
A Comprehensive Review on Solitary Fibrous Tumor: New Insights for New Horizons



Abstract
:Simple Summary
Abstract
1. Introduction
2. Clinical Aspects
2.1. Paraneoplastic Syndromes
2.2. Radiological Characteristics
2.3. Risk Classification
3. Pathogenesis and Pathology
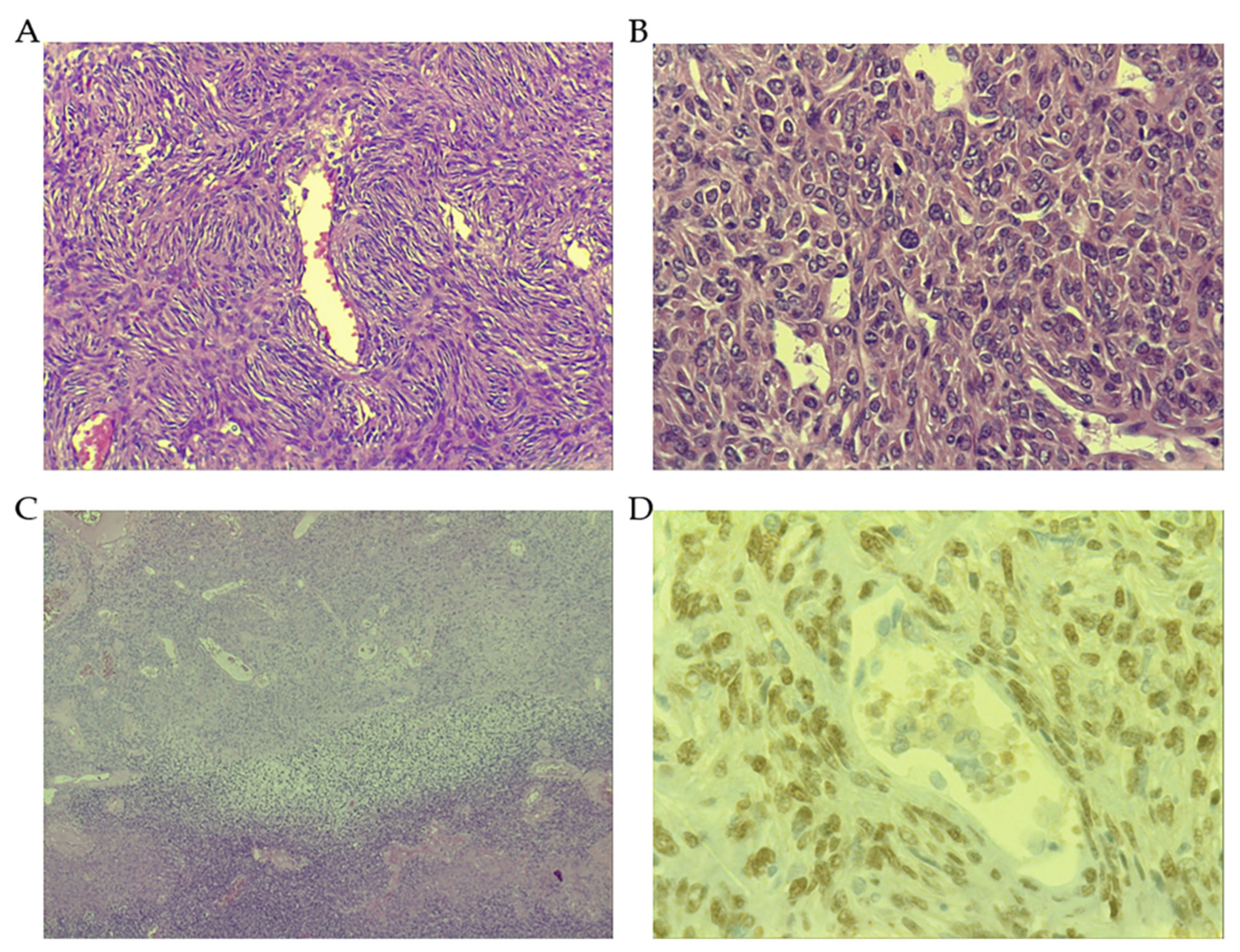
3.1. Morphological Features
3.2. The Role of Signal Transducer and Activator of Transcription 6 (STAT6)
3.3. Other Immunohistochemical Markers
4. Dedifferentiated SFT (DD-SFT)
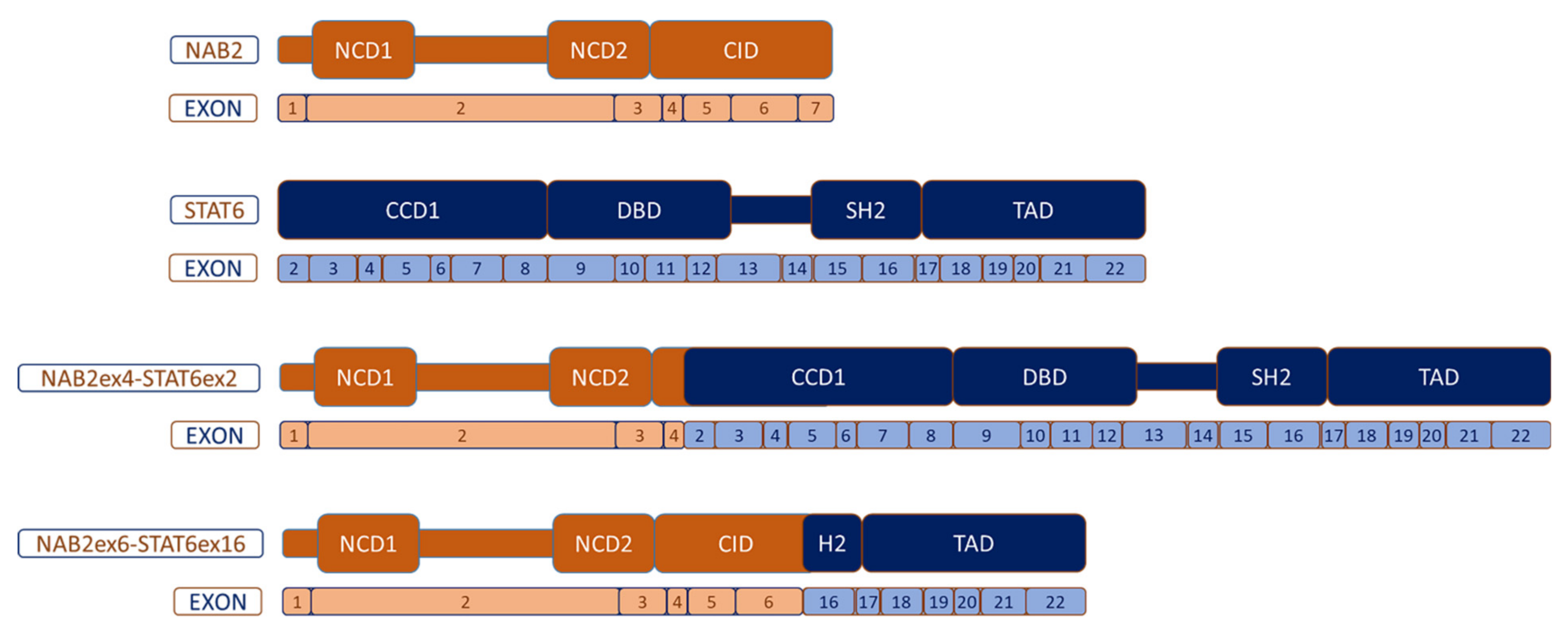
5. Molecular Biology
6. Biological Rationale for Antiangiogenic Therapy in SFT
7. Biological Rationale for Not Using Doxorubicin in SFT (Except for DD-SFT)
8. Treatment in SFT
8.1. Surgery and Radiation Therapy
8.2. Chemotherapy
8.3. Antiangiogenics
9. Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kinslow, C.J.; Wang, T.J.C. Incidence of extrameningeal solitary fibrous tumors. Cancer 2020, 126, 4067. [Google Scholar] [CrossRef]
- Kinslow, C.J.; Bruce, S.S.; Rae, A.I.; Sheth, S.A.; McKhann, G.M.; Sisti, M.B.; Bruce, J.N.; Sonabend, A.M.; Wang, T.J.C. Solitary-fibrous tumor/hemangiopericytoma of the central nervous system: A population-based study. J. Neuro-Oncol. 2018, 138, 173–182. [Google Scholar] [CrossRef]
- Fletcher, C.D. The evolving classification of soft tissue tumours—An update based on the new 2013 WHO classification. Histopathology 2014, 64, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Kallen, M.E.; Hornick, J.L. The 2020 WHO Classification: What’s New in Soft Tissue Tumor Pathology? Am. J. Surg. Pathol. 2021, 45, e1–e23. [Google Scholar] [CrossRef]
- O’Neill, A.C.; Tirumani, S.H.; Do, W.S.; Keraliya, A.R.; Hornick, J.L.; Shinagare, A.B.; Ramaiya, N.H. Metastatic Patterns of Solitary Fibrous Tumors: A Single-Institution Experience. AJR. Am. J. Roentgenol. 2017, 208, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Demicco, E.G.; Park, M.S.; Araujo, D.M.; Fox, P.S.; Bassett, R.L.; Pollock, R.E.; Lazar, A.J.; Wang, W.L. Solitary fibrous tumor: A clinicopathological study of 110 cases and proposed risk assessment model. Mod. Pathol. 2012, 25, 1298–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldi, G.G.; Stacchiotti, S.; Mauro, V.; Dei Tos, A.P.; Gronchi, A.; Pastorino, U.; Duranti, L.; Provenzano, S.; Marrari, A.; Libertini, M.; et al. Solitary fibrous tumor of all sites: Outcome of late recurrences in 14 patients. Clin. Sarcoma Res. 2013, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Fritchie, K.; Jensch, K.; Moskalev, E.A.; Caron, A.; Jenkins, S.; Link, M.; Brown, P.D.; Rodriguez, F.J.; Guajardo, A.; Brat, D.; et al. The impact of histopathology and NAB2-STAT6 fusion subtype in classification and grading of meningeal solitary fibrous tumor/hemangiopericytoma. Acta Neuropathol. 2019, 137, 307–319. [Google Scholar] [CrossRef] [Green Version]
- Salas, S.; Resseguier, N.; Blay, J.Y.; Le Cesne, A.; Italiano, A.; Chevreau, C.; Rosset, P.; Isambert, N.; Soulie, P.; Cupissol, D.; et al. Prediction of local and metastatic recurrence in solitary fibrous tumor: Construction of a risk calculator in a multicenter cohort from the French Sarcoma Group (FSG) database. Ann. Oncol. 2017, 28, 1979–1987. [Google Scholar] [CrossRef]
- Callemeyn, J.; Van Haecke, P.; Peetermans, W.E.; Blockmans, D. Clubbing and hypertrophic osteoarthropathy: Insights in diagnosis, pathophysiology, and clinical significance. Acta Clin. Belg. 2016, 71, 123–130. [Google Scholar] [CrossRef]
- Dickinson, C.J.; Martin, J.F. Megakaryocytes and platelet clumps as the cause of finger clubbing. Lancet 1987, 2, 1434–1435. [Google Scholar] [CrossRef]
- Atkinson, S.; Fox, S.B. Vascular endothelial growth factor (VEGF)-A and platelet-derived growth factor (PDGF) play a central role in the pathogenesis of digital clubbing. J. Pathol. 2004, 203, 721–728. [Google Scholar] [CrossRef]
- Olan, F.; Portela, M.; Navarro, C.; Gaxiola, M.; Silveira, L.H.; Ruiz, V.; Martinez-Lavin, M. Circulating vascular endothelial growth factor concentrations in a case of pulmonary hypertrophic osteoarthropathy. Correlation with disease activity. J. Rheumatol. 2004, 31, 614–616. [Google Scholar]
- Le Roith, D. Tumor-induced hypoglycemia. N. Engl. J. Med. 1999, 341, 757–758. [Google Scholar] [CrossRef] [PubMed]
- Wignall, O.J.; Moskovic, E.C.; Thway, K.; Thomas, J.M. Solitary fibrous tumors of the soft tissues: Review of the imaging and clinical features with histopathologic correlation. Am. J. Roentgenol. 2010, 195, W55–W62. [Google Scholar] [CrossRef] [PubMed]
- Helage, S.; Revel, M.P.; Chabi, M.L.; Audureau, E.; Ferretti, G.; Laurent, F.; Alifano, M.; Mansuet-Lupo, A.; Buy, J.N.; Vadrot, D. Solitary fibrous tumor of the pleura: Can computed tomography features help predict malignancy? A series of 56 patients with histopathological correlates. Diagn. Interv. Imaging 2016, 97, 347–353. [Google Scholar] [CrossRef]
- Rosado-de-Christenson, M.L.; Abbott, G.F.; McAdams, H.P.; Franks, T.J.; Galvin, J.R. From the archives of the AFIP: Localized fibrous tumor of the pleura. Radiographics 2003, 23, 759–783. [Google Scholar] [CrossRef] [PubMed]
- Ginat, D.T.; Bokhari, A.; Bhatt, S.; Dogra, V. Imaging features of solitary fibrous tumors. Am. J. Roentgenol. 2011, 196, 487–495. [Google Scholar] [CrossRef]
- Tazeler, Z.; Tan, G.; Aslan, A.; Tan, S. The utility of 18F-FDG PET/CT in solitary fibrous tumors of the pleura. Rev. Esp. Med. Nucl. Imagen Mol. 2016, 35, 165–170. [Google Scholar] [CrossRef]
- WHO Classification of Tumours Editorial Board. Soft Tissue and Bone Tumours. In WHO Classification of Tumours Series, 5th ed.; International Agency for Research on Cancer: Lyon, France, 2020; Volume 3, Available online: https://publications.Iarc.fr/588 (accessed on 10 June 2021).
- Martin-Broto, J.; Cruz, J.; Penel, N.; Le Cesne, A.; Hindi, N.; Luna, P.; Moura, D.S.; Bernabeu, D.; de Alava, E.; Lopez-Guerrero, J.A.; et al. Pazopanib for treatment of typical solitary fibrous tumours: A multicentre, single-arm, phase 2 trial. Lancet Oncol. 2020, 21, 456–466. [Google Scholar] [CrossRef]
- Pasquali, S.; Gronchi, A.; Strauss, D.; Bonvalot, S.; Jeys, L.; Stacchiotti, S.; Hayes, A.; Honore, C.; Collini, P.; Renne, S.L.; et al. Resectable extra-pleural and extra-meningeal solitary fibrous tumours: A multi-centre prognostic study. Eur. J. Surg. Oncol. 2016, 42, 1064–1070. [Google Scholar] [CrossRef]
