of miR-15a and miR-16-1 at 13q14 in Chronic Lymphocytic Leukemia. Clin. Chem. 2016, 62, 655–656. [Google Scholar] [CrossRef] [PubMed]
- Rampazzo, E.; Bojnik, E.; Trentin, L.; Bonaldi, L.; Del Bianco, P.; Frezzato, F.; Visentin, A.; Facco, M.; Semenzato, G.; De Rossi, A. Role of miR-15a/miR-16-1 and the TP53 axis in regulating telomerase expression in chronic lymphocytic leukemia. Haematologica 2017, 102, e253–e256. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.P.; Ismail, N.; Zhang, X.; Aguda, B.D.; Lee, E.J.; Yu, L.; Xiao, T.; Schafer, J.; Lee, M.-L.T.; Schmittgen, T.D.; et al. Detection of microRNA Expression in Human Peripheral Blood Microvesicles. PLoS ONE 2008, 3, e3694. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Takahashi, R.; Hiura, Y.; Hirokawa, G.; Fukushima, Y.; Iwai, N. Plasma miR-208 as a Biomarker of Myocardial Injury. Clin. Chem. 2009, 55, 1944–1949. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Taylor, D.D.; Gercel-Taylor, C. MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecol. Oncol. 2008, 110, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xu, S.; Liu, X. MicroRNA profiling of plasma exosomes from patients with ovarian cancer using high-throughput sequencing. Oncol. Lett. 2019, 17, 5601–5607. [Google Scholar] [CrossRef]
- Shapira, I.; Oswald, M.; Lovecchio, J.; Khalili, H.; Menzin, A.; Whyte, J.; Dos Santos, L.; Liang, S.; Bhuiya, T.; Keogh, M.; et al. Circulating biomarkers for detection of ovarian cancer and predicting cancer outcomes. Br. J. Cancer 2014, 110, 976–983. [Google Scholar] [CrossRef]
- Maeda, K.; Sasaki, H.; Ueda, S.; Miyamoto, S.; Terada, S.; Konishi, H.; Kogata, Y.; Ashihara, K.; Fujiwara, S.; Tanaka, Y.; et al. Serum exosomal microRNA-34a as a potential biomarker in epithelial ovarian cancer. J. Ovarian Res. 2020, 13, 47. [Google Scholar] [CrossRef]
- Nakamura, K.; Sawada, K.; Yoshimura, A.; Kinose, Y.; Nakatsuka, E.; Kimura, T. Clinical relevance of circulating cell-free microRNAs in ovarian cancer. Mol. Cancer 2016, 15, 48. [Google Scholar] [CrossRef]
- Staicu, C.E.; Predescu, D.-V.; Rusu, C.M.; Radu, B.M.; Cretoiu, D.; Suciu, N.; Crețoiu, S.M.; Voinea, S.-C. Role of microRNAs as Clinical Cancer Biomarkers for Ovarian Cancer: A Short Overview. Cells 2020, 9, 169. [Google Scholar] [CrossRef]
- Vaksman, O.; Tropé, C.; Davidson, B.; Reich, R. Exosome-derived miRNAs and ovarian carcinoma progression. Carcinogenesis 2014, 35, 2113–2120. [Google Scholar] [CrossRef]
- Zhou, J.; Gong, G.; Tan, H.; Dai, F.; Zhu, X.; Chen, Y.; Wang, J.; Liu, Y.; Chen, P.; Wu, X.; et al. Urinary microRNA-30a-5p is a potential biomarker for ovarian serous adenocarcinoma. Oncol. Rep. 2015, 33, 2915–2923. [Google Scholar] [CrossRef]
- Meng, X.; Müller, V.; Milde-Langosch, K.; Trillsch, F.; Pantel, K.; Schwarzenbach, H. Diagnostic and prognostic relevance of circulating exosomal miR-373, miR-200a, miR-200b and miR-200c in patients with epithelial ovarian cancer. Oncotarget 2016, 7, 16923–16935. [Google Scholar] [CrossRef]
- Kapetanakis, N.-I.; Uzan, C.; Jimenez-Pailhes, A.-S.; Gouy, S.; Bentivegna, E.; Morice, P.; Caron, O.; Gourzones-Dmitriev, C.; Le Teuff, G.; Busson, P. Plasma miR-200b in ovarian carcinoma patients: Distinct pattern of pre/post-treatment variation compared to CA-125 and potential for prediction of progression-free survival. Oncotarget 2015, 6, 36815–36824. [Google Scholar] [CrossRef]
- Chan, S.-H.; Wang, L.-H. Regulation of cancer metastasis by microRNAs. J. Biomed. Sci. 2015, 22, 9. [Google Scholar] [CrossRef]
- Huang, Q.; Gumireddy, K.; Schrier, M.; Le Sage, C.; Nagel, R.; Nair, S.; Egan, D.A.; Li, A.; Huang, G.; Klein-Szanto, A.J.; et al. The microRNAs miR-373 and miR-520c promote tumour invasion and metastasis. Nat. Cell Biol. 2008, 10, 202–210. [Google Scholar] [CrossRef]
- Li, N.; Yang, L.; Sun, Y.; Wu, X. MicroRNA-16 inhibits migration and invasion via regulation of the Wnt/β-catenin signaling pathway in ovarian cancer. Oncol. Lett. 2019, 17, 2631–2638. [Google Scholar] [CrossRef]
- Lezina, L.; Purmessur, N.; Antonov, A.V.; Ivanova, T.; Karpova, E.; Krishan, K.; Ivan, M.; Aksenova, V.; Tentler, D.; Garabadgiu, A.V.; et al. miR-16 and miR-26a target checkpoint kinases Wee1 and Chk1 in response to p53 activation by genotoxic stress. Cell Death Dis. 2013, 4, e953. [Google Scholar] [CrossRef]
- Choi, P.-W.; Wong, K.-K. The Functions of MicroRNA-200 Family in Ovarian Cancer: Beyond Epithelial-Mesenchymal Transition. Int. J. Mol. Sci. 2017, 18, 1207. [Google Scholar] [CrossRef]
