
Osteopontin as a Regulator of Colorectal Cancer Progression and Its Clinical Applications


 ,
, 
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
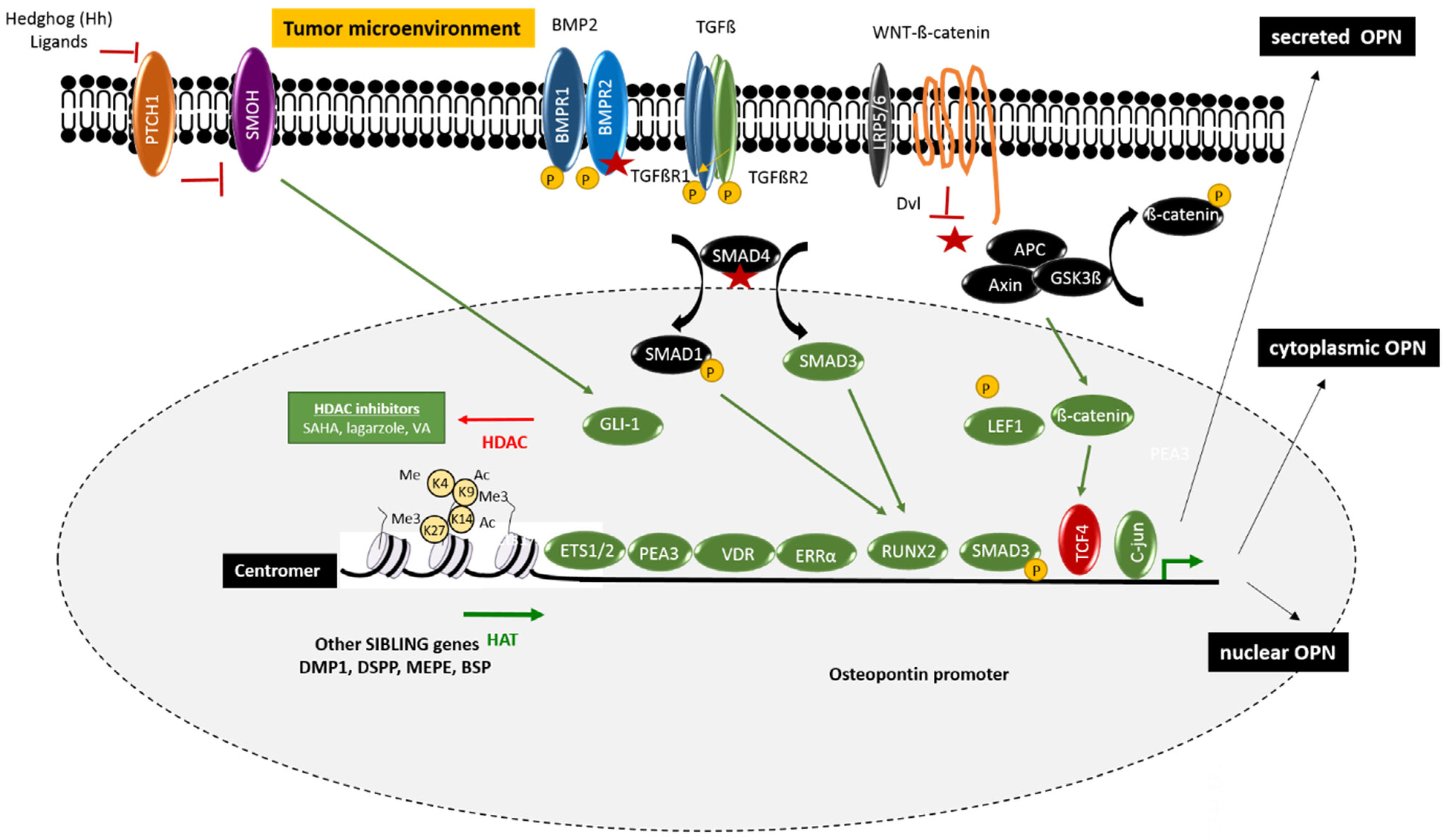
2. Regulation of Osteopontin
2.1. Epigenetic Regulators
2.1.1. Promoter Methylation, Histone Modifiers, and Chromatin Remodeling
2.1.2. Non-Coding RNAs
2.2. Gene Expression and Signaling Pathways
2.2.1. Wnt/Wingless Pathway
2.2.2. Bone Morphogen Protein/Transforming Growth Factor Pathway
2.2.3. Vitamin D Receptor (VDR) Pathway
2.2.4. Nuclear Receptor Pathway
2.2.5. Hedgehog Pathway
2.3. Alternative Splicing, Alternative Translation, and Protein Localization
2.4. Post-Translational Modifications
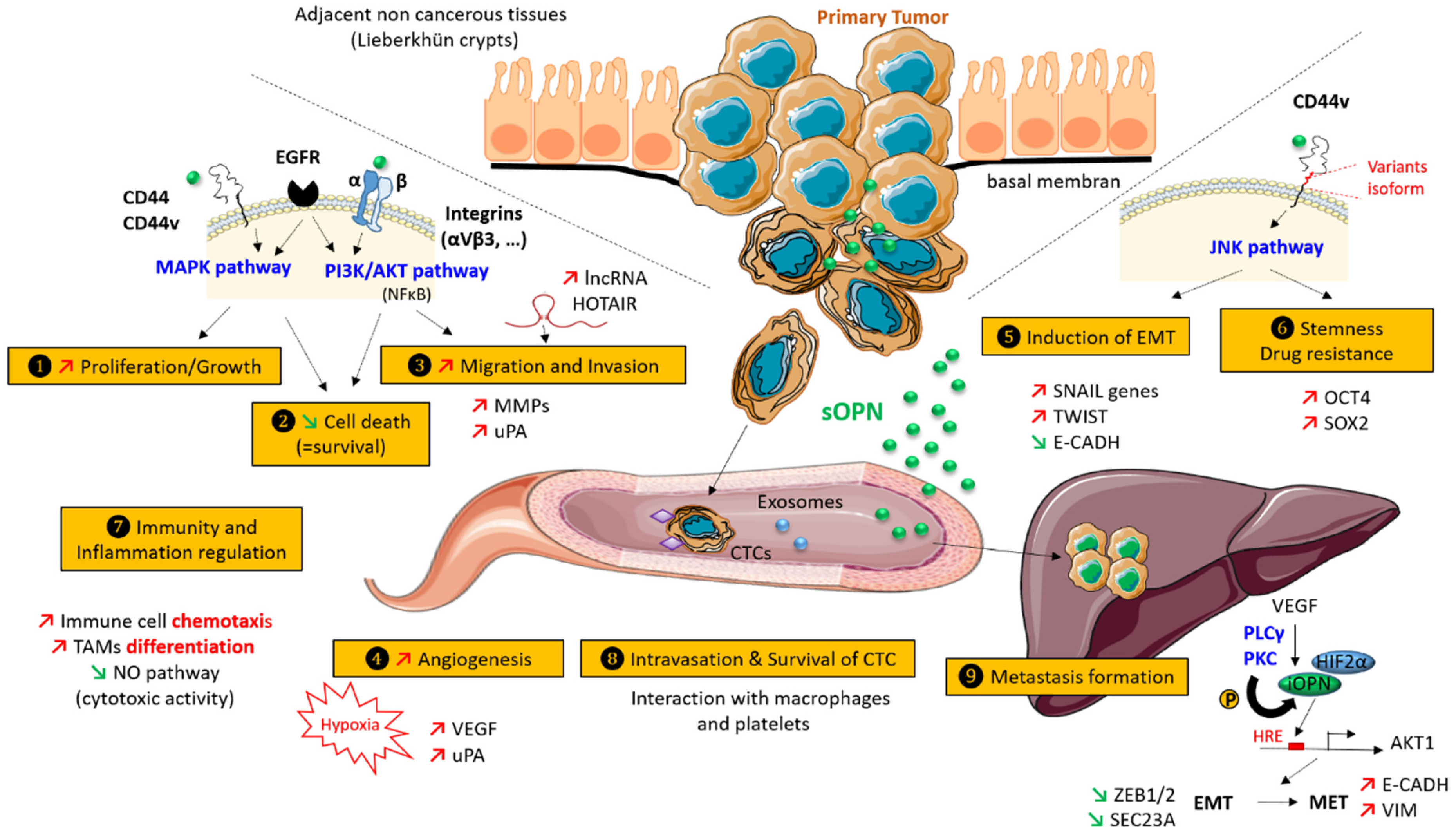
3. Role of Osteopontin in CRC
3.1. Increased Proliferation and Cell Growth
3.2. Inhibition of Cell Death
3.3. Migration and Invasion
3.4. Hypoxia and Neo-Angiogenesis
3.5. Induction of Epithelial to Mesenchymal Transition (EMT)
3.6. Stemness and Drug-Resistance
3.7. Immunity and Inflammation
3.8. Survival, Protection, and Intravasation of Circulating Tumor Cells (CTCs)
3.9. Metastasis Formation
4. Potential Applications of OPN as a Biomarker
4.1. Screening and Early Detection
4.2. Prognostic Value in CRC Patients
4.3. Evaluation of Recurrence Rate after Surgery
4.4. Potential Immune Marker
5. Potential Applications of OPN as a Therapeutic Target
6. Concluding Remarks and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021. [Google Scholar] [CrossRef]
- Zhong, W.; Yu, Z.; Zhan, J.; Yu, T.; Lin, Y.; Xia, Z.-S.; Yuan, Y.-H.; Chen, Q.-K. Association of Serum Levels of CEA, CA199, CA125, CYFRA21-1 and CA72-4 and Disease Characteristics in Colorectal Cancer. Pathol. Oncol. Res. POR 2015, 21. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, D.; Chen, T.; Irby, R.; Quackenbush, J.; Chambers, A.F.; Szabo, M.; Cantor, A.; Coppola, D.; Yeatman, T.J. Osteopontin Identified as Lead Marker of Colon Cancer Progression, Using Pooled Sample Expression Profiling. J. Natl. Cancer Inst. 2002, 94, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Coppola, D.; Szabo, M.; Boulware, D.; Muraca, P.; Alsarraj, M.; Chambers, A.F.; Yeatman, T.J. Correlation of Osteopontin Protein Expression and Pathological Stage across a Wide Variety of Tumor Histologies. Clin. Cancer Res. 2004, 10, 184–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denhardt, D.T.; Mistretta, D.; Chambers, A.F.; Krishna, S.; Porter, J.F.; Raghuram, S.; Rittling, S.R. Transcriptional Regulation of Osteopontin and the Metastatic Phenotype: Evidence for a Ras-Activated Enhancer in the Human OPN Promoter. Clin. Exp. Metastasis 2003, 20. [Google Scholar] [CrossRef] [PubMed]
- Bellahcène, A.; Castronovo, V.; Ogbureke, K.U.E.; Fisher, L.W.; Fedarko, N.S. Small Integrin-Binding Ligand N-Linked Glycoproteins (SIBLINGs): Multifunctional Proteins in Cancer. Nat. Rev. Cancer 2008, 8, 212–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Chen, Q.; Alam, A.; Cui, J.; Suen, K.C.; Soo, A.P.; Eguchi, S.; Gu, J.; Ma, D. The Role of Osteopontin in the Progression of Solid Organ Tumour. Cell Death Dis. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Senger, D.R.; Wirth, D.F.; Hynes, R.O. Transformed Mammalian Cells Secrete Specific Proteins and Phosphoproteins. Cell 1979, 16, 885–893. [Google Scholar] [CrossRef]
- Oldberg, A.; Franzén, A.; Heinegård, D. Cloning and Sequence Analysis of Rat Bone Sialoprotein (osteopontin) cDNA Reveals an Arg-Gly-Asp Cell-Binding Sequence. Proc. Natl. Acad. Sci. USA 1986, 83, 8819–8823. [Google Scholar] [CrossRef] [Green Version]
- Craig, A.M.; Smith, J.H.; Denhardt, D.T. Osteopontin, a Transformation-Associated Cell Adhesion Phosphoprotein, Is Induced by 12-O-Tetradecanoylphorbol 13-Acetate in Mouse Epidermis. J. Biol. Chem. 1989, 264, 9682–9689. [Google Scholar] [CrossRef]
- Patarca, R.; Freeman, G.J.; Singh, R.P.; Wei, F.Y.; Durfee, T.; Blattner, F.; Regnier, D.C.; Kozak, C.A.; Mock, B.A.; Morse, H.C., 3rd; et al. Structural and Functional Studies of the Early T Lymphocyte Activation 1 (Eta-1) Gene. Definition of a Novel T Cell-Dependent Response Associated with Genetic Resistance to Bacterial Infection. J. Exp. Med. 1989, 170, 145–161. [Google Scholar] [CrossRef] [Green Version]
- Arnsdorf, E.J.; Tummala, P.; Castillo, A.B.; Zhang, F.; Jacobs, C.R. The Epigenetic Mechanism of Mechanically Induced Osteogenic Differentiation. J. Biomech. 2010, 43, 2881–2886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, G.; Ren, A.; Qiu, Y.; Zhang, Y. Epigenetic Regulation of Osteogenic Differentiation of Mesenchymal Stem Cells. Curr. Stem Cell Res. Ther. 2016, 11, 235–246. [Google Scholar] [CrossRef]