- Tapias, L.F.; Mino-Kenudson, M.; Lee, H.; Wright, C.; Gaissert, H.A.; Wain, J.C.; Mathisen, D.J.; Lanuti, M. Risk factor analysis for the recurrence of resected solitary fibrous tumours of the pleura: A 33-year experience and proposal for a scoring system. Eur. J. Cardio-Thorac. Surg. 2013, 44, 111–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgiesh, T.; Boye, K.; Bjerkehagen, B. A novel risk score to predict early and late recurrence in solitary fibrous tumour. Histopathology 2020, 77, 123–132. [Google Scholar] [CrossRef]
- Gholami, S.; Cassidy, M.R.; Kirane, A.; Kuk, D.; Zanchelli, B.; Antonescu, C.R.; Singer, S.; Brennan, M. Size and Location are the Most Important Risk Factors for Malignant Behavior in Resected Solitary Fibrous Tumors. Ann. Surg. Oncol. 2017, 24, 3865–3871. [Google Scholar] [CrossRef] [PubMed]
- Demicco, E.G.; Wagner, M.J.; Maki, R.G.; Gupta, V.; Iofin, I.; Lazar, A.J.; Wang, W.L. Risk assessment in solitary fibrous tumors: Validation and refinement of a risk stratification model. Mod. Pathol. 2017, 30, 1433–1442. [Google Scholar] [CrossRef]
- Reisenauer, J.S.; Mneimneh, W.; Jenkins, S.; Mansfield, A.S.; Aubry, M.C.; Fritchie, K.J.; Allen, M.S.; Blackmon, S.H.; Cassivi, S.D.; Nichols, F.C.; et al. Comparison of Risk Stratification Models to Predict Recurrence and Survival in Pleuropulmonary Solitary Fibrous Tumor. J. Thorac. Oncol. 2018, 13, 1349–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.C.; Fletcher, C.D. Malignant fat-forming solitary fibrous tumor (so-called “lipomatous hemangiopericytoma”): Clinicopathologic analysis of 14 cases. Am. J. Surg. Pathol. 2011, 35, 1177–1185. [Google Scholar] [CrossRef]
- Dong, S.S.; Wang, N.; Yang, C.P.; Zhang, G.C.; Liang, W.H.; Zhao, J.; Qi, Y. Giant Cell-Rich Solitary Fibrous Tumor in the Nasopharynx: Case Report and Literature Review. OncoTargets Ther. 2020, 13, 6819–6826. [Google Scholar] [CrossRef]
- Thway, K.; Hayes, A.; Ieremia, E.; Fisher, C. Heterologous osteosarcomatous and rhabdomyosarcomatous elements in dedifferentiated solitary fibrous tumor: Further support for the concept of dedifferentiation in solitary fibrous tumor. Ann. Diagn. Pathol. 2013, 17, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.A.; Vivero, M.; Fletcher, C.D.; Mertens, F.; Hornick, J.L. Nuclear expression of STAT6 distinguishes solitary fibrous tumor from histologic mimics. Mod. Pathol. 2014, 27, 390–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagrada, G.P.; Spagnuolo, R.D.; Mauro, V.; Tamborini, E.; Cesana, L.; Gronchi, A.; Stacchiotti, S.; Pierotti, M.A.; Negri, T.; Pilotti, S. Solitary fibrous tumors: Loss of chimeric protein expression and genomic instability mark dedifferentiation. Mod. Pathol. 2015, 28, 1074–1083. [Google Scholar] [CrossRef] [Green Version]
- Barron, L.; Wynn, T.A. Fibrosis is regulated by Th2 and Th17 responses and by dynamic interactions between fibroblasts and macrophages. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G723–G728. [Google Scholar] [CrossRef] [Green Version]
- Ingram, J.L.; Antao-Menezes, A.; Mangum, J.B.; Lyght, O.; Lee, P.J.; Elias, J.A.; Bonner, J.C. Opposing actions of Stat1 and Stat6 on IL-13-induced up-regulation of early growth response-1 and platelet-derived growth factor ligands in pulmonary fibroblasts. J. Immunol. 2006, 177, 4141–4148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demicco, E.G.; Harms, P.W.; Patel, R.M.; Smith, S.C.; Ingram, D.; Torres, K.; Carskadon, S.L.; Camelo-Piragua, S.; McHugh, J.B.; Siddiqui, J.; et al. Extensive survey of STAT6 expression in a large series of mesenchymal tumors. Am. J. Clin. Pathol. 2015, 143, 672–682. [Google Scholar] [CrossRef] [Green Version]
- Doyle, L.A.; Tao, D.; Marino-Enriquez, A. STAT6 is amplified in a subset of dedifferentiated liposarcoma. Mod. Pathol. 2014, 27, 1231–1237. [Google Scholar] [CrossRef] [Green Version]
- Ouladan, S.; Trautmann, M.; Orouji, E.; Hartmann, W.; Huss, S.; Buttner, R.; Wardelmann, E. Differential diagnosis of solitary fibrous tumors: A study of 454 soft tissue tumors indicating the diagnostic value of nuclear STAT6 relocation and ALDH1 expression combined with in situ proximity ligation assay. Int. J. Oncol. 2015, 46, 2595–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoi, T.; Tsuzuki, T.; Yatabe, Y.; Suzuki, M.; Kurumaya, H.; Koshikawa, T.; Kuhara, H.; Kuroda, M.; Nakamura, N.; Nakatani, Y.; et al. Solitary fibrous tumour: Significance of p53 and CD34 immunoreactivity in its malignant transformation. Histopathology 1998, 32, 423–432. [Google Scholar] [CrossRef]
- Waller, E.K.; Olweus, J.; Lund-Johansen, F.; Huang, S.; Nguyen, M.; Guo, G.R.; Terstappen, L. The “common stem cell” hypothesis reevaluated: Human fetal bone marrow contains separate populations of hematopoietic and stromal progenitors. Blood 1995, 85, 2422–2435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quirici, N.; Soligo, D.; Bossolasco, P.; Servida, F.; Lumini, C.; Deliliers, G.L. Isolation of bone marrow mesenchymal stem cells by anti-nerve growth factor receptor antibodies. Exp. Hematol. 2002, 30, 783–791. [Google Scholar] [CrossRef]
- De Francesco, F.; Tirino, V.; Desiderio, V.; Ferraro, G.; D’Andrea, F.; Giuliano, M.; Libondi, G.; Pirozzi, G.; De Rosa, A.; Papaccio, G. Human CD34/CD90 ASCs are capable of growing as sphere clusters, producing high levels of VEGF and forming capillaries. PLoS ONE 2009, 4, e6537. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, T.; Matsuno, Y.; Shimoda, T.; Hirohashi, S.; Hirose, T.; Sano, T. Frequent expression of bcl-2 protein in solitary fibrous tumors. Jpn. J. Clin. Oncol. 1998, 28, 86–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takizawa, I.; Saito, T.; Kitamura, Y.; Arai, K.; Kawaguchi, M.; Takahashi, K.; Hara, N. Primary solitary fibrous tumor (SFT) in the retroperitoneum. Urol. Oncol. 2008, 26, 254–259. [Google Scholar] [CrossRef]
- Suster, S.; Fisher, C.; Moran, C.A. Expression of bcl-2 oncoprotein in benign and malignant spindle cell tumors of soft tissue, skin, serosal surfaces, and gastrointestinal tract. Am. J. Surg. Pathol. 1998, 22, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Baer, R. Bcl-2 breathes life into embryogenesis. Am. J. Pathol. 1994, 145, 7–10. [Google Scholar]