- Chung, V.Y.; Tan, T.Z.; Tan, M.; Wong, M.K.; Kuay, K.T.; Yang, Z.; Ye, J.; Muller, J.; Koh, C.M.; Guccione, E.; et al. GRHL2-miR-200-ZEB1 maintains the epithelial status of ovarian cancer through transcriptional regulation and histone modification. Sci. Rep. 2016, 6, 19943. [Google Scholar] [CrossRef]
- Shi, C.; Yang, Y.; Zhang, L.; Yu, J.; Qin, S.; Xu, H.; Gao, Y. MiR-200a-3p promoted the malignant behaviors of ovarian cancer cells through regulating PCDH9. OncoTargets Ther. 2019, 12, 8329–8338. [Google Scholar] [CrossRef]
- Brozovic, A.; Duran, G.E.; Wang, Y.C.; Francisco, E.B.; Sikic, B.I. The miR-200 family differentially regulates sensitivity to paclitaxel and carboplatin in human ovarian carcinoma OVCAR-3 and MES-OV cells. Mol. Oncol. 2015, 9, 1678–1693. [Google Scholar] [CrossRef] [PubMed]
- Jiajie, T.; Yanzhou, Y.; Hoi-Hung, A.C.; Zi-Jiang, C.; Wai-Yee, C. Conserved miR-10 family represses proliferation and induces apoptosis in ovarian granulosa cells. Sci. Rep. 2017, 7, 41304. [Google Scholar] [CrossRef]
- Yu, P.-N.; Yan, M.-D.; Lai, H.-C.; Huang, R.-L.; Chou, Y.-C.; Lin, W.-C.; Yeh, L.-T.; Lin, Y.-W. Downregulation ofmiR-29contributes to cisplatin resistance of ovarian cancer cells. Int. J. Cancer 2013, 134, 542–551. [Google Scholar] [CrossRef] [PubMed]
- To, S.K.Y.; Mak, A.S.C.; Fung, Y.M.E.; Che, C.-M.; Li, S.-S.; Deng, W.; Ru, B.; Zhang, J.; Wong, A.S.T. β-catenin downregulates Dicer to promote ovarian cancer metastasis. Oncogene 2017, 36, 5927–5938. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Cai, J.; Cai, L.; Jia, J.; Xie, L.; Zhu, Y.; Huang, B.; Jin, D.; Wang, Z. Let-7e sensitizes epithelial ovarian cancer to cisplatin through repressing DNA double strand break repair. J. Ovarian Res. 2017, 10, 24. [Google Scholar] [CrossRef]
- Roush, S.; Slack, F.J. The let-7 family of microRNAs. Trends Cell Biol. 2008, 18, 505–516. [Google Scholar] [CrossRef]
- Biamonte, F.; Santamaria, G.; Sacco, A.; Perrone, F.M.; Di Cello, A.; Battaglia, A.M.; Salatino, A.; Di Vito, A.; Aversa, I.; Venturella, R.; et al. MicroRNA let-7g acts as tumor suppressor and predictive biomarker for chemoresistance in human epithelial ovarian cancer. Sci. Rep. 2019, 9, 5668. [Google Scholar] [CrossRef]
- Wang, X.; Cao, L.; Wang, Y.; Wang, X.; Liu, N.; You, Y. Regulation of let-7 and its target oncogenes (Review). Oncol. Lett. 2012, 3, 955–960. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, L.; Li, J.; Zhu, S.; Tai, M.; Mason, C.W.; Chapman, J.A.; Reynolds, E.A.; Weiner, C.P.; Zhou, H.H. MicroRNA-205 promotes cell invasion by repressing TCF21 in human ovarian cancer. J. Ovarian Res. 2017, 10, 33. [Google Scholar] [CrossRef]
- Zhu, D.; Huang, X.; Liang, F.; Zhao, L. LncRNA miR503HG interacts with miR-31-5p through multiple ways to regulate cancer cell invasion and migration in ovarian cancer. J. Ovarian Res. 2020, 13, 3. [Google Scholar] [CrossRef]
- Ibrahim, F.F.; Jamal, R.; Syafruddin, S.E.; Ab Mutalib, N.S.; Saidin, S.; MdZin, R.R.; Mollah, M.M.H.; Mokhtar, N.M. MicroRNA-200c and microRNA-31 regulate proliferation, colony formation, migration and invasion in serous ovarian cancer. J. Ovarian Res. 2015, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Xiang, G.; Cheng, Y. MiR-126-3p inhibits ovarian cancer proliferation and invasion via targeting PLXNB2. Reprod. Biol. 2018, 18, 218–224. [Google Scholar] [CrossRef]
- Xia, X.Y.; Yu, Y.J.; Ye, F.; Peng, G.Y.; Li, Y.J.; Zhou, X.M. MicroRNA-506-3p inhibits proliferation and promotes apoptosis in ovarian cancer cell via targeting SIRT1/AKT/FOXO3a signaling pathway. Neoplasma 2020, 67, 344–353. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, C.; Zhou, B.; Jiang, D. miR-183 modulated cell proliferation and apoptosis in ovarian cancer through the TGF-β/Smad4 signaling pathway. Int. J. Mol. Med. 2019, 43, 1734–1746. [Google Scholar] [CrossRef]
- Li, T.; Li, Y.; Gan, Y.; Tian, R.; Wu, Q.; Shu, G.; Yin, G. Methylation-mediated repression of MiR-424/503 cluster promotes proliferation and migration of ovarian cancer cells through targeting the hub gene KIF23. Cell Cycle 2019, 18, 1601–1618. [Google Scholar] [CrossRef]
- Liao, Y.; Deng, Y.; Liu, J.; Ye, Z.; You, Z.; Yao, S.; He, S. MiR-760 overexpression promotes proliferation in ovarian cancer by downregulation of PHLPP2 expression. Gynecol. Oncol. 2016, 143, 655–663. [Google Scholar] [CrossRef]
- Feng, Y.; Hang, W.; Sang, Z.; Li, S.; Xu, W.; Miao, Y.; Xi, X.; Huang, Q. Identification of exosomal and non-exosomal microRNAs associated with the drug resistance of ovarian cancer. Mol. Med. Rep. 2019, 19, 3376–3392. [Google Scholar] [CrossRef]