- Huynh, N.C.-N.; Everts, V.; Ampornaramveth, R.S. Histone Deacetylases and Their Roles in Mineralized Tissue Regeneration. Bone Rep. 2017, 7, 33–40. [Google Scholar] [CrossRef]
- Janson, W.T.; Jenkins, L.J.; Chionh, F.; Mariadason, J.M. Aberrant DNA Methylation in Colorectal Cancer: What Should We Target? Trends Cancer 2017, 3, 698–712. [Google Scholar] [CrossRef]
- Haller, F.; Zhang, J.D.; Moskalev, E.A.; Braun, A.; Otto, C.; Geddert, H.; Riazalhosseini, Y.; Ward, A.; Balwierz, A.; Schaefer, I.-M.; et al. Combined DNA Methylation and Gene Expression Profiling in Gastrointestinal Stromal Tumors Reveals Hypomethylation of SPP1 as an Independent Prognostic Factor. Int. J. Cancer 2015, 136, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.-J.; Tsou, Y.-A.; Chen, H.-L.; Huang, H.-J.; Wu, S.-C.; Cheng, W.T.K.; Chen, C.Y.-C.; Chen, C.-M. Osteoponin Promoter Controlled by DNA Methylation: Aberrant Methylation in Cloned Porcine Genome. BioMed Res. Int. 2014, 2014, 1–16. [Google Scholar] [CrossRef]
- Sato, A.; Saito, Y.; Sugiyama, K.; Sakasegawa, N.; Muramatsu, T.; Fukuda, S.; Yoneya, M.; Kimura, M.; Ebinuma, H.; Hibi, T.; et al. Suppressive Effect of the Histone Deacetylase Inhibitor Suberoylanilide Hydroxamic Acid (SAHA) on Hepatitis C Virus Replication. J. Cell. Biochem. 2013, 114, 1987–1996. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, T.; Yan, H.; Qi, H.; Deng, C.; Ye, T.; Zhou, S.; Li, F.-R. Role of Histone Deacetylase Inhibitors in the Aging of Human Umbilical Cord Mesenchymal Stem Cells. J. Cell. Biochem. 2013, 114, 2231–2239. [Google Scholar] [CrossRef] [PubMed]
- Lund, R.J.; Emani, M.R.; Barbaric, I.; Kivinen, V.; Jones, M.; Baker, D.; Gokhale, P.; Nykter, M.; Lahesmaa, R.; Andrews, P.W. Karyotypically Abnormal Human ESCs Are Sensitive to HDAC Inhibitors and Show Altered Regulation of Genes Linked to Cancers and Neurological Diseases. Stem Cell Res. 2013, 11, 1022–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paino, F.; la Noce, M.; Tirino, V.; Naddeo, P.; Desiderio, V.; Pirozzi, G.; De Rosa, A.; Laino, L.; Altucci, L.; Papaccio, G. Histone Deacetylase Inhibition with Valproic Acid Downregulates Osteocalcin Gene Expression in Human Dental Pulp Stem Cells and Osteoblasts: Evidence for HDAC2 Involvement. Stem Cells Dayt. Ohio 2014, 32, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Zhang, P.; Ge, J.; Cheng, J.; Dong, W.; Yuan, H.; Du, Y.; Yang, M.; Sun, R.; Jiang, H. Histone Deacetylase 8 Suppresses Osteogenic Differentiation of Bone Marrow Stromal Cells by Inhibiting Histone H3K9 Acetylation and RUNX2 Activity. Int. J. Biochem. Cell Biol. 2014, 54, 68–77. [Google Scholar] [CrossRef]
- Cai, M.; Bompada, P.; Atac, D.; Laakso, M.; Groop, L.; De Marinis, Y. Epigenetic Regulation of Glucose-Stimulated Osteopontin (OPN) Expression in Diabetic Kidney. Biochem. Biophys. Res. Commun. 2016, 469, 108–113. [Google Scholar] [CrossRef]
- Xu, J.; Yu, B.; Hong, C.; Wang, C.-Y. KDM6B Epigenetically Regulates Odontogenic Differentiation of Dental Mesenchymal Stem Cells. Int. J. Oral Sci. 2013, 5, 200–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Matsui, Y.; Higashimoto, M.; Kawaguchi, Y.; Seki, S.; Motomura, H.; Hori, T.; Yahara, Y.; Kanamori, M.; Kimura, T. Myxoid Liposarcoma-Associated EWSR1-DDIT3 Selectively Represses Osteoblastic and Chondrocytic Transcription in Multipotent Mesenchymal Cells. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.D.; Garrison, J.; Guo, H.; Mi, Z.; Markovic, J.; Kim, V.M.; Kuo, P.C. Micro-RNA-181a Regulates Osteopontin Dependent Metastatic Function In Hepatocellular Cancer Cell Lines. Surgery 2010, 148, 291–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.-X.; Xia, Y.-H.; Xue, T.-C.; Ye, S.-L. Suppression of microRNA-96 Expression Inhibits the Invasion of Hepatocellular Carcinoma Cells. Mol. Med. Rep. 2012, 5, 800–804. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Liu, N.; Yang, Y.; Qiu, X. Regulation of Osteosarcoma Cell Invasion through Osteopontin Modification by miR-4262. Tumor Biol. 2016, 37, 6493–6499. [Google Scholar] [CrossRef]
- Taipaleenmäki, H.; Farina, N.H.; van Wijnen, A.J.; Stein, J.L.; Hesse, E.; Stein, G.S.; Lian, J.B. Antagonizing miR-218-5p Attenuates Wnt Signaling and Reduces Metastatic Bone Disease of Triple Negative Breast Cancer Cells. Oncotarget 2016, 7, 79032–79046. [Google Scholar] [CrossRef] [Green Version]
- Colden, M.; Dar, A.A.; Saini, S.; Dahiya, P.V.; Shahryari, V.; Yamamura, S.; Tanaka, Y.; Stein, G.; Dahiya, R.; Majid, S. MicroRNA-466 Inhibits Tumor Growth and Bone Metastasis in Prostate Cancer by Direct Regulation of Osteogenic Transcription Factor RUNX2. Cell Death Dis. 2017, 8, e2572. [Google Scholar] [CrossRef]