- Aronica, M.A.; Goenka, S.; Boothby, M. IL-4-dependent induction of BCL-2 and BCL-X(L)IN activated T lymphocytes through a STAT6- and pi 3-kinase-independent pathway. Cytokine 2000, 12, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, N.; Naka, T.; Yamamoto, I.; Fukuda, T.; Tsuneyoshi, M. Overexpression of bcl-2 protein in synovial sarcoma: A comparative study of other soft tissue spindle cell sarcomas and an additional analysis by fluorescence in situ hybridization. Hum. Pathol. 1996, 27, 1060–1065. [Google Scholar] [CrossRef]
- Manara, M.C.; Pasello, M.; Scotlandi, K. CD99: A Cell Surface Protein with an Oncojanus Role in Tumors. Genes 2018, 9, 159. [Google Scholar] [CrossRef] [Green Version]
- Maitra, A.; Hansel, D.E.; Argani, P.; Ashfaq, R.; Rahman, A.; Naji, A.; Deng, S.; Geradts, J.; Hawthorne, L.; House, M.G.; et al. Global expression analysis of well-differentiated pancreatic endocrine neoplasms using oligonucleotide microarrays. Clin. Cancer Res. 2003, 9, 5988–5995. [Google Scholar]
- Manara, M.C.; Bernard, G.; Lollini, P.L.; Nanni, P.; Zuntini, M.; Landuzzi, L.; Benini, S.; Lattanzi, G.; Sciandra, M.; Serra, M.; et al. CD99 acts as an oncosuppressor in osteosarcoma. Mol. Biol. Cell 2006, 17, 1910–1921. [Google Scholar] [CrossRef]
- Scotlandi, K.; Zuntini, M.; Manara, M.C.; Sciandra, M.; Rocchi, A.; Benini, S.; Nicoletti, G.; Bernard, G.; Nanni, P.; Lollini, P.L.; et al. CD99 isoforms dictate opposite functions in tumour malignancy and metastases by activating or repressing c-Src kinase activity. Oncogene 2007, 26, 6604–6618. [Google Scholar] [CrossRef] [Green Version]
- Bremond, A.; Meynet, O.; Mahiddine, K.; Coito, S.; Tichet, M.; Scotlandi, K.; Breittmayer, J.P.; Gounon, P.; Gleeson, P.A.; Bernard, A.; et al. Regulation of HLA class I surface expression requires CD99 and p230/golgin-245 interaction. Blood 2009, 113, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Olson, N.J.; Linos, K. Dedifferentiated Solitary Fibrous Tumor: A Concise Review. Arch. Pathol. Lab. Med. 2018, 142, 761–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Zhang, Q.; Yu, X.; Han, X.; Wang, H.; Xu, Y.; Qiu, X.; Jin, F. Immunohistochemical detection of STAT6, CD34, CD99 and BCL-2 for diagnosing solitary fibrous tumors/hemangiopericytomas. Int. J. Clin. Exp. Pathol. 2015, 8, 13166–13175. [Google Scholar]
- Fletcher, C.D.M.; Bridge, J.A.; Lee, J.C. Extrapleural solitary fibrous tumor. In WHO Classification of Tumors of Soft Tissue and Bone; Fletcher, C.D.M., Bridge, J.A., Hogendoom, P.C.W., Mertens, F., Eds.; IARC: Lyon, France, 2013; pp. 80–82. [Google Scholar]
- Yang, J.W.; Song, D.H.; Jang, I.S.; Ko, G.H. Dedifferentiated solitary fibrous tumor of thoracic cavity. Korean J. Pathol. 2014, 48, 250–253. [Google Scholar] [CrossRef]
- Schneider, N.; Hallin, M.; Thway, K. STAT6 Loss in Dedifferentiated Solitary Fibrous Tumor. Int. J. Surg. Pathol. 2017, 25, 58–60. [Google Scholar] [CrossRef]
- Schirosi, L.; Lantuejoul, S.; Cavazza, A.; Murer, B.; Yves Brichon, P.; Migaldi, M.; Sartori, G.; Sgambato, A.; Rossi, G. Pleuro-pulmonary solitary fibrous tumors: A clinicopathologic, immunohistochemical, and molecular study of 88 cases confirming the prognostic value of de Perrot staging system and p53 expression, and evaluating the role of c-kit, BRAF, PDGFRs (alpha/beta), c-met, and EGFR. Am. J. Surg. Pathol. 2008, 32, 1627–1642. [Google Scholar] [CrossRef] [PubMed]
- Kurisaki-Arakawa, A.; Akaike, K.; Hara, K.; Arakawa, A.; Takahashi, M.; Mitani, K.; Yao, T.; Saito, T. A case of dedifferentiated solitary fibrous tumor in the pelvis with TP53 mutation. Virchows Arch. 2014, 465, 615–621. [Google Scholar] [CrossRef]
- Akaike, K.; Kurisaki-Arakawa, A.; Hara, K.; Suehara, Y.; Takagi, T.; Mitani, K.; Kaneko, K.; Yao, T.; Saito, T. Distinct clinicopathological features of NAB2-STAT6 fusion gene variants in solitary fibrous tumor with emphasis on the acquisition of highly malignant potential. Hum. Pathol. 2015, 46, 347–356. [Google Scholar] [CrossRef]
- Hu, F.; Xue, M.; Li, Y.; Jia, Y.J.; Zheng, Z.J.; Yang, Y.L.; Guan, M.P.; Sun, L.; Xue, Y.M. Early Growth Response 1 (Egr1) Is a Transcriptional Activator of NOX4 in Oxidative Stress of Diabetic Kidney Disease. J. Diabetes Res. 2018, 2018, 3405695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakawa, Y.; Miyake, H.; Horiguchi, H.; Inokuchi, T.; Hino, N.; Ogasawara, T.; Kuroda, T.; Yamasaki, S. Overexpression of IGF2 and IGF2 receptor in malignant solitary fibrous tumor with hypoglycemia: A case report. Surg. Case Rep. 2018, 4, 106. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Kohashi, K.; Kinoshita, I.; Yamamoto, H.; Iwasaki, T.; Yoshimoto, M.; Ishihara, S.; Toda, Y.; Itou, Y.; Koga, Y.; et al. Clinicopathological review of solitary fibrous tumors: Dedifferentiation is a major cause of patient death. Virchows Arch. 2019, 475, 467–477. [Google Scholar] [CrossRef] [PubMed]
- England, D.M.; Hochholzer, L.; McCarthy, M.J. Localized benign and malignant fibrous tumors of the pleura. A clinicopathologic review of 223 cases. Am. J. Surg. Pathol. 1989, 13, 640–658. [Google Scholar] [CrossRef] [PubMed]
- Martin-Broto, J.; Stacchiotti, S.; Lopez-Pousa, A.; Redondo, A.; Bernabeu, D.; de Alava, E.; Casali, P.G.; Italiano, A.; Gutierrez, A.; Moura, D.S.; et al. Pazopanib for treatment of advanced malignant and dedifferentiated solitary fibrous tumour: A multicentre, single-arm, phase 2 trial. Lancet Oncol. 2019, 20, 134–144. [Google Scholar] [CrossRef]
- Chen, R.H.; Du, Y.; Han, P.; Wang, H.B.; Liang, F.Y.; Feng, G.K.; Zhou, A.J.; Cai, M.Y.; Zhong, Q.; Zeng, M.S.; et al. ISG15 predicts poor prognosis and promotes cancer stem cell phenotype in nasopharyngeal carcinoma. Oncotarget 2016, 7, 16910–16922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahrami, A.; Lee, S.; Schaefer, I.M.; Boland, J.M.; Patton, K.T.; Pounds, S.; Fletcher, C.D. TERT promoter mutations and prognosis in solitary fibrous tumor. Mod. Pathol. 2016, 29, 1511–1522. [Google Scholar] [CrossRef] [Green Version]
- Demicco, E.G.; Wani, K.; Ingram, D.; Wagner, M.; Maki, R.G.; Rizzo, A.; Meeker, A.; Lazar, A.J.; Wang, W.L. TERT promoter mutations in solitary fibrous tumour. Histopathology 2018, 73, 843–851. [Google Scholar] [CrossRef]