- Lv, Y.L.F.; Liu, P.S. MiR-151 promotes ovarian cancer through activation of akt/mTOR signaling pathway by decreasing RhoGDIA. Int. J. Clin. Exp. Pathol. 2016, 9, 11222–11229. [Google Scholar]
- Liu, X.; Yao, B.; Wu, Z. miRNA-199a-5p suppresses proliferation and invasion by directly targeting NF-κB1 in human ovarian cancer cells. Oncol. Lett. 2018, 16, 4543–4550. [Google Scholar] [CrossRef]
- Meng, F.; Henson, R.; Wehbe–Janek, H.; Ghoshal, K.; Jacob, S.T.; Patel, T. MicroRNA-21 Regulates Expression of the PTEN Tumor Suppressor Gene in Human Hepatocellular Cancer. Gastroenterology 2007, 133, 647–658. [Google Scholar] [CrossRef]
- Misso, G.; Di Martino, M.T.; De Rosa, G.; Farooqi, A.A.; Lombardi, A.; Campani, V.; Zarone, M.R.; Gullà, A.; Tagliaferri, P.; Tassone, P.; et al. Mir-34: A New Weapon Against Cancer? Mol. Ther. Nucleic Acids 2014, 3, e195. [Google Scholar] [CrossRef] [PubMed]
- Frixa, T.; Donzelli, S.; Blandino, G. Oncogenic MicroRNAs: Key Players in Malignant Transformation. Cancers 2015, 7, 2466–2485. [Google Scholar] [CrossRef]
- Kumar, D.; Gorain, M.; Kundu, G.; Kundu, G.C. Therapeutic implications of cellular and molecular biology of cancer stem cells in melanoma. Mol. Cancer 2017, 16, 7. [Google Scholar] [CrossRef]
- Yeh, Y.-M.; Chuang, C.-M.; Chao, K.-C.; Wang, L.-H. MicroRNA-138 suppresses ovarian cancer cell invasion and metastasis by targeting SOX4 and HIF-1α. Int. J. Cancer 2013, 133, 867–878. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; An, Q.; Niu, B.; Lu, X.; Zhang, N.; Cao, X. Role of miR-221/222 in Tumor Development and the Underlying Mechanism. J. Oncol. 2019, 2019, 7252013. [Google Scholar] [CrossRef]
- Amini-Farsani, Z.; Sangtarash, M.H.; Shamsara, M.; Teimori, H. MiR-221/222 promote chemoresistance to cisplatin in ovarian cancer cells by targeting PTEN/PI3K/AKT signaling pathway. Cytotechnology 2018, 70, 203–213. [Google Scholar] [CrossRef]
- Sun, C.; Cao, W.; Qiu, C.; Li, C.; Dongol, S.; Zhang, Z.; Dong, R.; Song, K.; Yang, X.; Zhang, Q.; et al. MiR-509-3 augments the synthetic lethality of PARPi by regulating HR repair in PDX model of HGSOC. J. Hematol. Oncol. 2020, 13, 9. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Huang, H.; Li, Y.; Li, L.; Hou, W.; You, Z. Overexpression of miRNA-221 promotes cell proliferation by targeting the apoptotic protease activating factor-1 and indicates a poor prognosis in ovarian cancer. Int. J. Oncol. 2017, 50, 1087–1096. [Google Scholar] [CrossRef]
- Liu, R.; Guo, H.; Lu, S. MiR-335-5p restores cisplatin sensitivity in ovarian cancer cells through targeting BCL2L2. Cancer Med. 2018, 7, 4598–4609. [Google Scholar] [CrossRef]
- Guan, R.; Cai, S.; Sun, M.; Xu, M. Upregulation of miR-520b promotes ovarian cancer growth. Oncol. Lett. 2017, 14, 3155–3161. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Lü, M.; Yao, G.; Tian, H.; Lian, J.; Liu, L.; Liang, M.; Wang, Y.; Sun, F. Transactivation of microRNA-383 by Steroidogenic Factor-1 Promotes Estradiol Release from Mouse Ovarian Granulosa Cells by Targeting RBMS1. Mol. Endocrinol. 2012, 26, 1129–1143. [Google Scholar] [CrossRef]
- Nakayama, I.; Shibazaki, M.; Yashima-Abo, A.; Miura, F.; Sugiyama, T.; Masuda, T.; Maesawa, C. Loss of HOXD10 expression induced by upregulation of miR-10b accelerates the migration and invasion activities of ovarian cancer cells. Int. J. Oncol. 2013, 43, 63–71. [Google Scholar] [CrossRef]
- Wang, J.; He, J.; Su, F.; Ding, N.; Hu, W.; Yao, B.; Wang, W.; Zhou, G. Repression of ATR pathway by miR-185 enhances radiation-induced apoptosis and proliferation inhibition. Cell Death Dis. 2013, 4, e699. [Google Scholar] [CrossRef]
- Boele, J.; Persson, H.; Shin, J.W.; Ishizu, Y.; Newie, I.S.; Søkilde, R.; Hawkins, S.M.; Coarfa, C.; Ikeda, K.; Takayama, K.-I.; et al. PAPD5-mediated 3’ adenylation and subsequent degradation of miR-21 is disrupted in proliferative disease. Proc. Natl. Acad. Sci. USA 2014, 111, 11467–11472. [Google Scholar] [CrossRef]
- Bao, J.; Yu, Y.; Chen, J.; He, Y.; Chen, X.; Ren, Z.; Xue, C.; Liu, L.; Hu, Q.; Li, J.; et al. MiR-126 negatively regulates PLK-4 to impact the development of hepatocellular carcinoma via ATR/CHEK1 pathway. Cell Death Dis. 2018, 9, 1045. [Google Scholar] [CrossRef]
- Zou, X.; Zhu, C.; Zhang, L.; Zhang, Y.; Fu, F.; Chen, Y.; Zhou, J. MicroRNA-708 Suppresses Cell Proliferation and Enhances Chemosensitivity of Cervical Cancer Cells to cDDP by Negatively Targeting Timeless. OncoTargets Ther. 2020, 13, 225–235. [Google Scholar] [CrossRef]