- Arffa, M.L.; Zapf, M.A.; Kothari, A.N.; Chang, V.; Gupta, G.N.; Ding, X.; Al-Gayyar, M.M.; Syn, W.; Elsherbiny, N.M.; Kuo, P.C.; et al. Epigallocatechin-3-Gallate Upregulates miR-221 to Inhibit Osteopontin-Dependent Hepatic Fibrosis. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Pu, X.; Huang, G.; Guo, H.; Guo, C.; Li, H.; Ye, S.; Ling, S.; Jiang, L.; Tian, Y.; Lin, T. Circulating miR-221 Directly Amplified from Plasma Is a Potential Diagnostic and Prognostic Marker of Colorectal Cancer and Is Correlated with p53 Expression. J. Gastroenterol. Hepatol. 2010, 25, 1674–1680. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, T.; Watanabe, K.; Hara, E.S.; Ono, M.; Kuboki, T.; Calderwood, S.K. OstemiR: A Novel Panel of MicroRNA Biomarkers in Osteoblastic and Osteocytic Differentiation from Mesencymal Stem Cells. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.; Fu, J.; Zhou, W.; Li, W.; Dong, S.; Yu, S.; Hu, Z.; Wang, H.; Xie, Z. MicroRNA-125b Regulates Osteogenic Differentiation of Mesenchymal Stem Cells by Targeting Cbfβ in Vitro. Biochimie 2014, 102. [Google Scholar] [CrossRef]
- Shirafkan, N.; Mansoori, B.; Mohammadi, A.; Shomali, N.; Ghasbi, M.; Baradaran, B. MicroRNAs as Novel Biomarkers for Colorectal Cancer: New Outlooks. Biomed. Pharmacother. 2018, 97, 1319–1330. [Google Scholar] [CrossRef]
- Žlajpah, M.; Boštjančič, E.; Zidar, N. (Epi)genetic Regulation of Osteopontin in Colorectal Cancerogenesis. Epigenomics 2020, 12, 1389–1403. [Google Scholar] [CrossRef]
- Iacona, J.R.; Lutz, C.S. miR-146a-5p: Expression, Regulation, and Functions in Cancer. WIREs RNA 2019, 10, e1533. [Google Scholar] [CrossRef] [PubMed]
- Bleau, A.-M.; Redrado, M.; Nistal-Villan, E.; Villalba, M.; Exposito, F.; Redin, E.; de Aberasturi, A.L.; Larzabal, L.; Freire, J.; Gomez-Roman, J.; et al. miR-146a Targets c-Met and Abolishes Colorectal Cancer Liver Metastasis. Cancer Lett. 2018, 414, 257–267. [Google Scholar] [CrossRef]
- El Bairi, K.; Tariq, K.; Himri, I.; Jaafari, A.; Smaili, W.; Kandhro, A.H.; Gouri, A.; Ghazi, B. Decoding Colorectal Cancer Epigenomics. Cancer Genet. 2018, 220, 49–76. [Google Scholar] [CrossRef]
- Baretti, M.; Azad, N.S. The Role of Epigenetic Therapies in Colorectal Cancer. Curr. Probl. Cancer 2018, 42, 530–547. [Google Scholar] [CrossRef]
- Hijiya, N.; Setoguchi, M.; Matsuura, K.; Higuchi, Y.; Akizuki, S.; Yamamoto, S. Cloning and Characterization of the Human Osteopontin Gene and Its Promoter. Biochem. J. 1994, 303, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Yamamoto, S.; Hijiya, N.; Benveniste, E.N.; Gladson, C.L. Transcriptional Regulation of the Human Osteopontin Promoter: Functional Analysis and DNA-Protein Interactions. Oncogene 2000, 19, 5801–5809. [Google Scholar] [CrossRef] [Green Version]
- The Cancer Genome Atlas Network. Comprehensive Molecular Characterization of Human Colon and Rectal Cancer. Nature 2012, 487, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Rohde, F.; Rimkus, C.; Friederichs, J.; Rosenberg, R.; Marthen, C.; Doll, D.; Holzmann, B.; Siewert, J.-R.; Janssen, K.-P. Expression of Osteopontin, a Target Gene of de-Regulated Wnt Signaling, Predicts Survival in Colon Cancer. Int. J. Cancer 2007, 121, 1717–1723. [Google Scholar] [CrossRef]
- Ravindranath, A.; O’Connell, A.; Johnston, P.G.; El-Tanani, M.K. The Role of LEF/TCF Factors in Neoplastic Transformation. Curr. Mol. Med. 2008, 8. [Google Scholar] [CrossRef]
- Martinez, C.; Churchman, M.; Freeman, T.; Ilyas, M. Osteopontin Provides Early Proliferative Drive and May Be Dependent upon Aberrant c-Myc Signalling in Murine Intestinal Tumours. Exp. Mol. Pathol. 2010, 88, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Rennoll, S.; Yochum, G. Regulation of MYC Gene Expression by Aberrant Wnt/β-Catenin Signaling in Colorectal Cancer. World J. Biol. Chem. 2015, 6, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.Y.; Chua, M.-S.; Choi, Y.-L.; Yeh, W.; Kim, Y.H.; Azzi, R.; Adams, G.A.; Sainani, K.; van de Rijn, M.; So, S.K.; et al. Comparative Profiling of Primary Colorectal Carcinomas and Liver Metastases Identifies LEF1 as a Prognostic Biomarker. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashida, R.; Tominaga, K.; Sasaki, E.; Watanabe, T.; Fujiwara, Y.; Oshitani, N.; Higuchi, K.; Mitsuyama, S.; Iwao, H.; Arakawa, T. AP-1 and Colorectal Cancer. Inflammopharmacology 2005, 13, 113–125. [Google Scholar] [CrossRef]