- Vogels, R.; Macagno, N.; Griewank, K.; Groenen, P.; Verdijk, M.; Fonville, J.; Kusters, B.; Figarella-Branger, D.; Wesseling, P.; Bouvier, C.; et al. Prognostic significance of NAB2-STAT6 fusion variants and TERT promotor mutations in solitary fibrous tumors/hemangiopericytomas of the CNS: Not (yet) clear. Acta Neuropathol. 2019, 137, 679–682. [Google Scholar] [CrossRef]
- Chmielecki, J.; Crago, A.M.; Rosenberg, M.; O’Connor, R.; Walker, S.R.; Ambrogio, L.; Auclair, D.; McKenna, A.; Heinrich, M.C.; Frank, D.A.; et al. Whole-exome sequencing identifies a recurrent NAB2-STAT6 fusion in solitary fibrous tumors. Nat. Genet. 2013, 45, 131–132. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.R.; Wu, Y.M.; Kalyana-Sundaram, S.; Cao, X.; Lonigro, R.J.; Sung, Y.S.; Chen, C.L.; Zhang, L.; Wang, R.; Su, F.; et al. Identification of recurrent NAB2-STAT6 gene fusions in solitary fibrous tumor by integrative sequencing. Nat. Genet. 2013, 45, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Mohajeri, A.; Tayebwa, J.; Collin, A.; Nilsson, J.; Magnusson, L.; von Steyern, F.V.; Brosjo, O.; Domanski, H.A.; Larsson, O.; Sciot, R.; et al. Comprehensive genetic analysis identifies a pathognomonic NAB2/STAT6 fusion gene, nonrandom secondary genomic imbalances, and a characteristic gene expression profile in solitary fibrous tumor. Genes Chromosomes Cancer 2013, 52, 873–886. [Google Scholar] [CrossRef]
- Lai, A.Y.; Wade, P.A. Cancer biology and NuRD: A multifaceted chromatin remodelling complex. Nat. Rev. Cancer 2011, 11, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, N.; Latos, P.; Hynes-Allen, A.; Loos, R.; Leaford, D.; O’Shaughnessy, A.; Mosaku, O.; Signolet, J.; Brennecke, P.; Kalkan, T.; et al. NuRD suppresses pluripotency gene expression to promote transcriptional heterogeneity and lineage commitment. Cell Stem Cell 2012, 10, 583–594. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Rodriguez, J.P.; Niu, F.; Pu, M.; Wang, J.; Hung, L.W.; Shao, Q.; Zhu, Y.; Ding, W.; Liu, Y.; et al. Structural basis for DNA recognition by STAT6. Proc. Nat. Acad. Sci. USA 2016, 113, 13015–13020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barthelmess, S.; Geddert, H.; Boltze, C.; Moskalev, E.A.; Bieg, M.; Sirbu, H.; Brors, B.; Wiemann, S.; Hartmann, A.; Agaimy, A.; et al. Solitary fibrous tumors/hemangiopericytomas with different variants of the NAB2-STAT6 gene fusion are characterized by specific histomorphology and distinct clinicopathological features. Am. J. Pathol. 2014, 184, 1209–1218. [Google Scholar] [CrossRef]
- Tai, H.C.; Chuang, I.C.; Chen, T.C.; Li, C.F.; Huang, S.C.; Kao, Y.C.; Lin, P.C.; Tsai, J.W.; Lan, J.; Yu, S.C.; et al. NAB2-STAT6 fusion types account for clinicopathological variations in solitary fibrous tumors. Mod. Pathol. 2015, 28, 1324–1335. [Google Scholar] [CrossRef]
- Bieg, M.; Moskalev, E.A.; Will, R.; Hebele, S.; Schwarzbach, M.; Schmeck, S.; Hohenberger, P.; Jakob, J.; Kasper, B.; Gaiser, T.; et al. Gene Expression in Solitary Fibrous Tumors (SFTs) Correlates with Anatomic Localization and NAB2-STAT6 Gene Fusion Variants. Am. J. Pathol. 2021. [Google Scholar] [CrossRef]
- Hatva, E.; Bohling, T.; Jaaskelainen, J.; Persico, M.G.; Haltia, M.; Alitalo, K. Vascular growth factors and receptors in capillary hemangioblastomas and hemangiopericytomas. Am. J. Pathol. 1996, 148, 763–775. [Google Scholar]
- Sawada, N.; Ishiwata, T.; Naito, Z.; Maeda, S.; Sugisaki, Y.; Asano, G. Immunohistochemical localization of endothelial cell markers in solitary fibrous tumor. Pathol. Int. 2002, 52, 769–776. [Google Scholar] [CrossRef]
- Demicco, E.G.; Wani, K.; Fox, P.S.; Bassett, R.L.; Young, E.D.; Lev, D.; Aldape, K.D.; Lazar, A.J.; Wang, W.L. Histologic variability in solitary fibrous tumors reflects angiogenic and growth factor signaling pathway alterations. Hum. Pathol. 2015, 46, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Silverman, E.S.; Khachigian, L.M.; Santiago, F.S.; Williams, A.J.; Lindner, V.; Collins, T. Vascular smooth muscle cells express the transcriptional corepressor NAB2 in response to injury. Am. J. Pathol. 1999, 155, 1311–1317. [Google Scholar] [CrossRef] [Green Version]
- Houston, P.; Campbell, C.J.; Svaren, J.; Milbrandt, J.; Braddock, M. The transcriptional corepressor NAB2 blocks Egr-1-mediated growth factor activation and angiogenesis. Biochem. Biophys. Res. Commun. 2001, 283, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Lucerna, M.; Mechtcheriakova, D.; Kadl, A.; Schabbauer, G.; Schafer, R.; Gruber, F.; Koshelnick, Y.; Muller, H.D.; Issbrucker, K.; Clauss, M.; et al. NAB2, a corepressor of EGR-1, inhibits vascular endothelial growth factor-mediated gene induction and angiogenic responses of endothelial cells. J. Biol. Chem. 2003, 278, 11433–11440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punetha, M.; Chouhan, V.S.; Sonwane, A.; Singh, G.; Bag, S.; Green, J.A.; Whitworth, K.; Sarkar, M. Early growth response gene mediates in VEGF and FGF signaling as dissected by CRISPR in corpus luteum of water buffalo. Sci. Rep. 2020, 10, 6849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.L.; Ripamonte, P.; Buratini, J.; Portela, V.M.; Price, C.A. Fibroblast growth factor-2 regulation of Sprouty and NR4A genes in bovine ovarian granulosa cells. J. Cell. Physiol. 2011, 226, 1820–1827. [Google Scholar] [CrossRef]
- Bae, S.K.; Bae, M.H.; Ahn, M.Y.; Son, M.J.; Lee, Y.M.; Bae, M.K.; Lee, O.H.; Park, B.C.; Kim, K.W. Egr-1 mediates transcriptional activation of IGF-II gene in response to hypoxia. Cancer Res. 1999, 59, 5989–5994. [Google Scholar] [PubMed]
- Pierscianek, D.; Michel, A.; Hindy, N.E.; Keyvani, K.; Dammann, P.; Oezkan, N.; Mueller, O.; Sure, U.; Zhu, Y. Activation of multiple angiogenic signaling pathways in hemangiopericytoma. Brain Tumor Pathol. 2016, 33, 200–208. [Google Scholar] [CrossRef]
- Baetta, R.; Soma, M.; De-Fraja, C.; Comparato, C.; Teruzzi, C.; Magrassi, L.; Cattaneo, E. Upregulation and activation of Stat6 precede vascular smooth muscle cell proliferation in carotid artery injury model. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 931–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Q.; Zhang, T.; Jiang, L.; Wang, L.; Luo, S. Expression and significance of STAT3 and VEGF with MVD in the nasal polyps. J. Clin. Otorhinolaryngol. Head Neck Surg. 2010, 24, 641–644, 647. [Google Scholar]
- Tang, X.; Yang, Y.; Yuan, H.; You, J.; Burkatovskaya, M.; Amar, S. Novel transcriptional regulation of VEGF in inflammatory processes. J. Cell. Mol. Med. 2013, 17, 386–397. [Google Scholar] [CrossRef]