- Choi, Y.E.; Meghani, K.; Brault, M.-E.; Leclerc, L.; He, Y.; Day, T.A.; Elias, K.M.; Drapkin, R.; Weinstock, D.M.; Dao, F.; et al. Platinum and PARP Inhibitor Resistance Due to Overexpression of MicroRNA-622 in BRCA1-Mutant Ovarian Cancer. Cell Rep. 2016, 14, 429–439. [Google Scholar] [CrossRef]
- Marchini, S.; Cavalieri, D.; Fruscio, R.; Calura, E.; Garavaglia, D.; Nerini, I.F.; Mangioni, C.; Cattoretti, G.; Clivio, L.; Beltrame, L.; et al. Association between miR-200c and the survival of patients with stage I epithelial ovarian cancer: A retrospective study of two independent tumour tissue collections. Lancet Oncol. 2011, 12, 273–285. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, X.; Huang, Y.; Zhang, Q.; Zhou, J.; Zhang, X.; Wang, X. miR-200b and miR-200c co-contribute to the cisplatin sensitivity of ovarian cancer cells by targeting DNA methyltransferases. Oncol. Lett. 2018, 17, 1453–1460. [Google Scholar] [CrossRef]
- Cha, S.Y.; Choi, Y.H.; Hwang, S.; Jeong, J.-Y.; An, H.J. Clinical Impact of microRNAs Associated with Cancer Stem Cells as a Prognostic Factor in Ovarian Carcinoma. J. Cancer 2017, 8, 3538–3547. [Google Scholar] [CrossRef]
- Kan, C.W.S.; Hahn, M.A.; Gard, G.B.; Maidens, J.; Huh, J.Y.; Marsh, D.J.; Howell, V.M. Elevated levels of circulating microRNA-200 family members correlate with serous epithelial ovarian cancer. BMC Cancer 2012, 12, 627. [Google Scholar] [CrossRef]
- Iorio, M.V.; Visone, R.; Di Leva, G.; Donati, V.; Petrocca, F.; Casalini, P.; Taccioli, C.; Volinia, S.; Liu, C.-G.; Alder, H.; et al. MicroRNA Signatures in Human Ovarian Cancer. Cancer Res. 2007, 67, 8699–8707. [Google Scholar] [CrossRef]
- Nam, E.J.; Yoon, H.; Kim, S.W.; Kim, H.; Kim, Y.T.; Kim, J.H.; Kim, S. MicroRNA Expression Profiles in Serous Ovarian Carcinoma. Clin. Cancer Res. 2008, 14, 2690–2695. [Google Scholar] [CrossRef]
- Calura, E.; Fruscio, R.; Paracchini, L.; Bignotti, E.; Ravaggi, A.; Martini, P.; Sales, G.; Beltrame, L.; Clivio, L.; Ceppi, L.; et al. miRNA Landscape in Stage I Epithelial Ovarian Cancer Defines the Histotype Specificities. Clin. Cancer Res. 2013, 19, 4114–4123. [Google Scholar] [CrossRef] [PubMed]
- Elgaaen, B.V.; Olstad, O.K.; Haug, K.B.F.; Brusletto, B.; Sandvik, L.; Staff, A.C.; Gautvik, K.M.; Davidson, B. Global miRNA expression analysis of serous and clear cell ovarian carcinomas identifies differentially expressed miRNAs including miR-200c-3p as a prognostic marker. BMC Cancer 2014, 14, 80. [Google Scholar] [CrossRef]
- Yeom, K.-H.; Lee, Y.; Han, J.; Suh, M.R.; Kim, V.N. Characterization of DGCR8/Pasha, the essential cofactor for Drosha in primary miRNA processing. Nucleic Acids Res. 2006, 34, 4622–4629. [Google Scholar] [CrossRef]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef]
- Palmero, E.; De Campos, S.G.P.; Campos, M.; De Souza, N.C.N.; Guerreiro, I.D.C.; Carvalho, A.L.; Marques, M.M.C. Mechanisms and role of microRNA deregulation in cancer onset and progression. Genet. Mol. Biol. 2011, 34, 363–370. [Google Scholar] [CrossRef]
- Yin, F.; Liu, X.; Li, D.; Wang, Q.; Zhang, W.; Li, L. Tumor suppressor genes associated with drug resistance in ovarian cancer (Review). Oncol. Rep. 2013, 30, 3–10. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cobb, G.; Anderson, T.A. microRNAs as oncogenes and tumor suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef]
- Lu, L.; Schwartz, P.; Scarampi, L.; Rutherford, T.; Canuto, E.M.; Yu, H.; Katsaros, D. MicroRNA let-7a: A potential marker for selection of paclitaxel in ovarian cancer management. Gynecol. Oncol. 2011, 122, 366–371. [Google Scholar] [CrossRef]
- Wang, Y.; Lei, X.; Gao, C.; Xue, Y.; Li, X.; Wang, H.; Feng, Y. MiR-506-3p suppresses the proliferation of ovarian cancer cells by negatively regulating the expression of MTMR6. J. Biosci. 2019, 44, 126. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.; Mitra, R.; Maulik, U.; Zhang, M.Q. Development of the human cancer microRNA network. Silence 2010, 1, 6. [Google Scholar] [CrossRef] [PubMed]
- Jansson, M.D.; Lund, A.H. MicroRNA and cancer. Mol. Oncol. 2012, 6, 590–610. [Google Scholar] [CrossRef] [PubMed]
- Berchuck, A.; Kohler, M.F.; Bast, R.C. Oncogenes in Ovarian Cancer. Hematol. Clin. N. Am. 1992, 6, 813–827. [Google Scholar] [CrossRef]
- Vescarelli, E.; Gerini, G.; Megiorni, F.; Anastasiadou, E.; Pontecorvi, P.; Solito, L.; De Vitis, C.; Camero, S.; Marchetti, C.; Mancini, R.; et al. MiR-200c sensitizes Olaparib-resistant ovarian cancer cells by targeting Neuropilin 1. J. Exp. Clin. Cancer Res. 2020, 39, 3. [Google Scholar] [CrossRef]