- El-Tanani, M.; Platt-Higgins, A.; Rudland, P.S.; Campbell, F.C. Ets Gene PEA3 Cooperates with β-Catenin-Lef-1 and c-Jun in Regulation of Osteopontin Transcription. J. Biol. Chem. 2004, 279, 20794–20806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wai, P.Y.; Mi, Z.; Gao, C.; Guo, H.; Marroquin, C.; Kuo, P.C. Ets-1 and Runx2 Regulate Transcription of a Metastatic Gene, Osteopontin, in Murine Colorectal Cancer Cells. J. Biol. Chem. 2006, 281, 18973–18982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssef, N.S.; Osman, W.M. Relationship between Osteopontin and β-Catenin Immunohistochemical Expression and Prognostic Parameters of Colorectal Carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 1503–1514. [Google Scholar] [PubMed]
- Shi, X.; Bai, S.; Li, L.; Cao, X. Hoxa-9 Represses Transforming Growth Factor-β-Induced Osteopontin Gene Transcription. J. Biol. Chem. 2001, 276, 850–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georges, R.; Adwan, H.; Zhivkova, M.; Eyol, E.; Bergmann, F.; Berger, M.R. Regulation of Osteopontin and Related Proteins in Rat CC531 Colorectal Cancer Cells. Int. J. Oncol. 2010, 37, 249–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sase, T.; Suzuki, T.; Miura, K.; Shiiba, K.; Sato, I.; Nakamura, Y.; Takagi, K.; Onodera, Y.; Miki, Y.; Watanabe, M.; et al. Runt-Related Transcription Factor 2 in Human Colon Carcinoma: A Potent Prognostic Factor Associated with Estrogen Receptor. Int. J. Cancer 2012, 131, 2284–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, D.; Li, S.; Jiang, W.; Zhu, J.; Liu, J.; Zhao, S. miR-539 Inhibits Human Colorectal Cancer Progression by Targeting RUNX2. Biomed. Pharmacother. 2017, 95, 1314–1320. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.-C.; Lin, J.-K. Liquiritigenin Inhibits Colorectal Cancer Proliferation, Invasion, and Epithelial-to-Mesenchymal Transition by Decreasing Expression of Runt-Related Transcription Factor 2. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2019, 27, 139–146. [Google Scholar] [CrossRef]
- Xu, H.; McCann, M.; Zhang, Z.; Posner, G.H.; Bingham, V.; El-Tanani, M.; Campbell, F.C. Vitamin D Receptor Modulates the Neoplastic Phenotype through Antagonistic Growth Regulatory Signals. Mol. Carcinog. 2009, 48, 758–772. [Google Scholar] [CrossRef]
- Matusiak, D.; Murillo, G.; Carroll, R.E.; Mehta, R.G.; Benya, R.V. Expression of Vitamin D Receptor and 25-Hydroxyvitamin D3-1α-Hydroxylase in Normal and Malignant Human Colon. Cancer Epidemiol. Prev. Biomark. 2005, 14, 2370–2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheinin, Y.; Kaserer, K.; Wrba, F.; Wenzl, E.; Kriwanek, S.; Peterlik, M.; Cross, H.S. In Situ mRNA Hybridization Analysis and Immunolocalization of the Vitamin D Receptor in Normal and Carcinomatous Human Colonic Mucosa: Relation to Epidermal Growth Factor Receptor Expression. Virchows Arch. 2000, 437, 501–507. [Google Scholar] [CrossRef]
- Lechner, D.; Kállay, E.; Cross, H.S. 1α,25-Dihydroxyvitamin D3 Downregulates CYP27B1 and Induces CYP24A1 in Colon Cells. Mol. Cell. Endocrinol. 2007, 263, 55–64. [Google Scholar] [CrossRef]
- Labudzynskyi, D.; Shymanskyy, I.; Veliky, M. Role of Vitamin D3 in Regulation of Interleukin-6 and Osteopontin Expression in Liver of Diabetic Mice. Eur Rev Med Pharmacol Sci. 2016, 20, 2916–2919. [Google Scholar]
- Zagani, R.; Hamzaoui, N.; Cacheux, W.; de Reyniès, A.; Terris, B.; Chaussade, S.; Romagnolo, B.; Perret, C.; Lamarque, D. Cyclooxygenase-2 Inhibitors Down-Regulate Osteopontin and Nr4a2—New Therapeutic Targets for Colorectal Cancers. Gastroenterology 2009, 137, 1358–1366. [Google Scholar] [CrossRef]
- Boudjadi, S.; Bernatchez, G.; Beaulieu, J.-F.; Carrier, J.C. Control of the Human Osteopontin Promoter by ERRα in Colorectal Cancer. Am. J. Pathol. 2013, 183, 266–276. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Harris, L.G.; Metge, B.J.; Liu, S.; Riker, A.I.; Samant, R.S.; Shevde, L.A. The Hedgehog Pathway Transcription Factor GLI1 Promotes Malignant Behavior of Cancer Cells by Up-Regulating Osteopontin. J. Biol. Chem. 2009, 284, 22888–22897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchett, J.; Harvey, E.; Athwal, V.; Berry, A.; Rowe, C.; Oakley, F.; Moles, A.; Mann, D.A.; Bobola, N.; Sharrocks, A.D.; et al. Osteopontin Is a Novel Downstream Target of SOX9 with Diagnostic Implications for Progression of Liver Fibrosis in Humans. Hepatology 2012, 56, 1108–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimba, E.R.; Tilli, T.M. Human Osteopontin Splicing Isoforms: Known Roles, Potential Clinical Applications and Activated Signaling Pathways. Cancer Lett. 2013, 331, 11–17. [Google Scholar] [CrossRef]