- Judson, I.; Verweij, J.; Gelderblom, H.; Hartmann, J.T.; Schoffski, P.; Blay, J.Y.; Kerst, J.M.; Sufliarsky, J.; Whelan, J.; Hohenberger, P.; et al. Doxorubicin alone versus intensified doxorubicin plus ifosfamide for first-line treatment of advanced or metastatic soft-tissue sarcoma: A randomised controlled phase 3 trial. Lancet Oncol. 2014, 15, 415–423. [Google Scholar] [CrossRef]
- Ryan, C.W.; Merimsky, O.; Agulnik, M.; Blay, J.Y.; Schuetze, S.M.; Van Tine, B.A.; Jones, R.L.; Elias, A.D.; Choy, E.; Alcindor, T.; et al. PICASSO III: A Phase III, Placebo-Controlled Study of Doxorubicin with or Without Palifosfamide in Patients With Metastatic Soft Tissue Sarcoma. J. Clin. Oncol. 2016, 34, 3898–3905. [Google Scholar] [CrossRef] [PubMed]
- Tap, W.D.; Papai, Z.; Van Tine, B.A.; Attia, S.; Ganjoo, K.N.; Jones, R.L.; Schuetze, S.; Reed, D.; Chawla, S.P.; Riedel, R.F.; et al. Doxorubicin plus evofosfamide versus doxorubicin alone in locally advanced, unresectable or metastatic soft-tissue sarcoma (TH CR-406/SARC021): An international, multicentre, open-label, randomised phase 3 trial. Lancet Oncol. 2017, 18, 1089–1103. [Google Scholar] [CrossRef]
- Tap, W.D.; Wagner, A.J.; Schoffski, P.; Martin-Broto, J.; Krarup-Hansen, A.; Ganjoo, K.N.; Yen, C.C.; Abdul Razak, A.R.; Spira, A.; Kawai, A.; et al. Effect of Doxorubicin Plus Olaratumab vs Doxorubicin Plus Placebo on Survival in Patients With Advanced Soft Tissue Sarcomas: The ANNOUNCE Randomized Clinical Trial. JAMA 2020, 323, 1266–1276. [Google Scholar] [CrossRef]
- Gewirtz, D.A. A critical evaluation of the mechanisms of action proposed for the antitumor effects of the anthracycline antibiotics adriamycin and daunorubicin. Biochem. Pharmacol. 1999, 57, 727–741. [Google Scholar] [CrossRef]
- Chang, Y.L.; Lee, Y.C.; Wu, C.T. Thoracic solitary fibrous tumor: Clinical and pathological diversity. Lung Cancer 1999, 23, 53–60. [Google Scholar] [CrossRef]
- Harrison-Phipps, K.M.; Nichols, F.C.; Schleck, C.D.; Deschamps, C.; Cassivi, S.D.; Schipper, P.H.; Allen, M.S.; Wigle, D.A.; Pairolero, P.C. Solitary fibrous tumors of the pleura: Results of surgical treatment and long-term prognosis. J. Thorac. Cardiovasc. Surg. 2009, 138, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Li, W.; Shao, J.; Zhao, J. Thoracic solitary fibrous tumors: An analysis of 70 patients who underwent surgical resection in a single institution. J. Cancer Res. Clin. Oncol. 2020, 146, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Aridi, T.; Tawil, A.; Hashem, M.; Khoury, J.; Raad, R.A.; Youssef, P. Unique Presentation and Management Approach of Pleural Solitary Fibrous Tumor. Case Rep. Surg. 2019, 2019, 9706825. [Google Scholar] [CrossRef] [Green Version]
- Dingley, B.; Fiore, M.; Gronchi, A. Personalizing surgical margins in retroperitoneal sarcomas: An update. Expert Rev. Anticancer. Ther. 2019, 19, 613–631. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, R.; Ji, T.; Chen, Z.; Guo, W. Surgical treatment of primary solitary fibrous tumors involving the pelvic ring. PLoS ONE 2018, 13, e0207581. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, S.S.; Scheithauer, B.W.; Nascimento, A.G.; Hirose, T.; Davis, D.H. Solitary fibrous tumor of the meninges: A lesion distinct from fibrous meningioma. A clinicopathologic and immunohistochemical study. Am. J. Clin. Pathol. 1996, 106, 217–224. [Google Scholar] [CrossRef]
- Kim, J.M.; Choi, Y.L.; Kim, Y.J.; Park, H.K. Comparison and evaluation of risk factors for meningeal, pleural, and extrapleural solitary fibrous tumors: A clinicopathological study of 92 cases confirmed by STAT6 immunohistochemical staining. Pathol. Res. Pract. 2017, 213, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Haas, R.L.; Walraven, I.; Lecointe-Artzner, E.; van Houdt, W.J.; Scholten, A.N.; Strauss, D.; Schrage, Y.; Hayes, A.J.; Raut, C.P.; Fairweather, M.; et al. Management of meningeal solitary fibrous tumors/hemangiopericytoma; surgery alone or surgery plus postoperative radiotherapy? Acta Oncol. 2021, 60, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Haas, R.L.; Walraven, I.; Lecointe-Artzner, E.; van Houdt, W.J.; Strauss, D.; Schrage, Y.; Hayes, A.J.; Raut, C.P.; Fairweather, M.; Baldini, E.H.; et al. Extrameningeal solitary fibrous tumors-surgery alone or surgery plus perioperative radiotherapy: A retrospective study from the global solitary fibrous tumor initiative in collaboration with the Sarcoma Patients EuroNet. Cancer 2020, 126, 3002–3012. [Google Scholar] [CrossRef]
- Lee, J.H.; Jeon, S.H.; Park, C.K.; Park, S.H.; Yoon, H.I.; Chang, J.H.; Suh, C.O.; Kang, S.J.; Lim, D.H.; Kim, I.A.; et al. The Role of Postoperative Radiotherapy in Intracranial Solitary Fibrous Tumor/Hemangiopericytoma: A Multi-Institutional Retrospective Study (KROG 18-11). Cancer Res. Treat. 2021. [Google Scholar] [CrossRef]
- Krengli, M.; Cena, T.; Zilli, T.; Jereczek-Fossa, B.A.; De Bari, B.; Villa Freixa, S.; Kaanders, J.; Torrente, S.; Pasquier, D.; Sole, C.V.; et al. Radiotherapy in the treatment of extracranial hemangiopericytoma/solitary fibrous tumor: Study from the Rare Cancer Network. Radiother. Oncol. 2020, 144, 114–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, R.L.; Walraven, I.; Lecointe-Artzner, E.; Scholten, A.N.; van Houdt, W.J.; Griffin, A.M.; Ferguson, P.C.; Miah, A.B.; Zaidi, S.; DeLaney, T.F.; et al. Radiation Therapy as Sole Management for Solitary Fibrous Tumors (SFT): A Retrospective Study From the Global SFT Initiative in Collaboration With the Sarcoma Patients EuroNet. Int. J. Radiat. Oncol. Biol. Phys. 2018, 101, 1226–1233. [Google Scholar] [CrossRef]
- Martin-Broto, J.; Hindi, N.; Lopez-Pousa, A.; Peinado-Serrano, J.; Alvarez, R.; Alvarez-Gonzalez, A.; Italiano, A.; Sargos, P.; Cruz-Jurado, J.; Isern-Verdum, J.; et al. Assessment of Safety and Efficacy of Combined Trabectedin and Low-Dose Radiotherapy for Patients with Metastatic Soft-Tissue Sarcomas: A Nonrandomized Phase 1/2 Clinical Trial. JAMA Oncol. 2020, 6, 535–541. [Google Scholar] [CrossRef]
- Fletcher, C.D.M.; Bridge, J.A.; Hogendoorn, P.C.W.; Mertens, F. WHO Classification of Tumours of Soft Tissue and Bone, 4th ed.; IARC Press: Lyon, France, 2013; Volume 5. [Google Scholar]