- Duran, G.E.; Wang, Y.C.; Moisan, F.; Francisco, E.B.; Sikic, B.I. Decreased levels of baseline and drug-induced tubulin polymerisation are hallmarks of resistance to taxanes in ovarian cancer cells and are associated with epithelial-to-mesenchymal transition. Br. J. Cancer 2017, 116, 1318–1328. [Google Scholar] [CrossRef]
- Huang, R.-X.; Zhou, P.-K. DNA damage response signaling pathways and targets for radiotherapy sensitization in cancer. Signal Transduct. Target. Ther. 2020, 5, 60. [Google Scholar] [CrossRef]
- Taylor, R.; Cullen, S.P.; Martin, S. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef]
- Yousefi, B.; Rahmati, M.; Ahmadi, Y. The roles of p53R2 in cancer progression based on the new function of mutant p53 and cytoplasmic p21. Life Sci. 2014, 99, 14–17. [Google Scholar] [CrossRef]
- Fukuda, T.; Yamagata, K.; Fujiyama, S.; Matsumoto, T.; Koshida, I.; Yoshimura, K.; Mihara, M.; Naitou, M.; Endoh, H.; Nakamura, T.; et al. DEAD-box RNA helicase subunits of the Drosha complex are required for processing of rRNA and a subset of microRNAs. Nat. Cell Biol. 2007, 9, 604–611. [Google Scholar] [CrossRef]
- Gralewska, P.; Gajek, A.; Marczak, A.; Rogalska, A. Participation of the ATR/CHK1 pathway in replicative stress targeted therapy of high-grade ovarian cancer. J. Hematol. Oncol. 2020, 13, 39. [Google Scholar] [CrossRef]
- Saravi, S.; Katsuta, E.; Jeyaneethi, J.; Amin, H.A.; Kaspar, M.; Takabe, K.; Pados, G.; Drenos, F.; Hall, M.; Karteris, E. H2A Histone Family Member X (H2AX) Is Upregulated in Ovarian Cancer and Demonstrates Utility as a Prognostic Biomarker in Terms of Overall Survival. J. Clin. Med. 2020, 9, 2844. [Google Scholar] [CrossRef] [PubMed]
- Medina, P.P.; Slack, F.J. MicroRNAs and cancer: An overview. Cell Cycle 2008, 7, 2485–2492. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, J.-W.; Li, M.; Cavenee, W.K.; Mitchell, P.; Zhou, X.; Tewari, M.; Furnari, F.B.; Taniguchi, T. MicroRNA-138 Modulates DNA Damage Response by Repressing Histone H2AX Expression. Mol. Cancer Res. 2011, 9, 1100–1111. [Google Scholar] [CrossRef]
- Georges, S.A.; Biery, M.C.; Kim, S.-Y.; Schelter, J.M.; Guo, J.; Chang, A.N.; Jackson, A.L.; Carleton, M.O.; Linsley, P.S.; Cleary, M.A.; et al. Coordinated Regulation of Cell Cycle Transcripts by p53-Inducible microRNAs, miR-192 and miR-215. Cancer Res. 2008, 68, 10105–10112. [Google Scholar] [CrossRef]
- Agostini, A.; Brunetti, M.; Davidson, B.; Tropé, C.G.; Eriksson, A.G.Z.; Heim, S.; Panagopoulos, I.; Micci, F. The microRNA miR-192/215 family is upregulated in mucinous ovarian carcinomas. Sci. Rep. 2018, 8, 11069. [Google Scholar] [CrossRef]
- Stucki, M.; Clapperton, J.A.; Mohammad, D.; Yaffe, M.B.; Smerdon, S.J.; Jackson, S.P. MDC1 Directly Binds Phosphorylated Histone H2AX to Regulate Cellular Responses to DNA Double-Strand Breaks. Cell 2005, 123, 1213–1226. [Google Scholar] [CrossRef]
- Xiao, A.; Li, H.; Shechter, D.; Ahn, S.H.; Fabrizio, L.A.; Erdjument-Bromage, H.; Ishibe-Murakami, S.; Wang, B.; Tempst, P.; Hofmann, K.; et al. WSTF regulates the H2A.X DNA damage response via a novel tyrosine kinase activity. Nat. Cell Biol. 2008, 457, 57–62. [Google Scholar] [CrossRef]
- Coster, G.; Goldberg, M. The cellular response to DNA damage: A focus on MDC1 and its interacting proteins. Nucleus 2010, 1, 166–178. [Google Scholar] [CrossRef]
- Chapman, R.; Jackson, S.P. Phospho-dependent interactions between NBS1 and MDC1 mediate chromatin retention of the MRN complex at sites of DNA damage. EMBO Rep. 2008, 9, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Lukas, J.; Lukas, C.; Bartek, J. More than just a focus: The chromatin response to DNA damage and its role in genome integrity maintenance. Nat. Cell Biol. 2011, 13, 1161–1169. [Google Scholar] [CrossRef]
- Nakada, S. Opposing roles of RNF8/RNF168 and deubiquitinating enzymes in ubiquitination-dependent DNA double-strand break response signaling and DNA-repair pathway choice. J. Radiat. Res. 2016, 57, i33–i40. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Qin, B.; Lou, Z. Ubiquitin and ubiquitin-like molecules in DNA double strand break repair. Cell Biosci. 2020, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Price, B.D.; D’Andrea, A.D. Chromatin Remodeling at DNA Double-Strand Breaks. Cell 2013, 152, 1344–1354. [Google Scholar] [CrossRef]
- Wang, Z.; Yin, H.; Zhang, Y.; Feng, Y.; Yan, Z.; Jiang, X.; Bukhari, I.; Iqbal, F.; Cooke, H.J.; Shi, Q. miR-214-mediated downregulation of RNF8 induces chromosomal instability in ovarian cancer cells. Cell Cycle 2014, 13, 3519–3528. [Google Scholar] [CrossRef][Green Version]