- Briones-Orta, M.A.; Avendaño-Vázquez, S.E.; Aparicio-Bautista, D.I.; Coombes, J.D.; Weber, G.F.; Syn, W.-K. Osteopontin Splice Variants and Polymorphisms in Cancer Progression and Prognosis. Biochim. Biophys. Acta BBA-Rev. Cancer 2017, 1868, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Mirza, M.; Shaughnessy, E.; Hurley, J.K.; Vanpatten, K.A.; Pestano, G.A.; He, B.; Weber, G.F. Osteopontin-c Is a Selective Marker of Breast Cancer. Int. J. Cancer 2008, 122, 889–897. [Google Scholar] [CrossRef]
- Zduniak, K.; Ziolkowski, P.; Ahlin, C.; Agrawal, A.; Agrawal, S.; Blomqvist, C.; Fjällskog, M.-L.; Weber, G.F. Nuclear Osteopontin-c Is a Prognostic Breast Cancer Marker. Br. J. Cancer 2015, 112, 729–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, C.; Wang, Z.; Gu, Y.; Jiang, W.G.; Cheng, S. Prognostic Value of Osteopontin Splice Variant-c Expression in Breast Cancers: A Meta-Analysis. BioMed Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Weber, G.F. The Osteopontin-c Splice Junction Is Important for Anchorage-Independent Growth. Mol. Carcinog. 2014, 53, 480–487. [Google Scholar] [CrossRef]
- Boguslawska, J.; Sokol, E.; Rybicka, B.; Czubaty, A.; Rodzik, K.; Piekielko-Witkowska, A. microRNAs Target SRSF7 Splicing Factor to Modulate the Expression of Osteopontin Splice Variants in Renal Cancer Cells. Gene 2016, 595, 142–149. [Google Scholar] [CrossRef]
- Chang, S.; Huang, J.; Niu, H.; Wang, J.; Si, Y.; Bai, Z.; Cheng, S.; Ding, W. Epigenetic Regulation of Osteopontin Splicing Isoform c Defines Its Role as a Microenvironmental Factor to Promote the Survival of Colon Cancer Cells from 5-FU Treatment. Cancer Cell Int. 2020, 20, 452. [Google Scholar] [CrossRef]
- Shinohara, M.L.; Kim, H.-J.; Kim, J.-H.; Garcia, V.A.; Cantor, H. Alternative Translation of Osteopontin Generates Intracellular and Secreted Isoforms That Mediate Distinct Biological Activities in Dendritic Cells. Proc. Natl. Acad. Sci. USA 2008, 105, 7235–7239. [Google Scholar] [CrossRef] [Green Version]
- Inoue, M.; Shinohara, M.L. Intracellular Osteopontin (iOPN) and Immunity. Immunol. Res. 2011, 49, 160–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zohar, R.; Lee, W.; Arora, P.; Cheifetz, S.; McCulloch, C.; Sodek, J. Single Cell Analysis of Intracellular Osteopontin in Osteogenic Cultures of Fetal Rat Calvarial Cells. J. Cell. Physiol. 1997, 170, 88–100. [Google Scholar] [CrossRef]
- Kim, H.-J.; Lee, H.-J.; Jun, J.-I.; Oh, Y.; Choi, S.-G.; Kim, H.; Chung, C.-W.; Kim, I.-K.; Park, I.-S.; Chae, H.-J.; et al. Intracellular Cleavage of Osteopontin by Caspase-8 Modulates Hypoxia/reoxygenation Cell Death through p53. Proc. Natl. Acad. Sci. USA 2009, 106, 15326–15331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junaid, A.; Moon, M.C.; Harding, G.E.J.; Zahradka, P. Osteopontin Localizes to the Nucleus of 293 Cells and Associates with Polo-like Kinase-1. Am. J. Physiol.-Cell Physiol. 2007, 292, C919–C926. [Google Scholar] [CrossRef] [Green Version]
- Sodek, J.; Ganss, B.; McKee, M.D. Osteopontin. Crit. Rev. Oral Biol. Med. 2000, 11, 279–303. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Baba, O.; Butler, W.T. Post-Translational Modifications of Sibling Proteins and Their Roles in Osteogenesis and Dentinogenesis. Crit. Rev. Oral Biol. Med. Off. Publ. Am. Assoc. Oral Biol. 2004, 15, 126–136. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.; Kazanecki, C.C.; Petersen, T.E.; Rittling, S.R.; Denhardt, D.T.; Sørensen, E.S. Cell Type-Specific Post-Translational Modifications of Mouse Osteopontin Are Associated with Different Adhesive Properties. J. Biol. Chem. 2007, 282, 19463–19472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashkar, S.; Weber, G.F.; Panoutsakopoulou, V.; Sanchirico, M.E.; Jansson, M.; Zawaideh, S.; Rittling, S.R.; Denhardt, D.T.; Glimcher, M.J.; Cantor, H. Eta-1 (Osteopontin): An Early Component of Type-1 (Cell-Mediated) Immunity. Science 2000, 287, 860–864. [Google Scholar] [CrossRef]
- Weber, G.F.; Zawaideh, S.; Hikita, S.; Kumar, V.A.; Cantor, H.; Ashkar, S. Phosphorylation-Dependent Interaction of Osteopontin with Its Receptors Regulates Macrophage Migration and Activation. J. Leukoc. Biol. 2002, 72, 752–761. [Google Scholar] [CrossRef]
- Kazanecki, C.C.; Kowalski, A.J.; Ding, T.; Rittling, S.R.; Denhardt, D.T. Characterization of Anti-Osteopontin Monoclonal Antibodies: Binding Sensitivity to Post-Translational Modifications. J. Cell. Biochem. 2007, 102, 925–935. [Google Scholar] [CrossRef]