- Stacchiotti, S.; Saponara, M.; Frapolli, R.; Tortoreto, M.; Cominetti, D.; Provenzano, S.; Negri, T.; Dagrada, G.P.; Gronchi, A.; Colombo, C.; et al. Patient-derived solitary fibrous tumour xenografts predict high sensitivity to doxorubicin/dacarbazine combination confirmed in the clinic and highlight the potential effectiveness of trabectedin or eribulin against this tumour. Eur. J. Cancer 2017, 76, 84–92. [Google Scholar] [CrossRef]
- Stacchiotti, S.; Libertini, M.; Negri, T.; Palassini, E.; Gronchi, A.; Fatigoni, S.; Poletti, P.; Vincenzi, B.; Dei Tos, A.P.; Mariani, L.; et al. Response to chemotherapy of solitary fibrous tumour: A retrospective study. Eur. J. Cancer 2013, 49, 2376–2383. [Google Scholar] [CrossRef]
- Constantinidou, A.; Jones, R.L.; Olmos, D.; Thway, K.; Fisher, C.; Al-Muderis, O.; Judson, I. Conventional anthracycline-based chemotherapy has limited efficacy in solitary fibrous tumour. Acta Oncol. 2012, 51, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Levard, A.; Derbel, O.; Meeus, P.; Ranchere, D.; Ray-Coquard, I.; Blay, J.Y.; Cassier, P.A. Outcome of patients with advanced solitary fibrous tumors: The Centre Leon Berard experience. BMC Cancer 2013, 13, 109. [Google Scholar] [CrossRef]
- Schoffski, P.; Timmermans, I.; Hompes, D.; Stas, M.; Sinnaeve, F.; De Leyn, P.; Coosemans, W.; Van Raemdonck, D.; Hauben, E.; Sciot, R.; et al. Clinical Presentation, Natural History, and Therapeutic Approach in Patients with Solitary Fibrous Tumor: A Retrospective Analysis. Sarcoma 2020, 2020, 1385978. [Google Scholar] [CrossRef] [Green Version]
- Park, M.S.; Ravi, V.; Conley, A.; Patel, S.R.; Trent, J.C.; Lev, D.C.; Lazar, A.J.; Wang, W.L.; Benjamin, R.S.; Araujo, D.M. The role of chemotherapy in advanced solitary fibrous tumors: A retrospective analysis. Clin. Sarcoma Res. 2013, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Outani, H.; Kobayashi, E.; Wasa, J.; Saito, M.; Takenaka, S.; Hayakawa, K.; Endo, M.; Takeuchi, A.; Kobayashi, H.; Kito, M.; et al. Clinical Outcomes of Patients with Metastatic Solitary Fibrous Tumors: A Japanese Musculoskeletal Oncology Group (JMOG) Multiinstitutional Study. Ann. Surg. Oncol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Stacchiotti, S.; Tortoreto, M.; Bozzi, F.; Tamborini, E.; Morosi, C.; Messina, A.; Libertini, M.; Palassini, E.; Cominetti, D.; Negri, T.; et al. Dacarbazine in solitary fibrous tumor: A case series analysis and preclinical evidence vis-a-vis temozolomide and antiangiogenics. Clin. Cancer Res. 2013, 19, 5192–5201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaigneau, L.; Kalbacher, E.; Thiery-Vuillemin, A.; Fagnoni-Legat, C.; Isambert, N.; Aherfi, L.; Pauchot, J.; Delroeux, D.; Servagi-Vernat, S.; Mansi, L.; et al. Efficacy of trabectedin in metastatic solitary fibrous tumor. Rare Tumors 2011, 3, e29. [Google Scholar] [CrossRef]
- Khalifa, J.; Ouali, M.; Chaltiel, L.; Le Guellec, S.; Le Cesne, A.; Blay, J.Y.; Cousin, P.; Chaigneau, L.; Bompas, E.; Piperno-Neumann, S.; et al. Efficacy of trabectedin in malignant solitary fibrous tumors: A retrospective analysis from the French Sarcoma Group. BMC Cancer 2015, 15, 700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, H.; Iwata, S.; Wakamatsu, T.; Hayakawa, K.; Yonemoto, T.; Wasa, J.; Oka, H.; Ueda, T.; Tanaka, S. Efficacy and safety of trabectedin for patients with unresectable and relapsed soft-tissue sarcoma in Japan: A Japanese Musculoskeletal Oncology Group study. Cancer 2020, 126, 1253–1263. [Google Scholar] [CrossRef]
- Park, M.S.; Patel, S.R.; Ludwig, J.A.; Trent, J.C.; Conrad, C.A.; Lazar, A.J.; Wang, W.L.; Boonsirikamchai, P.; Choi, H.; Wang, X.; et al. Activity of temozolomide and bevacizumab in the treatment of locally advanced, recurrent, and metastatic hemangiopericytoma and malignant solitary fibrous tumor. Cancer 2011, 117, 4939–4947. [Google Scholar] [CrossRef] [PubMed]
- Kawai, A.; Araki, N.; Sugiura, H.; Ueda, T.; Yonemoto, T.; Takahashi, M.; Morioka, H.; Hiraga, H.; Hiruma, T.; Kunisada, T.; et al. Trabectedin monotherapy after standard chemotherapy versus best supportive care in patients with advanced, translocation-related sarcoma: A randomised, open-label, phase 2 study. Lancet Oncol. 2015, 16, 406–416. [Google Scholar] [CrossRef]
- Forni, C.; Minuzzo, M.; Virdis, E.; Tamborini, E.; Simone, M.; Tavecchio, M.; Erba, E.; Grosso, F.; Gronchi, A.; Aman, P.; et al. Trabectedin (ET-743) promotes differentiation in myxoid liposarcoma tumors. Mol. Cancer Ther. 2009, 8, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Grohar, P.J.; Segars, L.E.; Yeung, C.; Pommier, Y.; D’Incalci, M.; Mendoza, A.; Helman, L.J. Dual targeting of EWS-FLI1 activity and the associated DNA damage response with trabectedin and SN38 synergistically inhibits Ewing sarcoma cell growth. Clin. Cancer Res. 2014, 20, 1190–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoffski, P.; Chawla, S.; Maki, R.G.; Italiano, A.; Gelderblom, H.; Choy, E.; Grignani, G.; Camargo, V.; Bauer, S.; Rha, S.Y.; et al. Eribulin versus dacarbazine in previously treated patients with advanced liposarcoma or leiomyosarcoma: A randomised, open-label, multicentre, phase 3 trial. Lancet 2016, 387, 1629–1637. [Google Scholar] [CrossRef]
- Schoffski, P.; Ray-Coquard, I.L.; Cioffi, A.; Bui, N.B.; Bauer, S.; Hartmann, J.T.; Krarup-Hansen, A.; Grunwald, V.; Sciot, R.; Dumez, H.; et al. Activity of eribulin mesylate in patients with soft-tissue sarcoma: A phase 2 study in four independent histological subtypes. Lancet Oncol. 2011, 12, 1045–1052. [Google Scholar] [CrossRef]
- Kawai, A.; Araki, N.; Naito, Y.; Ozaki, T.; Sugiura, H.; Yazawa, Y.; Morioka, H.; Matsumine, A.; Saito, K.; Asami, S.; et al. Phase 2 study of eribulin in patients with previously treated advanced or metastatic soft tissue sarcoma. Jpn. J. Clin. Oncol. 2017, 47, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Stacchiotti, S.; Negri, T.; Libertini, M.; Palassini, E.; Marrari, A.; De Troia, B.; Gronchi, A.; Dei Tos, A.P.; Morosi, C.; Messina, A.; et al. Sunitinib malate in solitary fibrous tumor (SFT). Ann. Oncol. 2012, 23, 3171–3179. [Google Scholar] [CrossRef] [PubMed]