- Wan, G.; Mathur, R.; Hu, X.; Zhang, X.; Lu, X. miRNA response to DNA damage. Trends Biochem. Sci. 2011, 36, 478–484. [Google Scholar] [CrossRef]
- Lal, A.; Pan, Y.; Navarro, F.; Dykxhoorn, D.M.; Moreau, L.; Meire, E.; Bentwich, Z.; Lieberman, J.; Chowdhury, D. miR-24–mediated downregulation of H2AX suppresses DNA repair in terminally differentiated blood cells. Nat. Struct. Mol. Biol. 2009, 16, 492–498. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, L.; Duan, L.; Lu, C. MicroRNA-3196 is inhibited by H2AX phosphorylation and attenuates lung cancer cell apoptosis by downregulating PUMA. Oncotarget 2016, 7, 77764–77776. [Google Scholar] [CrossRef]
- Wu, J.; Sun, Y.; Zhang, P.-Y.; Qian, M.; Zhang, H.; Chen, X.; Ma, D.; Xu, Y.; Chen, X.; Tang, K.-F. The Fra-1–miR-134–SDS22 feedback loop amplifies ERK/JNK signaling and reduces chemosensitivity in ovarian cancer cells. Cell Death Dis. 2016, 7, e2384. [Google Scholar] [CrossRef]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA damage, repair, and mutagenesis. Environ. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nat. Cell Biol. 2009, 461, 1071–1078. [Google Scholar] [CrossRef]
- Dietlein, F.; Thelen, L.; Reinhardt, H.C. Cancer-specific defects in DNA repair pathways as targets for personalized therapeutic approaches. Trends Genet. 2014, 30, 326–339. [Google Scholar] [CrossRef] [PubMed]
- Alhmoud, J.F.; Woolley, J.F.; Al Moustafa, A.-E.; Malki, M.I. DNA Damage/Repair Management in Cancers. Cancers 2020, 12, 1050. [Google Scholar] [CrossRef]
- Van Gent, D.C.; Hoeijmakers, J.H.J.; Kanaar, R. Chromosomal stability and the DNA double-stranded break connection. Nat. Rev. Genet. 2001, 2, 196–206. [Google Scholar] [CrossRef]
- Lieber, M.R. The Mechanism of Double-Strand DNA Break Repair by the Nonhomologous DNA End-Joining Pathway. Annu. Rev. Biochem. 2010, 79, 181–211. [Google Scholar] [CrossRef]
- Brandsma, I.; van Gent, D.C. Pathway choice in DNA double strand break repair: Observations of a balancing act. Genome Integr. 2012, 3, 9. [Google Scholar] [CrossRef]
- Zhao, X.; Wei, C.; Li, J.; Xing, P.; Li, J.; Zheng, S.; Chen, X. Cell cycle-dependent control of homologous recombination. Acta Biochim. Biophys. Sin. 2017, 49, 655–668. [Google Scholar] [CrossRef]
- Northall, S.J.; Ivančić-Baće, I.; Soultanas, P.; Bolt, E.L. Remodeling and Control of Homologous Recombination by DNA Helicases and Translocases that Target Recombinases and Synapsis. Genes 2016, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Kolinjivadi, A.M.; Sannino, V.; De Antoni, A.; Técher, H.; Baldi, G.; Costanzo, V. Moonlighting at replication forks—A new life for homologous recombination proteins BRCA1, BRCA2 and RAD51. FEBS Lett. 2017, 591, 1083–1100. [Google Scholar] [CrossRef] [PubMed]
- Moskwa, P.; Buffa, F.M.; Pan, Y.; Panchakshari, R.; Gottipati, P.; Muschel, R.J.; Beech, J.; Kulshrestha, R.; Abdelmohsen, K.; Weinstock, D.M.; et al. miR-182-Mediated Downregulation of BRCA1 Impacts DNA Repair and Sensitivity to PARP Inhibitors. Mol. Cell 2011, 41, 210–220. [Google Scholar] [CrossRef]
- Pannunzio, N.R.; Watanabe, G.; Lieber, M.R. Nonhomologous DNA end-joining for repair of DNA double-strand breaks. J. Biol. Chem. 2018, 293, 10512–10523. [Google Scholar] [CrossRef] [PubMed]
- Chiruvella, K.K.; Liang, Z.; Wilson, T.E. Repair of Double-Strand Breaks by End Joining. Cold Spring Harb. Perspect. Biol. 2013, 5, a012757. [Google Scholar] [CrossRef]
- Yan, D.; Ng, W.L.; Zhang, X.; Wang, P.; Zhang, Z.; Mo, Y.-Y.; Mao, H.; Hao, C.; Olson, J.J.; Curran, W.J.; et al. Targeting DNA-PKcs and ATM with miR-101 Sensitizes Tumors to Radiation. PLoS ONE 2010, 5, e11397. [Google Scholar] [CrossRef]
- Chakraborty, A.; Uechi, T.; Kenmochi, N. Guarding the ‘translation apparatus’: Defective ribosome biogenesis and the p53 signaling pathway. Wiley Interdiscip. Rev. RNA 2011, 2, 507–522. [Google Scholar] [CrossRef]
- Dancy, B.M.; Cole, P.A. Protein Lysine Acetylation by p300/CBP. Chem. Rev. 2015, 115, 2419–2452. [Google Scholar] [CrossRef]
- Hasty, P.; Christy, B.A. p53 as an intervention target for cancer and aging. Pathobiol. Aging Age-Relat. Dis. 2013, 3. [Google Scholar] [CrossRef]
- Chen, X.; Che, T. Roles of MicroRNA in DNA Damage and Repair. In DNA Repair; Kruman, I., Ed.; InTech; ISBN 978-953-307-697-3. Available online: http://www.intechopen.com/books/dnarepair/roles-of-microrna-in-dna-damage-and-repair (accessed on 13 April 2021).