- Castellano, G.; Malaponte, G.; Mazzarino, M.C.; Figini, M.; Marchese, F.; Gangemi, P.; Travali, S.; Stivala, F.; Canevari, S.; Libra, M. Activation of the Osteopontin/Matrix Metalloproteinase-9 Pathway Correlates with Prostate Cancer Progression. Clin. Cancer Res. 2008, 14, 7470–7480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokosaki, Y.; Matsuura, N.; Sasaki, T.; Murakami, I.; Schneider, H.; Higashiyama, S.; Saitoh, Y.; Yamakido, M.; Taooka, Y.; Sheppard, D. The Integrin α9β1 Binds to a Novel Recognition Sequence (SVVYGLR) in the Thrombin-Cleaved Amino-Terminal Fragment of Osteopontin. J. Biol. Chem. 1999, 274, 36328–36334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, N.; Sakai, F.; Kon, S.; Morimoto, J.; Kimura, C.; Yamazaki, H.; Okazaki, I.; Seki, N.; Fujii, T.; Uede, T. Essential Role of the Cryptic Epitope SLAYGLR within Osteopontin in a Murine Model of Rheumatoid Arthritis. J. Clin. Investig. 2003, 112, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.; Kläning, E.; Nielsen, M.S.; Andersen, M.H.; Sørensen, E.S. C-Terminal Modification of Osteopontin Inhibits Interaction with the αVβ3-Integrin. J. Biol. Chem. 2012, 287, 3788–3797. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Sun, L.; Jiang, C.; Zhou, H.; Gu, L.; Liu, Y.; Xu, Q. SPP1, Analyzed by Bioinformatics Methods, Promotes the Metastasis in Colorectal Cancer by Activating EMT Pathway. Biomed. Pharmacother. 2017, 91, 1167–1177. [Google Scholar] [CrossRef]
- Ishigamori, R.; Komiya, M.; Takasu, S.; Mutoh, M.; Imai, T.; Takahashi, M. Osteopontin Deficiency Suppresses Intestinal Tumor Development in Apc-Deficient Min Mice. Int. J. Mol. Sci. 2017, 18, 1058. [Google Scholar] [CrossRef]
- Harada, N.; Mizoi, T.; Kinouchi, M.; Hoshi, K.; Ishii, S.; Shiiba, K.; Sasaki, I.; Matsuno, S. Introduction of Antisense CD44s cDNA down-Regulates Expression of Overall CD44 Isoforms and Inhibits Tumor Growth and Metastasis in Highly Metastatic Colon Carcinoma Cells. Int. J. Cancer 2001, 91, 67–75. [Google Scholar] [CrossRef]
- Sinicrope, F.A.; Roddey, G.; McDonnell, T.J.; Shen, Y.; Cleary, K.R.; Stephens, L.C. Increased Apoptosis Accompanies Neoplastic Development in the Human Colorectum. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 1996, 2, 1999–2006. [Google Scholar]
- Huang, R.; Quan, Y.; Chen, J.; Wang, T.; Xu, M.; Ye, M.; Yuan, H.; Zhang, C.; Liu, X.; Min, Z. Osteopontin Promotes Cell Migration and Invasion, and Inhibits Apoptosis and Autophagy in Colorectal Cancer by Activating the p38 MAPK Signaling Pathway. Cell. Physiol. Biochem. 2017, 41, 1851–1864. [Google Scholar] [CrossRef]
- Cao, D.-X. Osteopontin as Potential Biomarker and Therapeutic Target in Gastric and Liver Cancers. World J. Gastroenterol. 2012, 18, 3923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Zhang, S.; Gao, F.; Liu, Z.; Lu, M.; Peng, S.; Zhang, T.; Zhang, F. Osteopontin Enhances the Expression of HOTAIR in Cancer Cells via IRF1. Biochim. Biophys. Acta BBA-Gene Regul. Mech. 2014, 1839, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducreux, M.; Österlund, P.; Pignon, J.P. Angiogenesis Inhibition in the Second-Line Treatment of Metastatic Colorectal cancer—A Definite Conclusion? Semin. Oncol. 2017, 44, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Wohlleben, G.; Hauff, K.; Gasser, M.; Waaga-Gasser, A.; Grimmig, T.; Flentje, M.; Polat, B. Hypoxia Induces Differential Expression Patterns of Osteopontin and CD44 in Colorectal Carcinoma. Oncol. Rep. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.-L.; Lin, K.-J.; Bai, A.-P.; Wang, W.-X.; Meng, X.-K.; Su, X.-L.; Hou, M.-X.; Dong, P.-D.; Zhang, J.-J.; Wang, Z.-Y.; et al. Osteopontin Knockdown Suppresses the Growth and Angiogenesis of Colon Cancer Cells. World J. Gastroenterol. WJG 2014, 20, 10440–10448. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.-S.; Chen, W.-S.; Chen, L.-L.; Chen, C.-C.; Hsu, Y.-T.; Chua, K.V.; Wang, H.-D.; Huang, T.-S. Osteopontin–integrin Engagement Induces HIF-1α–TCF12-Mediated Endothelial-Mesenchymal Transition to Exacerbate Colorectal Cancer. Oncotarget 2017, 9, 4998–5015. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.Z.; Miow, Q.H.; Miki, Y.; Noda, T.; Mori, S.; Huang, R.Y.; Thiery, J.P. Epithelial-mesenchymal Transition Spectrum Quantification and Its Efficacy in Deciphering Survival and Drug Responses of Cancer Patients. EMBO Mol. Med. 2014, 6, 1279–1293. [Google Scholar] [CrossRef] [PubMed]
- Acloque, H.; Adams, M.S.; Fishwick, K.; Bronner-Fraser, M.; Nieto, M.A. Epithelial-Mesenchymal Transitions: The Importance of Changing Cell State in Development and Disease. J. Clin. Investig. 2009, 119, 1438–1449. [Google Scholar] [CrossRef] [Green Version]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The Consensus Molecular Subtypes of Colorectal Cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef]