- Maruzzo, M.; Martin-Liberal, J.; Messiou, C.; Miah, A.; Thway, K.; Alvarado, R.; Judson, I.; Benson, C. Pazopanib as first line treatment for solitary fibrous tumours: The Royal Marsden Hospital experience. Clin. Sarcoma Res. 2015, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Mulamalla, K.; Truskinovsky, A.M.; Dudek, A.Z. Rare case of hemangiopericytoma responds to sunitinib. Transl. Res. 2008, 151, 129–133. [Google Scholar] [CrossRef]
- George, S.; Merriam, P.; Maki, R.G.; Van den Abbeele, A.D.; Yap, J.T.; Akhurst, T.; Harmon, D.C.; Bhuchar, G.; O’Mara, M.M.; D’Adamo, D.R.; et al. Multicenter phase II trial of sunitinib in the treatment of nongastrointestinal stromal tumor sarcomas. J. Clin. Oncol. 2009, 27, 3154–3160. [Google Scholar] [CrossRef] [Green Version]
- Domont, J.; Massard, C.; Lassau, N.; Armand, J.P.; Le Cesne, A.; Soria, J.C. Hemangiopericytoma and antiangiogenic therapy: Clinical benefit of antiangiogenic therapy (sorafenib and sunitinib) in relapsed malignant haemangioperyctoma /solitary fibrous tumour. Investig. New Drugs 2010, 28, 199–202. [Google Scholar] [CrossRef]
- Martin-Broto, J.; Hindi, N.; Grignani, G.; Martinez-Trufero, J.; Redondo, A.; Valverde, C.; Stacchiotti, S.; Lopez-Pousa, A.; D’Ambrosio, L.; Gutierrez, A.; et al. Nivolumab and sunitinib combination in advanced soft tissue sarcomas: A multicenter, single-arm, phase Ib/II trial. J. Immunother. Cancer 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Valentin, T.; Fournier, C.; Penel, N.; Bompas, E.; Chaigneau, L.; Isambert, N.; Chevreau, C. Sorafenib in patients with progressive malignant solitary fibrous tumors: A subgroup analysis from a phase II study of the French Sarcoma Group (GSF/GETO). Investig. New Drugs 2013, 31, 1626–1627. [Google Scholar] [CrossRef]
- Xie, C.; Wan, X.; Quan, H.; Zheng, M.; Fu, L.; Li, Y.; Lou, L. Preclinical characterization of anlotinib, a highly potent and selective vascular endothelial growth factor receptor-2 inhibitor. Cancer Sci. 2018, 109, 1207–1219. [Google Scholar] [CrossRef] [Green Version]
- Hu-Lowe, D.D.; Zou, H.Y.; Grazzini, M.L.; Hallin, M.E.; Wickman, G.R.; Amundson, K.; Chen, J.H.; Rewolinski, D.A.; Yamazaki, S.; Wu, E.Y.; et al. Nonclinical antiangiogenesis and antitumor activities of axitinib (AG-013736), an oral, potent, and selective inhibitor of vascular endothelial growth factor receptor tyrosine kinases 1, 2, 3. Clin. Cancer Res. 2008, 14, 7272–7283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, W.K.; Sennino, B.; Williamson, C.W.; Falcón, B.; Hashizume, H.; Yao, L.C.; Aftab, D.T.; McDonald, D.M. VEGF and c-Met blockade amplify angiogenesis inhibition in pancreatic islet cancer. Cancer Res. 2011, 71, 4758–4768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, J.; Yamamoto, Y.; Funahashi, Y.; Tsuruoka, A.; Watanabe, T.; Wakabayashi, T.; Uenaka, T.; Asada, M. E7080, a novel inhibitor that targets multiple kinases, has potent antitumor activities against stem cell factor producing human small cell lung cancer H146, based on angiogenesis inhibition. Int. J. Cancer 2008, 122, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.A.; Boloor, A.; Cheung, M.; Kumar, R.; Crosby, R.M.; Davis-Ward, R.G.; Epperly, A.H.; Hinkle, K.W.; Hunter, R.N., 3rd; Johnson, J.H.; et al. Discovery of 5-[[4-[(2,3-dimethyl-2H-indazol-6-yl)methylamino]-2-pyrimidinyl]amino]-2-methyl-benzenesulfonamide (Pazopanib), a novel and potent vascular endothelial growth factor receptor inhibitor. J. Med. Chem. 2008, 51, 4632–4640. [Google Scholar] [CrossRef]
- Wilhelm, S.M.; Dumas, J.; Adnane, L.; Lynch, M.; Carter, C.A.; Schütz, G.; Thierauch, K.H.; Zopf, D. Regorafenib (BAY 73-4506): A new oral multikinase inhibitor of angiogenic, stromal and oncogenic receptor tyrosine kinases with potent preclinical antitumor activity. Int. J. Cancer 2011, 129, 245–255. [Google Scholar] [CrossRef]
- Wilhelm, S.M.; Carter, C.; Tang, L.; Wilkie, D.; McNabola, A.; Rong, H.; Chen, C.; Zhang, X.; Vincent, P.; McHugh, M.; et al. BAY 43-9006 exhibits broad spectrum oral antitumor activity and targets the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in tumor progression and angiogenesis. Cancer Res. 2004, 64, 7099–7109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, L.Q.M.; Eckhardt, S.G. Sunitinib: From Rational Design to Clinical Efficacy. J. Clin. Oncol. 2007, 25, 884–896. [Google Scholar] [CrossRef]
- Gitenay, D.; Baron, V.T. Is EGR1 a potential target for prostate cancer therapy? Future Oncol. 2009, 5, 993–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.L.; Gokgoz, N.; Samman, B.; Andrulis, I.L.; Wunder, J.S.; Demicco, E.G. RNA expression profiling reveals PRAME, a potential immunotherapy target, is frequently expressed in solitary fibrous tumors. Mod. Pathol. 2020. [Google Scholar] [CrossRef]
- Chalmers, Z.R.; Connelly, C.F.; Fabrizio, D.; Gay, L.; Ali, S.M.; Ennis, R.; Schrock, A.; Campbell, B.; Shlien, A.; Chmielecki, J.; et al. Analysis of 100,000 human cancer genomes reveals the landscape of tumor mutational burden. Genome Med. 2017, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, J.A.; Federman, N.; Anderson, P.; Macy, M.E.; Davis, L.E.; Riedel, R.F.; Muscal, J.A.; Daw, N.C.; Ratan, R.; Toretsky, J.; et al. Phase I study of TK216, a novel anti-ETS agent for Ewing sarcoma. Ann. Oncol. 2020, 31 (Suppl. S4), S914–S933. [Google Scholar] [CrossRef]
- Huijbers, E.J.M.; van der Werf, I.M.; Faber, L.D.; Sialino, L.D.; van der Laan, P.; Holland, H.A.; Cimpean, A.M.; Thijssen, V.; van Beijnum, J.R.; Griffioen, A.W. Targeting Tumor Vascular CD99 Inhibits Tumor Growth. Front. Immunol. 2019, 10, 651. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Author | N Total/Selected | External Validation | Subtypes | RFS/MFS/DFS | Overall Survival | ||
|---|---|---|---|---|---|---|---|
| Univariate | Multivariate | Univariate | Multivariate | ||||
| Salas et al. [9] | 214/162 | Yes | Extrameningeal | Age, Mitosis, Necrosis | Mitosis | Age, Mitosis | Age |
| Pasquali et al. [22] | 269/243 | No | Extrameningeal and Extrapleural | Mitosis, Necrosis, Hypercellularity, Pleomorphism, Positive margins | Mitosis, Hypercellularity, Pleomorphism | Mitosis, Necrosis, Hypercellularity, Pleomorphism, Age, deep-seated | Pleomorphism, Hypercellularity |
| Demicco et al. [6] | 110 | Yes | Extrameningeal | Age, Mitosis, Size, Necrosis | Mitosis, Size | Age, Size, Mitosis | NA |
| Gholami et al. [25] | 219 | No | Extrameningeal | Size | NA | T location, Size | T location, Size |