- Jones, M.F.; Lal, A. MicroRNAs, wild-type and mutant p53: More questions than answers. RNA Biol. 2012, 9, 781–791. [Google Scholar] [CrossRef]
- Yamakuchi, M.; Ferlito, M.; Lowenstein, C.J. miR-34a repression of SIRT1 regulates apoptosis. Proc. Natl. Acad. Sci USA 2008, 105, 13421–13426. [Google Scholar] [CrossRef]
- Park, S.-Y.; Lee, J.H.; Ha, M.; Nam, J.-W.; Kim, V.N. miR-29 miRNAs activate p53 by targeting p85α and CDC42. Nat. Struct. Mol. Biol. 2008, 16, 23–29. [Google Scholar] [CrossRef]
- Fornari, F.; Gramantieri, L.; Ferracin, M.; Veronese, A.; Sabbioni, S.; A Calin, G.; Grazi, G.L.; Giovannini, C.; Croce, C.M.; Bolondi, L.; et al. MiR-221 controls CDKN1C/p57 and CDKN1B/p27 expression in human hepatocellular carcinoma. Oncogene 2008, 27, 5651–5661. [Google Scholar] [CrossRef] [PubMed]
- Fornari, F.; Gramantieri, L.; Giovannini, C.; Veronese, A.; Ferracin, M.; Sabbioni, S.; Calin, G.A.; Grazi, G.L.; Croce, C.M.; Tavolari, S.; et al. MiR-122/Cyclin G1 Interaction Modulates p53 Activity and Affects Doxorubicin Sensitivity of Human Hepatocarcinoma Cells. Cancer Res. 2009, 69, 5761–5767. [Google Scholar] [CrossRef]
- Corney, D.C.; Flesken-Nikitin, A.; Godwin, A.K.; Wang, W.; Nikitin, A.Y. MicroRNA-34b and MicroRNA-34c Are Targets of p53 and Cooperate in Control of Cell Proliferation and Adhesion-Independent Growth. Cancer Res. 2007, 67, 8433–8438. [Google Scholar] [CrossRef]
- Raver-Shapira, N.; Marciano, E.; Meiri, E.; Spector, Y.; Rosenfeld, N.; Moskovits, N.; Bentwich, Z.; Oren, M. Transcriptional Activation of miR-34a Contributes to p53-Mediated Apoptosis. Mol. Cell 2007, 26, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.-C.; Wentzel, E.A.; Kent, O.A.; Ramachandran, K.; Mullendore, M.; Lee, K.H.; Feldmann, G.; Yamakuchi, M.; Ferlito, M.; Lowenstein, C.J.; et al. Transactivation of miR-34a by p53 Broadly Influences Gene Expression and Promotes Apoptosis. Mol. Cell 2007, 26, 745–752. [Google Scholar] [CrossRef]
- Asslaber, D.; Piñón, J.D.; Seyfried, I.; Desch, P.; Stöcher, M.; Tinhofer, I.; Egle, A.; Merkel, O.; Greil, R. microRNA-34a expression correlates with MDM2 SNP309 polymorphism and treatment-free survival in chronic lymphocytic leukemia. Blood 2010, 115, 4191–4197. [Google Scholar] [CrossRef]
- Bryant, H.E.; Schultz, N.; Thomas, H.D.; Parker, K.M.; Flower, D.; Lopez, E.; Kyle, S.; Meuth, M.; Curtin, N.J.; Helleday, T. Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) polymerase. Nat. Cell Biol. 2005, 434, 913–917. [Google Scholar] [CrossRef]
- Meghani, K.; Fuchs, W.; Detappe, A.; Drané, P.; Gogola, E.; Rottenberg, S.; Jonkers, J.; Matulonis, U.; Swisher, E.M.; Konstantinopoulos, P.A.; et al. Multifaceted Impact of MicroRNA 493-5p on Genome-Stabilizing Pathways Induces Platinum and PARP Inhibitor Resistance in BRCA2-Mutated Carcinomas. Cell Rep. 2018, 23, 100–111. [Google Scholar] [CrossRef]
- Neijenhuis, S.; Bajrami, I.; Miller, R.; Lord, C.J.; Ashworth, A. Identification of miRNA modulators to PARP inhibitor response. DNA Repair 2013, 12, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, J.-W.; Calses, P.; Kemp, C.J.; Taniguchi, T. MiR-96 Downregulates REV1 and RAD51 to Promote Cellular Sensitivity to Cisplatin and PARP Inhibition. Cancer Res. 2012, 72, 4037–4046. [Google Scholar] [CrossRef]
- Liu, Y.; Niu, Z.; Lin, X.; Tian, Y. MiR-216b increases cisplatin sensitivity in ovarian cancer cells by targeting PARP1. Cancer Gene Ther. 2017, 24, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Kaur, S.; Volinia, S.; Greshock, J.; Lassus, H.; Hasegawa, K.; Liang, S.; Leminen, A.; Deng, S.; Smith, L.; et al. MicroRNA Microarray Identifies Let-7i as a Novel Biomarker and Therapeutic Target in Human Epithelial Ovarian Cancer. Cancer Res. 2008, 68, 10307–10314. [Google Scholar] [CrossRef] [PubMed]
- Cortez, D.; Guntuku, S.; Qin, J.; Elledge, S.J. ATR and ATRIP: Partners in Checkpoint Signaling. Science 2001, 294, 1713–1716. [Google Scholar] [CrossRef]
- Cimprich, K.A.; Cortez, D. ATR: An essential regulator of genome integrity. Nat. Rev. Mol. Cell Biol. 2008, 9, 616–627. [Google Scholar] [CrossRef]