- Vu, T.; Datta, P. Regulation of EMT in Colorectal Cancer: A Culprit in Metastasis. Cancers 2017, 9, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boesch, M.; Spizzo, G.; Seeber, A. Concise Review: Aggressive Colorectal Cancer: Role of Epithelial Cell Adhesion Molecule in Cancer Stem Cells and Epithelial-to-Mesenchymal Transition. Stem Cells Transl. Med. 2018, 7, 495–501. [Google Scholar] [CrossRef]
- Kothari, A.N.; Arffa, M.L.; Chang, V.; Blackwell, R.H.; Syn, W.-K.; Zhang, J.; Mi, Z.; Kuo, P.C. Osteopontin—A Master Regulator of Epithelial-Mesenchymal Transition. J. Clin. Med. 2016, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Ng, L.; Wan, T.; Chow, A.; Iyer, D.; Man, J.; Chen, G.; Yau, T.C.-C.; Lo, O.; Foo, C.-C.; Poon, J.T.-C.; et al. Osteopontin Overexpression Induced Tumor Progression and Chemoresistance to Oxaliplatin through Induction of Stem-Like Properties in Human Colorectal Cancer. Stem Cells Int. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Rao, G.; Du, L.; Chen, Q. Osteopontin, a Possible Modulator of Cancer Stem Cells and Their Malignant Niche. Oncoimmunology 2013, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todaro, M.; Gaggianesi, M.; Catalano, V.; Benfante, A.; Iovino, F.; Biffoni, M.; Apuzzo, T.; Sperduti, I.; Volpe, S.; Cocorullo, G.; et al. CD44v6 Is a Marker of Constitutive and Reprogrammed Cancer Stem Cells Driving Colon Cancer Metastasis. Cell Stem Cell 2014, 14, 342–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayadi, M.; Bouygues, A.; Ouaret, D.; Ferrand, N.; Chouaib, S.; Thiery, J.-P.; Muchardt, C.; Sabbah, M.; Larsen, A.K. Chronic Chemotherapeutic Stress Promotes Evolution of Stemness and WNT/beta-Catenin Signaling in Colorectal Cancer Cells: Implications for Clinical Use of WNT-Signaling Inhibitors. Oncotarget 2015, 6, 18518–18533. [Google Scholar] [CrossRef]
- Kumar, S.; Sharma, P.; Kumar, D.; Chakraborty, G.; Gorain, M.; Kundu, G.C. Functional Characterization of Stromal Osteopontin in Melanoma Progression and Metastasis. PLoS ONE 2013, 8, e69116. [Google Scholar] [CrossRef] [Green Version]
- Iida, T.; Wagatsuma, K.; Hirayama, D. Hiroshi Nakase Is Osteopontin a Friend or Foe of Cell Apoptosis in Inflammatory Gastrointestinal and Liver Diseases? Int. J. Mol. Sci. 2017, 19, 7. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Wu, H. The Significant Prognostic Value of Circulating Tumor Cells in Colorectal Cancer: A Systematic Review and Meta-Analysis. Curr. Probl. Cancer 2018, 42, 95–106. [Google Scholar] [CrossRef]
- Cayrefourcq, L.; Mazard, T.; Joosse, S.; Solassol, J.; Ramos, J.; Assenat, E.; Schumacher, U.; Costes, V.; Maudelonde, T.; Pantel, K.; et al. Establishment and Characterization of a Cell Line from Human Circulating Colon Cancer Cells. Cancer Res. 2015, 75, 892–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; von Au, A.; Schnölzer, M.; Hackert, T.; Zöller, M. CD44v6-Competent Tumor Exosomes Promote Motility, Invasion and Cancer-Initiating Cell Marker Expression in Pancreatic and Colorectal Cancer Cells. Oncotarget 2016, 7, 55409–55436. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, M.; Bæk, R.; Pedersen, S.; Søndergaard, E.K.L.; Kristensen, S.R.; Varming, K. Extracellular Vesicle (EV) Array: Microarray Capturing of Exosomes and Other Extracellular Vesicles for Multiplexed Phenotyping. J. Extracell. Vesicles 2013, 2, 20920. [Google Scholar] [CrossRef] [PubMed]
- Minciacchi, V.R.; Freeman, M.R.; Di Vizio, D. Extracellular Vesicles in Cancer: Exosomes, Microvesicles and the Emerging Role of Large Oncosomes. Semin. Cell Dev. Biol. 2015, 40, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Jeong, S.; Xia, Q.; Kong, X. Role of Osteopontin in Liver Diseases. Int. J. Biol. Sci. 2016, 12, 1121–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Pan, C.; Hu, H.; Zheng, S.; Ding, L. Osteopontin-Enhanced Hepatic Metastasis of Colorectal Cancer Cells. PLoS ONE 2012, 7, e47901. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-B.; Qi, Q.-R.; Wu, K.-L.; Xie, Q.-Z. Role of Osteopontin in Decidualization and Pregnancy Success. Reproduction 2018, 155, 423–432. [Google Scholar] [CrossRef]
- Jia, R.; Liang, Y.; Chen, R.; Liu, G.; Wang, H.; Tang, M.; Zhou, X.; Wang, H.; Yang, Y.; Wei, H.; et al. Osteopontin Facilitates Tumor Metastasis by Regulating Epithelial–mesenchymal Plasticity. Cell Death Dis. 2016, 7, e2564. [Google Scholar] [CrossRef]