| Tapias et al. [23] | 59 | No | Pleural | Size, Hypercellularity, Necrosis, Mitosis, Pleural effusion, Parietal (vs. visceral) | Size | NA | NA |
| Reisenauer et al. [27] | 147 | NA | Pleuro-Pulmonary | NA | NA | Age, Sex, Pattern *, Pleomorphism, Necrosis, Mitosis, Size | NA |
| Georgiesh et al. [24] | 100 | No | Extrameningeal | Sex, Necrosis, Mitosis, Pleomorphism | Mitosis, Necrosis, Sex | Age, Mitosis, Necrosis, Pleomorphism | Age, Mitosis, Necrosis, |
| Marker | Normal Tissues/Precursors | Mesenchymal Benign Entities | Mesenchymal Malignant Tumors |
|---|---|---|---|
| CD 34 | -Early hematopoietic stem cells | - Giant cell fibroblastoma - Lipoma | -SFT * |
| -Mesenchymal stem cells | -Kaposi sarcoma | ||
| -Small-vessels endothelial cells | -Low-grade myofibroblastic sarcoma | ||
| -Embryonic fibroblasts | -Inflammatory myofibroblastic tumors | ||
| -Endoneurial cells, dermal dendritic interstitial fibroblastic cells | -DFSP | ||
| -Adipocitic cells | -GIST | ||
| bcl 2 | -Stem cells -Endocrine tissue -Long-lived cells | -Schwannoma -Spindle cell lipoma -Dendritic fibromyxolipoma -Neurofibromas (focal) | -SFT * -Fibrosarcoma -Low-grade myxofibrosarcoma -Dedifferentiated liposarcoma -Synovial sarcoma -DFSP -GIST -Kaposi sarcoma -MPNST |
| CD99 | -Bone marrow cells -Leukocytes | -Fibroma -Giant cell angiofibroma | -SFT * -Ewing sarcoma -Synovial sarcoma -Rhabdomyosarcoma -Osteosarcoma |
| STAT 6 | -SFT * -Unclassified sarcomas of spindle cell or epithelioid morphology (12%) -Desmoid tumors (8%) -Neurofibromas (5%) -Clear cell sarcoma (5%) -Well-diff./dedifferentiated liposarcoma |
| Series | Type of Study | n | Classification of SFT | Drug/Regimen | Responses | mPFS (Months) | mOS (Months) |
|---|---|---|---|---|---|---|---|
| Stacchiotti et al. 2013 [113] | Retrospective | 31 | Typical: 1 MSFT: 17 DSFT: 12 NA: 1 | Anthracycline-based (30 pts): 8 monotherapy 23 Anthracycline + Ifosfamide High-dose Ifosfamide (19 pts) | PR: 6 (20%); SD: 8 (27%); PD: 16 (53%) PR: 2 (10%) SD: 5 (26%) PD: 12 (63%) | 3.5 (MSFT) 5 (DSFT) 3 | 11.5 NR |
| Levard et al. 2013 [115] | Retrospective | 23 | NR | Anthracycline-based (19 pts): 9 monotherapy 9 Doxo-Ifo/palifosfamide 1 Pegylated liposomal doxorubicin | PR: 2/19 (10.5%) | 4 (Doxo) 6.7 (combination) | NR |
| Constantinidou et al. 2012 [114] | Retrospective | 24 | NR | Anthracycline-based (17 pts): 14 monotherapy 3 Anthracycline + IFO Ifosfamide (4) | PR: 1/17 (6%) PR: 0; SD: 3/4 (75%); PD: 1 (25%) | 4.2 NR | 14.6 NR |
| Schöffski et al. 2020 [116] | Retrospective | 94 (26 treated) | NR | Doxorubicin (15 pts) | PR: 2 (13%) SD: 4 (26%) PD: 7 (47%) NE: 2 (13%) | 4.8 | NR |
| Stacchiotti et al. 2017 [112] | Retrospective | 12 | MSFT: 7 DSFT: 5 | Doxorubicin + Dacarbazine | PR: 6 (50%) SD: 1 (8.3%) PD: 5 (41.7%) | 6 (MSFT) 10 (DSFT) | 19 |
| Outani et al. 2020 [118] | Retrospective | 60 (31 treated) | Typical: 7 (12%) MSFT: 35 (58%) NA: 18 (30%) | Anthracycline-based (11 pts) Gemcitabine–docetaxel (10 pts) Ifosfamide-based (7 pts) Trabectedin (6 pts) Eribulin (4 pts) | NR | 3 * 8 * 2 * 3.5 * 8 * | NR |
| Park et al. 2013 [117] | Retrospective | 21 | NR | Anthracycline-based (15 pts) 1 monotherapy 14 Doxorubicin combos Gemcitabine-based (5 pts) Paclitaxel (3 pts) | PR: 0; SD: 14/15 (93%); PD: 1/15 (7%) SD: 2/5 (40%); PD: 3/5 (40%) SD: 4/5 (80%); PD: 1/5 (20%) | 4.6 ** | 10.3 *** |
| Stacchiotti et al. 2013 [119] | Retrospective | 8 | MSFT: 3 DSFT: 5 | Dacarbazine | PR: 3 (38%) SD: 4 (50%) PD: 1 (12%) | 7 | NR |
| Chaigneau et al. 2011 [120] | Case report | 1 | MSFT | Trabectedin | PR | 8 | NR |
| Khalifa et al. 2015 [121] | Retrospective | 11 | Typical: 1 MSFT: 10 | Trabectedin | PR: 1 (9.1%) SD: 8 (72.7%) | 11.6 | 22.3 |
| Kobayashi et al. 2020 [122] | Retrospective | 6 | NR | Trabectedin | PR: 1 (16.7%) | 2.3 | NR |
| Author | N | Design | Scheme | mPFS (Months) | 6mPFSR | SFT Subtype | Response | |
|---|---|---|---|---|---|---|---|---|
| RECIST | Choi | |||||||
| Stacchiotti et al. 2013 [113] | 31 | Retrospective | Anthracycline-based | 3.5 (M SFT) 4.0 (DD-SFT) | 20% | 17 M SFT; 12 DD-SFT | PR 20% SD 27% | UNK |
| Park et al. 2013 [117] | 25 | Retrospective | Anthracycline-based and others | 4.6 | 28% | UNK | SD 80% | UNK |
| Park et al. 2011 [123] | 14 | Retrospective | Temozolomide and Bevacizumab | 9.7 | 78.6% | 3 T SFT; 5 M SFT; 6 UNK | PR 14% SD 86% | PR 79% SD 14% PD 7% |
| Stacchiotti et al. 2012 [130] | 35 | Retrospective | Sunitinib | 6 | 45% | 22 M SFT; 13 DD-SFT | PR 6% SD 54% PD 40% | PR 48% SD 17% PD 34% |
| Maruzzo et al. 2015 [131] | 13 | Prospective collection | Pazopanib | 4.7 | 44.9% | UNK | PR 9% SD 73% PD 18% | PR 46% SD 36% PD 18% |
| IC50 (nM) in Cell-Free Kinase Assay | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Inhibitor | VEGFR1 | VEGFR2 | VEGFR3 | PDGFRα | PDGFRβ | FGFR1 | FGFR2 | FGFR3 | FGFR4 | C-Met |
| Anlotinib [137] | 26.9 | 0.2 | 0.7 | ND | 115 | ND | ND | ND | ND | >1000 |
| Axitinib [138] | ND | 0.2 | ND | 5 | 1.6 | ND | ND | ND | ND | ND |
| Cabozantinib [139] | ND | 0.035 | ND | ND | 234 | >1000 | ND | ND | ND | 1.3 |
| Lenvatinib [140] | 22 | 4 | 5.2 | 51 | 39 | 46 | ND | ND | ND | ND |
| Pazopanib [141] | 10 | 30 | 47 | 71 | 84 | 140 | ND | 130 | 800 * | ND |
| Regorafenib [142] | 13 | 4.2 | 46 | ND | 22 | 202 | ND | ND | ND | ND |
| Sorafenib [143] | ND | 90 | ND | ND | 57 | 580 | ND | ND | ND | >1000 |
| Sunitinib [144] | 21 | 34 | 3 | ND | 75 | 437 | ND | ND | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Broto, J.; Mondaza-Hernandez, J.L.; Moura, D.S.; Hindi, N. A Comprehensive Review on Solitary Fibrous Tumor: New Insights for New Horizons. Cancers 2021, 13, 2913. https://doi.org/10.3390/cancers13122913
Martin-Broto J, Mondaza-Hernandez JL, Moura DS, Hindi N. A Comprehensive Review on Solitary Fibrous Tumor: New Insights for New Horizons. Cancers. 2021; 13(12):2913. https://doi.org/10.3390/cancers13122913
Chicago/Turabian StyleMartin-Broto, Javier, Jose L. Mondaza-Hernandez, David S. Moura, and Nadia Hindi. 2021. "A Comprehensive Review on Solitary Fibrous Tumor: New Insights for New Horizons" Cancers 13, no. 12: 2913. https://doi.org/10.3390/cancers13122913
APA StyleMartin-Broto, J., Mondaza-Hernandez, J. L., Moura, D. S., & Hindi, N. (2021). A Comprehensive Review on Solitary Fibrous Tumor: New Insights for New Horizons. Cancers, 13(12), 2913. https://doi.org/10.3390/cancers13122913