- Feng, Y.-J.; Tian, R.; You, A.-G.; Wang, J.; Wu, Y.-J.; Wang, W.; Zhou, A.-Y.; Wei, X.-L.; He, Q.-D.; Feng, X.; et al. Expression and significance of DNA methyltransferase in sera of patients with lung cancer. Zhonghua Yi Xue Za Zhi 2013, 93, 3822–3825. [Google Scholar]
- Bartek, J.; Lukas, J. Chk1 and Chk2 kinases in checkpoint control and cancer. Cancer Cell 2003, 3, 421–429. [Google Scholar] [CrossRef]
- Buisson, R.; Boisvert, J.L.; Benes, C.H.; Zou, L. Distinct but Concerted Roles of ATR, DNA-PK, and Chk1 in Countering Replication Stress during S Phase. Mol. Cell 2015, 59, 1011–1024. [Google Scholar] [CrossRef]
- Xu, J.; Li, Y.; Wang, F.; Wang, X.; Cheng, B.; Ye, F.; Xie, X.; Zhou, C.; Lu, W. Suppressed miR-424 expression via upregulation of target gene Chk1 contributes to the progression of cervical cancer. Oncogene 2012, 32, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yao, L.; Liu, F.; Hong, J.; Chen, L.; Zhang, B.; Zhang, W. Characterization of microRNA expression in serous ovarian carcinoma. Int. J. Mol. Med. 2014, 34, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Qu, J.; Xu, F.; Guo, Y.; Wang, Y.; Yu, H.; Qian, B. MiR-195 suppresses non-small cell lung cancer by targeting CHEK1. Oncotarget 2015, 6, 9445–9456. [Google Scholar] [CrossRef]
- Su, B.-B.; Zhou, S.-W.; Gan, C.-B.; Zhang, X.-N. MiR-330-5p regulates tyrosinase and PDIA3 expression and suppresses cell proliferation and invasion in cutaneous malignant melanoma. J. Surg. Res. 2016, 203, 434–440. [Google Scholar] [CrossRef]
- Lee, K.-H.; Chen, Y.-L.; Yeh, S.-D.; Hsiao, M.; Lin, J.-T.; Goan, Y.-G.; Lu, P.-J. MicroRNA-330 acts as tumor suppressor and induces apoptosis of prostate cancer cells through E2F1-mediated suppression of Akt phosphorylation. Oncogene 2009, 28, 3360–3370. [Google Scholar] [CrossRef]
- Bibby, B.A.S.; Reynolds, J.V.; Maher, S.G. MicroRNA-330-5p as a Putative Modulator of Neoadjuvant Chemoradiotherapy Sensitivity in Oesophageal Adenocarcinoma. PLoS ONE 2015, 10, e0134180. [Google Scholar] [CrossRef]
- Kong, R.; Liu, W.; Guo, Y.; Feng, J.; Cheng, C.; Zhang, X.; Ma, Y.; Li, S.; Jiang, J.; Zhang, J.; et al. Inhibition of NOB1 by microRNA-330-5p overexpression represses cell growth of non-small cell lung cancer. Oncol. Rep. 2017, 38, 2572–2580. [Google Scholar] [CrossRef]
- Shao, S.; Tian, J.; Zhang, H.; Wang, S. LncRNA myocardial infarction-associated transcript promotes cell proliferation and inhibits cell apoptosis by targeting miR-330-5p in epithelial ovarian cancer cells. Arch. Med. Sci. 2018, 14, 1263–1270. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Feng, P.-C.; Ke, X.-F.; Kuang, H.-L.; Pan, L.-L.; Ye, Q.; Wu, J.-B. Silencing Long Non-coding RNA LINC01224 Inhibits Hepatocellular Carcinoma Progression via MicroRNA-330-5p-Induced Inhibition of CHEK1. Mol. Ther. Nucleic Acids 2020, 19, 482–497. [Google Scholar] [CrossRef]
- Xie, Y.; Wei, R.-R.; Huang, G.-L.; Zhang, M.-Y.; Yuan, Y.-F.; Wang, H.-Y. Checkpoint kinase 1 is negatively regulated by miR-497 in hepatocellular carcinoma. Med. Oncol. 2014, 31, 844. [Google Scholar] [CrossRef]
- Wei, L.; He, Y.; Bi, S.; Li, X.; Zhang, J.; Zhang, S. miRNA-199b-3p suppresses growth and progression of ovarian cancer via the CHK1/E-cadherin/EMT signaling pathway by targeting ZEB1. Oncol. Rep. 2020, 45, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; James, J.; Annunziata, C.M. Topotecan synergizes with CHEK1 (CHK1) inhibitor to induce apoptosis in ovarian cancer cells. BMC Cancer 2015, 15, 196. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wang, G.; Zhang, X.; Wu, D.; Yang, L.; Wang, G.; Hao, D. The expression of miRNAs is associated with tumour genome instability and predicts the outcome of ovarian cancer patients treated with platinum agents. Sci. Rep. 2017, 7, 14736. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.; Roela, R.A.; Lopez, R.V.M.; Estevez-Diz, M.D.P. The prognostic role of microRNA in epithelial ovarian cancer: A systematic review of literature with an overall survival meta-analysis. Oncotarget 2020, 11, 1085–1095. [Google Scholar] [CrossRef] [PubMed][Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).