- Limani, P.; Linecker, M.; Schneider, M.A.; Kron, P.; Tschuor, C.; Kachaylo, E.; Ungethuem, U.; Nicolau, C.; Lehn, J.-M.; Graf, R.; et al. The Allosteric Hemoglobin Effector ITPP Inhibits Metastatic Colon Cancer in Mice. Ann. Surg. 2017, 266, 746–753. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.S.; Piper, M.A.; Perdue, L.A.; Rutter, C.M.; Webber, E.M.; O’Connor, E.; Smith, N.; Whitlock, E.P. Screening for Colorectal Cancer: Updated Evidence Report and Systematic Review for the US Preventive Services Task Force. JAMA 2016, 315, 2576. [Google Scholar] [CrossRef] [Green Version]
- Wild, N.; Andres, H.; Rollinger, W.; Krause, F.; Dilba, P.; Tacke, M.; Karl, J. A Combination of Serum Markers for the Early Detection of Colorectal Cancer. Clin. Cancer Res. 2010, 16, 6111–6121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, S.; Krause, F.; Rolny, V.; Strobl, M.; Morgenstern, D.; Datz, C.; Chen, H.; Brenner, H. Evaluation of a 5-Marker Blood Test for Colorectal Cancer Early Detection in a Colorectal Cancer Screening Setting. Clin. Cancer Res. 2016, 22, 1725–1733. [Google Scholar] [CrossRef] [Green Version]
- Dressen, K.; Hermann, N.; Manekeller, S.; Walgenbach-Bruenagel, G.; Schildberg, F.A.; Hettwer, K.; Uhlig, S.; Kalff, J.C.; Hartmann, G.; Holdenrieder, S. Diagnostic Performance of a Novel Multiplex Immunoassay in Colorectal Cancer. Anticancer Res. 2017, 37, 2477–2486. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.D.; Li, L.; Wang, Y.; Thoburn, C.; Afsari, B.; Danilova, L.; Douville, C.; Javed, A.A.; Wong, F.; Mattox, A.; et al. Detection and Localization of Surgically Resectable Cancers with a Multi-Analyte Blood Test. Science 2018, 359, 926–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Liang, F.; Zhang, B.; Yan, W.; Zhang, J. The Impact of Osteopontin on Prognosis and Clinicopathology of Colorectal Cancer Patients: A Systematic Meta-Analysis. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef]
- Klement, J.D.; Paschall, A.V.; Redd, P.S.; Ibrahim, M.L.; Lu, C.; Yang, D.; Celis, E.; Abrams, S.I.; Ozato, K.; Liu, K. An osteopontin/CD44 Immune Checkpoint Controls CD8+ T Cell Activation and Tumor Immune Evasion. J. Clin. Investig. 2018, 128, 5549–5560. [Google Scholar] [CrossRef] [PubMed]
- Moorman, H.R.; Poschel, D.; Klement, J.D.; Lu, C.; Redd, P.S.; Liu, K. Osteopontin: A Key Regulator of Tumor Progression and Immunomodulation. Cancers 2020, 12, 3379. [Google Scholar] [CrossRef]
- Klement, J.D.; Poschel, D.B.; Lu, C.; Merting, A.D.; Yang, D.; Redd, P.S.; Liu, K. Osteopontin Blockade Immunotherapy Increases Cytotoxic T Lymphocyte Lytic Activity and Suppresses Colon Tumor Progression. Cancers 2021, 13, 1006. [Google Scholar] [CrossRef]
- Bandopadhyay, M.; Bulbule, A.; Butti, R.; Chakraborty, G.; Ghorpade, P.; Ghosh, P.; Gorain, M.; Kale, S.; Kumar, D.; Kumar, S.; et al. Osteopontin as a Therapeutic Target for Cancer. Expert Opin. Ther. Targets 2014, 18, 883–895. [Google Scholar] [CrossRef]
- Hao, C.; Cui, Y.; Owen, S.; Li, W.; Cheng, S.; Jiang, W.G. Human Osteopontin: Potential Clinical Applications in Cancer (Review). Int. J. Mol. Med. 2017, 39, 1327–1337. [Google Scholar] [CrossRef] [Green Version]
- Talbot, L.J.; Mi, Z.; Bhattacharya, S.D.; Kim, V.; Guo, H.; Kuo, P.C. Pharmacokinetic Characterization of an RNA Aptamer against Osteopontin and Demonstration of in Vivo Efficacy in Reversing Growth of Human Breast Cancer Cells. Surgery 2011, 150, 224–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, N.I.F.; Gunasekharan, V.K.; Ravindranath, A.; O’Connell, C.; Johnston, P.G.; El-Tanani, M.K. Osteopontin as a Target for Cancer Therapy. Front. Biosci. 2008, 13, 4361–4372. [Google Scholar] [CrossRef] [Green Version]
- Weber, G.F. The Metastasis Gene Osteopontin: A Candidate Target for Cancer Therapy. Biochim. Biophys. Acta 2001, 1552, 61–85. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amilca-Seba, K.; Sabbah, M.; Larsen, A.K.; Denis, J.A. Osteopontin as a Regulator of Colorectal Cancer Progression and Its Clinical Applications. Cancers 2021, 13, 3793. https://doi.org/10.3390/cancers13153793
Amilca-Seba K, Sabbah M, Larsen AK, Denis JA. Osteopontin as a Regulator of Colorectal Cancer Progression and Its Clinical Applications. Cancers. 2021; 13(15):3793. https://doi.org/10.3390/cancers13153793
Chicago/Turabian StyleAmilca-Seba, Katyana, Michèle Sabbah, Annette K. Larsen, and Jérôme A. Denis. 2021. "Osteopontin as a Regulator of Colorectal Cancer Progression and Its Clinical Applications" Cancers 13, no. 15: 3793. https://doi.org/10.3390/cancers13153793
APA StyleAmilca-Seba, K., Sabbah, M., Larsen, A. K., & Denis, J. A. (2021). Osteopontin as a Regulator of Colorectal Cancer Progression and Its Clinical Applications. Cancers, 13(15), 3793. https://doi.org/10.3390/cancers13153793